Трихобилгарция регенти
| Трихобилгарция регенти | |
|---|---|
 | |
| Trichobilharzia regenti , церкарии | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Платихельминты |
| Сорт: | Трематоды |
| Заказ: | Диплостомида |
| Семья: | Шистосоматиды |
| Род: | Трихобилгарция |
| Разновидность: | Т. регент |
| Биномиальное имя | |
| Трихобилгарция регенти Горак, Коларжова и Дворжак, 1998 г. [1] | |
Trichobilharzia regenti — нейропатогенный паразитический плоский червь птиц , который также вызывает церкариальный дерматит у людей. [2] Впервые вид был описан в 1998 году в Чехии. [1] а впоследствии он был обнаружен и в других европейских странах, например в Дании , [3] Германия , [4] Франция , [5] Исландия , [6] Польша , [7] [8] Швейцария , [9] или Россия , [10] и даже в Иране . [11] [12] Благодаря его уникальному нейротропному поведению у позвоночных-хозяев, взаимодействия хозяин-паразит широко изучаются с точки зрения молекулярной биологии, биохимии и иммунологии. [13] [14] [15]
Жизненный цикл
[ редактировать ]Жизненный цикл T. regent аналогичен жизненному циклу шистосом человека . Взрослые сосальщики спариваются в слизистой оболочке носа птиц-анатид (например, Anas platyrhynchos , Spatula clypeata или Cairina moschata ) и производят яйца с мирацидиями , которые вылупляются непосредственно в тканях хозяина и вытекают наружу, когда птица пьет/кормится. [1] Оказавшись в воде, мирацидии плавают, используя свои реснички , и активно ищут подходящего -моллюска промежуточного хозяина ( Radix lagotis , Radix labiata , Radix peregra ). [16] У улитки мирацидии развиваются в первичную спороцисту , в которой образуются вторичные спороцисты , которые позже дают начало церкариям. [17]
Церкарии , инфекционные личинки, выходят из улитки и проникают в кожу птицы-хозяина. После проникновения в кожу хозяина они теряют иммуногенный поверхностный гликокаликс. [18] и превращаются в шистосомулу (подвзрослая стадия, ср. шистосомулум). Затем шистосомулы ищут периферические нервы, чтобы использовать их для доступа к спинному мозгу. Через него они продолжают миграцию в мозг. [19] [20] и, наконец, носовая ткань клюва. Здесь они созревают, совокупляются и откладывают яйца, вызывая патологию (воспалительную инфильтрацию, кровоизлияния). [21]
Если церкариями заражаются млекопитающие (а не птицы), паразиты погибают в коже, захватываясь иммунным ответом. [22] Клиническое проявление такой инфекции известно как запущенное аллергическое заболевание, называемое церкариальным дерматитом (или зудом пловца ). [2] [23] У мышей, особенно иммунодефицитных, наблюдалась миграция паразита в спинной мозг. [24] [25]
Полный жизненный цикл T. regent можно поддерживать в лабораторных условиях, используя Radix lagotis и домашнюю утку ( Anas platyrhynchos f. Domestica) в качестве промежуточных и окончательных хозяев соответственно. [1] Интересно, что домашние утки также могут служить резервуарными хозяевами на участках аквакультуры, например на рисовых полях. [12] Для изучения биологии T. regent у млекопитающих C57BL/6 , BALB/c a SCID . в качестве случайных хозяев используют линии мышей [25] [26] [27]
Миграция у позвоночных хозяев
[ редактировать ]Когда церкарии T. regenti находят хозяина у птиц или млекопитающих, они проникают в его кожу. Для этой цели они оснащены цистеиновыми пептидазами, присутствующими в их экскреторных/секреторных продуктах, которые способны расщеплять кератин и коллаген . [28] [29] Эксперименты с лабораторно полученной рекомбинантной формой цистеинпептидазы катепсина В2 T. regenti (TrCB2) подтвердили ее способность расщеплять белки кожи (коллаген, кератин и эластин). [30]
