Адаптивная иммунная система
Эта статья нуждается в дополнительных цитатах для проверки . ( март 2024 г. ) |

Адаптивная иммунная система , также известная как приобретенная иммунная система или специфическая иммунная система, представляет собой подсистему иммунной системы , состоящую из специализированных системных клеток и процессов, которые уничтожают патогены или предотвращают их рост. Приобретенная иммунная система — одна из двух основных стратегий иммунитета , обнаруженных у позвоночных (вторая — врожденная иммунная система ).

Как и врожденная система, адаптивная иммунная система включает в себя как гуморального иммунитета компоненты , так и компоненты клеточного иммунитета и уничтожает вторгшиеся патогены. В отличие от врожденной иммунной системы , которая заранее запрограммирована реагировать на общие категории патогенов, адаптивная иммунная система очень специфична для каждого конкретного патогена, с которым столкнулся организм. [1]
Адаптивный иммунитет создает иммунологическую память после первоначальной реакции на конкретный патоген и приводит к усилению реакции на будущие встречи с этим патогеном. Антитела являются важной частью адаптивной иммунной системы. Адаптивный иммунитет может обеспечить длительную защиту, иногда на всю жизнь человека. Например, человек, выздоровевший от кори, теперь защищен от кори на всю жизнь; в других случаях он не обеспечивает пожизненной защиты, как при ветряной оспе . Этот процесс адаптивного иммунитета лежит в основе вакцинации . [ нужна ссылка ]
Клетки, которые осуществляют адаптивный иммунный ответ, представляют собой лейкоциты, известные как лимфоциты . В-клетки и Т-клетки , два разных типа лимфоцитов, выполняют основные функции: реакцию антител и клеточно-опосредованный иммунный ответ. В ответ на антитела В-клетки активируются и секретируют антитела , которые представляют собой белки, также известные как иммуноглобулины. Антитела проходят через кровоток и связываются с чужеродным антигеном, вызывая его инактивацию, что не позволяет антигену связываться с хозяином. [2] Антигены – это любые вещества, вызывающие адаптивный иммунный ответ. Иногда адаптивная система не способна отличить вредные чужеродные молекулы от безвредных; Последствиями этого могут быть сенная лихорадка , астма или любая другая аллергия .
При адаптивном иммунитете специфичные к возбудителю рецепторы «приобретаются» в течение жизни организма (тогда как при врожденном иммунитете специфичные к возбудителю рецепторы уже закодированы в геноме ). Эта приобретенная реакция называется «адаптивной», поскольку она готовит иммунную систему организма к будущим вызовам (хотя на самом деле она также может быть неадаптивной , когда приводит к аллергии или аутоиммунитету ).
Система легко адаптируется по двум причинам. Во-первых, соматическая гипермутация — это процесс ускоренных случайных генетических мутаций в генах, кодирующих антитела, который позволяет создавать антитела с новой специфичностью. Во-вторых, рекомбинация V(D)J случайным образом выбирает одну переменную (V), одно разнообразие (D),и одна соединительная область (J) для генетической рекомбинации и отбрасывание остальных, что создает уникальную комбинацию сегментов гена антиген-рецептор в каждом лимфоците. Этот механизм позволяет небольшому количеству генетических сегментов генерировать огромное количество различных антигенных рецепторов, которые затем уникальным образом экспрессируются на каждом отдельном лимфоците . Поскольку перестройка генов приводит к необратимым изменениям в ДНК каждой клетки, все потомки (потомки) этой клетки наследуют гены, которые кодируют одну и ту же специфичность рецептора, включая В-клетки памяти и Т-клетки памяти , которые являются ключом к долгоживущим. специфический иммунитет.
Мы
[ редактировать ]Термин «адаптивный» впервые был использован Робертом Гудом в отношении реакции антител у лягушек в качестве синонима «приобретенного иммунного ответа» в 1964 году. Гуд признал, что использовал эти термины как синонимы, но объяснил лишь, что предпочитает использовать термин «адаптивный». ". Возможно, он имел в виду тогда еще не невероятную теорию образования антител, согласно которой антитела были пластичными и могли адаптироваться к молекулярной форме антигенов, и/или к концепции «адаптивных ферментов», описанной Моно у бактерий, т.е. , ферменты, экспрессия которых может быть индуцирована их субстратами. Эту фразу использовали почти исключительно Гуд, его ученики и несколько других иммунологов, работающих с маргинальными организмами, до 1990-х годов, когда она стала широко использоваться в тандеме с термином «врожденный иммунитет», который стал популярной темой после открытия рецепторной системы Толля. у дрозофилы , ранее маргинального организма для изучения иммунологии. Термин «адаптивный», используемый в иммунологии, проблематичен, поскольку приобретенные иммунные реакции могут быть как адаптивными, так и дезадаптивными в физиологическом смысле. Действительно, как приобретенные, так и врожденные иммунные реакции могут быть как адаптивными, так и дезадаптивными в эволюционном смысле. Большинство учебников сегодня, после раннего использования Джейнвей , почти исключительно используйте слово «адаптивный» и отмечайте в глоссариях, что этот термин является синонимом «приобретенного». [ нужна ссылка ]
стал означать Классический смысл «приобретенного иммунитета» со времени открытия Тонегавы «антиген-специфический иммунитет, опосредованный соматическими перестройками генов, которые создают клон-определяющие антигенные рецепторы». В последнее десятилетие термин «адаптивный» все чаще применяется к другому классу иммунного ответа, пока еще не связанному с перестройками соматических генов. К ним относятся экспансия естественных киллеров (NK) клеток с пока необъяснимой специфичностью к антигенам, экспансия NK-клеток, экспрессирующих рецепторы, кодируемые зародышевой линией, и активация других врожденных иммунных клеток в активированное состояние, которое обеспечивает кратковременную «иммунную память». ". В этом смысле «адаптивный иммунитет» больше напоминает понятие «активированное состояние» или «гетеростаз», возвращаясь, таким образом, по смыслу к физиологическому смыслу «адаптации» к изменениям окружающей среды. [ нужна ссылка ]
Функции
[ редактировать ]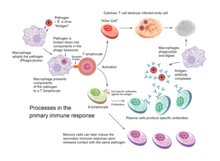
Приобретенный иммунитет запускается у позвоночных, когда патоген уклоняется от врожденной иммунной системы и (1) генерирует пороговый уровень антигена и (2) генерирует сигналы «незнакомца» или «опасности», активирующие дендритные клетки . [3]
К основным функциям приобретенной иммунной системы относятся: [ нужна ссылка ]
- Распознавание специфических «чужих» антигенов в присутствии «своих» в процессе презентации антигена .
