Регуляторная Т-клетка
Регуляторные Т-клетки ( Treg / ˈ t iː r ɛ ɡ / или T reg- клетки), ранее известные как Т-супрессоры , представляют собой субпопуляцию Т-клеток , которые модулируют иммунную систему , поддерживают толерантность к аутоантигенам и предотвращают аутоиммунные заболевания. . T- reg -клетки обладают иммуносупрессивным действием и обычно подавляют или подавляют индукцию и пролиферацию эффекторных T-клеток . [1] T- reg- клетки экспрессируют биомаркеры CD4 , FOXP3 и CD25 и, как полагают, происходят из той же линии , что и наивный CD4. + клетки . [2] Поскольку эффекторные Т-клетки также экспрессируют CD4 и CD25, Т- reg -клетки очень трудно эффективно отличить от эффекторных CD4. + , что затрудняет их изучение. Исследования показали, что цитокин- трансформирующий фактор роста бета (TGF-β) необходим для T- reg -клеток от наивных CD4. дифференцировки + клеток и важен для поддержания Т- рег- клеток гомеостаза . [3]
Мышиные модели показали, что модуляция T- reg -клеток может лечить аутоиммунные заболевания и рак, а также может способствовать трансплантации органов. [4] и заживление ран . [5] Их последствия для рака сложны. У больных раком T- reg -клетки, как правило, активируются, и, по-видимому, они рекрутируются в места возникновения многих опухолей . Исследования как на людях, так и на животных моделях показали, что большое количество T- reg -клеток в микроокружении опухоли указывает на плохой прогноз , а T- reg -клетки, как полагают, подавляют опухолевый иммунитет, тем самым препятствуя врожденной способности организма контролировать рост раковых клеток. клетки. [6] Исследования в области иммунотерапии изучают, как регуляция Т-клеток может быть использована при лечении рака. [7]
Население
[ редактировать ]Т-регуляторные клетки являются компонентом иммунной системы, подавляющим иммунные реакции других клеток. Это важная «самопроверка», встроенная в иммунную систему для предотвращения чрезмерных реакций. Регуляторные Т-клетки существуют во многих формах, наиболее изученными из которых являются те, которые экспрессируют CD4, CD25 и FOXP3 (CD4 + CD25 + регуляторные Т-клетки). Эти Т- рег -клетки отличаются от Т-хелперов . [8] Другой субпопуляцией регуляторных Т-клеток являются клетки T reg 17. [9] Регуляторные Т-клетки участвуют в подавлении иммунных реакций после успешной ликвидации вторгшихся организмов, а также в предотвращении аутоиммунитета. [10]
CD4 + ФОКС3 + Регуляторные Т-клетки CD25 (высокого уровня) были названы «естественными» регуляторными Т-клетками. [11] чтобы отличить их от популяций «супрессорных» Т-клеток, которые генерируются in vitro . Дополнительные популяции регуляторных Т-клеток включают Tr1 , Th3 , CD8. + CD28 − и Т-клетки, ограниченные Qa-1. Вклад этих популяций в аутотолерантность и иммунный гомеостаз менее определен. FOXP3 можно использовать как хороший маркер мышиного CD4. + CD25 + Т-клетки, хотя недавние исследования также показали доказательства экспрессии FOXP3 в CD4. + CD25 − Т-клетки. У людей FOXP3 также экспрессируется недавно активированными обычными Т-клетками и, таким образом, не идентифицирует специфически человеческие T- reg . [12]
Разработка
[ редактировать ]Все Т-клетки происходят из клеток-предшественников в костном мозге , которые становятся приверженными своей линии в тимусе . Все Т-клетки начинаются с CD4 - CD8 - TCR -клеток на стадии DN (двойной отрицательный результат), когда отдельная клетка перестраивает свои гены рецепторов Т-клеток, чтобы сформировать уникальную функциональную молекулу, которую они, в свою очередь, тестируют на клетках. в коре тимуса для минимального уровня взаимодействия с собственным MHC . Если они получают эти сигналы, они пролиферируют и экспрессируют как CD4, так и CD8, становясь дважды положительными клетками. Селекция T reg происходит на радиорезистентных гематопоэтических клетках, экспрессирующих MHC класса II, в мозговом веществе или на тельцах Гассаля в тимусе. На стадии DP (двойной положительный результат) они отбираются путем взаимодействия с клетками тимуса, начинают транскрипцию Foxp3 и становятся T- reg -клетками, хотя они могут начать экспрессировать Foxp3 только на стадии одиночного положительного результата. в этот момент они являются функциональными T reg s. T reg не имеют ограниченной экспрессии TCR NKT или γδ Т-клеток; Т reg имеют большее разнообразие TCR, чем эффекторные Т-клетки, склонные к собственным пептидам.