После проникновения через кожу церкарии превращаются в шистосомулы и начинают миграцию по организму хозяина. Они избегают проникновения в кровеносные капилляры и предпочитают проникать в периферические нервы конечностей хозяина. Шистосомулы обнаруживаются в периферических нервах уток и мышей уже через 1,5 и 1 день после заражения (DPI) соответственно. [20] У обоих типов хозяев шистосомулы проявляют высокое сродство к центральной нервной системе , в которую они проникают через спинномозговые корешки . [26] Согласно недавним наблюдениям с помощью методов 3D-визуализации (ультрамикроскопия и микроКТ ), шистосомулы, по-видимому, мигрируют преимущественно через белое вещество спинного мозга как у птиц, так и у млекопитающих. [27]
Дальнейшее течение инфекции различается у окончательных и случайных хозяев. У уток шистосомулы наблюдаются в синсакральных сегментах спинного мозга через 3 ДПИ и через 7–8 дней (10–11 ДПИ) достигают головного мозга. В окончательной локализации (ткани носа) они возникают на 13–14 ДПИ, а откладка яиц начинается на 15 ДПИ. [20] [21] У мышей первые шистосомулы обнаруживаются в поясничном отделе спинного мозга уже на 2 ДПИ, а продолговатый мозг на следующий день инвазируются в , но только у некоторых особей. Большинство шистосомул остаются локализованными в грудном и шейном отделах спинного мозга и лишь в исключительных случаях мигрируют в головной мозг. [19] [20] В полости носа не обнаружено ни наличие червей, ни их созревание в нервной ткани. Развитие шистосомул у мышей подавляется, вероятно, из-за иммунного ответа хозяина и/или присутствия/отсутствия некоторых существенных (питательных, стимулирующих) факторов хозяина. [25]
Патология у позвоночных-хозяев
[ редактировать ]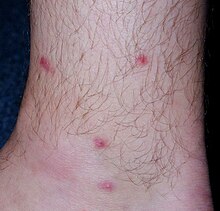
У позвоночных-хозяев, инфицированных T. regenti , патологические состояния могут быть вызваны:
- проникающие в кожу церкарии, превращающиеся в шистосомулы,
- шистосомулы, мигрирующие через центральную нервную систему (ЦНС),
- взрослые особи откладывают яйца в слизистую оболочку носа (только у птиц-хозяев).

Хотя мыши являются случайными хозяевами, большинство исследований патологического действия T. regent было проведено на этой модели.
Патологии кожи
[ редактировать ]В начальной фазе инфекции раннетрансформированные шистосомулы локализуются в коже. Информация о патологии кожи птиц пока не полная. У мышей немедленный отек и утолщение участка появляются уже через 30 мин после проникновения церкарий; также проявляется эритема. В течение 48 часов вокруг паразитов развиваются воспалительные очаги, содержащие нейтрофилы, эозинофилы, макрофаги, CD4+ лимфоциты и дегранулирующие тучные клетки. [22] [24]
При повторных инфекциях клеточная инфильтрация существенно увеличивается, а обширное воспаление может привести к образованию крупных абсцессов или даже эпидермального и/или дермального некроза. [22] У человека клинические симптомы проникновения церкариоза заключаются в образовании пятен/папул в местах проникновения паразита на кожу, сопровождающихся интенсивным зудом. Проявление более тяжелое у ранее сенсибилизированных людей. Это заболевание, вызываемое не только T. regenti , но и церкариями других видов шистосом птиц, называется церкариальным дерматитом (он же « зуд пловца» ). Считается забытым аллергическим заболеванием. [2] [23]

патология ЦНС
[ редактировать ]Следующая фаза заражения T. regenti представлена миграцией шистосомул в центральную нервную систему. [15] Это сопровождается серьезными неврологическими нарушениями у птиц, страдающих параличом ног и нарушением равновесия. [19] У уток отмечен эозинофильный менингит с паразитами, окруженными эозинофилами и гетерофилами. Кроме того, лейкоциты инфильтрировали периваскулярные пространства и ткани, прилегающие к центральному каналу. [31]