- Генерация ответов, направленных на максимальное устранение конкретных патогенов или инфицированных патогенами клеток.
- Развитие иммунологической памяти , при которой патогены «запоминаются» посредством В-клеток памяти и Т-клеток памяти .
У людей адаптивной иммунной системе требуется 4–7 дней, чтобы обеспечить значимый ответ. [4]
Лимфоциты
[ редактировать ]Т- и В- лимфоциты являются клетками адаптивной иммунной системы. В организме человека имеется около 2 триллионов лимфоцитов, которые составляют 20–40% лейкоцитов; их общая масса примерно такая же, как у мозга или печени . Периферический кровоток содержит только 2% всех циркулирующих лимфоцитов; остальные 98% перемещаются в тканях и лимфатической системе , включая лимфатические узлы и селезенку . [2] У людей примерно 1–2% пула лимфоцитов циркулирует каждый час, что увеличивает возможность клеток встретиться с конкретным патогеном и антигеном, на которые они реагируют. [5]
В-клетки и Т-клетки происходят из одних и тех же мультипотентных гемопоэтических стволовых клеток и выглядят одинаково друг с другом до тех пор, пока не будут активированы. В-клетки играют большую роль в гуморальном иммунном ответе , тогда как Т-клетки тесно вовлечены в клеточно-опосредованные иммунные реакции . У всех позвоночных, кроме Agnatha , В- и Т-клетки производятся стволовыми клетками костного мозга. [6] Предшественники Т-клеток затем мигрируют из костного мозга в тимус , где развиваются дальше.
У взрослого животного периферические лимфоидные органы содержат смесь В- и Т-клеток, находящихся как минимум на трех стадиях дифференцировки: [ нужна ссылка ]
- Наивные В- и наивные Т-клетки , которые покинули костный мозг или тимус и попали в лимфатическую систему, но еще не встретили соответствующий антиген.
- Эффекторные клетки , которые активируются соответствующим антигеном и активно участвуют в уничтожении патогена.
- Клетки памяти, пережившие прошлые инфекции
Презентация антигена
[ редактировать ]Приобретенный иммунитет основан на способности иммунных клеток различать собственные клетки организма и нежелательные захватчики.Клетки хозяина экспрессируют «собственные» антигены . Эти антигены отличаются от антигенов на поверхности бактерий или на поверхности инфицированных вирусом клеток-хозяев («чужие» или «чужие» антигены). Приобретенный иммунный ответ запускается путем распознавания чужеродного антигена в клеточном контексте активированной дендритной клетки. [ нужна ссылка ]
За исключением безъядерных клеток (включая эритроциты ), все клетки способны презентировать антиген посредством функции молекул главного комплекса гистосовместимости (MHC). [6] Некоторые клетки специально оборудованы для презентации антигена и для первичной активации наивных Т-клеток. Дендритные клетки , В-клетки и макрофаги оснащены специальными «костимулирующими» лигандами, распознаваемыми костимулирующими рецепторами на Т-клетках, и называются профессиональными антигенпрезентирующими клетками (APC). [ нужна ссылка ]
Несколько подгрупп Т-клеток могут быть активированы профессиональными APC, и каждый тип Т-клеток специально оснащен для борьбы с каждым уникальным токсином или микробным патогеном. Тип активированных Т-клеток и тип генерируемого ответа частично зависят от контекста, в котором APC впервые встретил антиген. [3]
Экзогенные антигены
[ редактировать ]
Дендритные клетки поглощают экзогенные патогены, такие как бактерии, паразиты или токсины, в тканях, а затем мигрируют посредством хемотаксических сигналов в лимфатические узлы, обогащенные Т-клетками. Во время миграции дендритные клетки подвергаются процессу созревания, в ходе которого они теряют большую часть своей способности поглощать другие патогены и развивают способность взаимодействовать с Т-клетками. Дендритная клетка использует ферменты для измельчения патогена на более мелкие кусочки, называемые антигенами . В лимфатическом узле дендритная клетка отображает эти чужеродные антигены на своей поверхности, связывая их с рецептором, называемым главным комплексом гистосовместимости , или MHC (также известным у людей как человеческий лейкоцитарный антиген (HLA)). Этот комплекс MHC-антиген распознается Т-клетками, проходящими через лимфатический узел. Экзогенные антигены обычно отображаются на молекулах MHC класса II , которые активируют CD4 + Т-хелперные клетки . [3]
Эндогенные антигены
[ редактировать ]Эндогенные антигены продуцируются внутриклеточными бактериями и вирусами, реплицирующимися внутри клетки-хозяина. Клетка-хозяин использует ферменты для переваривания белков, связанных с вирусом, и отображает эти фрагменты на своей поверхности Т-клеткам, связывая их с MHC. Эндогенные антигены обычно отображаются на молекулах MHC класса I и активируют CD8 + цитотоксические Т-клетки. За исключением безъядерных клеток (включая эритроциты ), MHC класса I экспрессируется всеми клетками-хозяевами. [3]
Т-лимфоциты
[ редактировать ]CD8+ Т-лимфоциты и цитотоксичность
[ редактировать ]Цитотоксические Т-клетки (также известные как ТС, Т-клетки-киллеры или цитотоксические Т-лимфоциты (ЦТЛ)) представляют собой подгруппу Т-клеток, которые вызывают гибель клеток, инфицированных вирусами (и другими патогенами) или иным образом поврежден или неработоспособен. [3]
Наивные цитотоксические Т-клетки активируются, когда их Т-клеточный рецептор (TCR) сильно взаимодействует с связанной с пептидом молекулой MHC класса I. Это сродство зависит от типа и ориентации комплекса антиген/MHC и является тем, что удерживает CTL и инфицированную клетку вместе. [3] После активации CTL подвергается процессу, называемому клональной селекцией , в ходе которого он приобретает функции и быстро делится, образуя армию «вооруженных» эффекторных клеток. Активированный CTL затем путешествует по всему организму в поисках клеток, несущих этот уникальный пептид MHC класса I +. [ нужна ссылка ]
При воздействии этих инфицированных или дисфункциональных соматических клеток эффекторные CTL выделяют перфорин и гранулизин клетки-мишени : цитотоксины, которые образуют поры в плазматической мембране , позволяя ионам и воде проникать в инфицированную клетку и вызывая ее разрыв или лизис . CTL высвобождают гранзим , сериновую протеазу, инкапсулированную в гранулу, которая проникает в клетки через поры, вызывая апоптоз (гибель клеток). Чтобы ограничить обширное повреждение тканей во время инфекции, активация CTL строго контролируется и, как правило, требует очень сильного сигнала активации MHC/антигена или дополнительных сигналов активации, обеспечиваемых «помощниками» Т-клетками (см. ниже). [3]