Процесс выбора T- reg определяется аффинностью взаимодействия с аутопептидным комплексом MHC. Отбор на получение статуса T- reg - это процесс " Златовласки " - т.е. не слишком высокий, не слишком низкий, но в самый раз; [13] Т-клетка, получающая очень сильные сигналы, подвергается апоптотической гибели; клетка, получившая слабый сигнал, выживет и станет эффекторной клеткой. Если Т-клетка получит промежуточный сигнал, она станет регуляторной клеткой. Из-за стохастического характера процесса активации Т-клеток все популяции Т-клеток с данным TCR в конечном итоге будут иметь смесь T eff и T reg – относительных пропорций, определяемых сродством Т-клеток к собственному пептиду. -МХК. Даже в мышиных моделях с TCR-трансгенными клетками, отобранными на строме, секретирующей специфический антиген, делеция или конверсия не являются полными.
После взаимодействия с собственным пептидным комплексом MHC Т-клетка должна активировать IL-2R , CD25 и TNFR членов суперсемейства GITR , OX40 и TNFR2, чтобы стать CD25. + ФОКС3 - Предшественник Т- рег- клетки. Чтобы стать зрелым T reg , необходимо активировать транскрипционный фактор FOXP3, что приводится в действие зависимыми от γ-цепи (CD132) цитокинами, в частности IL-2 и/или IL-15. [14] [15] Только IL-2 недостаточно для стимуляции экспрессии Foxp3, необходимы другие цитокины. В то время как IL-2 продуцируется аутореактивными тимоцитами, IL-15 продуцируется стромальными клетками тимуса, главным образом mTEC и cTEC . [14]
другое подмножество предшественников T reg Недавно было идентифицировано . В этой подгруппе отсутствует CD25 и наблюдается низкая экспрессия Foxp3. Его развитие в основном зависит от IL-15. Эта подгруппа имеет более низкое сродство к аутоантигенам, чем CD25. + Фоксp3 высокий подмножество. Обе подгруппы генерируют зрелые T -reg -клетки после стимуляции IL-2 с сопоставимой эффективностью как in vitro , так и in vivo . CD25 + Фоксp3 высокий предшественники демонстрируют повышенный апоптоз и развиваются в зрелые T- reg -клетки с более быстрой кинетикой, чем Foxp3. низкий прародители. [16] T- reg, полученные из CD25 + Фоксp3 высокий предшественники защищают от экспериментального аутоиммунного энцефаломиелита, тогда как предшественники CD25 + Фоксp3 низкий индуцированного Т-клетками предшественники защищают от колита, . [14]
Зрелые CD25+Foxp3+ Treg также можно разделить на два разных подмножества в зависимости от уровня экспрессии CD25, GITR и PD-1 . Treg, экспрессирующие низкие количества CD25, GITR и PD-1, ограничивают развитие колита, способствуя конверсии обычного CD4. + Т-клетки в pTreg. Treg с высокой экспрессией CD25, GITR и PD-1 более самореактивны и контролируют лимфопролиферацию в периферических лимфатических узлах - они могут обладать способностью защищать от аутоиммунных заболеваний. [14]
Фоксp3 + Генерация T reg в тимусе задерживается на несколько дней по сравнению с клетками T eff и не достигает уровня взрослых ни в тимусе, ни на периферии примерно в течение трех недель после родов. T- reg -клеткам требуется CD28 костимуляция , а экспрессия B7.2 в значительной степени ограничена мозговым веществом, развитие которого, по-видимому, параллельно развитию Foxp3. + клетки. Было высказано предположение, что эти два процесса связаны, но окончательная связь между этими процессами пока не доказана. TGF-β не требуется для функциональности T reg в тимусе, поскольку тимические T reg от TGF-β нечувствительных мышей TGFβRII-DN являются функциональными.