На этом этапе шистосомулы питаются нервной тканью, о чем свидетельствует обнаружение олигодендроцитов и нейронов в просвете кишечника паразита. [26] Цистеинпептидаза катепсин В1 T. regenti (TrCB1), локализованная в кишечнике мигрирующих шистосомул, способна расщеплять основные белки миелина , что, вероятно, служит для пищеварения нервной ткани. [32] Тем не менее, проглатывание нервной ткани, вероятно, оказывает лишь незначительное патогенное воздействие на центральную нервную ткань хозяина. [26] Это подтверждается наблюдениями паралича ног только у хозяев с ослабленным иммунитетом. [24] [26] тогда как в экспериментах с иммунокомпетентными линиями мышей у инфицированных животных не выявлено неврологических нарушений. [20] [24] [26] Неврологическая симптоматика возникает, вероятно, вследствие механического повреждения нервной ткани, приводящего к дистрофическим или даже некротическим изменениям нейронов и повреждению аксонов. Причиной этого являются крупные мигрирующие шистосомулы (приблизительно 340×80 мкм), которые не уничтожаются должным иммунным ответом. [24] [26] [31] Недавно было высказано предположение, что массивное подавление нейрофизиологических путей ответственно за моторную дисфункцию, обнаруженную также у тщательно обследованных иммунокомпетентных мышей. [15]
Носовая патология
[ редактировать ]У птиц-хозяев T. regent достигает тканей носа, где спаривается и откладывает яйца. Крупная патология на этом участке представляет собой очаговые кровоизлияния, разбросанные по всей слизистой оболочке. Вокруг яиц присутствуют инфильтраты лимфоцитов и на более поздних стадиях формируются даже гранулемы, содержащие лимфоциты, эозинофилы и гетерофилы. Подобные инфильтраты присутствуют вокруг свободных мирацидий, но образования гранулем не отмечено. Вблизи взрослых червей клеточной реакции не отмечено. [21]
Иммунный ответ у позвоночных-хозяев
[ редактировать ]Утки
[ редактировать ]Сведения о клеточном иммунном ответе на T. regent у уток довольно скудны. Клеточная инфильтрация пораженных участков кожи у повторно инфицированных уток была отмечена только без дальнейшей характеристики инфильтрирующих клеток. [33] В ЦНС эозинофилы и гетерофилы окружают паразита, но не препятствуют его миграции к конечной локализации в слизистой оболочке носа. [19]
Учитывая реакцию антител, антицеркариальный IgM достигает кульминации 15 DPI, а антицеркариальный IgY достигает пика 30 DPI. Утки, инфицированные в более старшем возрасте, имеют более высокие уровни антицеркариального IgY , чем у уток, инфицированных в более молодом возрасте. Однако уровни антицеркариального IgY в значительной степени не зависят от инфекционной дозы. Некоторые антигены хозяина, паразитов, специфически распознаваемые IgY рассматриваются как кандидаты для иммунодиагностики. [34]
Мыши
[ редактировать ]Инфекция проявляется как ранняя реакция гиперчувствительности I типа и поздняя фаза кожного воспаления.
Клеточный иммунный ответ представлен продукцией провоспалительных ( IL-1β , IL-6 и IL-12p40 ) и противовоспалительных ( IL-10 ) цитокинов в коже мышей, инфицированных впервые. [22] Лимфоциты из кожных дренирующих лимфатических узлов демонстрируют смешанную поляризацию Th1 / Th2 после воздействия антигенов паразита. [22] [35] Напротив, противовоспалительные IL-4 и IL-10 доминируют у повторно инфицированных мышей, которые также секретируют большое количество гистамина из тучных клеток . Лимфоциты из кожных дренирующих лимфатических узлов производят IL-4 и IL-5 после стимуляции антигенами паразита, что демонстрирует Th2- поляризацию иммунного ответа хозяина. [22]