После разрешения инфекции большинство эффекторных клеток погибают, и фагоциты уничтожают их, но некоторые из этих клеток остаются в качестве клеток памяти. [6] При более поздней встрече с тем же антигеном эти клетки памяти быстро дифференцируются в эффекторные клетки, что значительно сокращает время, необходимое для формирования эффективного ответа. [ нужна ссылка ]
Т-хелперы
[ редактировать ]
CD4+-лимфоциты, также называемые Т-хелперами, являются медиаторами иммунного ответа и играют важную роль в установлении и максимизации возможностей приобретенного иммунного ответа. [3] Эти клетки не обладают цитотоксической или фагоцитарной активностью; и не может убивать инфицированные клетки или уничтожать патогены, но, по сути, «управляет» иммунным ответом, направляя другие клетки на выполнение этих задач. [ нужна ссылка ]
Т-хелперы экспрессируют Т-клеточные рецепторы (TCR), которые распознают антиген, связанный с молекулами MHC класса II. Активация наивной Т-хелперной клетки заставляет ее высвобождать цитокины, которые влияют на активность многих типов клеток, включая APC (антигенпрезентирующую клетку), которая ее активировала. Т-хелперам требуется гораздо более мягкий стимул активации, чем цитотоксическим Т-клеткам. Т-хелперы могут подавать дополнительные сигналы, которые «помогают» активировать цитотоксические клетки. [6]
Th1 и Th2: ответы Т-хелперов
[ редактировать ]Классически выделяют два типа эффектора CD4. + Реакции Т-хелперных клеток могут быть индуцированы профессиональными АПК, обозначенными Th1 и Th2, каждый из которых предназначен для устранения различных типов патогенов. Факторы, которые определяют, вызывает ли инфекция ответ типа Th1 или Th2, до конца не изучены, но возникающий ответ действительно играет важную роль в устранении различных патогенов. [3]
Ответ Th1 характеризуется выработкой интерферона-гамма , который активирует бактерицидную активность макрофагов и индуцирует В-клетки для выработки опсонизирующих (маркирующих для фагоцитоза) и связывающих комплемент антител и приводит к клеточно-опосредованному иммунитету . [3] В целом, ответы Th1 более эффективны против внутриклеточных патогенов (вирусов и бактерий, находящихся внутри клеток-хозяев).
Ответ Th2 характеризуется высвобождением интерлейкина 5 , который индуцирует эозинофилы в удалении паразитов. [8] Th2 также производит интерлейкин 4 , который облегчает переключение изотипа B-клеток . [3] В целом, ответы Th2 более эффективны против внеклеточных бактерий, паразитов, включая гельминтов и токсинов . [3] Как и цитотоксические Т-клетки, большая часть CD4 + хелперные клетки умирают после разрешения инфекции, при этом некоторые из них остаются в виде CD4. + ячейки памяти.
Научные исследования на мышах и людях все чаще дают убедительные доказательства более широкого разнообразия CD4. + подмножества эффекторных Т-хелперов. [9] [10] Регуляторные Т-клетки (Treg) были идентифицированы как важные негативные регуляторы адаптивного иммунитета, поскольку они ограничивают и подавляют иммунную систему, контролируя аберрантные иммунные реакции на аутоантигены; важный механизм контроля развития аутоиммунных заболеваний. [6] Фолликулярные Т-хелперы (Tfh) представляют собой еще одну популяцию эффекторных CD4. + Т-клетки, которые развиваются из наивных Т-клеток после активации антигена. Клетки Tfh специализируются на поддержке гуморального иммунитета В-клеток , поскольку они обладают уникальной способностью мигрировать к фолликулярным В-клеткам во вторичных лимфоидных органах и передавать им положительные паракринные сигналы, позволяющие генерировать и вызывать выработку высококачественных антител, созревших по аффинности . Подобно Treg, клетки Tfh также играют роль в иммунологической толерантности , поскольку аномальное увеличение количества клеток Tfh может привести к неограниченной выработке аутореактивных антител, вызывая тяжелые системные аутоиммунные нарушения. [11]
Актуальность CD4 + Т-хелперные клетки выделяются во время ВИЧ- инфекции. ВИЧ способен разрушить иммунную систему, специфически атакуя CD4. + Т-клетки, именно те клетки, которые могут способствовать выведению вируса, а также клетки, обеспечивающие иммунитет против всех других патогенов, встречающихся в течение жизни организма. [6]
Гамма-дельта Т-клетки
[ редактировать ]Гамма-дельта-Т-клетки (γδ-Т-клетки) обладают альтернативным Т-клеточным рецептором (TCR) в отличие от CD4+ и CD8+ αβ-Т-клеток и имеют общие характеристики Т-хелперов, цитотоксических Т-клеток и естественных клеток-киллеров. Подобно другим «нетрадиционным» подмножествам Т-клеток, несущих инвариантные TCR, таким как CD1d -рестриктированные естественные Т-киллеры Т-клетки , γδ-Т-клетки обладают характеристиками, которые ставят их на границу между врожденным и приобретенным иммунитетом. С одной стороны, γδ Т-клетки можно рассматривать как компонент адаптивного иммунитета, поскольку они перестраивают гены TCR посредством рекомбинации V(D)J , что также приводит к разнообразию соединений , и развивают фенотип памяти. С другой стороны, однако, различные подгруппы также можно рассматривать как часть врожденной иммунной системы, где ограниченные рецепторы TCR или NK могут использоваться в качестве рецепторов распознавания образов . Например, согласно этой парадигме, большое количество Т-клеток Vγ9/Vδ2 в течение нескольких часов реагируют на обычные молекулы, продуцируемые микробами, а сильно ограниченные внутриэпителиальные Т-клетки Vδ1 реагируют на стрессированные эпителиальные клетки. [ нужна ссылка ]
B-лимфоциты и продукция антител
[ редактировать ]
В-клетки — это основные клетки, участвующие в выработке антител , циркулирующих в плазме крови и лимфе, известных как гуморальный иммунитет . Антитела (также известные как иммуноглобулины, Ig) представляют собой большие Y-образные белки, используемые иммунной системой для идентификации и нейтрализации инородных объектов. У млекопитающих имеется пять типов антител: IgA , IgD , IgE , IgG и IgM , различающихся биологическими свойствами; каждый из них эволюционировал, чтобы справляться с разными видами антигенов. При активации В-клетки вырабатывают антитела, каждое из которых распознает уникальный антиген и нейтрализует определенные патогены. [3]
Связывание антигена и антитела может вызвать пять различных защитных механизмов:
- Агглютинация: уменьшает количество инфекционных единиц, с которыми необходимо иметь дело.