Тимическая рециркуляция
[ редактировать ]Было замечено, что некоторые FOXP3 + T- reg -клетки возвращаются в тимус, где они и развились. Этот T- reg в основном присутствовал в мозговом веществе тимуса, который является основным местом дифференцировки T- reg -клеток. [17] Присутствие этих клеток в тимусе или добавление в культуру ткани тимуса плода подавляют развитие новых Т- рег- клеток на 34–60%, [17] но клетки Tconv не затрагиваются. Это означает, что рециркуляция Т- рег в тимус ингибирует клеток только de novo развитие Т- рег- .Молекулярный механизм этого процесса работает благодаря способности T- reg адсорбировать IL-2 из микроокружения, тем самым способствуя индуцированию апоптоза других Т-клеток, которым необходим IL-2 в качестве основного фактора роста. [18] Рециркулирующие T-reg-клетки в тимусе экспрессируют большое количество α-цепи высокоаффинного рецептора IL-2 ( CD25 ), кодируемой геном Il2ra , которая собирает IL-2 из мозгового вещества тимуса и снижает его концентрацию. Новый сгенерированный FOXP3 + T reg- клетки тимуса имеют не столь высокую степень экспрессии Il2ra . [17] IL-2 представляет собой цитокин, необходимый для развития Т- рег- клеток в тимусе. Он важен для пролиферации и выживания Т-клеток, но в случае его дефицита IL-15 может быть заменен. Однако развитие Т- рег- клеток зависит от IL-2. [19] У человека в тимусе обнаружена популяция CD31- отрицательных Т- reg -клеток. [17] CD31 можно использовать в качестве маркера вновь созданных Т- рег -клеток, как и других Т-лимфоцитов. Зрелые и периферические Т- рег- клетки снизили его экспрессию. [20] Поэтому вполне возможно, что этот регуляторный механизм развития Т- рег- клеток тимуса также функционален и у человека.
Вероятно, существует также положительная регуляция развития Т- рег- клеток тимуса, вызванная рециркуляцией Т- рег- клеток в тимус. Обнаружена популяция CD24 с низким содержанием FOXP3. + в тимусе с повышенной экспрессией IL-1R2 ( Il1r2 ) по сравнению с периферическими Т- рег- клетками. [21] [22] Высокая концентрация IL-1β , вызванная воспалением, снижает de novo развитие T- reg -клеток в тимусе. [22] Присутствие рециркулирующих T -reg- клеток в тимусе с высокой экспрессией IL1R2 во время воспалительных состояний помогает поглощать IL-1β и снижать его концентрацию в микроокружении мозгового вещества, таким образом, они способствуют развитию клеток de novo T -reg- . [22] Высокая концентрация IL-1β , вызванная воспалением, снижает de novo развитие T- reg -клеток в тимусе. [22] Связывание IL-1β с IL1R2 на поверхности T -reg -клеток не вызывает какой-либо передачи сигнала, поскольку отсутствует внутриклеточный ( TIR ) домен рецептора Toll-интерлейкина-1, который обычно присутствует в клетках врожденного иммунитета. [23]
Функция
[ редактировать ]Иммунная система должна уметь различать свое и чужое. Когда распознавание себя и чужого не удается, иммунная система разрушает клетки и ткани организма и в результате вызывает аутоиммунные заболевания . Регуляторные Т-клетки активно подавляют активацию иммунной системы и предотвращают патологическую самореактивность, т.е. аутоиммунное заболевание. О критической роли регуляторных Т-клеток в иммунной системе свидетельствует тяжелый аутоиммунный синдром, возникающий в результате генетической недостаточности регуляторных Т-клеток ( синдром IPEX – см. также ниже).