В спинном мозге сильный клеточный иммунный ответ, состоящий из гранулоцитов , плазматических клеток , макрофагов и Т-клеток мышей развивается у иммунокомпетентных , особенно вокруг поврежденной шистосомулы. У мышей с дефицитом CD3 воспаление отсутствует или развивается лишь в легкой форме, которое сопровождается неврологическими симптомами из-за механического повреждения нервной ткани. [24] [26] [31] Активированная микроглия локализуется в миграционных путях шистосомул и в воспалительных очагах, содержащих остатки паразитов. Хотя предполагается, что микроглия участвует в разрушении шистосомул, она обладает противовоспалительным фенотипом М2 и скорее способствует восстановлению тканей. [15] Гипертрофированные астроциты расположены на миграционных путях и вблизи шистосомулы, что указывает на их роль в иммунном ответе и репарации тканей. [26] Было показано, что мышиные астроциты и микроглия продуцируют провоспалительные цитокины ( IL-6 и TNF-α ) и оксид азота после воздействия in vitro антигенов паразита, что подтверждает их роль в иммунном ответе хозяина. [36]
IgM Ответ антител направлен главным образом на углеводные эпитопы молекул паразита. [37] Паразитоспецифические IgG1 и IgG2a появляются уже через 7 ДПИ. [35] Высокие уровни IgG1 и IgG2b , но не IgG2a , специфичные в основном к белковым эпитопам гомогената церкария , обнаруживаются при 150 DPI у повторно инфицированных мышей. Уровень общего IgE повышается уже после 10 ДПИ и остается высоким до 150 ДПИ у реинфицированных мышей. [37] Увеличение продукции антигенспецифического IgG1 и общего IgE , но небольшое снижение антигенспецифического IgG2b подтверждает иммунную поляризацию Th2 у повторно инфицированных лиц. [22] [37]
Люди
[ редактировать ]Клиническое проявление иммунного ответа человека на инфекцию T. regenti известно как церкариальный дерматит ( также известный как зуд пловца ). [2] [23] У большинства людей (82% взрослых, 57% детей), перенесших церкариальный дерматит (вызванный неопределенными видами птичьих шистосом), наблюдаются повышенные уровни T. regent антигенспецифических IgG , но не IgE . Церкариальный гомогенат и экскреторно-секреторные продукты T. regenti индуцируют базофилов людей, не перенесших дерматит дегрануляцию церкариальный и высвобождение IL-4 в анамнезе . [37]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Горак П., Коларова Л., Дворжак Ю. (декабрь 1998 г.). «Trichobilharzia regenti n. sp. (Schistosomatidae, Bilharziellinae), новая носовая шистосома из Европы» . Паразит . 5 (4): 349–57. дои : 10.1051/паразит/1998054349 . ПМИД 9879557 .

- ^ Jump up to: а б с д Горак П., Микеш Л., Лихтенбергова Л., Скала В., Солданова М., Брант С.В. (январь 2015 г.). «Птичьи шистосомы и вспышки церкариального дерматита» . Обзоры клинической микробиологии . 28 (1): 165–90. дои : 10.1128/CMR.00043-14 . ПМЦ 4284296 . ПМИД 25567226 .
- ^ Кристиансен А.О., Олсен А., Бухманн К., Кания П.В., Нейсум П., Веннервальд Б.Дж. (март 2016 г.). «Молекулярное разнообразие птичьих шистосом датских пресноводных улиток». Паразитологические исследования . 115 (3): 1027–37. дои : 10.1007/s00436-015-4830-3 . ПМИД 26573519 . S2CID 18519058 .
- ^ Прютер Х., Ситко Дж., Кроне О. (март 2017 г.). «Имея в виду шистосомы птиц - первое обнаружение Bilharziella polonica (Kowalewski 1895) в нервной системе птиц». Паразитологические исследования . 116 (3): 865–870. дои : 10.1007/s00436-016-5359-9 . ПМИД 28012027 . S2CID 22625424 .
- ^ Жуэ Д., Скирниссон К., Коларова Л., Ферте Х. (сентябрь 2010 г.). «Окончательные хозяева и изменчивость Trichobilharzia regent в природных условиях». Паразитологические исследования . 107 (4): 923–30. дои : 10.1007/s00436-010-1953-4 . ПМИД 20556426 . S2CID 10383377 .
- ^ Скирниссон К., Коларжова Л., Хорак П., Ферте Х., Жуэ Д. (май 2012 г.). «Морфологические особенности носового кровяного сосальщика Trichobilharzia regenti (Schistosomatidae, Digenea) от естественно инфицированных хозяев». Паразитологические исследования . 110 (5): 1881–92. дои : 10.1007/s00436-011-2713-9 . ПМИД 22146993 . S2CID 18351593 .