- Активация комплемента: вызывает воспаление и лизис клеток.
- Опсонизация: покрытие антигена антителом усиливает фагоцитоз.
- Антителозависимая клеточно-опосредованная цитотоксичность: антитела, прикрепленные к клетке-мишени, вызывают разрушение макрофагами, эозинофилами и NK-клетками.
- Нейтрализация: блокирует прилипание бактерий и вирусов к слизистой оболочке.
Как и Т-клетка, В-клетки экспрессируют уникальный В-клеточный рецептор (BCR), в данном случае мембраносвязанную молекулу антитела. Все BCR любого клона В-клеток распознают и связываются только с одним конкретным антигеном. Критическая разница между В-клетками и Т-клетками заключается в том, как каждая клетка «видит» антиген. Т-клетки распознают родственный антиген в процессированной форме – как пептид в контексте молекулы MHC. [3] тогда как B-клетки распознают антигены в их нативной форме. [3] Как только В-клетка встречает родственный (или специфический) антиген (и получает дополнительные сигналы от Т-хелперов (преимущественно типа Th2)), она далее дифференцируется в эффекторную клетку, известную как плазматическая клетка. [3]
Плазматические клетки — это короткоживущие клетки (2–3 дня), секретирующие антитела. Эти антитела связываются с антигенами, делая их более легкой мишенью для фагоцитов, и запускают каскад комплемента . [3] Около 10% плазматических клеток выживают и становятся долгоживущими антигенспецифическими В-клетками памяти . [3] Эти клетки, уже подготовленные к выработке специфических антител, могут быть призваны быстро отреагировать, если тот же патоген повторно заразит хозяина, в то время как у хозяина возникнет мало симптомов или они вообще возникнут.
Альтернативные системы
[ редактировать ]У бесчелюстных позвоночных
[ редактировать ]Примитивные бесчелюстные позвоночные , такие как минога и миксина , имеют адаптивную иммунную систему, которая демонстрирует 3 различных клеточных линии, каждая из которых имеет общее происхождение с В-клетками, αβ Т-клетками и врожденными γΔ Т-клетками. [12] [13] Вместо классических антител и Т-клеточных рецепторов эти животные обладают большим набором молекул, называемых вариабельными лимфоцитными рецепторами (сокращенно VLR), которые, как и антигенные рецепторы челюстных позвоночных, производятся лишь из небольшого количества (одного или двух) гены . Считается, что эти молекулы связывают патогенные антигены аналогично антителам и с той же степенью специфичности. [14]
У насекомых
[ редактировать ]Долгое время считалось, что насекомые и другие беспозвоночные обладают только врожденной иммунной системой . Однако в последние годы у насекомых были обнаружены некоторые основные признаки адаптивного иммунитета. Этими чертами являются иммунная память и специфичность. Хотя отличительные признаки присутствуют, механизмы отличаются от таковых у позвоночных .
Иммунная память у насекомых была открыта благодаря феномену прайминга. Когда насекомые подвергаются воздействию несмертельной дозы или убитых при нагревании бактерий, у них может развиться память об этой инфекции, которая позволяет им противостоять в противном случае смертельной дозе тех же бактерий, воздействию которых они подвергались раньше. [15] [16] В отличие от позвоночных, у насекомых нет клеток, специфичных для адаптивного иммунитета. Вместо этого эти механизмы опосредуются гемоцитами . Гемоциты функционируют аналогично фагоцитам и после праймирования способны более эффективно распознавать и поглощать возбудитель. [17] Также было показано, что можно передать память потомству. Например, у медоносных пчел , если матка заражена бактериями, новорожденные рабочие обладают повышенными способностями в борьбе с теми же бактериями. [18] Другая экспериментальная модель, основанная на красном мучном жуке, также продемонстрировала передачу памяти, специфичной для патогена, потомству как от матери, так и от отца. [19]
Наиболее общепринятая теория специфичности основана на гене Dscam . Ген Dscam, также известный как адгезивная молекула клеток синдрома Дауна, представляет собой ген, который содержит 3 вариабельных домена Ig . Эти домены можно альтернативно соединять, достигая большого количества вариаций. [20] Показано, что после воздействия различных патогенов образуются разные сплайсинговые формы dscam. После того как животные с разными формами сплайсинга подвергаются воздействию одного и того же патогена, выживают только особи со специфичной для этого патогена формой сплайсинга. [20]
Другим механизмом, поддерживающим специфичность иммунитета насекомых, является РНК-интерференция (РНКи). РНКи представляет собой форму противовирусного иммунитета с высокой специфичностью. [21] У него есть несколько различных путей, каждый из которых заканчивается тем, что вирус не может размножаться. Одним из путей является siRNA, в которой длинная двухцепочечная РНК разрезается на части, которые служат матрицами для белкового комплекса Ago2-RISC, который находит и разрушает комплементарную РНК вируса. Путь микроРНК в цитоплазме связывается с комплексом Ago1-RISC и действует как матрица для деградации вирусной РНК. Последний — piRNA , где небольшая РНК связывается с семейством белков Piwi и контролирует транспозоны и другие мобильные элементы. [22] Несмотря на исследования, точные механизмы, ответственные за иммунную реакцию и специфичность у насекомых, недостаточно хорошо описаны.
Иммунологическая память
[ редактировать ]Когда В-клетки и Т-клетки активируются, некоторые из них становятся В-клетками памяти , а некоторые Т-клетками памяти . На протяжении всей жизни животного эти клетки памяти формируют базу данных эффективных В- и Т-лимфоцитов. При взаимодействии с ранее встреченным антигеном соответствующие клетки памяти выбираются и активируются. Таким образом, второе и последующие воздействия антигена вызывают более сильный и быстрый иммунный ответ. Это «адаптивно» в том смысле, что иммунная система организма готовится к будущим вызовам, но, конечно, она «неадаптивна», если рецепторы аутоиммунны. Иммунологическая память может быть в форме пассивной кратковременной памяти или активной долговременной памяти. [ нужна ссылка ]
Пассивная память
[ редактировать ]Пассивная память обычно кратковременна и длится от нескольких дней до нескольких месяцев. Новорожденные младенцы ранее не подвергались воздействию микробов и особенно уязвимы к инфекциям. Мать обеспечивает несколько уровней пассивной защиты. Внутриутробно , материнские IgG транспортируются непосредственно через плаценту поэтому при рождении у младенцев наблюдается высокий уровень антител с тем же диапазоном антигенной специфичности, что и у их матери. [3] Грудное молоко содержит антитела (в основном IgA), которые передаются в кишечник ребенка, защищая от бактериальных инфекций, пока новорожденный не сможет синтезировать собственные антитела. [3]
Это пассивный иммунитет , поскольку плод на самом деле не вырабатывает никаких клеток памяти или антител: он только заимствует их. Кратковременный пассивный иммунитет также может передаваться искусственно от одного человека к другому через сыворотку , богатую антителами . [ нужна ссылка ]
Активная память
[ редактировать ]Как правило, активный иммунитет является долгосрочным и может быть приобретен путем заражения с последующей активацией В- и Т-клеток или искусственно приобретен с помощью вакцин в процессе, называемом иммунизацией.