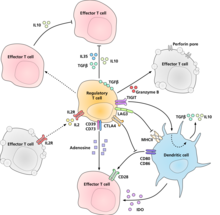
Молекулярный механизм, с помощью которого регуляторные Т-клетки проявляют свою супрессорную/регуляторную активность, окончательно не охарактеризован и является предметом интенсивных исследований. Эксперименты in vitro дали неоднозначные результаты относительно необходимости межклеточного контакта с подавляемой клеткой. Ниже представлены некоторые из предполагаемых механизмов иммуносупрессии:
- Регуляторные Т-клетки продуцируют ряд ингибирующих цитокинов. К ним относятся TGF-β, [24] Интерлейкин 35 , [25] и Интерлейкин 10 . [26] Также оказывается, что регуляторные Т-клетки могут побуждать другие типы клеток экспрессировать интерлейкин-10. [27]
- Регуляторные Т-клетки могут продуцировать гранзим B , который, в свою очередь, может индуцировать апоптоз эффекторных клеток. Сообщается, что регуляторные Т-клетки мышей с дефицитом гранзима B являются менее эффективными супрессорами активации эффекторных Т-клеток. [28]
- Обратная передача сигналов посредством прямого взаимодействия с дендритными клетками и индукции иммуносупрессивной индоламин-2,3-диоксигеназы . [29]
- Передача сигналов через эктоферменты CD39 и CD73 с продукцией иммуносупрессивного аденозина . [30] [31]
- Путем прямого взаимодействия с дендритными клетками посредством LAG3 и TIGIT . [32] [33] Этот обзор взаимодействий T reg с дендритными клетками обеспечивает различие между механизмами, описанными для клеток человека и клеток мыши. [34]
- Другой механизм управления осуществляется через петлю обратной связи IL-2. Активированные антигеном Т-клетки производят IL-2, который затем действует на рецепторы IL-2 регуляторных Т-клеток, предупреждая их о том, что в этой области происходит высокая активность Т-клеток, и они вызывают супрессорную реакцию против них. Это петля отрицательной обратной связи , гарантирующая отсутствие чрезмерной реакции. Если присутствует фактическая инфекция, другие воспалительные факторы подавляют подавление. Нарушение петли приводит к гиперреактивности, регуляция может изменить силу иммунного ответа. [35] Связанное с этим предположение в отношении интерлейкина 2 заключается в том, что активированные регуляторные Т-клетки поглощают интерлейкин 2 настолько жадно, что лишают эффекторные Т-клетки достаточного количества, чтобы избежать апоптоза. [18]
- Основным механизмом подавления регуляторными Т-клетками является предотвращение костимуляции через CD28 эффекторных Т-клеток под действием молекулы CTLA-4 . [36]
Естественные и индуцированные регуляторные Т-клетки
[ редактировать ]Т-регуляторные лимфоциты развиваются в процессе онтогенеза либо в тимусе , либо на периферии. Соответственно их разделяют на естественные и индуцированные Т-регуляторные клетки. [37]
Природные Т-регуляторные лимфоциты (tTregs, nTregs) характеризуются постоянной экспрессией FoxP3 и Т-клеточного рецептора (TCR) с относительно высоким аутоаффинностью. Эти клетки преимущественно обнаруживаются в организме в кровотоке или лимфатических узлах и служат главным образом для придания толерантности к аутоантигенам. [37]
Индуцированные (периферические) Т-регуляторные клетки (iTregs, pTregs) возникают в определенных ситуациях в присутствии IL-2 и TGF-b на периферии и начинают индуцируемо экспрессировать FoxP3, становясь таким образом функциональным эквивалентом клеток tTreg. Однако iTreg обнаруживаются преимущественно в периферических барьерных тканях, где они в первую очередь участвуют в предотвращении воспаления в присутствии внешних антигенов. [37]