- ^ Рудольфова Дж., Литтлвуд Д.Т., Ситко Дж., Хорак П. (июнь 2007 г.). «Птичьи шистосомы диких птиц Чехии и Польши» . Фолиа Паразитологическая . 54 (2): 88–93. дои : 10.14411/fp.2007.011 . ПМИД 17886736 .
- ^ Маршевска А, Стшала Т, Цичи А, Домбровска ГБ, Жбиковска Е (декабрь 2018 г.). «Агенты опасного для зуда меньшинства пловцов при вторжении Digenea Lymnaeidae в водоемы и первое сообщение о Trichobilharzia regenti в Польше» . Паразитологические исследования . 117 (12): 3695–3704. дои : 10.1007/s00436-018-6068-3 . ПМК 6224017 . ПМИД 30215139 .
- ^ Пикард Д., Жуссон О (сентябрь 2001 г.). «Генетическая изменчивость церкарий Schistosomatidae (Trematoda: Digenea), вызывающая зуд пловца в Европе» . Паразит . 8 (3): 237–242. дои : 10.1051/паразит/2001083237 . ПМИД 11584754 .
- ^ Корсуненко А.В., Хрисанфова Г.Г., Рысков А.П., Мовсесян С.О., Васильев В.А., Семенова СК (август 2010 г.). «Обнаружение шистосом европейских Trichobilharzia (T. Franki, T. szidati и T. regenti) на основе новых последовательностей генома». Журнал паразитологии . 96 (4): 802–6. дои : 10.1645/GE-2297.1 . ПМИД 20677938 . S2CID 20445108 .
- ^ Фахар М., Гобадитара М., Брант С.В., Карамян М., Гохардехи С., Бастани Р. (апрель 2016 г.). «Филогенетический анализ носовых шистосом птиц (Trichobilharzia) водоплавающих птиц в провинции Мазандаран, северный Иран». Международная Паразитология . 65 (2): 151–8. дои : 10.1016/j.parint.2015.11.009 . ПМИД 26631753 .
- ^ Jump up to: а б Ашрафи К., Нуроста А., Шарифдини М., Махмуди М.Р., Рахмати Б., Брант С.В. (декабрь 2018 г.). «Генетическое разнообразие носовой шистосомы птиц, вызывающей церкариальный дерматит на черноморско-средиземноморском миграционном пути». Паразитологические исследования . 117 (12): 3821–3833. дои : 10.1007/s00436-018-6087-0 . ПМИД 30343420 . S2CID 53043270 .
- ^ «Лаборатория гельминтологии» . Чехия: Карлов университет в Праге . Проверено 20 марта 2016 г.
- ^ Леонтович Р., Янг Н.Д., Корхонен П.К., Холл Р.С., Тан П., Микеш Л., Кашни М., Горак П., Гассер Р.Б. (февраль 2016 г.). «Сравнительное транскриптомное исследование выявило уникальные молекулярные адаптации нейропатогенной трихобилгарции к вторжению и паразитированию на ее окончательном птичьем хозяине» . PLOS Забытые тропические болезни . 10 (2): e0004406. дои : 10.1371/journal.pntd.0004406 . ПМЦ 4749378 . ПМИД 26863542 .
- ^ Jump up to: а б с д Махачек Т, Леонтович Р, Шмидова Б, Майер М, Вондрачек О, Войтехова И и др. (февраль 2022 г.). «Механизмы иммунного ответа хозяина и гельминтоиндуцированная патология при нейроинвазии Trichobilharzia regenti (Schistosomatidae) у мышей» . PLoS Патогены . 18 (2): e1010302. дои : 10.1371/journal.ppat.1010302 . ПМЦ 8849443 . ПМИД 35120185 .
- ^ Гунева К, Кашни М, Хампл В, Леонтович Р, Кубена А, Микеш Л, Горак П (сентябрь 2012 г.). «Radix spp.: Идентификация промежуточных хозяев трематод в Чехии» . Акта Паразитологическая . 57 (3): 273–84. дои : 10.2478/s11686-012-0040-7 . ПМИД 22875675 . S2CID 235463 .
- ^ Пештова Ю (2015). Дифференцировка тотипотентных зародышевых клеток у личинок шистосом птиц [на чешском языке] (Магистерская диссертация). Прага, Чехия: Карлов университет.
