Макрофаг
| Макрофаг | |
|---|---|
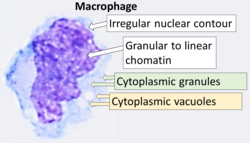 Цитология макрофага с типичными особенностями. Райт пятна . | |
| Подробности | |
| Произношение | / Mac (ʊ) feɪdʒ / |
| Система | Иммунная система |
| Функция | Фагоцитоз |
| Идентификаторы | |
| латинский | макрофагоцит |
| Аббревиатура (ы) | M φ , mφ |
| Сетка | D008264 |
| Тур | H2.00.03.0.01007 |
| FMA | 63261 |
| Анатомические термины микроанатомии | |
Макрофаги ( / ˈ M æ K r oʊ f eɪ dʒ / ; сокращенно M φ , Mφ или Mp ) являются типом лейкоцитов врожденной иммунной системы , которая охватывает и переваривает патогены, такие как раковые клетки , микробы , клеточный мусор, и посторонние вещества, которые не имеют белков, которые специфичны для здоровых клеток тела на их поверхности. [ 1 ] [ 2 ] Этот процесс называется фагоцитозом , который действует для защиты хозяина от инфекции и травм. [ 3 ]
Макрофаги обнаружены практически во всех тканях, [ 4 ] где они патрулируют для потенциальных патогенов с помощью амебоидного движения . Они принимают различные формы (с различными именами) по всему телу (например, гистиоциты , клетки Купфера , альвеолярные макрофаги , микроглия и другие), но все они являются частью мононуклеарной фагоцитарной системы . Помимо фагоцитоза, они играют решающую роль в неспецифической защите ( врожденный иммунитет ), а также помогают инициировать конкретные защитные механизмы ( адаптивный иммунитет ) путем рекрута других иммунных клеток, таких как лимфоциты . Например, они важны в качестве докладчиков антигена для Т -клеток . У людей дисфункциональные макрофаги вызывают тяжелые заболевания, такие как хронические гранулематозные заболевания , которые приводят к частым инфекциям.
Помимо увеличения воспаления и стимуляции иммунной системы, макрофаги также играют важную противовоспалительную роль и могут уменьшить иммунные реакции посредством высвобождения цитокинов . Макрофаги, которые поощряют воспаление, называются макрофагами M1, тогда как те, которые уменьшают воспаление и стимулируют восстановление тканей, называются макрофагами M2. [ 5 ] Эта разница отражается в их метаболизме; M1 -макрофаги обладают уникальной способностью метаболизировать аргинин с молекулой «убийца» оксида азота , тогда как макрофаги M2 обладают уникальной способностью метаболизировать аргинин с орнитином «восстановления» молекулы . [ 6 ] Тем не менее, эта дихотомия была недавно подвергнута сомнению, поскольку была обнаружена дальнейшая сложность. [ 7 ]
Человеческие макрофаги имеют около 21 микрометра (0,00083 дюйма) в диаметре [ 8 ] и продуцируются дифференцировкой моноцитов в тканях. Они могут быть идентифицированы с использованием проточной цитометрии или иммуногистохимического окрашивания с помощью их специфической экспрессии белков, таких как CD14 , CD40 , CD11B , (человек), лизоцим M, MAC-1/MAC-3 и CD68 (человека) , CD64, F4/80 (мыши)/ EMR1 (человек), лизоцим M, MAC-1 /MAC-3 и лизоцим M, MAC-1/MAC-3 и CD68 (человек), лизоцим M, MAC-1/MAC-3 и CD68 CD68 (человек Полем [ 9 ]
В 1884 году макрофаги были впервые обнаружены и названы Эли Метчникофф , российской империи. [ 10 ] [ 11 ]
Структура
[ редактировать ]Типы
[ редактировать ]
Большинство макрофагов размещаются в стратегических точках, где может возникнуть микробная инвазия или накопление иностранных частиц. Эти клетки вместе как группа известны как мононуклеарная система фагоцитов и ранее были известны как ретикулоендотелиальная система. Каждый тип макрофагов, определяемый его местоположением, имеет определенное имя:
| Название ячейки | Анатомическое расположение |
| Макрофаги жировой ткани | Жировая ткань (жир) |
| Моноциты | Костный мозг / кровь |
| Купффер -клетки | Печень |
| Синусовые гистиоциты | Лимфатические узлы |
| Альвеолярные макрофаги (пылевые клетки) | Легочные альвеолы |
| Макрофаги ткани (гистиоциты), ведущие к гигантским клеткам | Соединительная ткань |
| Микроглия | Центральная нервная система |
| Хофбауэр -клетки | Плацента |
| Внутригломерулярные мезангиальные клетки [ 12 ] | Почка |
| Остеокласты [ 13 ] | Кость |
| Лангерганс Клетки | Кожа |
| Эпителиоидные клетки | Гранулемы |
| Макрофаги красной пульпы ( синусоидальные слизистые клетки) | Красная пульпа селезенки |
| Перитонеальные макрофаги | Брюшная полость |
| периваскулярные макрофаги [ 14 ] | тесно связано с кровеносными сосудами |
Исследования, касающиеся клеток Купффера, затруднены, потому что у людей клетки Купффера доступны только для иммуногистохимического анализа биопсий или вскрытий. От крыс и мышей их трудно изолировать, и после очистки можно получить только приблизительно 5 миллионов клеток от одной мыши.
Макрофаги могут экспрессировать паракриновые функции в органах, которые специфичны для функции этого органа. Например, в яичке макрофаги способны взаимодействовать с клетками Leydig, секретируя 25-гидроксихолестерин , оксистерол , который может быть преобразован в тестостерон соседними клетками Leydig. [ 15 ] Кроме того, макрофаги яичка могут участвовать в создании иммунной привилегированной среды в яичке и в опосредовании бесплодия во время воспаления яичка.
Макрофаги резидентов сердца участвуют в электрической проводимости посредством связи с пробелом с миоцитами сердца . [ 16 ]
Макрофаги могут быть классифицированы на основе фундаментальной функции и активации. Согласно этой группировке, существуют классически активированные (M1) макрофаги , макрофаги, подвергающегося ране (также известные как альтернативно-активированные (M2) макрофаги ) и регуляторные макрофаги (MREGS). [ 17 ]
Разработка
[ редактировать ]Макрофаги, которые проживают в здоровых тканях взрослых, происходят либо из циркулирующих моноцитов , либо устанавливаются до рождения, а затем поддерживаются во время жизни взрослого независимо от моноцитов. [ 18 ] [ 19 ] Напротив, большинство макрофагов, которые накапливаются на больных участках, обычно происходят из циркулирующих моноцитов. [ 20 ] Экстравазация лейкоцитов описывает проникновение моноцитов в поврежденную ткань через эндотелий кровеносных сосудов , когда они становятся макрофагами. Моноциты притягиваются к поврежденному участку химическими веществами посредством хемотаксиса , запускаемых рядом стимулов, включая поврежденные клетки, патогены и цитокины, выделяемые макрофагами, уже на месте. На некоторых сайтах, таких как яичка, было показано, что макрофаги заполняют орган посредством пролиферации. [ 21 ] В отличие от короткоживущих нейтрофилов , макрофаги выживают дольше в организме, до нескольких месяцев.
Функция
[ редактировать ]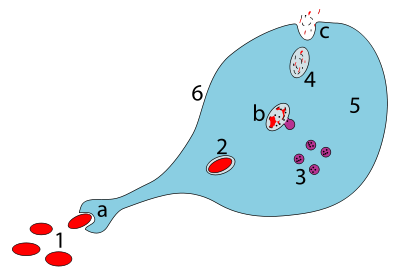
а Проглатывание через фагоцитоз, образуется фагосома
беременный Слияние лизосом с фагосомой создает фаголизосому ; Патоген разбит ферментами
в Отходы изгнаны или ассимилируются (последнее не изображено)
Части:
1. Патогены
2. фагосома
3. Лизосомы
4. Отходы
5. Цитоплазма
6. клеточная мембрана
Фагоцитоз
[ редактировать ]Макрофаги представляют собой профессиональные фагоциты и являются высокоспециализированными по удалению умирающих или мертвых клеток и клеточного мусора. Эта роль важна в хроническом воспалении, так как на ранних стадиях воспаления преобладают нейтрофилы, которые принимаются макрофагами, если они достигают совершеннолетия (см. CD31 для описания этого процесса). [ 22 ]
Нейтрофилы сначала привлечены к месту, где они выполняют свою функцию и умирают, прежде чем они или их нейтрофильные внеклеточные ловушки фагоцитируются макрофагами. [ 22 ] [ 23 ] На месте, первая волна нейтрофилов, после процесса старения и после первых 48 часов, стимулирует появление макрофагов, в результате чего эти макрофаги будут проходить пожилые нейтрофилы. [ 22 ]
Удаление умирающих клеток, в большей степени, обрабатывается фиксированными макрофагами , которые будут оставаться в стратегических местах, таких как легкие, печень, нервная ткань , кость, селезенка и соединительная ткань, проглатывание иностранных материалов, таких как патогены и рекрутирующие дополнительные макрофаги При необходимости. [ 24 ]
Когда макрофаг принимает патоген, патоген попадает в фагосому , которая затем сливается с лизосомой . В рамках фаголизосомы ферменты и токсичные пероксиды переваривают патоген. Однако некоторые бактерии, такие как Mycobacterium Tuberculosis , стали устойчивыми к этим методам пищеварения. Тифоидные лосомоны индуцируют свой собственный фагоцитоз макрофагами хозяина in vivo и ингибируют расщепление лизосомальным действием, тем самым используя макрофаги для их собственной репликации и вызывая апоптоз макрофагов. [ 25 ] Макрофаги могут переваривать более 100 бактерий, прежде чем они наконец умрут из -за своих собственных пищеварительных соединений.