Иммунизация
[ редактировать ]Исторически инфекционные заболевания были основной причиной смертности населения. За последнее столетие были разработаны два важных фактора борьбы с их распространением: санитария и иммунизация . [6] Иммунизация (обычно называемая вакцинацией ) представляет собой преднамеренную индукцию иммунного ответа и представляет собой единственную наиболее эффективную манипуляцию иммунной системой, разработанную учеными. [6] Иммунизация успешна, потому что она использует как естественную специфичность иммунной системы, так и ее индуцируемость. [ нужна ссылка ]
Принцип иммунизации заключается во введении антигена, полученного из болезнетворного организма, который стимулирует иммунную систему к выработке защитного иммунитета против этого организма, но сам по себе не вызывает патогенных эффектов этого организма. Антиген . (сокращение от антител » «генератор ) определяется как любое вещество, которое связывается со специфическим антителом и вызывает адаптивный иммунный ответ [2]
Большинство вирусных вакцин основано на живых аттенуированных вирусах, тогда как многие бактериальные вакцины основаны на бесклеточных компонентах микроорганизмов, в том числе на безвредных токсинных компонентах. [2] Многие антигены, полученные из бесклеточных вакцин, не вызывают сильного адаптивного ответа, а большинство бактериальных вакцин требуют добавления адъювантов , которые активируют антигенпрезентирующие клетки врожденной иммунной системы для повышения иммуногенности . [6]
Иммунологическое разнообразие
[ редактировать ]
большинство крупных молекул, включая практически все белки и многие полисахариды . Антигенами могут служить [3] Части антигена, которые взаимодействуют с молекулой антитела или рецептором лимфоцита, называются эпитопами или антигенными детерминантами. Большинство антигенов содержат множество эпитопов и могут стимулировать выработку антител, специфические реакции Т-клеток или и то, и другое. [3] Очень небольшая часть (менее 0,01%) всех лимфоцитов способна связываться с определенным антигеном, что позволяет предположить, что только несколько клеток реагируют на каждый антиген. [6]
Чтобы приобретенная реакция «запоминала» и устраняла большое количество патогенов, иммунная система должна уметь различать множество различных антигенов. [2] а рецепторы, распознающие антигены, должны создаваться в огромном разнообразии конфигураций, по сути, по одному рецептору (по крайней мере) для каждого отдельного патогена, который может когда-либо встретиться. Даже при отсутствии антигенной стимуляции человек может производить более 1 триллиона различных молекул антител. [6] Для хранения генетической информации, которая производит эти рецепторы, потребуются миллионы генов, но весь геном человека содержит менее 25 000 генов . [23]
Множество рецепторов производятся посредством процесса, известного как клональная селекция . [2] [3] Согласно теории клонального отбора, при рождении животное случайным образом генерирует огромное разнообразие лимфоцитов (каждый из которых несет уникальный антигенный рецептор) на основе информации, закодированной в небольшом семействе генов. Чтобы создать каждый уникальный антигенный рецептор, эти гены подверглись процессу, называемому рекомбинацией V(D)J , или комбинаторной диверсификацией , при котором один сегмент гена рекомбинируется с другими сегментами гена, образуя один уникальный ген. Этот процесс сборки генерирует огромное разнообразие рецепторов и антител еще до того, как организм встретит антигены, и позволяет иммунной системе реагировать на почти неограниченное разнообразие антигенов. [3] На протяжении всей жизни животного лимфоциты, которые могут реагировать на антигены, с которыми животное действительно сталкивается, отбираются для действия, направленного против всего, что экспрессирует этот антиген. [ нужна ссылка ]
Врожденная и приобретенная части иммунной системы работают вместе, а не вопреки друг другу. Приобретенные руки, В- и Т-клетки не могли функционировать без участия врожденной системы. Т-клетки бесполезны без антигенпрезентирующих клеток, которые их активируют, а В-клетки отключаются без помощи Т-клеток. С другой стороны, врожденная система, скорее всего, будет переполнена патогенами без специального действия адаптивного иммунного ответа.
Приобретенный иммунитет во время беременности
[ редактировать ]Краеугольным камнем иммунной системы является распознавание «своего» и «чужого». Поэтому особый интерес представляют механизмы, защищающие человеческий плод (который считается «чужим») от атаки иммунной системы. Хотя не было найдено исчерпывающего объяснения этого загадочного и часто повторяющегося отсутствия отторжения, две классические причины могут объяснить, как переносится плод. Во-первых, плод занимает часть тела, защищенную неиммунологическим барьером — маткой , которую иммунная система обычно не патрулирует. [3] Во-вторых, сам плод может способствовать местной иммуносупрессии у матери, возможно, за счет активного истощения питательных веществ. [3] Более современное объяснение индукции толерантности состоит в том, что специфические гликопротеины, экспрессируемые в матке во время беременности , подавляют иммунный ответ матки (см. eu-FEDS ). [ нужна ссылка ]
Во время беременности у живородящих млекопитающих (все млекопитающие, кроме Monotremes ) эндогенные ретровирусы (ERV) активируются и производятся в больших количествах во время имплантации эмбриона. В настоящее время известно, что они обладают иммуносупрессивными свойствами, что позволяет предположить их роль в защите эмбриона от иммунной системы матери. Кроме того, слитые вирусные белки вызывают образование плацентарного синцития. [24] ограничить обмен мигрирующими клетками между развивающимся эмбрионом и телом матери (чего эпителий не может сделать в достаточной степени, поскольку определенные клетки крови специализируются на внедрении между соседними эпителиальными клетками). Иммунодепрессивное действие представляло собой исходное нормальное поведение вируса, подобное ВИЧ. Слитые белки были способом распространения инфекции на другие клетки путем простого слияния их с инфицированной (ВИЧ тоже делает это). Считается, что предки современных живородящих млекопитающих развились после заражения этим вирусом, что позволило плоду пережить иммунную систему матери. [25]
Проект генома человека обнаружил несколько тысяч ERV, разделенных на 24 семейства. [26]
Теория иммунной сети
[ редактировать ]Теоретической основой, объясняющей работу приобретенной иммунной системы, является теория иммунной сети , основанная на взаимодействии между идиотипами (уникальными молекулярными особенностями одного клонотипа, т.е. уникальным набором антигенных детерминант вариабельной части антитела) и «анти- идиотипы (антигенные рецепторы, которые реагируют с идиотипом, как если бы это был чужеродный антиген). Эта теория, которая основывается на существующей гипотезе клонального отбора и с 1974 года разрабатывается в основном Нильсом Йерне и Джеффри В. Хоффманном , рассматривается как имеющая отношение к пониманию патогенеза ВИЧ и поиску вакцины против ВИЧ .