- ^ Ржимначова Ю, Микеш Л, Турьяникова Л, Булантова Ю, Горак П (15 марта 2017 г.). «Изменения поверхностного гликозилирования и отщепления гликокаликса у Trichobilharzia regenti (Schistosomatidae) во время трансформации церкарий в шистосомулум» . ПЛОС ОДИН . 12 (3): e0173217. Бибкод : 2017PLoSO..1273217R . дои : 10.1371/journal.pone.0173217 . ПМК 5351870 . ПМИД 28296924 .
- ^ Jump up to: а б с д Хорак П., Дворжак Ю., Коларова Л., Трефил Л. (декабрь 1999 г.). «Trichobilharzia regenti, возбудитель центральной нервной системы птиц и млекопитающих». Паразитология . 119 (6): 577–81. дои : 10.1017/s0031182099005132 . ПМИД 10633919 . S2CID 33919554 .
- ^ Jump up to: а б с д и Градкова К., Горак П. (июнь 2002 г.). «Нейротропное поведение Trichobilharzia regenti у уток и мышей». Журнал гельминтологии . 76 (2): 137–41. дои : 10.1079/JOH2002113 . ПМИД 12015826 . S2CID 6387025 .
- ^ Jump up to: а б с Чанова М., Горак П. (июнь 2007 г.). «Терминальная фаза шистосомоза птиц, вызванного Trichobilharzia regenti (Schistosomatidae) у уток (Anas platyrhynchos f. Domestica)» . Фолиа Паразитологическая . 54 (2): 105–7. дои : 10.14411/fp.2007.014 . ПМИД 17886739 .
- ^ Jump up to: а б с д и ж г Курилова П., Хогг К.Г., Коларова Л., Маунтфорд А.П. (март 2004 г.). «Церкариальный дерматит, вызванный шистосомами птиц, включает кожные реакции гиперчувствительности как немедленной, так и поздней фазы» . Журнал иммунологии . 172 (6): 3766–74. дои : 10.4049/jimmunol.172.6.3766 . ПМИД 15004181 .
- ^ Jump up to: а б с Коларжова Л., Горак П., Скирниссон К., Маречкова Х., Дёнхофф М. (август 2013 г.). «Церкариальный дерматит – забытое аллергическое заболевание». Клинические обзоры по аллергии и иммунологии . 45 (1): 63–74. дои : 10.1007/s12016-012-8334-y . ПМИД 22915284 . S2CID 32704545 .
- ^ Jump up to: а б с д и ж Курилова П., Сыручек М., Коларова Л. (май 2004 г.). «Тяжесть патологий мышей, вызванных шистосомами птиц Trichobilharzia regenti, в зависимости от иммунного статуса хозяина». Паразитологические исследования . 93 (1): 8–16. дои : 10.1007/s00436-004-1079-7 . ПМИД 15034785 . S2CID 19462770 .
- ^ Jump up to: а б с Блазова К., Горак П. (сентябрь 2005 г.). «Trichobilharzia regenti: различия в развитии естественных и аномальных хозяев». Международная Паразитология . 54 (3): 167–172. дои : 10.1016/j.parint.2005.03.003 . ПМИД 15908263 .
- ^ Jump up to: а б с д и ж г час я Лихтенбергова Л., Лассманн Х., Джонс М.К., Коларжова Л., Горак П. (август 2011 г.). «Trichobilharzia regenti: иммунный ответ хозяина в патогенезе нейроинфекции у мышей». Экспериментальная паразитология . 128 (4): 328–35. дои : 10.1016/j.exppara.2011.04.006 . ПМИД 21554878 .
- ^ Jump up to: а б Булантова Ю., Махачек Т., Панска Л., Крейчи Ф., Карч Дж., Ярлинг Н., Сагафи С., Додт Х.У., Горак П. (апрель 2016 г.). «Trichobilharzia regenti (Schistosomatidae): методы трехмерной визуализации для характеристики миграции личинок через ЦНС позвоночных». Микрон . 83 : 62–71. дои : 10.1016/j.micron.2016.01.009 . ПМИД 26897588 .