Роль в врожденном иммунном ответе
[ редактировать ]Когда патоген вторгается, макрофаги, резидентные ткани, являются одними из первых клеток, которые реагируют. [ 26 ] Две основные роли резидентных макрофагов ткани заключаются в фагоцитозе входящего антигена и выделять провоспалительные цитокины, которые вызывают воспаление и рекрутируют другие иммунные клетки в сайт. [ 27 ]
Фагоцитоз патогенов
[ редактировать ]
Макрофаги могут усваивать антигены посредством рецепторного опосредованного фагоцитоза. [ 28 ] Макрофаги имеют широкий спектр рецепторов распознавания паттернов (PRR), которые могут распознавать микроб-ассоциированные молекулярные паттерны (MAMPS) из патогенов. Многие PRR, такие как рецепторы, похожие на Toll (TLR), рецепторы мусорщиков (SRS), рецепторы лектина C-типа, среди прочего, распознают патогены фагоцитоза. [ 28 ] Макрофаги могут также распознавать патогены для фагоцитоза косвенно через Opsonins , которые представляют собой молекулы, которые прикрепляются к патогенам и отмечают их для фагоцитоза. [ 29 ] Опсонины могут вызывать более сильную адгезию между макрофагом и патогеном во время фагоцитоза, следовательно, опсонины имеют тенденцию усилить фагоцитарную активность макрофагов. [ 30 ] Обе белки комплемента и антитела могут связываться с антигенами и их опсонизировать. Макрофаги имеют комплемента рецептор 1 (CR1) и 3 (CR3), которые распознают белки комплемента, связанные с патогеном C3B и IC3B, соответственно, а также кристаллируемые фрагментные γ-рецепторы (FCγR), которые распознают кристаллизованную (FC) область антигена-связанного иммуноглобулина иммуноглобулина , которые распознают фрагмент-кристаллизуем G (IgG) антитела. [ 29 ] [ 31 ] При фагоцитозировании и переваривании патогенов макрофаги проходят дыхательный взрыв , где потребляется больше кислорода для обеспечения энергии, необходимой для производства активных форм кислорода (АФК) и других противомикробных молекул, которые переваривают потребляемые патогены. [ 27 ] [ 32 ]
Секреция цитокинов
[ редактировать ]Распознавание MAMP с помощью PRR может активировать резидентные макрофаги тканей для выделения провоспалительных цитокинов, которые рекрутируют другие иммунные клетки. Среди PRR TLR играют важную роль в трансдукции сигнала, ведущей к производству цитокинов. [ 28 ] Связывание MAMP с TLR запускает серию последующих событий, которые в конечном итоге активирует фактор транскрипции NF-κB и приводит к транскрипции генов для нескольких провоспалительных цитокинов, включая IL-1β , IL-6 , TNF-α , IL-12B и Интерфероны типа I, такие как IFN-α и IFN-β. [ 33 ] Системно IL-1β, IL-6 и TNF-α вызывают лихорадку и инициируют острый фазовый ответ, в котором печень выделяет белки острой фазы . [ 26 ] [ 27 ] [ 34 ] Локально IL-1β и TNF-α вызывают вазодилатацию, где промежутки между эпителиальными клетками кровеносного сосуда расширяются, и повышенная регуляция молекул адгезии клеточной поверхности на эпителиальных клетках для индуцирования экстравазации лейкоцитов . [ 26 ] [ 27 ]
Нейтрофилы являются одними из первых иммунных клеток, рекрутированных макрофагами, для выхода из крови через экстравазацию и достижение места инфекции. [ 34 ] Макрофаги выделяют многие хемокины, такие как CXCL1 , CXCL2 и CXCL8 (IL-8), которые привлекают нейтрофилы к месту инфекции. [ 26 ] [ 34 ] После того, как нейтрофилы завершили фагоцитозирование и очистку антигена в конце иммунного ответа, они подвергаются апоптозу, а макрофаги рекрутируются из моноцитов крови, чтобы помочь очистить апоптотический мусор. [ 35 ]
Макрофаги также рекрутируют другие иммунные клетки, такие как моноциты, дендритные клетки, природные клетки -киллеры, базофилы, эозинофилы и Т -клетки, такие как хемокины, такие как CCL2 , CCL4 , CCL5 , CXCL8 , CXCL9 , CXCL10 и CXCL11 . [ 26 ] [ 34 ] Наряду с дендритными клетками макрофаги помогают активировать природные киллеры (NK) клетки посредством секреции интерферонов I типа (IFN-α и IFN-β) и IL-12 . IL-12 действует с IL-18 для стимуляции продукции провоспалительного цитокинового интерферона гамма (IFN-γ) NK-клетками, которые служат важным источником IFN-γ до активации адаптивной иммунной системы. [ 34 ] [ 36 ] IFN-γ усиливает врожденный иммунный ответ, индуцируя более агрессивный фенотип в макрофагах, позволяя макрофагам более эффективно убивать патогены. [ 34 ]
Некоторые из Т -клеточных хемоаттрактантов, секретируемых макрофагами, включают CCL5 , CXCL9 , CXCL10 и CXCL11 . [ 26 ]
Роль в адаптивном иммунитете
[ редактировать ]
Взаимодействие с CD4 + T Helper Cells
[ редактировать ]Макрофаги представляют собой профессиональный антиген, представляющие клетки (APC), что означает, что они могут представлять пептиды из фагоцитозированных антигенов на основных молекулах гистосовместимости (MHC) II на их клеточной поверхности для T -Helper -клеток. [ 38 ] Макрофаги не являются первичными активаторами наивных Т -вспомогательных клеток, которые никогда ранее активировались, поскольку макрофаги, резидентные ткани, не перемещаются в лимфатические узлы, где проживают наивные Т -хелперные клетки. [ 39 ] [ 40 ] Хотя макрофаги также обнаруживаются в вторичных лимфоидных органах, таких как лимфатические узлы, они не находятся в зонах Т -клеток и не эффективны в активации наивных Т -хелпер -клеток. [ 39 ] Макрофаги в лимфоидных тканях больше вовлечены в приема антигенов и предотвращение их вступления в кровь, а также для получения мусора из апоптотических лимфоцитов. [ 39 ] [ 41 ] Следовательно, макрофаги взаимодействуют в основном с ранее активированными Т -хелперными клетками, которые покинули лимфатический узел и прибыли в место инфекции или с Т -клетками памяти, резидентной ткани. [ 40 ]
Макрофаги подают оба сигнала, необходимые для активации T-Helper Cell: 1) макрофаги присутствуют молекулу MHC класса II класса антигена, которые должны быть распознаны соответствующим рецептором Т-клеток (TCR) и 2) распознавание патогенов PRR индуцируют макрофаги для регулирования CO). -stimulatory Molecules CD80 и CD86 (также известные как B7 ), которые связываются с CD28 на клетках T-Helper для подачи костимуляторного сигнала. [ 34 ] [ 38 ] Эти взаимодействия позволяют T -вспомогательным клеткам выполнять полную эффекторную функцию и обеспечивать Т -хелперные клетки с постоянными сигналами выживаемости и дифференцировки, мешающие им проходить апоптоз из -за отсутствия передачи сигналов TCR. [ 38 ] Например, передача сигналов IL-2 в Т-клетках активирует экспрессию антиапоптотического белка Bcl-2 , но выработка Т-клеток IL-2 и высокоаффинно-рецептор IL-2 IL-2RA требуют постоянного сигнала от распознавания TCR MHC-связанный антиген. [ 34 ] [ 42 ]
Активация
[ редактировать ]Макрофаги могут достигать различных фенотипов активации посредством взаимодействия с различными подмножествами T -Helper Cells, такими как T H 1 и T H 2. [ 17 ] Хотя существует широкий спектр фенотипов активации макрофагов, есть два основных фенотипа, которые обычно признаются. [ 17 ] Это классически активированные макрофаги или макрофаги M1, а также альтернативно активированные макрофаги или макрофаги M2. Макрофаги M1 являются провоспалительными, в то время как макрофаги M2 в основном противовоспалительны. [ 17 ]
Классический
[ редактировать ]Клетки T H 1 играют важную роль в классической активации макрофагов как часть иммунного ответа типа 1 против внутриклеточных патогенов (таких как внутриклеточные бактерии ), которые могут выжить и повторять внутри клеток -хозяев, особенно те патогены, которые повторяются даже после фагоцитации макрофагами. [ 43 ] После того, как TCR TC-клеток T - 1 распознает специфические антиген-пептидные молекулы MHC класса II на макрофагах, клетки T H 1) секретируют IFN-γ и 2) активируют экспрессию лиганда CD40 (CD40L), который связывается с CD40 на макрофагах. Полем [ 44 ] [ 34 ] Эти 2 сигнала активируют макрофаги и усиливают их способность убивать внутриклеточные патогены за счет увеличения продукции антимикробных молекул, таких как оксид азота (NO) и супероксид (O 2- ). [ 26 ] [ 34 ] Это усиление антимикробной способности макрофагов с помощью клеток T H 1 известно как классическая активация макрофагов, а активированные макрофаги известны как классически активированные макрофаги или макрофаги M1. Макрофаги M1 в приверженности усиливают молекулы B7 и презентацию антигена с помощью молекул MHC класса II, чтобы обеспечить сигналы, которые поддерживают помощь Т -клеток. [ 44 ] Активация макрофагов T H 1 и M1 является положительной петлей обратной связи, причем IFN-γ из T H клеток 1 активирует экспрессию CD40 на макрофагах; Взаимодействие между CD40 на макрофагах и CD40L на Т-клетках активирует макрофаги, чтобы секретировать IL-12; и IL-12 способствует большей секреции IFN-γ из T H 1. клеток [ 34 ] [ 44 ] Первоначальный контакт между MHC II II и TCR, связанным с антигеном, и TCR, который служит контактной точкой между двумя клетками, где большая часть секреции IFN-γ и CD-40L на Т-клетках концентрируется, поэтому только макрофаги непосредственно взаимодействуют с клетками T H 1, являются клетками T H 1. вероятно, будет активирован. [ 34 ]
В дополнение к активации макрофагов M1, T H клетки 1 экспрессируют Fas-лиганд (FASL) и бета-лимфотоксин (LT-β), чтобы помочь убить хронически инфицированные макрофаги, которые больше не могут убивать патогены. [ 34 ] Убивание хронически инфицированных макрофагов высвобождает патогены во внеклеточном пространстве, которое затем может быть убито другими активированными макрофагами. [ 34 ] Клетки T H 1 также помогают рекрутировать больше моноцитов, предшественника макрофагов, в сайт инфекции. T H Секреция 1 TNF-α и LT-α, чтобы облегчить кровеносные сосуды для моноцитов и выходить. [ 34 ] T H 1 Секреция CCL2 в качестве хемоаттрактанта для моноцитов. IL-3 и GM-CSF, высвобождаемые клетками T H 1, стимулируют больше продукции моноцитов в костном мозге. [ 34 ]