Стимуляция адаптивного иммунитета
[ редактировать ]Одним из наиболее интересных достижений биомедицинской науки за последние несколько десятилетий стало выяснение механизмов, обеспечивающих врожденный иммунитет. Один набор врожденных иммунных механизмов является гуморальным, например, активация комплемента . Другой набор включает рецепторы распознавания образов, такие как toll-подобные рецепторы , которые индуцируют выработку интерферонов и других цитокинов, повышающих устойчивость клеток, таких как моноциты, к инфекциям. [27] Цитокины, вырабатываемые при врожденных иммунных реакциях, относятся к активаторам адаптивных иммунных реакций. [27] Антитела оказывают аддитивное или синергическое действие с механизмами врожденного иммунитета. Нестабильные кластеры HbS Band-3, основной интегральный белок эритроцитов; [28] антитела распознают эти кластеры и ускоряют их удаление фагоцитирующими клетками. Кластерные белки Band 3 с присоединенными антителами активируют комплемент, а фрагменты C3 комплемента представляют собой опсонины, распознаваемые рецептором комплемента CR1 на фагоцитирующих клетках. [29]
Популяционное исследование показало, что защитный эффект серповидноклеточного признака против малярии falciparum включает усиление приобретенных, а также врожденных иммунных реакций на малярийного паразита, что иллюстрирует ожидаемый переход от врожденного к приобретенному иммунитету. [30]
Повторные инфекции малярии укрепляют приобретенный иммунитет и расширяют его действие против паразитов, экспрессирующих различные поверхностные антигены . К школьному возрасту у большинства детей вырабатывается эффективный адаптивный иммунитет против малярии. Эти наблюдения поднимают вопросы о механизмах, которые способствуют выживанию большинства детей в Африке, в то же время позволяя некоторым из них развивать потенциально смертельные инфекции.
При малярии, как и при других инфекциях, [27] врожденные иммунные реакции приводят к адаптивным иммунным реакциям и стимулируют их. Генетический контроль врожденного и приобретенного иммунитета в настоящее время является крупной и процветающей дисциплиной.
Гуморальные и клеточно-опосредованные иммунные реакции ограничивают размножение малярийных паразитов, а многие цитокины способствуют патогенезу малярии, а также разрешению инфекций. [31]
Эволюция
[ редактировать ]Приобретенная иммунная система, которая лучше всего изучена у млекопитающих, возникла у челюстных рыб примерно 500 миллионов лет назад. Большинство молекул, клеток, тканей и связанных с ними механизмов этой системы защиты обнаружено у хрящевых рыб . [32] Рецепторы лимфоцитов , Ig и TCR, обнаружены у всех челюстных позвоночных. Самый древний класс Ig, IgM, связан с мембраной и затем секретируется при стимуляции В-клеток хрящевых рыб. Другой изотип, акульий IgW, родственен IgD млекопитающих. TCR, как α/β, так и γ/δ, обнаружены у всех животных, от челюстноротых до млекопитающих. Организация генных сегментов, претерпевающих генную перестройку , у хрящевых рыб отличается, они имеют кластерную форму по сравнению с транслоконной формой у костных рыб и млекопитающих. Подобно TCR и Ig, MHC обнаружен только у челюстных позвоночных. Гены, участвующие в антигена процессинге и презентации , а также гены класса I и класса II тесно связаны в MHC почти всех изученных видов. [ нужна ссылка ]
Лимфоидные клетки можно обнаружить у некоторых вторичноротых предпозвоночных ( например, у морских ежей). [33] Они связывают антиген с рецепторами распознавания образов (PRR) врожденной иммунной системы . У бесчелюстных рыб две подгруппы лимфоцитов используют вариабельные лимфоцитарные рецепторы (VLR) для связывания антигена. [34] Разнообразие создается за счет опосредованной цитозиндезаминазой перестройки сегментов ДНК на основе LRR . [35] Нет никаких доказательств существования генов, активирующих рекомбинацию (RAG), которые перестраивают сегменты генов Ig и TCR у челюстных позвоночных.
Считается, что эволюция AIS, основанная на молекулах Ig, TCR и MHC, возникла в результате двух основных эволюционных событий: переноса RAG транспозона (возможно, вирусного происхождения) и двух дупликаций целого генома . [32] Хотя молекулы АИС хорошо консервативны, они также быстро развиваются. Тем не менее, сравнительный подход показывает, что многие характеристики довольно одинаковы для всех таксонов. Все основные функции АИС возникли рано и быстро. У бесчелюстных рыб другой AIS , который основан на перестройке генов для генерации разнообразных иммунных рецепторов с функциональной дихотомией, аналогичной молекулам Ig и TCR. [36] Врожденная иммунная система , играющая важную роль в активации АИС, является наиболее важной защитной системой беспозвоночных и растений .
Виды приобретенного иммунитета
[ редактировать ]Иммунитет может быть приобретен как активно, так и пассивно. Иммунитет приобретается активно, когда человек подвергается воздействию посторонних веществ, и иммунная система реагирует. Пассивный иммунитет – это когда антитела передаются от одного хозяина к другому. Как активно приобретенный, так и пассивно приобретенный иммунитет можно получить естественным или искусственным путем.
- Естественно приобретенный активный иммунитет – когда человек естественным образом подвергается воздействию антигенов, заболевает, а затем выздоравливает.
- Естественно приобретенный пассивный иммунитет – предполагает естественную передачу антител от матери к ребенку. Антитела проникают через плаценту женщины к плоду. Антитела также могут передаваться через грудное молоко с выделениями молозива.
- Искусственно приобретенный активный иммунитет – достигается путем вакцинации (введение мертвого или ослабленного антигена в клетку хозяина).