- ^ Майкс Л., Зидкова Л., Касны М., Дворжак Дж., Горак П. (июнь 2005 г.). «Стимуляция опорожнения пенетрационной железы in vitro церкариями Trichobilharzia szidati и T. regenti (Schistosomatidae). Количественный сбор и частичная характеристика продуктов». Паразитологические исследования . 96 (4): 230–41. дои : 10.1007/s00436-005-1347-1 . ПМИД 15868186 . S2CID 765928 .
- ^ Касны М., Майкс Л., Далтон Дж.П., Маунтфорд А.П., Хорак П. (октябрь 2007 г.). «Сравнение активности цистеинпептидазы у Trichobilharzia regenti и Schistosoma mansoni cercariae» (PDF) . Паразитология . 134 (Часть 11): 1599–609. дои : 10.1017/S0031182007002910 . hdl : 10453/5909 . ПМИД 17517170 . S2CID 14440074 .
- ^ Долецкова К., Касны М., Майкс Л., Картрайт Дж., Едельски П., Шнайдер Э.Л., Дворак Дж., Маунтфорд А.П., Крейк К.С., Хорак П. (январь 2009 г.). «Функциональное выражение и характеристика цистеиновой пептидазы инвазивной стадии нейропатогенных шистосом Trichobilharzia regenti» . Международный журнал паразитологии . 39 (2): 201–11. дои : 10.1016/j.ijpara.2008.06.010 . ПМЦ 2625449 . ПМИД 18708063 .
- ^ Jump up to: а б с Коларова Л., Хорак П., Када Ф (август 2001 г.). «Гистопатология ЦНС и назальные инфекции, вызванные Trichobilharzia regenti у позвоночных». Паразитологические исследования . 87 (8): 644–50. дои : 10.1007/s004360100431 . ПМИД 11511002 . S2CID 2895174 .
- ^ Дворак Дж., Делькруа М., Росси А., Вопаленский В., Посписек М., Сединова М., Майкес Л., Саджид М., Сали А., МакКерроу Дж.Х., Хорак П., Кэффри Ч.Р. (июль 2005 г.). «Множественные изоформы катепсина B у шистосомул Trichobilharzia regenti: идентификация, характеристика и предполагаемая роль в миграции и питании». Международный журнал паразитологии . 35 (8): 895–910. дои : 10.1016/j.ijpara.2005.02.018 . ПМИД 15950230 .
- ^ Печ В. (2013). Пероральные инфекции птиц и млекопитающих нейропатогенным сосальщиком Trichobilharzia regenti (магистерская диссертация). Чехия: Карлов университет.
- ^ Турьяникова Л., Микеш Л., Пецкова М., Горак П. (июль 2015 г.). «Антитильный ответ окончательных хозяев против антигенов двух стадий жизни нейропатогенной шистосомы Trichobilharzia regenti» . Паразиты и переносчики . 8 (1): 400. doi : 10.1186/s13071-015-1007-y . ПМЦ 4517386 . ПМИД 26216102 .
- ^ Jump up to: а б Майер М., Махачек Т., Сукеникова Л., Хрды Ю., Горак П. (июнь 2020 г.). «Периферический иммунный ответ мышей, инфицированных нейропатогенной шистосомой». Иммунология паразитов . 42 (6): e12710. дои : 10.1111/pim.12710 . ПМИД 32145079 . S2CID 212629117 .
- ^ Махачек Т., Панска Л., Дворжакова Х., Горак П. (ноябрь 2016 г.). «Продукция оксида азота и цитокинов глиальными клетками, подвергшимися in vitro воздействию нейропатогенных шистосом Trichobilharzia regenti» . Паразиты и переносчики . 9 (1): 579. doi : 10.1186/s13071-016-1869-7 . ПМК 5109812 . ПМИД 27842570 .
- ^ Jump up to: а б с д Лихтенбергова Л, Кольбекова П, Курилова П, Касны М, Майкс Л, Хаас Х, Шрамм Г, Горак П, Коларова Л, Маунтфорд А.П. (01 ноября 2008 г.). «Реакция антител, индуцированная антигенами Trichobilharzia regenti у мышей и людей с церкариальным дерматитом» . Иммунология паразитов . 30 (11–12): 585–95. дои : 10.1111/j.1365-3024.2008.01059.x . ПМК 2680328 . ПМИД 19067839 .