Когда внутриклеточные патогены не могут быть устранены, например, в случае микобактерии туберкулеза , патоген содержится посредством образования гранулемы , агрегации инфицированных макрофагов, окруженных активированными Т -клетками. [ 45 ] Макрофаги, граничащие с активированными лимфоцитами, часто сливаются, образуя многоядерные гигантские клетки, которые, по -видимому, обладают повышенной антимикробной способностью из -за их близости к клеткам T H 1, но со временем клетки в центре начинают умирать и образовывать некротическую ткань. [ 40 ] [ 45 ]
Альтернатива
[ редактировать ]Клетки T H 2 играют важную роль в альтернативной активации макрофагов как часть иммунного ответа 2 типа против больших внеклеточных патогенов, таких как гельминт . [ 34 ] [ 46 ] Клетки T H 2 секретируют IL-4 и IL-13, которые активируют макрофаги, чтобы стать макрофагами M2, также известные как альтернативно активированные макрофаги. [ 46 ] [ 47 ] M2 макрофаги экспрессируют аргиназу-1 , фермент, который превращает аргинин в орнитин и мочевину . [ 46 ] Орнитин помогает увеличить сокращение гладких мышц, чтобы выгнать червя, а также участвует в восстановлении тканей и раны. Орнитин может быть дополнительно метаболизирован на пролин , что необходимо для синтеза коллагена . [ 46 ] M2 макрофаги также могут уменьшить воспаление путем продуцирования антагониста рецептора IL-1 (IL-1RA) и рецепторов IL-1, которые не приводят к нижней воспалительной передаче сигналов (IL-1RII). [ 34 ] [ 48 ]
Взаимодействие с CD8 + цитотоксические Т -клетки
[ редактировать ]Другая часть активации адаптивного иммунитета включает стимулирование CD8 + через перекрестное представление антигенов пептидов на молекулах класса I MHC. Исследования показали, что провоспалительные макрофаги способны к перекрестному представлению антигенов на молекулах класса I MHC, но играет ли макрофаги поперечная презентация в наивном или память CD8 + Активация Т -клеток до сих пор неясна. [ 27 ] [ 49 ] [ 41 ]
Взаимодействия с B -клетками
[ редактировать ]Было показано, что макрофаги секретируют цитокины BAFF и April, которые важны для переключения изотипа плазменных клеток. Апрель и IL-6, секретируемые предшественниками макрофагов в костном мозге, помогают поддерживать выживание плазматических клеток, дома, подходящих к костному мозгу. [ 50 ]
Подтипы
[ редактировать ]
Есть несколько активированных форм макрофагов. [ 17 ] Несмотря на спектр способов активации макрофагов, существуют две основные группы, обозначенные M1 и M2 . M1 макрофаги: как упоминалось ранее (ранее называлось классически активированными макрофагами), [ 52 ] M1 "Killer" макрофаги активируются LPS и IFN-гамма и секретируют высокие уровни IL-12 и низкие уровни IL-10 . M1 макрофаги выполняют провоспалительные, бактерицидные и фагоцитарные функции. [ 53 ] Напротив, обозначение M2 восстановления »(также называемое альтернативно активированными макрофагами) в целом относится к макрофагам, которые функционируют в конструктивных процессах, таких как заживление ран и восстановление тканей, и те, которые отключают повреждение активации иммунной системы, продуцируя противовоспалительные цитокины, такие как такие IL-10 . M2-это фенотип резидентных тканевых макрофагов, который может быть дополнительно повышен с помощью IL-4 . M2 макрофаги продуцируют высокий уровень IL-10, TGF-бета и низкий уровень IL-12. Связанные с опухолью макрофаги в основном из фенотипа М2 и, по-видимому, активно способствуют росту опухоли. [ 54 ]
Макрофаги существуют в различных фенотипах, которые определяются роли, которую они играют в созревании раны. Фенотипы могут быть преимущественно разделены на две основные категории; M1 и M2. M1 макрофаги являются доминирующим фенотипом, наблюдаемым на ранних стадиях воспаления, и активируются четырьмя ключевыми медиаторами: интерферон-γ (IFN-γ), фактор некроза опухоли (TNF) и связанные с повреждением молекулярные паттерны (DEAMS). Эти медиаторные молекулы создают провоспалительный ответ, который в ответ продуцирует провоспалительные цитокины, такие как интерлейкин-6 и TNF. В отличие от макрофагов M1, макрофаги M2 выделяют противовоспалительный ответ посредством добавления интерлейкина-4 или интерлейкина-13. Они также играют роль в заживлении ран и необходимы для реваскуляризации и реэпителизации. M2 макрофаги разделены на четыре основных типа, основанные на их ролях: M2A, M2B, M2C и M2D. То, как фенотипы M2 определяются, все еще не подходит для обсуждения, но исследования показали, что их среда позволяет им приспосабливаться к тому, какой фенотип наиболее подходит для эффективного заживления раны. [ 53 ]
M2 макрофаги необходимы для сосудистой стабильности. Они продуцируют сосудистый эндотелиальный фактор роста-А и TGF-β1 . [ 53 ] Существует сдвиг фенотипа с макрофагов M1 к M2 в острых ранх, однако этот сдвиг нарушен для хронических ран. Эта дисрегуляция приводит к недостаточным макрофагам M2 и соответствующим факторам роста, которые помогают при восстановлении раны. Из-за отсутствия этих факторов роста/противовоспалительных цитокинов и переизбыток провоспалительных цитокинов из макрофагов M1 хронические раны не могут своевременно заживать. Обычно, после нейтрофилов есть мусор/патогенные микроорганизмы, они выполняют апоптоз и удаляются. На этом этапе воспаление не требуется, и M1 подвергается переходу на M2 (противовоспалительный). Тем не менее, дисрегуляция возникает, когда макрофаги M1 не способны/не являются фагоцитозными нейтрофилами, которые подвергались апоптозу, приводящим к увеличению миграции макрофагов и воспаления. [ 53 ]
Как макрофаги M1 и M2 играют роль в продвижении атеросклероза . M1 макрофаги способствуют атеросклерозу за счет воспаления. M2 макрофаги могут удалять холестерин из кровеносных сосудов, но когда холестерин окисляется, макрофаги M2 становятся апоптотическими пенистыми клетками, способствующими атероматозным бляшкам атеросклероза. [ 55 ] [ 56 ]
Роль в регенерации мышц
[ редактировать ]Первым шагом к пониманию важности макрофагов в восстановлении, росте и регенерации мышц является то, что существуют две «волны» макрофагов с появлением разрушаемого использования мышц - субпопуляции, которые делают и не оказывают непосредственного влияния на восстановление мышц. Первоначальная волна - это фагоцитарная популяция, которая появляется в периоды увеличения использования мышц, которые достаточны для того, чтобы вызвать лизис мышечной мембраны и воспаление мембраны, которые могут вводить и разрушать содержимое поврежденных мышечных волокон. [ 57 ] [ 58 ] [ 59 ] Эти раноинвадирующие фагоцитарные макрофаги достигают самой высокой концентрации примерно через 24 часа после появления какой-либо формы повреждения мышечных клеток или перезагрузки. [ 60 ] Их концентрация быстро снижается через 48 часов. [ 58 ] Вторая группа-это нефагоцитарные типы, которые распределены вблизи регенеративных волокон. Это пик от двух до четырех дней и остается повышенным в течение нескольких дней во время восстановления мышечной ткани. [ 58 ] Первая субпопуляция не имеет прямой пользы для восстановления мышц, в то время как вторая нефагоцитарная группа делает.
Считается, что макрофаги выделяют растворимые вещества, которые влияют на пролиферацию, дифференцировку, рост, восстановление и регенерацию мышц, но в настоящее время фактор, который производится для опосредования этих эффектов, неизвестен. [ 60 ] Известно, что участие макрофагов в стимулирование восстановления тканей не является специфичным для мышц; Они накапливаются в многочисленных тканях во время фазы процесса заживления после травмы. [ 61 ]
Роль в заживлении ран
[ редактировать ]Макрофаги необходимы для заживления ран . [ 62 ] Они заменяют полиморфноядерные нейтрофилы в качестве преобладающих клеток в ране ко второму дню после повреждения. [ 63 ] Притягиваемые к месту раны по факторам роста, высвобождаемым тромбоцитами и другими клетками, моноциты из кровотока попадают в область через стенки кровеносных сосудов. [ 64 ] Количество моноцитов в ране пика от полутора с половиной дней после травмы. Как только они находятся в месте раны, моноциты разразились в макрофаги. Селезенка . содержит половину моноцитов тела в резерве, готовых к развертыванию в поврежденную ткань [ 65 ] [ 66 ]
Основная роль макрофага заключается в фагоцитировании бактерий и поврежденной ткани, [ 62 ] И они также обезведите поврежденную ткань, выпустив протеазы. [ 67 ] Макрофаги также секретируют ряд таких факторов, как факторы роста и другие цитокины, особенно в третий и четвертый пост-ваульный дни. Эти факторы привлекают клетки, участвующие в стадии пролиферации заживления в область. [ 68 ] Макрофаги могут также сдерживать фазу сокращения. [ 69 ] Макрофаги стимулируются низким содержанием кислорода в их окружении, чтобы вызвать факторы, которые вызывают и ускоряют ангиогенез [ 70 ] И они также стимулируют клетки, которые реэпителинизируют рану, создают грануляционную ткань и устанавливают новый внеклеточный матрикс . [ 71 ] [ Лучший источник необходим ] Секстрируя эти факторы, макрофаги способствуют продвижению процесса заживления ран в следующий этап.