- Искусственно приобретенный пассивный иммунитет – предполагает введение в организм человека антител, а не антигенов. Эти антитела получены от животного или человека, у которого уже есть иммунитет к заболеванию.
| Естественно приобретенный | Искусственно приобретенный |
|---|---|
| Активный – антиген поступает в организм естественным путем. | Активный – антигены вводятся в состав вакцин. |
| Пассивный – антитела передаются от матери к плоду через плаценту или к ребенку через материнское молоко. | Пассивный – предварительно сформированные антитела в иммунную сыворотку вводятся путем инъекции. |
См. также
[ редактировать ]Примечания и ссылки
[ редактировать ]- Примечания
- Ссылки
- ^ Врожденная и адаптивная иммунная система . Институт качества и эффективности здравоохранения (IQWiG). 4 августа 2016 г.
- ^ Jump up to: а б с д и ж Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтерс П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк и Лондон: Garland Science. ISBN 0-8153-3218-1 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Джейнвей Калифорния, Трэверс П., Уолпорт М., Шломчик М.Дж. (2001). Иммунобиология (5-е изд.). Нью-Йорк и Лондон: Garland Science. ISBN 0-8153-4101-6 .
- ^ Чарльз Джейнвей-младший; Трэверс, Пол; Уолпорт, Марк; Шломчик, Марк Дж. (2001). «Принципы врожденного и адаптивного иммунитета» . Иммунобиология: иммунная система в здоровье и болезни. 5-е издание .
- ^ «Онлайн-учебник по микробиологии и иммунологии» . Медицинский факультет Университета Южной Каролины. Архивировано из оригинала 2 сентября 2008 года.
- ^ Jump up to: а б с д и ж г час я дж к Джейнвей Калифорния, Трэверс П., Уолпорт М., Шломчик М.Дж. (2005). Иммунобиология (6-е изд.). Гирляндная наука. ISBN 0-443-07310-4 .
- ^ Jump up to: а б с «Понимание того, как работает иммунная система» (PDF) . Публикация НИЗ № 03-5423 . Национальный институт аллергии и инфекционных заболеваний, Национальный институт рака, Министерство здравоохранения и социальных служб США, Национальные институты здравоохранения. Сентябрь 2003 г. Архивировано из оригинала (PDF) 3 января 2007 г.
- ^ Спенсер Л.А., Веллер П.Ф. (2010). «Эозинофилы и иммунитет Th2: современные идеи» . Иммунология и клеточная биология . 88 (3): 250–56. дои : 10.1038/icb.2009.115 . ПМЦ 3589820 . ПМИД 20065995 .
- ^ Сабо П.А., Левитин Х.М., Мирон М., Снайдер М.Е., Сенда Т., Юань Дж. и др. (октябрь 2019 г.). «Одноклеточная транскриптомика Т-клеток человека выявляет ткани и признаки активации в здоровых и болезненных состояниях» . Природные коммуникации . 10 (1): 4706. Бибкод : 2019NatCo..10.4706S . дои : 10.1038/s41467-019-12464-3 . ПМК 6797728 . ПМИД 31624246 .
- ^ Маген А., Ни Дж., Чиуччи Т., Тамутоунур С., Чжао Ю., Мехта М. и др. (декабрь 2019 г.). «+ Т-клетки» . Отчеты по ячейкам . 29 (10): 3019–3032.e6. дои : 10.1016/j.celrep.2019.10.131 . ПМЦ 6934378 . ПМИД 31801070 .
- ^ Вайнштейн Дж. С., Эрнандес С. Г., Крафт Дж. (май 2012 г.). «Т-клетки, которые способствуют созреванию B-клеток при системном аутоиммунитете» . Иммунологические обзоры . 247 (1): 160–71. дои : 10.1111/j.1600-065x.2012.01122.x . ПМЦ 3334351 . ПМИД 22500839 .
- ^ Флайник, Мартин Ф. (19 марта 2018 г.). «Хладнокровный взгляд на адаптивный иммунитет» . Обзоры природы Иммунология . 18 (7): 438–53. дои : 10.1038/s41577-018-0003-9 . ПМК 6084782 . ПМИД 29556016 .
- ^ Касамацу, Дзюн (январь 2013 г.). «Эволюция врожденной и адаптивной иммунной системы у бесчелюстных позвоночных» . Микробиология и иммунология . 57 (1): 1–12. дои : 10.1111/j.1348-0421.2012.00500.x . ISSN 0385-5600 . ПМИД 22924515 .
- ^ Олдер М.Н., Рогозин И.Б., Айер Л.М., Глазко Г.В., Купер М.Д., Панцер З. (декабрь 2005 г.). «Разнообразие и функции рецепторов адаптивного иммунитета у бесчелюстных позвоночных» . Наука . 310 (5756): 1970–73. Бибкод : 2005Sci...310.1970A . дои : 10.1126/science.1119420 . ПМИД 16373579 .
- ^ Миконранта Л., Маппес Дж., Каукониитти М., Фрейтак Д. (март 2014 г.). «Иммунитет к насекомым: пероральный контакт с бактериальным патогеном вызывает реакцию свободных радикалов и защищает от повторной инфекции» . Границы в зоологии . 11 (1): 23. дои : 10.1186/1742-9994-11-23 . ПМЦ 3975449 . ПМИД 24602309 .
- ^ Садд Б.М., Шмид-Хемпель П. (июнь 2006 г.). «Иммунитет насекомых проявляет специфичность в защите при вторичном воздействии патогена» . Современная биология . 16 (12): 1206–10. Бибкод : 2006CBio...16.1206S . дои : 10.1016/j.cub.2006.04.047 . ПМИД 16782011 . S2CID 14436004 .
- ^ Фам Л.Н., Дионн М.С., Ширасу-Хиза М., Шнайдер Д.С. (март 2007 г.). «Специфический иммунный ответ у дрозофилы зависит от фагоцитов» . ПЛОС Патогены . 3 (3): е26. дои : 10.1371/journal.ppat.0030026 . ПМК 1817657 . ПМИД 17352533 .
- ^ Эрнандес Лопес Х., Шуэли В., Крайльсхайм К., Риссбергер-Галле Ю. (июнь 2014 г.). «Межпоколенческий иммунный прайминг у медоносных пчел» . Слушания. Биологические науки . 281 (1785): 20140454. doi : 10.1098/rspb.2014.0454 . ПМК 4024302 . ПМИД 24789904 .
- ^ Рот О, Йооп Г, Эггерт Х, Хилберт Дж, Дэниел Дж, Шмид-Хемпель П, Курц Дж (март 2010 г.). «Отцовский иммунный прайминг для потомства красного мучного жука Tribolium castaneum» . Журнал экологии животных . 79 (2): 403–13. Бибкод : 2010JAnEc..79..403R . дои : 10.1111/j.1365-2656.2009.01617.x . ПМИД 19840170 .
- ^ Jump up to: а б Донг Ю, Тейлор Х.Э., Димопулос Г. (июль 2006 г.). «AgDscam, гипервариабельный домен иммуноглобулина, содержащий рецептор врожденной иммунной системы Anopheles gambiae» . ПЛОС Биология . 4 (7): е229. дои : 10.1371/journal.pbio.0040229 . ПМК 1479700 . ПМИД 16774454 .
- ^ Меки И.К., Кариити Х.М., Паркер А.Г., Врейсен М.Дж., Рос В.И., Влак Дж.М., ван Орс М.М., Абд-Алла А.М. (ноябрь 2018 г.). «Противовирусный иммунный ответ, основанный на РНК-интерференции, против вируса гипертрофии слюнных желез у Glossina pallidipes» . БМК Микробиология . 18 (Приложение 1): 170. doi : 10.1186/s12866-018-1298-1 . ПМК 6251114 . ПМИД 30470195 .
- ^ Рубио М., Маэстро Дж.Л., Доктор Пиулахс, Belles X (июнь 2018 г.). «Консервативная ассоциация белков Argonaute 1 и 2 с путями микроРНК и миРНК на протяжении всей эволюции насекомых, от тараканов до мух». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1861 (6): 554–60. дои : 10.1016/j.bbagrm.2018.04.001 . hdl : 10261/168051 . ПМИД 29656113 . S2CID 4890483 .
- ^ Международный консорциум по секвенированию генома человека (октябрь 2004 г.). «Завершение эухроматической последовательности генома человека» . Природа . 431 (7011): 931–45. Бибкод : 2004Natur.431..931H . дои : 10.1038/nature03001 . ПМИД 15496913 .
- ^ Ми С., Ли Х, Ли Х, Вельдман ГМ, Финнерти Х, Рэйси Л., ЛаВалли Е, Тан XY, Эдуард П., Хоуз С., Кейт Дж. К., Маккой Дж. М. (февраль 2000 г.). «Синцитин представляет собой пленный белок оболочки ретровируса, участвующий в морфогенезе плаценты человека». Природа . 403 (6771): 785–89. Бибкод : 2000Natur.403..785M . дои : 10.1038/35001608 . ПМИД 10693809 . S2CID 4367889 .
- ^ Вильярреал ЛП. «Вирусы, которые создают нас: роль эндогенного ретровируса в эволюции плацентарных видов» . Калифорнийский университет в Ирвине (конспекты лекций). Архивировано из оригинала 15 июля 2007 г. Проверено 3 февраля 2008 г.
- ^ Вильярреал LP (октябрь 2001 г.). «Сохраняющиеся вирусы могут играть роль в эволюции хостов» . Новости АСМ . Архивировано из оригинала 8 мая 2009 г.
- ^ Jump up to: а б с Уэмацу С., Акира С. (май 2007 г.). «Толл-подобные рецепторы и интерфероны I типа» . Журнал биологической химии . 282 (21): 15319–23. дои : 10.1074/jbc.R700009200 . ПМИД 17395581 .
- ^ Куросс С.А., ранг BH, Hebbel RP (апрель 1988 г.). «Избыток гема во внутренних мембранах серповидных эритроцитов: возможная роль в окислении тиолов» (PDF) . Кровь . 71 (4): 876–82. дои : 10.1182/blood.V71.4.876.876 . ПМИД 3355895 .
- ^ Арезе П., Туррини Ф., Шварцер Э. (2005). «Диапазон 3/комплемент-опосредованное распознавание и удаление нормально стареющих и патологических эритроцитов человека» . Клеточная физиология и биохимия . 16 (4–6): 133–46. дои : 10.1159/000089839 . ПМИД 16301814 .
- ^ Уильямс Т.Н., Мванги Т.В., Робертс DJ, Александр Н.Д., Weatherall DJ, Вамбуа С., Корток М., Сноу Р.В., Марш К. (май 2005 г.). «Иммунная основа защиты от малярии с помощью серповидноклеточной анемии» . ПЛОС Медицина . 2 (5): е128. doi : 10.1371/journal.pmed.0020128 . ПМЦ 1140945 . ПМИД 15916466 .
- ^ Шофилд Л., Грау Г.Е. (сентябрь 2005 г.). «Иммунологические процессы в патогенезе малярии» . Обзоры природы. Иммунология . 5 (9): 722–35. дои : 10.1038/nri1686 . ПМИД 16138104 . S2CID 19594405 .
- ^ Jump up to: а б Флажник М.Ф., Касахара М. (январь 2010 г.). «Происхождение и эволюция адаптивной иммунной системы: генетические события и селективное давление» . Обзоры природы. Генетика . 11 (1): 47–59. дои : 10.1038/nrg2703 . ПМК 3805090 . ПМИД 19997068 .
- ^ Хибино Т., Лоза-Колл М., Мессье С., Мажеске А.Дж., Коэн А.Х., Тервиллигер Д.П., Бакли К.М., Броктон В., Наир С.В., Берни К., Фугманн С.Д., Андерсон М.К., Пансер З., Кэмерон Р.А., Смит Л.К., Раст Дж.П. ( декабрь 2006 г.). «Репертуар иммунных генов, закодированный в геноме пурпурного морского ежа» (PDF) . Биология развития . 300 (1): 349–65. дои : 10.1016/j.ydbio.2006.08.065 . ПМИД 17027739 .
- ^ Пансер З., Амемия К.Т., Эрхардт Г.Р., Цейтлин Дж., Гартланд Г.Л., Купер, доктор медицинских наук (июль 2004 г.). «Соматическая диверсификация вариабельных рецепторов лимфоцитов у бесчелюстной морской миноги» (PDF) . Природа 430 (6996): 174–80. Бибкод : 2004Nature.430..174P . дои : 10.1038/nature02740 . hdl : 2027.42/62870 . ПМИД 15241406 . S2CID 876413 .
- ^ Рогозин И.Б., Айер Л.М., Лян Л., Глазко Г.В., Листон В.Г., Павлов Ю.И., Аравинд Л., Панцер З. (июнь 2007 г.). «Эволюция и диверсификация антигенных рецепторов миноги: доказательства участия цитозиндезаминазы семейства AID-APOBEC». Природная иммунология . 8 (6): 647–56. дои : 10.1038/ni1463 . PMID 17468760 . S2CID 3658963 .
- ^ Бём Т. (май 2011 г.). «Принципы проектирования адаптивных иммунных систем». Обзоры природы. Иммунология . 11 (5): 307–17. дои : 10.1038/nri2944 . ПМИД 21475308 . S2CID 25989912 .