Роль в регенерации конечностей
[ редактировать ]Ученые выяснили, что, как и питание материального мусора, макрофаги участвуют в типичной регенерации конечностей в саламандре. [ 72 ] [ 73 ] Они обнаружили, что удаление макрофагов из саламандры привело к неудаче регенерации конечностей и реакции рубцов. [ 72 ] [ 73 ]
Роль в железном гомеостазе
[ редактировать ]Как описано выше, макрофаги играют ключевую роль в удалении умирающих или мертвых клеток и клеточного мусора. Эритроциты имеют в среднем срок службы в среднем 120 дней, и поэтому они постоянно разрушаются макрофагами в селезенке и печени. Макрофаги также будут поглощать макромолекулы и, таким образом, играть ключевую роль в фармакокинетике парентеральных утюгов . [ Цитация необходима ]
Железо, выделяемое из гемоглобина, либо хранится внутри, либо выпускается в циркуляцию через ферропортин . В тех случаях, когда повышаются системные уровни железа или где присутствует воспаление, повышающие уровни гепсидина действуют на макрофагах ферропортин -каналах, что приводит к тому, что железо оставалось в макрофагах. [ Цитация необходима ]
Роль в сохранении пигмента
[ редактировать ]
Меланофаги представляют собой подмножество макрофагов с тканевым резидентом, способным поглощать пигмент, родом из организма или экзогенного (например, татуировки ), из внеклеточного пространства. В отличие от дендритных меланоцитов , которые синтезируют меланосомы и содержат различные этапы их развития, меланофаги накапливают только фагоцитозированный меланин в лизосомоподобных фагосомах. [ 74 ] [ 75 ] Это происходит неоднократно, когда пигмент из мертвых кожных макрофагов фагоцитируется их преемниками, сохраняя татуировку в том же месте. [ 76 ]
Роль в гомеостазе ткани
[ редактировать ]Каждая ткань содержит свою собственную специализированную популяцию резидентных макрофагов, которые развлекают взаимные взаимосвязи со стромой и функциональной тканью. [ 77 ] [ 78 ] Эти резидентные макрофаги являются сидячими (не-мигрирующими), обеспечивают важные факторы роста для поддержки физиологической функции ткани (например, макрофаги-нейрональные перекрестные помехи в кишечниках), [ 79 ] и может активно защищать ткань от воспалительного повреждения. [ 80 ]
Нерв ассоциированные макрофаги
[ редактировать ]Нерво, ассоциированные с макрофагами или NAM, являются теми макрофагами тканевых резидентов, которые связаны с нервами. Известно, что некоторые из них имеют удлиненную морфологию до 200 мкм [ 81 ]
Клиническое значение
[ редактировать ]Из -за их роли в фагоцитозе макрофаги участвуют во многих заболеваниях иммунной системы. Например, они участвуют в формировании гранулем , воспалительных поражениях, которые могут быть вызваны большим количеством заболеваний. Например, были описаны некоторые расстройства, в основном редкие, неэффективного фагоцитоза и функции макрофагов. [ 82 ]
Как хозяин внутриклеточных патогенов
[ редактировать ]В роли фагоцитарных иммунных клеток макрофаги ответственны за то, чтобы их уничтожить патогены. Некоторые патогенные микроорганизмы подрывают этот процесс и вместо этого живут внутри макрофагов. Это обеспечивает среду, в которой патоген скрыт от иммунной системы и позволяет его воспроизвести. [ Цитация необходима ]
Заболевания с этим типом поведения включают туберкулез (вызванный микобактерием туберкулеза ) и лейшманиозом (вызванный видами лейшмании ). [ Цитация необходима ]
Чтобы свести к минимуму возможность стать хозяином внутриклеточных бактерий, макрофаги развивали защитные механизмы, такие как индукция оксида азота и реактивные кислородные промежуточные продукты, [ 83 ] которые токсичны для микробов. Макрофаги также развили способность ограничивать питательный поставщик микроба и вызывать аутофагию . [ 84 ]
Туберкулез
[ редактировать ]После охвата макрофагом, возбудителя туберкулеза, Mycobacterium tuberculosis , [ 85 ] избегает сотовой защиты и использует ячейку для воспроизведения. Последние данные свидетельствуют о том, что в ответ на легочную инфекцию микобактерий туберкулеза периферические макрофаги превращаются в фенотип M1. Фенотип макрофага M1 характеризуется повышенной секрецией провоспалительных цитокинов (IL-1β, TNF-α и IL-6) и повышенной гликолитической активностью, необходимой для клиренса инфекции. [ 1 ]
Лейшманиоз
[ редактировать ]После фагоцитоза макрофагом паразит лейшмании оказывается в фагоцитарной вакуоле. При нормальных обстоятельствах эта фагоцитарная вакуоль будет развиваться в лизосому, и ее содержание будет переваривано. Лейшмания изменяет этот процесс и избегает разрушения; Вместо этого они делают дом в вакуоле. [ Цитация необходима ]
Чикунгунья
[ редактировать ]Инфекция макрофагов в суставах связана с локальным воспалением во время и после острой фазы чикунгунья (вызванного вирусом чикв или чикунгунья). [ 86 ]
Другие
[ редактировать ]Аденовирус (наиболее распространенная причина розового глаза) может оставаться скрытым в макрофагах -хозяине, причем продолжающийся вирусный выпадок через 6–18 месяцев после первоначальной инфекции. [ Цитация необходима ]
Brucella spp. может оставаться скрытым в макрофаге посредством ингибирования фагосомы - лизосомы слияния ; вызывает бруцеллез (нерадосленная лихорадка). [ Цитация необходима ]
Legionella pneumophila , возбудителя болезни Легионеров , также устанавливает резиденцию в макрофагах. [ Цитация необходима ]
Сердечное заболевание
[ редактировать ]Макрофаги являются преобладающими клетками, участвующими в создании прогрессирующих поражений бляшек атеросклероза . [ 87 ]
Фокальный рекрутинг макрофагов происходит после появления острого инфаркта миокарда . Эти макрофаги функционируют для удаления мусора, апоптотических клеток и для подготовки к регенерации тканей . [ 88 ] Макрофаги защищают от индуцированной ишемией желудочковой тахикардии у мышей с гипокалиемией. [ 89 ]
ВИЧ -инфекция
[ редактировать ]Макрофаги также играют роль в инфекции вируса иммунодефицита человека (ВИЧ). Как и Т -клетки , макрофаги могут быть инфицированы ВИЧ и даже стать резервуаром постоянной репликации вируса по всему организму. ВИЧ может войти в макрофаг посредством связывания GP120 с CD4 и вторым мембранным рецептором, CCR5 (рецептор хемокинов). Как циркулирующие моноциты, так и макрофаги служат резервуаром для вируса. [ 90 ] Макрофаги лучше способны сопротивляться инфекции ВИЧ-1, чем CD4+ T-клетки, хотя восприимчивость к ВИЧ-инфекции отличается среди подтипов макрофагов. [ 91 ]
Рак
[ редактировать ]Макрофаги могут способствовать росту и прогрессированию опухоли, способствуя пролиферации и инвазии опухолевых клеток, содействуя ангиогенезу опухоли и подавляя противоопухолевые иммунные клетки. [ 92 ] [ 93 ] Воспалительные соединения, такие как фактор некроза опухоли (TNF) -альфа, высвобождаемая макрофагами, активируют ядерный фактор-переключатель генов B. Затем NF-κB попадает в ядро опухолевой клетки и поворачивает продукцию белков, которые останавливают апоптоз и способствуют пролиферации клеток и воспалению. [ 94 ] Кроме того, макрофаги служат источником для многих проангиогенных факторов, включая эндотелиальный фактор сосудов (VEGF), фактор некроза опухоли (TNF-альфа), макрофаги-стимулирующий фактор (M-CSF/CSF1) и IL-1 и IL- -6 , [ 95 ] способствуя росту опухоли.
Было показано, что макрофаги проникают в ряд опухолей. Их число коррелирует с плохим прогнозом при определенных видах рака, включая рак молочной железы, шейки матки, мочевого пузыря, мозга и простаты. [ 96 ] [ 97 ] Некоторые опухоли также могут вызывать факторы, в том числе M-CSF/CSF1, MCP-1/CCL2 и ангиотензин II , которые запускают амплификацию и мобилизацию макрофагов в опухолях. [ 98 ] [ 99 ] [ 100 ] Кроме того, субкапсулярные синусовые макрофаги в лимфатических узлах с опухолью могут подавлять прогрессирование рака, содержащая распространение материалов, полученных из опухоли. [ 101 ]
Терапия рака
[ редактировать ]Экспериментальные исследования показывают, что макрофаги могут влиять на все терапевтические методы, включая хирургию , химиотерапию , лучевую терапию , иммунотерапию и целевую терапию . [ 93 ] [ 102 ] [ 103 ] Макрофаги могут влиять на результаты лечения как положительно, так и отрицательно. Макрофаги могут быть защитными по -разному: они могут удалять мертвые опухолевые клетки (в процессе, называемом фагоцитозом ) после обработки, которые убивают эти клетки; Они могут служить в качестве лекарственных складов для некоторых противоопухолевых препаратов; [ 104 ] Они также могут быть активированы некоторыми методами лечения, чтобы способствовать противоопухолевому иммунитету. [ 105 ] Макрофаги также могут быть вредными по нескольким способам: например, они могут подавлять различные химиотерапии, [ 106 ] [ 107 ] Радиотерапия [ 108 ] [ 109 ] и иммунотерапия. [ 110 ] [ 111 ] Поскольку макрофаги могут регулировать прогрессирование опухоли, терапевтические стратегии для уменьшения количества этих клеток или манипулировать их фенотипами в настоящее время тестируются у пациентов с раком. [ 112 ] [ 113 ] Тем не менее, макрофаги также участвуют в цитотоксичности, опосредованной антителами (ADCC), и этот механизм был важен для определенных антител к иммунотерапии рака. [ 114 ]
Ожирение
[ редактировать ]Было отмечено, что увеличение числа провоспалительных макрофагов в жировой ткани с ожирением способствует осложнениям ожирения, включая резистентность к инсулину и диабет 2 типа. [ 115 ]
Поэтому модуляция воспалительного состояния макрофагов жировой ткани считалась возможной терапевтической мишенью для лечения заболеваний, связанных с ожирением. [ 116 ] Хотя макрофаги жировой ткани подвержены противовоспалительному гомеостатическому контролю с помощью симпатической иннервации, эксперименты с использованием мышей, нокаутированных геном ADRB2, указывают на то, что этот эффект косвенно проявляется посредством модуляции функции адипоцитов, а не посредством прямой активации адренергического адренергического бета-2 , предполагая, что адренергическая Стимуляция макрофагов может быть недостаточной для воздействия на воспаление или функцию жировой ткани при ожирении. [ 117 ]
В рамках жировой ( жировой ) ткани CCR2 с дефицитом мышей наблюдается увеличенное количество эозинофилов , большую альтернативную активацию макрофагов и склонность к экспрессии цитокинов 2 типа . Кроме того, этот эффект был преувеличен, когда мыши стали ожирением от диеты с высоким содержанием жира. [ 118 ] Это частично вызвано переключением фенотипа макрофагов, индуцированным некрозом жировых клеток ( адипоцитов ). У индивидуума с ожирением некоторые адипоциты ломаются и подвергаются некротической смерти, что заставляет жилые макрофаги M2 переключаться на фенотип M1. Это одна из причин низкоклассного системного хронического воспалительного состояния, связанного с ожирением. [ 119 ] [ 120 ]
Кишечные макрофаги
[ редактировать ]Хотя кишечные макрофаги очень похожи по структуре с тканевыми макрофагами, разработали специфические характеристики и функции, учитывая их природную среду, которая находится в пищеварительном тракте. Макрофаги и кишечные макрофаги обладают высокой пластичностью, вызывая изменение их фенотипа в их среде. [ 121 ] Как и макрофаги, кишечные макрофаги являются дифференцированными моноцитами, хотя кишечные макрофаги должны сосуществовать с микробиомом в кишечнике. Это задача, учитывая, что бактерии, обнаруженные в кишечнике, не признаются «я» и могут быть потенциальными мишенями для фагоцитоза макрофагом. [ 122 ]
Чтобы предотвратить разрушение кишечных бактерий, кишечные макрофаги развили ключевые различия по сравнению с другими макрофагами. Прежде всего, кишечные макрофаги не вызывают воспалительных реакций. Принимая во внимание, что макрофаги ткани выделяют различные воспалительные цитокины, такие как IL-1, IL-6 и TNF-α, кишечные макрофаги не продуцируют и не секретируют воспалительные цитокины. Это изменение напрямую вызвано средой кишечника макрофагов. Окружающие эпителиальные клетки кишечника выделяют TGF-β , что вызывает изменение от провоспалительного макрофага к невоспалительным макрофагам. [ 122 ]
Несмотря на то, что воспалительный ответ понижается в кишечных макрофагах, фагоцитоз все еще осуществляется. В эффективности фагоцитоза нет падения, поскольку кишечные макрофаги способны эффективно фагоцитизировать бактерии, S. typhimurium и E. coli , но кишечные макрофаги все еще не высвобождают цитокины, даже после фагоцитоза. Кроме того, кишечные макрофаги не экспрессируют рецепторы липополисахарида (LPS), IgA или IgG. [ 123 ] Отсутствие рецепторов LPS важно для кишечника, так как кишечные макрофаги не обнаруживают, связанные с микробоем молекулярные паттерны (MAMP/PAMP) кишечного микробиома. Они также не экспрессируют рецепторы фактора роста IL-2 и IL-3. [ 122 ]
Роль в болезнях
[ редактировать ]Было показано, что кишечные макрофаги играют роль в воспалительном заболевании кишечника (IBD), таких как болезнь Крона (CD) и язвенного колита (UC). В здоровом кишечнике кишечные макрофаги ограничивают воспалительный ответ в кишечнике, но в состоянии болезни, кишечные макрофаги и разнообразие изменяются. Это приводит к воспалению кишечника и симптомов заболевания ВЗК. Кишечные макрофаги имеют решающее значение для поддержания гомеостаза кишечника . Наличие воспаления или патогена изменяет этот гомеостаз и одновременно изменяет кишечные макрофаги. [ 124 ] Не было определенного механизма изменения кишечных макрофагов путем набора новых моноцитов или изменений в уже присутствующих кишечных макрофагах. [ 123 ]
Кроме того, новое исследование показывает, что макрофаги ограничивают доступ железа к бактериям путем высвобождения внеклеточных везикул, улучшая результаты сепсиса. [ 125 ]
СМИ
[ редактировать ]-
Активный макрофаг J774, как видно, занимает четыре конидию кооперативным образом. Клетки J774 обрабатывали 5-н/мл интерфероном-γ за одну ночь перед съемкой с конидиями. Наблюдения проводились каждые 30 -е годы в течение 2,5 -часового периода.
-
два высоко активных альвеолярных макрофага Можно увидеть, как проглатывают конидии . Промежуток времени составляет 30 с за кадр в течение 2,5 часов.
История
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Март 2018 г. ) |
Макрофаги были впервые обнаружены в конце 19 -го века Эли Метчникофф . [ 126 ] Эли Метчникофф произвела революцию в отрасли макрофагов, объединив философские идеи и эволюционное изучение жизни. [ 127 ] Позже Ван Форт в 1960 -х годах предложил идею о том, что циркулирующие моноциты крови у взрослых допускают происхождение всех тканевых макрофагов. [ 128 ] В последние годы публикация в отношении макрофагов заставила людей полагать, что множественные резидентные тканевые макрофаги не зависят от моноцитов крови, поскольку они образуются на эмбриональной стадии развития. [ 129 ] В течение 21 -го века все идеи, касающиеся происхождения макрофагов (присутствующих в тканях), составлены вместе, чтобы предположить, что физиологически сложные организмы образуют макрофаги независимо от механизмов, которые не должны зависеть от моноцитов крови. [ 130 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Махла Р.С., Кумар А., Тутин Х.Дж., Кришнаджи С.Т., Сатьямурти Б., Нурсадеги М. и др. (Январь 2021 г.). «NIX-опосредованная митофагия регулирует метаболическое перепрограммирование в фагоцитарных клетках во время микобактериальной инфекции». Туберкулез . 126 (январь): 102046. DOI : 10.1016/j.tube.2020.102046 . PMID 33421909 . S2CID 231437641 .
- ^ «Партнерство по регенеративной медицине в образовании» . Архивировано с оригинала 25 апреля 2015 года . Получено 7 мая 2015 года .
- ^ Nahrendorf M, Hoyer FF, Meerwaldt AE, Van Leent MM, Senders ML, Calcagno C, et al. (Октябрь 2020 г.). "Макрофаги с сердечно -сосудистыми и легкими с помощью датчика по позитронному эмиссионному томографии 64 Cu-Macrin у мышей, кроликов и свиней » . Циркуляция: сердечно-сосудистая визуализация . 13 (10): E010586. DOI : /Circimation.120.010586 . PMC 7583675. 10.1161 PMID 33076700 .
- ^ Овчинников Д.А. (сентябрь 2008 г.). «Макрофаги в эмбрионе и за его пределами: гораздо больше, чем просто гигантские фагоциты» . Бытие . 46 (9): 447–462. doi : 10.1002/dvg.20417 . PMID 18781633 . S2CID 38894501 .
Макрофаги присутствуют практически во всех тканях, начиная с эмбрионального развития и, в дополнение к их роли в защите хозяина и в области апоптотических клеток, все чаще признаются за их трофическую функцию и роль в регенерации.
- ^ CD Mills (2012). «Макрофаги M1 и M2: оракулы здоровья и болезней». Критические обзоры иммунологии . 32 (6): 463–488. doi : 10.1615/critrevimmunol.v32.i6.10 . PMID 23428224 .
- ^ Мерфи К., Уивер С. (2006). Иммунобиология Джейнвея . Гарлендская наука, Нью -Йорк. с. 464, 904. ISBN 978-0-8153-4551-0 .
- ^ Ransohoff RM (июль 2016 г.). «Поляризующий вопрос: существует ли микроглия M1 и M2?». Nature Neuroscience . 19 (8): 987–991. doi : 10.1038/nn.4338 . PMID 27459405 . S2CID 27541569 .
- ^ Krombach F, Münzing S, Allmeling AM, Gerlach JT, Behr J, Dörger M (сентябрь 1997 г.). «Размер клеток альвеолярных макрофагов: межвидовое сравнение» . Перспективы здоровья окружающей среды . 105 (Suppl 5): 1261–1263. doi : 10.2307/3433544 . JSTOR 3433544 . PMC 1470168 . PMID 9400735 .
- ^ Khazen W, M'bika JP, Tomkiewicz C, Benelli C, Chany C, Achour A, et al. (Октябрь 2005 г.). «Экспрессия макрофагов-селективных маркеров в адипоцитах человека и грызунов» . Письма Febs . 579 (25): 5631–5634. doi : 10.1016/j.febslet.2005.09.032 . PMID 16213494 . S2CID 6066984 .
- ^ Zalkind S (2001). Илья Мечников: Его жизнь и работа . Гонолулу, Гавайи: университетская пресса Тихого океана. с. 78, 210. ISBN 978-0-89875-622-7 .
- ^ Shapour-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Marani F, et al. (Сентябрь 2018 г.). «Макрофагская пластичность, поляризация и функции в отношении здоровья и заболеваний » Журнал клеточной физиологии 233 (9): 6425–6 Doi : 10.1002/ jcp.2 29319160PMID 3621509S2CID
- ^ Лото CJ. Принципы физиологии почек, 5 -е издание . Спрингер. п. 37
- ^ Ширази С., Равиндран С., Купер Л.Ф. (декабрь 2022 г.). «Опосредованная топографией иммуномодуляция в остеоинтеграции; союзник или враг» . Биоматериалы . 291 : 121903. DOI : 10.1016/j.biomaterial.2022.121903 . PMC 10148651 . PMID 36410109 .
- ^ Сире С., Ван Медлен М., Бавайс Дж., Чжон Х.В., Редди Самавар С.К., Капупара К. и др. (2022). «Расшифровка гетерогенности периваскулярных макрофагов Lyve1+ в мозге мыши» . Природная связь . 13 (1): 7366. Bibcode : 2022natco..13.7366S . doi : 10.1038/s41467-022-35166-9 . PMC 9712536 . PMID 36450771 .
- ^ Nes WD, Lukyanenko Yo, Jia ZH, Quideau S, Howald WN, Pratum TK, et al. (Март 2000 г.). «Идентификация липофильного фактора, продуцируемого макрофагами, которые стимулируют стероидогенез» . Эндокринология . 141 (3): 953–958. doi : 10.1210/endo.141.3.7350 . PMID 10698170 .
- ^ Hulsmans M, Clauss S, Xiao L, Aguirre AD, King KR, Hanley A, et al. (Апрель 2017). «Макрофаги облегчают электрическую проводимость в сердце» . Клетка . 169 (3): 510–522.E20. doi : 10.1016/j.cell.2017.03.050 . PMC 5474950 . PMID 28431249 .
- ^ Jump up to: а беременный в дюймовый и Моссер Д.М., Эдвардс Дж.П. (декабрь 2008 г.). «Изучение полного спектра активации макрофагов» . Природные обзоры. Иммунология . 8 (12): 958–969. doi : 10.1038/nri2448 . PMC 2724991 . PMID 19029990 .
- ^ Perdiguero Eg, Geissmann F (январь 2016 г.). «Разработка и обслуживание резидентных макрофагов » Природа иммунология 17 (1): 2–8 Doi : 10.1038/ ni.3341 PMC 4950995 26681456PMID
- ^ Джинхоу Ф., Гильямс М (март 2016 г.). «Онтогенство и гомеостаз с тканями-резидентами» . Иммунитет . 44 (3): 439–449. doi : 10.1016/j.immuni.2016.02.024 . PMID 26982352 .
- ^ Питтет М.Дж., Нахрендорф М., Свирски Ф.К. (июнь 2014 г.). «Путешествие от стволовых клеток к макрофагам» . Анналы нью -йоркской академии наук . 1319 (1): 1–18. Bibcode : 2014nyasa1319 .... 1p . doi : 10.1111/nyas.12393 . PMC 4074243 . PMID 24673186 .
- ^ Wang M, Yang Y, Cansever D, Wang Y, Kantores C, Messiaen S, et al. (Январь 2021 г.). «Две популяции самостоятельных моноцит-независимых макрофагов существуют у взрослых эпидидимис и яичка» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (1): E2013686117. Bibcode : 2021pnas..11813686W . doi : 10.1073/pnas.2013686117 . PMC 7817195 . PMID 33372158 .
- ^ Jump up to: а беременный в Eming SA, Krieg T, Davidson JM (март 2007 г.). «Воспаление при восстановлении раны: молекулярные и клеточные механизмы» . Журнал следственной дерматологии . 127 (3): 514–25. doi : 10.1038/sj.jid.5700701 . PMID 17299434 .
= Моноциты/макрофаги. Если стимулы для рекрутирования нейтрофилов не сохраняются в месте раны, инфильтрация нейтрофилов не прекращается через несколько дней, а затраченные нейтрофилы сами фагоцитозированы макрофагами, которые присутствуют на стороне раны в течение 2 дней после травмы.
- ^ Monteith AJ, Miller JM, Maxwell CN, Chazin WJ, Skaar EP (сентябрь 2021 г.). «Нейтрофильные внеклеточные ловушки усиливают убийство макрофагов бактериальных патогенов» . Наука достижения . 7 (37): EABJ2101. Bibcode : 2021scia .... 7.2101M . doi : 10.1126/sciadv.abj2101 . PMC 8442908 . PMID 34516771 .
- ^ Верма Н., Сараф С. (15 ноября 2017 г.). «Роль макрофагов: обзор» . Журнал доставки лекарств и терапии . 7 (6): 91–103. doi : 10.22270/jddt.v7i6.1521 . ISSN 2250-1177 .
- ^ Yashroy RC (2000). «Похищение макрофагов Salmonella (310R) через секрецию« типы III », такая как экзоцитотическая передача сигналов: механизм заражения куриной подвздошной кишки» . Индийский журнал птицеводства . 35 (3): 276–281.
- ^ Jump up to: а беременный в дюймовый и фон глин Arango Duque G, Descoteaux A (7 октября 2014 г.). «Макрофажные цитокины: участие в иммунитете и инфекционных заболеваниях» . Границы в иммунологии . 5 : 491. DOI : 10.3389/fimmu.2014.00491 . PMC 4188125 . PMID 25339958 .
- ^ Jump up to: а беременный в дюймовый и Punt J, Stranford S, Jones P, Owen J (25 мая 2018 г.). Kuby Immunology (8 -е изд.). Нью -Йорк, Нью -Йорк: WH Freeman. ISBN 978-1-4641-8978-4 .
- ^ Jump up to: а беременный в Fu yl, Harrison Re (29 апреля 2021 года). «Микробные фагоцитарные рецепторы и их потенциальное участие в индукции цитокинов в макрофагах» . Границы в иммунологии . 12 : 662063. DOI : 10.3389/fimmu.2021.662063 . PMC 8117099 . PMID 33995386 .
- ^ Jump up to: а беременный Hirayama D, Iida T, Nakase H (декабрь 2017 г.). «Фагоцитарная функция макрофагов, способствующего врожденному иммунитету и гомеостазу ткани» . Международный журнал молекулярных наук . 19 (1): 92. doi : 10.3390/ijms19010092 . PMC 5796042 . PMID 29286292 .
- ^ Uribe-Querol E, Rosales C (2 июня 2020 года). «Фагоцитоз: наше нынешнее понимание универсального биологического процесса» . Границы в иммунологии . 11 : 1066. DOI : 10.3389/fimmu.2020.01066 . PMC 7280488 . PMID 32582172 .
- ^ Закон SK (1 января 1988 г.). «Рецепторы C3 на макрофагах». Журнал сотовой науки. Добавка . 9 (Приложение_9): 67–97. doi : 10.1242/jcs.1988.supplement_9.4 . PMID 2978518 . S2CID 29387085 .
- ^ Forman HJ, Torres M (декабрь 2002 г.). «Реактивные формы кислорода и клеточная передача сигналов: дыхательный взрыв в передаче сигналов макрофагов». Американский журнал респираторной медицины и медицины интенсивной терапии . 166 (12 Pt 2): S4 - S8. doi : 10.1164/rccm.2206007 . PMID 12471082 . S2CID 22246117 .
- ^ Лю Т., Чжан Л., Джу Д., Сан (14 июля 2017 г.). «Передача сигналов NF-κB в воспалении» . Сигнальная трансдукция и целевая терапия . 2 (1): 17023–. doi : 10.1038/sigtrans.2017.23 . PMC 5661633 . PMID 29158945 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий Мерфи К., Уивер С, Берг Л (2022). Иммунобиология Джейнвея (10 -е изд.). Нью -Йорк, Нью -Йорк: WW Norton & Company. ISBN 978-0-393-88487-6 .
- ^ Eming SA, Krieg T, Davidson JM (март 2007 г.). «Воспаление при восстановлении раны: молекулярные и клеточные механизмы» . Журнал следственной дерматологии . 127 (3): 514–525. doi : 10.1038/sj.jid.5700701 . PMID 17299434 .
- ^ Mezouar S, Mege JL (июль 2020 г.). «Изменение парадигмы IFN-γ на границе между врожденным и адаптивным иммунитетом: IFN-γ с макрофагом». Журнал биологии лейкоцитов . 108 (1): 419–426. doi : 10.1002/jlb.4mir0420-619rr . PMID 32531848 . S2CID 219622032 .
- ^ Kress H, Stelzer EH, Holzer D, Buss F, Griffiths G, Rohrbach A (июль 2007 г.). «Филоподия действует как фагоцитарные щупальца и тянут с дискретными шагами и зависимой от нагрузки скорости» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (28): 11633–11638. Bibcode : 2007pnas..10411633K . doi : 10.1073/pnas.0702449104 . PMC 1913848 . PMID 17620618 .
- ^ Jump up to: а беременный в Guerriero JL (2019). «Макрофаги: их невыразительная история в активации и функции Т -клеток». Международный обзор клеточной и молекулярной биологии . 342 . Elsevier: 73–93. doi : 10.1016/bsicmb.2018.07.001 . ISBN 978-0-12-815381-9 Полем PMID 30635094 .
- ^ Jump up to: а беременный в Итано А.А., Дженкинс М.К. (август 2003 г.). «Презентация антигена на наивные CD4 T -клетки в лимфатическом узле». Природа иммунология . 4 (8): 733–739. doi : 10.1038/ni957 . PMID 12888794 . S2CID 10305140 .
- ^ Jump up to: а беременный в Мерфи К., Уивер С. (2016). Иммунобиология Джейнвея (9 -е изд.). Нью -Йорк, Нью -Йорк: Гарлендская наука. С. 363–364. ISBN 978-0-8153-4505-3 .
- ^ Jump up to: а беременный Grey EE, Cyster JG (2012). «Макрофаги лимфатического узла» . Журнал врожденного иммунитета . 4 (5–6): 424–436. doi : 10.1159/000337007 . PMC 3574571 . PMID 22488251 .
- ^ Аббас А.К. (сентябрь 2020 г.). «Удивительная история IL-2» . Американский журнал патологии . 190 (9): 1776–1781. doi : 10.1016/j.ajpath.2020.05.007 . PMID 32828360 . S2CID 221280663 .
- ^ Annunziato F, Romagnani C, Romagnani S (март 2015 г.). «3 основных типа врожденного и адаптивного клеточного эффекторного иммунитета» . Журнал аллергии и клинической иммунологии . 135 (3): 626–635. doi : 10.1016/j.jaci.2014.11.001 . PMID 25528359 .
- ^ Jump up to: а беременный в Cai H, Zhang Y, Wang J, Gu J (23 июня 2021 г.). «Дефекты в перепрограммировании макрофагов при терапии рака: негативное влияние PD-L1/PD-1» . Границы в иммунологии . 12 : 690869. DOI : 10.3389/fimmu.2021.690869 . PMC 8260839 . PMID 34248982 .
- ^ Jump up to: а беременный Hilhorst M, Shirai T, Berry G, Goronzy JJ, Weyand CM (2014). «Т-клеточные взаимодействия и образование гранулемы при васкулите» . Границы в иммунологии . 5 : 432. DOI : 10.3389/fimmu.2014.00432 . PMC 4162471 . PMID 25309534 .
- ^ Jump up to: а беременный в дюймовый Rolot M, Dewals BG (2 июля 2018 г.). «Активация и функционирование макрофагов во время гельминтовой инфекции: последние достижения лабораторной мыши» . Журнал иммунологических исследований . 2018 : 2790627. DOI : 10.1155/2018/2790627 . PMC 6051086 . PMID 30057915 .
- ^ Гордон С. (январь 2003 г.). «Альтернативная активация макрофагов». Природные обзоры. Иммунология . 3 (1): 23–35. doi : 10.1038/nri978 . PMID 12511873 . S2CID 23185583 .
- ^ Peters VA, Joesting JJ, Freund GG (август 2013 г.). «Рецептор 2 IL-1 (IL-1R2) и его роль в иммунной регуляции» . Мозг, поведение и иммунитет . 32 : 1–8. doi : 10.1016/j.bbi.2012.11.006 . PMC 3610842 . PMID 23195532 .
- ^ MuntJewerff EM, Meesters Ld, Van Den Bogaart G (8 июля 2020 года). «Антигеном перекрестная презентация макрофагами» . Границы в иммунологии . 11 : 1276. DOI : 10.3389/fimmu.2020.01276 . PMC 7360722 . PMID 32733446 .
- ^ Xu W, Banchereau J (январь 2014 г.). «Антиген, представляющие клетки, инструктируют дифференцировку плазматических клеток» . Границы в иммунологии . 4 : 504. DOI : 10.3389/fimmu.2013.00504 . PMC 3880943 . PMID 24432021 .
- ^ Котран Р.С., Кумар В., Коллинз Т. (1999). Роббинс патологическая основа болезни . Филадельфия: WB Saunders Company. ISBN 978-0-7216-7335-6 .
- ^ «История лимфоцитов» . Новый ученый (1605) . Получено 13 сентября 2007 года .
- ^ Jump up to: а беременный в дюймовый Hesketh M, Sahin KB, West Ze, Murray RZ (июль 2017 г.). «Фенотипы макрофагов регулируют образование рубцов и хроническое заживление ран» . Международный журнал молекулярных наук . 18 (7): 1545. DOI : 10.3390/IJMS18071545 . PMC 5536033 . PMID 28714933 .
- ^ Galdiero MR, Garlanda C, Jaillon S, Marone G, Mantovani A (июль 2013 г.). «Связанные с опухолями макрофаги и нейтрофилы при прогрессировании опухоли». Журнал клеточной физиологии . 228 (7): 1404–1412. doi : 10.1002/jcp.24260 . PMID 23065796 . S2CID 41189572 .
- ^ Hotamisligil GS (апрель 2010 г.). «Стресс эндоплазматического ретикулума и атеросклероз» . Природная медицина . 16 (4): 396–399. doi : 10.1038/nm0410-396 . PMC 2897068 . PMID 20376052 .
- ^ О, Дж, Рик А.Е., Вэн С., Петти М., Ким Д., Колонна М. и др. (Апрель 2012 г.). «Стресс эндоплазматического ретикулума контролирует дифференцировку макрофагов M2 и образование пенистых клеток» . Журнал биологической химии . 287 (15): 11629–11641. doi : 10.1074/jbc.m111.338673 . PMC 3320912 . PMID 22356914 .
- ^ Криппендорф Б.Б., Райли Да (январь 1993 г.). «Различающая разгрузка- по сравнению с индуцированными перезагруженными изменениями в мышцах подошвы крысы». Мышцы и нерв . 16 (1): 99–108. doi : 10.1002/mus.880160116 . PMID 8423838 . S2CID 23012375 .
- ^ Jump up to: а беременный в Сент -Пьер BA, Tidball JG (июль 1994 г.). «Дифференциальный ответ субпопуляций макрофагов на перезагрузку мышц подошва после суспензии задних конечностей крысы». Журнал прикладной физиологии . 77 (1): 290–297. doi : 10.1152/jappl.1994.77.1.290 . PMID 7961247 .
- ^ Tidball JG, Berchenko E, Frenette J (апрель 1999 г.). «Инвазия макрофагов не способствует повреждению мышечной мембраны во время воспаления» . Журнал биологии лейкоцитов . 65 (4): 492–498. doi : 10.1002/jlb.65.4.492 . PMID 10204578 . S2CID 23315528 .
- ^ Jump up to: а беременный Schiaffino S, Partridge T (2008). Восстановление скелетных мышц и регенерация . Достижения в области исследований мышц. Тол. 3
- ^ Brechot N, Gomez E, Bignon M, Khallou-Laschet J, Dussiot M, Cazes A, et al. (2008). «Модуляция состояния активации макрофагов защищает ткани от некроза во время критической ишемии конечностей у мышей с дефицитом тромбоспондина-1» . Plos один . 3 (12): E3950. Bibcode : 2008ploso ... 3.3950b . doi : 10.1371/journal.pone.0003950 . PMC 2597179 . PMID 19079608 .
- ^ Jump up to: а беременный Де Ла Торре Дж., Шолар А. (2006). Заживление раны: хронические раны . Emedicine.com. Доступ 20 января 2008 года.
- ^ «Фазы кожного заживления ран» (PDF) . Экспертные обзоры в молекулярной медицине . 5 Издательство Кембриджского университета. 21 марта 2003 года. Архивировано из оригинала (PDF) 17 декабря 2008 года.
- ^ Lorenz HP, Longaker MT (2003). «Раны: биология, патология и управление». (PDF) . В Li M, Norton JA, Bollinger RR, Chang AE, Lowry SF, Mulvihill SJ, Pass Hi, Thompson RW (Eds.). Основная практика хирургии . Нью -Йорк, Нью -Йорк: Спрингер. С. 77–88. ISBN 978-0-387-22744-3 Полем Архивировано из оригинала (PDF) 31 октября 2005 года.
- ^ Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, et al. (Июль 2009 г.). «Идентификация моноцитов селезеночного резервуара и их развертывание на воспалительные участки» . Наука . 325 (5940): 612–616. Bibcode : 2009Sci ... 325..612S . doi : 10.1126/science.1175202 . PMC 2803111 . PMID 19644120 .
- ^ Цзя Т., Памер Э.Г. (июль 2009 г.). «Иммунология. Необходимый, но не неуместный» . Наука . 325 (5940): 549–550. Bibcode : 2009Sci ... 325..549J . doi : 10.1126/science.1178329 . PMC 2917045 . PMID 19644100 .
- ^ Deodhar AK, Rana Re (1997). «Хирургическая физиология заживления ран: обзор» . Журнал аспирантуры . 43 (2): 52–56. PMID 10740722 .
- ^ Розенберг Л., Де Ла Торре Дж. (2006). Заживление ран, факторы роста . Emedicine.com. Доступ 20 января 2008 года.
- ^ Newton PM, Watson JA, Wolowacz RG, Wood EJ (август 2004 г.). «Макрофаги сдерживают сокращение модели заживления ран in vitro». Воспаление . 28 (4): 207–214. doi : 10.1023/b: ifla.0000049045.41784.59 . PMID 15673162 . S2CID 9612298 .
- ^ Greenhalgh DG (сентябрь 1998 г.). «Роль апоптоза в заживлении ран». Международный журнал биохимии и клеточной биологии . 30 (9): 1019–1030. doi : 10.1016/s1357-2725 (98) 00058-2 . PMID 9785465 .
- ^ Stashak TS, Farstvedt E, Othic A (июнь 2004 г.). «Обновление о заправках с ранами: показаниями и лучшим использованием». Клинические методы в практике лошадей . 3 (2): 148–163. doi : 10.1053/j.ctep.2004.08.006 .
- ^ Jump up to: а беременный Souppouris A (23 мая 2013 г.). «Ученые идентифицируют клетку, которая может хранить секрет регенерации конечностей» . Verge.com.
Исследователи определили клетку, которая помогает отрастать конечности в саламандре. Макрофаги - это тип восстановительной ячейки, которая пожирает мертвые клетки и патогены, и запускают другие иммунные клетки для реагирования на патогены.
- ^ Jump up to: а беременный Godwin JW, Pinto AR, Rosenthal NA (июнь 2013 г.). «Макрофаги необходимы для взрослых самолетов саламандры» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (23): 9415–9420. Bibcode : 2013pnas..110.9415G . doi : 10.1073/pnas.1300290110 . PMC 3677454 . PMID 23690624 .
- ^ Мишима Y (октябрь 1967 г.). «Лизосомы при малянинском фагоцитозе и синтезе» . Природа . 216 (5110): 67. Bibcode : 1967nater.216 ... 67M . doi : 10.1038/216067a0 . PMID 6050674 . S2CID 4285140 .
- ^ Мишима Y (январь 1966 г.). «Клеточная и субклеточная дифференциация фагоцитоза меланина и синтеза лизосомальной и меланосомной активностью» . Журнал следственной дерматологии . 46 (1): 70–75. doi : 10.1038/jid.1966.11 . PMID 5905254 .
- ^ Baranska A, Shawket A, Jouve M, Baratin M, Malosse C, Voluzan O, et al. (Апрель 2018). «Раскрывающая динамика макрофагов кожи объясняет как стойкость татуировки, так и напряженное удаление» . Журнал экспериментальной медицины . 215 (4): 1115–1133. doi : 10.1084/jem.20171608 . PMC 5881467 . PMID 29511065 .
- ^ Okabe Y, Medzhitov R (май 2014). «Специфичные для ткани сигналы контролируют обратимую программу локализации и функциональной поляризации макрофагов» . Клетка . 157 (4): 832–844. doi : 10.1016/j.cell.2014.04.016 . PMC 4137874 . PMID 24792964 .
- ^ Gosselin D, Link VM, Romanoski CE, Fonseca GJ, Eichenfield DZ, Spann NJ, et al. (Декабрь 2014). «Окружающая среда приводит к выбору и функции усилителей, контролирующих тканеспецифические идентичности макрофагов» . Клетка . 159 (6): 1327–1340. doi : 10.1016/j.cell.2014.11.023 . PMC 4364385 . PMID 25480297 .
- ^ Muller PA, Koscsó B, Rajani GM, Stevanovic K, Berres ML, Hashimoto D, et al. (Июль 2014). «Перекрестка между макрофагами Muscularis и кишечными нейронами регулирует желудочно -кишечную подвижность» . Клетка . 158 (2): 300–313. doi : 10.1016/j.cell.2014.04.050 . PMC 4149228 . PMID 25036630 .
- ^ Uderhardt S, Martins AJ, Tsang JS, Lämmermann T, Germain RN (апрель 2019 г.). «Резидентские макрофаги выключите ткань, чтобы предотвратить воспалительное повреждение, управляемое нейтрофилами» . Клетка . 177 (3): 541–555.e17. doi : 10.1016/j.cell.2019.02.028 . PMC 6474841 . PMID 30955887 .
- ^ Колтер Дж., Кирдорф К., Хеннеке П (январь 2020 г.). «Происхождение и дифференциация нервных макрофагов» . Журнал иммунологии . 204 (2): 271–279. doi : 10.4049/jimmunol.1901077 . PMID 31907269 . S2CID 210043405 .
- ^ Вольф А.Дж., Андерхилл Д.М. (2014). "Фагоцитоз". Макрофаги: биология и роль в патологии заболеваний . Спрингер Нью -Йорк. С. 91–109. doi : 10.1007/978-1-4939-1311-4_5 . ISBN 978-1-4939-1310-7 .
- ^ Herb M, Schramm M (февраль 2021 г.). «Функции АФК в макрофагах и антимикробном иммунитете» . Антиоксиданты . 10 (2): 313. doi : 10.3390/antiox10020313 . PMC 7923022 . PMID 33669824 .
- ^ Weiss G, Schaible UE (март 2015 г.). «Механизмы защиты макрофагов от внутриклеточных бактерий» . Иммунологические обзоры . 264 (1): 182–203. doi : 10.1111/imr.12266 . PMC 4368383 . PMID 25703560 .
- ^ Райан К.Дж., Рэй К.Г., ред. (2004). Шеррис Медицинская микробиология (4 -е изд.). МакГроу Хилл. ISBN 978-0-8385-8529-0 .
- ^ Dupuis-Maguiraga L, Noret M, Brun S, Le Grand R, Gras G, Roques P (2012). «Болезнь Чикунгунья: связанные с инфекцией маркеры от острой до хронической фазы артралгии, вызванной арбовирусом» . ПЛО не пренебрегали тропическими заболеваниями . 6 (3): E1446. doi : 10.1371/journal.pntd.0001446 . PMC 3313943 . PMID 22479654 .
- ^ Лукас А.Д., Гривз Д.Р. (ноябрь 2001 г.). «Атеросклероз: роль хемокинов и макрофагов». Экспертные обзоры в молекулярной медицине . 3 (25): 1–18. doi : 10.1017/s1462399401003696 . PMID 14585150 . S2CID 8952545 .
- ^ Frantz S, Nahrendorf M (май 2014). «Сердечные макрофаги и их роль в ишемической болезни сердца» . Сердечно -сосудистые исследования . 102 (2): 240–248. doi : 10.1093/cvr/cvu025 . PMC 3989449 . PMID 24501331 .
- ^ Grune J, Lewis AJ, Yamazoe M, Hulsmans M, Rohde D, Xiao L, et al. (Июль 2022 г.). «Нейтрофилы подстрекают и макрофаги предотвращают электрический шторм после инфаркта миокарда» . Природные сердечно -сосудистые исследования . 1 (7): 649–664. doi : 10.1038/s44161-022-00094-w . PMC 9410341 . PMID 36034743 . S2CID 250475623 .
- ^ Бол С.М., Кобос-Хименес В., Кутстра Н., Ван-не Вуут А.Б. (февраль 2011 г.). «Марофейс». Вирусология . 6 (2): 187–208. DOS : 10.2217/RVL.10.93 .
- ^ Koppensteiner H, Brack-Werner R, Schindler M (октябрь 2012 г.). «Макрофаги и их актуальность в инфекции вируса иммунодефицита человека» . Ретровирология . 9 (1): 82. doi : 10.1186/1742-4690-9-82 . PMC 3484033 . PMID 23035819 .
- ^ Qian BZ, Pollard JW (апрель 2010 г.). «Разнообразие макрофагов усиливает прогрессирование опухоли и метастазирование» . Клетка . 141 (1): 39–51. doi : 10.1016/j.cell.2010.03.014 . PMC 4994190 . PMID 20371344 .
- ^ Jump up to: а беременный Engblom C, Pfirschke C, Pittet MJ (июль 2016 г.). «Роль миелоидных клеток в терапии рака». Природные обзоры. Рак . 16 (7): 447–462. doi : 10.1038/nrc.2016.54 . PMID 27339708 . S2CID 21924175 .
- ^ Stix G (июль 2007 г.). «Злокачественное пламя. Понимание хронического воспаления, которое способствует болезням сердца, болезням Альцгеймера и множеству других заболеваний, может быть ключом к разблокировке загадок рака». Scientific American . 297 (1): 60–67. Bibcode : 2007sciam.297a..60s . doi : 10.1038/Scientificamerican0707-60 . PMID 17695843 .
- ^ Лин Эй, Ли Дж. Ф., Гнатовский Л., Денг Й., Чжу Л., Грзесик Д.А. и др. (Декабрь 2006 г.). «Макрофаги регулируют ангиогенный переключатель на мышиной модели рака молочной железы». РАНКА . 66 (23): 11238–11246. doi : 10.1158/0008-5472.can-06-1278 . PMID 17114237 . S2CID 12722658 .
- ^ Бингл Л., Браун Н.Дж., Льюис Се. Роль макрофагов, связанных с опухолем в прогрессировании опухоли: последствия для новых противоопухолевых методов лечения. J Pathol 2002; 196: 254–65.
- ^ De Groot AE (июль 2018 г.). «Модель макрофагов, ассоциированная с опухолью человека, включает в себя пролиферацию макрофагов как механизм поддержания ассоциированных с опухолью популяций макрофагов» . РАНКА . 78 (13 Дополнение): 4060. DOI : 10.1158/1538-7445.AM2018-4060 . S2CID 80769044 .
- ^ Лин Эй, Нгуен А.В., Рассел Р.Г., Поллард Дж.В. (март 2001 г.). «Колони-стимулирующий фактор 1 способствует прогрессированию опухолей молочной железы до злокачественных новообразований» . Журнал экспериментальной медицины . 193 (6): 727–740. doi : 10.1084/jem.193.6.727 . PMC 2193412 . PMID 11257139 .
- ^ Qian BZ, Li J, Zhang H, Kitamura T, Zhang J, Campion LR, et al. (Июнь 2011 г.). «CCL2 рекрутирует воспалительные моноциты, чтобы облегчить метастазирование грудина» . Природа . 475 (7355): 222–225. doi : 10.1038/nature10138 . PMC 3208506 . PMID 21654748 .
- ^ Cortez-Retamozo V, Etzrodt M, Newton A, Ryan R, Pucci F, SIO SW, et al. (Февраль 2013 г.). «Ангиотензин II ведет выработку макрофагов, способствующих опухоле,» . Иммунитет . 38 (2): 296–308. doi : 10.1016/j.immuni.2012.10.015 . PMC 3582771 . PMID 23333075 .
- ^ Pucci F, Garris C, Lai CP, Newton A, Pfirschke C, Engblom C, et al. (Апрель 2016 г.). «Макрофаги SCS подавляют меланому, ограничивая взаимодействия с пузырьками-b-клетками, полученные из опухоли» . Наука . 352 (6282): 242–246. Bibcode : 2016sci ... 352..242p . doi : 10.1126/science.aaf1328 . PMC 4960636 . PMID 26989197 .
- ^ Mantovani A, Allavena P (апрель 2015 г.). «Взаимодействие противоопухолевой терапии с связанными с опухолью макрофагов» . Журнал экспериментальной медицины . 212 (4): 435–445. doi : 10.1084/jem.20150295 . PMC 4387285 . PMID 25753580 .
- ^ De Palma M, Lewis CE (март 2013 г.). «Регуляция макрофагов опухолевых реакций на противоопухолевую терапию» . Раковая клетка . 23 (3): 277–286. doi : 10.1016/j.ccr.2013.02.013 . PMID 23518347 .
- ^ Miller MA, Zheng YR, Gadde S, Pfirschke C, Zope H, Engblom C, et al. (Октябрь 2015). «Макрофаги, ассоциированные с опухолом, действуют как резервуар с медленным высвобождением нанотерапевтического PT (IV) Pro-Srug» . Природная связь . 6 : 8692. Bibcode : 2015natco ... 6.8692m . doi : 10.1038/ncomms9692 . PMC 4711745 . PMID 26503691 .
- ^ Klug F, Prakash H, Huber PE, Seibel T, Bender N, Halama N, et al. (Ноябрь 2013). «Программы макрофагов с низкой дозой дифференцировки макрофагов в фенотип INOS⁺/M1, который организует эффективную иммунотерапию Т-клеток» . Раковая клетка . 24 (5): 589–602. doi : 10.1016/j.ccr.2013.09.014 . PMID 24209604 .
- ^ Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CM, Pryer N, et al. (Ноябрь 2014). «Macrophage IL-10 блокирует CD8+ Т-клеточные реакции на химиотерапию путем подавления экспрессии IL-12 во внутриопухолевых дендритных клетках» . Раковая клетка . 26 (5): 623–637. doi : 10.1016/j.ccell.2014.09.006 . PMC 4254570 . PMID 25446896 .
- ^ Денардо Д.Г., Бреннан Д.Дж., Рексхепай Э., Руффелл Б., Шиао С.Л., Мэдден С.Ф. и др. (Июнь 2011 г.). «Сложность лейкоцитов предсказывает выживаемость рака молочной железы и функционально регулирует реакцию на химиотерапию» . Открытие рака . 1 (1): 54–67. doi : 10.1158/2159-8274.cd-10-0028 . PMC 3203524 . PMID 22039576 .
- ^ Shiao SL, Ruffell B, Denardo DG, Faddegon BA, Park CC, Coussens LM (май 2015). «Th2-поляризованные CD4 (+) Т-клетки и макрофаги ограничивают эффективность лучевой терапии» . Раковая иммунология исследования . 3 (5): 518–525. doi : 10.1158/2326-6066.cir-14-0232 . PMC 4420686 . PMID 25716473 .
- ^ Козин С.В., Камоун В.С., Хуан Ю., Доусон М.Р., Джейн Р.К., Дуда Д.Г. (июль 2010 г.). «Рекрутирование миелоидных, но не эндотелиальных клеток -предшественников облегчает отражение опухоли после локального облучения» . РАНКА . 70 (14): 5679–5685. doi : 10.1158/0008-5472.can-09-4446 . PMC 2918387 . PMID 20631066 .
- ^ Arlauckas SP, Garris CS, Kohler RH, Kitaoka M, Cuccarese MF, Yang KS, et al. (Май 2017). «Визуализация in vivo выявляет, связанный с опухолью, опосредованным макрофагским пути резистентности при терапии против PD-1» . Научная трансляционная медицина . 9 (389): EAAL3604. doi : 10.1126/scitranslmed.aal3604 . PMC 5734617 . PMID 28490665 .
- ^ Zhu Y, Knolhoff BL, Meyer MA, Nywening TM, West BL, Luo J, et al. (Сентябрь 2014). «CSF1/CSF1R-блокада перепрограммирует инфильтрирующие опухоль макрофаги и улучшает реакцию на иммунотерапию контрольной точки Т-клеток в моделях рака поджелудочной железы» . РАНКА . 74 (18): 5057–5069. doi : 10.1158/0008-5472.can-13-3723 . PMC 4182950 . PMID 25082815 .
- ^ Ries CH, Cannarile MA, Hoves S, Benz J, Wartha K, Runza V, et al. (Июнь 2014 г.). «Нацеливание на макрофаги, ассоциированные с опухолью с антителом против CSF-1R, выявляет стратегию терапии рака» . Раковая клетка . 25 (6): 846–859. doi : 10.1016/j.ccr.2014.05.016 . PMID 24898549 .
- ^ Ruffell B, Coussens LM (апрель 2015 г.). «Макрофаги и терапевтическая устойчивость при раке» . Раковая клетка . 27 (4): 462–472. doi : 10.1016/j.ccell.2015.02.015 . PMC 4400235 . PMID 25858805 .
- ^ Sharma N, Vacher J, Allison JP (май 2019). «Лиганд TLR1/2 повышает противоопухолевую эффективность блокады CTLA-4 за счет увеличения внутриопухолевого истощения Treg» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (21): 10453–10462. Bibcode : 2019pnas..11610453S . doi : 10.1073/pnas.1819004116 . PMC 6534983 . PMID 31076558 .
- ^ Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante Aw. Ожирение связано с накоплением макрофагов в жировой ткани " Журнал клинических исследований 2003; 112: 1796–808.
- ^ Гилхерм А., Энрикес Ф., Бедард А.Х., Чешский депутат (апрель 2019 г.). «Молекулярные пути, связывающие жировую иннервацию с действием инсулина при ожирении и сахарном диабете» . Природные обзоры. Эндокринология . 15 (4): 207–225. doi : 10.1038/s41574-019-0165-y . PMC 7073451 . PMID 30733616 .
- ^ Petkevicius K, Bidault G, Wortue S, Newland SA, Dale M, Dugourd A, et al. (Июнь 2021 г.). «Макрофаг бета2-адренергический рецептор не подходит для воспаления и функции жировой ткани» . Молекулярный метаболизм . 48 : 101220. DOI : 10.1016/j.molmet.2021.101220 . PMC 8086137 . PMID 33774223 .
- ^ Bolus WR, Gutierrez DA, Kennedy AJ, Anderson-Baucum EK, Hasty AH (октябрь 2015 г.). «Дефицит CCR2 приводит к увеличению эозинофилов, альтернативной активации макрофагов и экспрессии цитокинов типа 2 в жировой ткани» . Журнал биологии лейкоцитов . 98 (4): 467–477. doi : 10.1189/jlb.3hi0115-018r . PMC 4763864 . PMID 25934927 .
- ^ Boutens L, Stienstra R (май 2016 г.). «Макрофаги жировой ткани: уход с пути во время ожирения» . Диабетология . 59 (5): 879–894. doi : 10.1007/s00125-016-3904-9 . PMC 4826424 . PMID 26940592 .
- ^ Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E, et al. (Ноябрь 2005 г.). «Смерть адипоцитов определяет локализацию и функцию макрофагов в жировой ткани у мышей с ожирением и людей» . Журнал липидных исследований . 46 (11): 2347–2355. doi : 10.1194/jlr.m500294-jlr200 . PMID 16150820 .
- ^ Kühl AA, Erben U, Kredel Li, Siegmund B (7 декабря 2015 г.). «Разнообразие кишечных макрофагов при воспалительных заболеваниях кишечника» . Границы в иммунологии . 6 : 613. DOI : 10.3389/fimmu.2015.00613 . PMC 4670857 . PMID 26697009 .
- ^ Jump up to: а беременный в Smythies LE, Sellers M, Clements RH, Mosteller-Barnum M, Meng G, Benjamin WH, et al. (Январь 2005 г.). «Человеческие кишечные макрофаги демонстрируют глубокую воспалительную анергию, несмотря на страстную фагоцитарную и бактериоцидную активность» . Журнал клинических исследований . 115 (1): 66–75. doi : 10.1172/jci19229 . PMC 539188 . PMID 15630445 .
- ^ Jump up to: а беременный Mowat AM, Bain CC (2011). «Макрофаги слизистой оболочки в кишечном гомеостазе и воспалении» . Журнал врожденного иммунитета . 3 (6): 550–564. doi : 10.1159/000329099 . PMC 3224516 . PMID 22025201 .
- ^ Bain CC, Mowat Am (июль 2014 г.). «Макрофаги в кишечном гомеостазе и воспалении» . Иммунологические обзоры . 260 (1): 102–117. doi : 10.1111/imr.12192 . PMC 4141699 . PMID 24942685 .
- ^ Вайс Г (январь 2023 г.). «Макрофажные пузырьки голодают бактерии железа». Природа метаболизм . 5 (1): 10–12. doi : 10.1038/s42255-022-00719-1 . PMID 36658401 . S2CID 256030791 .
- ^ Эпельман С., Лавин К.Дж., Рэндольф Г.Дж. (июль 2014 г.). «Происхождение и функции тканевых макрофагов» . Иммунитет . 41 (1): 21–35. doi : 10.1016/j.immuni.2014.06.013 . PMC 4470379 . PMID 25035951 .
- ^ Mass E, Lachmann N (сентябрь 2021 г.). «От биологии макрофагов до клеточных иммунотерапии на основе макрофагов» . Генная терапия . 28 (9): 473–476. doi : 10.1038/s41434-021-00221-5 . PMC 8455330 . PMID 33542457 .
- ^ Эпельман С., Лавин К.Дж., Рэндольф Г.Дж. (июль 2014 г.). «Происхождение и функции тканевых макрофагов» . Иммунитет . 41 (1): 21–35. doi : 10.1016/j.immuni.2014.06.013 . PMC 4470379 . PMID 25035951 .
- ^ Итальянский П, Бораски Д (август 2015 г.). «Новое понимание тканевых макрофагов: от их происхождения до развития памяти» . Иммунная сеть . 15 (4): 167–176. doi : 10.4110/in.2015.15.4.167 . PMC 4553254 . PMID 26330802 .
- ^ Lazarov T, Juarez-Carreño S, Cox N, Geissmann F (июнь 2023 г.). «Физиология и заболевания макрофагов с тканевыми резидентами» . Природа . 618 (7966): 698–707. Bibcode : 2023natur.618..698L . doi : 10.1038/s41586-023-06002-x . PMC 10649266 . PMID 37344646 .
| Базы данных управления авторитетом : национальный |
|---|