К основным признакам, дифференцирующим клетки tTreg и iTreg, относятся Helios и Neuropilin-1 , наличие которых предполагает происхождение из тимуса. Еще одной особенностью, отличающей эти две популяции Treg-клеток, является стабильность экспрессии FoxP3 в разных условиях. [37]
Индуцированные Т-регуляторные клетки
[ редактировать ]Индуцированные регуляторные Т-клетки (iT reg ) (CD4 + CD25 + ФОКС3 + ) представляют собой супрессорные клетки, участвующие в толерантности. iT reg Было показано, что клетки подавляют пролиферацию Т-клеток и экспериментальные аутоиммунные заболевания. Эти клетки включают клетки T reg 17 . iT reg -клетки развиваются из зрелого CD4 + обычные Т-клетки вне тимуса: определяющее различие между естественными регуляторными Т-клетками (nT reg ) и клетками iT reg . Хотя клетки iT reg и nT reg имеют схожую функцию, недавно было показано, что клетки iT reg представляют собой «важную неизбыточную регуляторную подгруппу, которая дополняет клетки nT reg , отчасти за счет расширения разнообразия TCR в рамках регуляторных ответов». [38] Резкое истощение пула iT- рег- клеток на мышиных моделях привело к воспалению и потере веса. Вклад клеток nT reg по сравнению с клетками iT reg в поддержание толерантности неизвестен, но оба они важны. Эпигенетические различия наблюдались между клетками nT reg и iT reg , причем первые имели более стабильную экспрессию FOXP3 и более широкое деметилирование .
Среда тонкого кишечника богата витамином А и является местом производства ретиноевой кислоты. [39] Ретиноевая кислота и TGF-бета, продуцируемые дендритными клетками в этой области, сигнализируют о выработке регуляторных Т-клеток. [39] противоположные , клеткам Th17 Витамин А и TGF-бета способствуют дифференцировке Т-клеток в регуляторные Т-клетки , даже в присутствии IL-6 . [40] [41] Кишечная среда может привести к индуцированию регуляторных Т-клеток с помощью TGF-бета и ретиноевой кислоты. [42] некоторые из них экспрессируют лектиноподобный рецептор CD161 и специализируются на поддержании целостности барьера за счет ускорения заживления ран. [43] T- рег в кишечнике дифференцируются от наивных Т-клеток после введения антигена. [44] Недавно было показано, что регуляторные Т-клетки человека могут быть индуцированы как из наивных, так и из предварительно коммитированных клеток Th1 и клеток Th17. [45] , полученного из паразита с использованием миметика TGF-β , секретируемого Heligmosomoides polygyrus и называемого Hp -TGM ( мимик TGF-β H.polygyrus ). [46] [47] Hp -TGM может индуцировать экспрессию регуляторных Т-клеток мышиного FOXP3 , которые были стабильны в присутствии воспаления in vivo . [48] Hp -TGM-индуцированные регуляторные Т-клетки FOXP3+ человека были стабильными в присутствии воспаления и имели повышенные уровни CD25 , CTLA4 и пониженное метилирование в FOXP3 T -reg -специфической деметилированной области по сравнению с TGF-β-индуцированными Treg . [45]
RORγt+ регуляторные Т-лимфоциты
[ редактировать ]Примерно 30–40% клеток FoxP3+ Treg толстой кишки экспрессируют фактор транскрипции RORγt. [49] iTreg способны дифференцироваться в клетки, экспрессирующие RORγt , и таким образом приобретать фенотип клеток Th17 . Эти клетки связаны с функциями лимфоидных тканей слизистой оболочки, таких как кишечный барьер. В собственной пластинке кишечника обнаружено 20–30% регуляторных клеток Foxp3+ T, экспрессирующих RORyt, и эта высокая доля сильно зависит от наличия сложного микробиома кишечника. У мышей без микробов (GF) популяция регуляторных клеток RORγt+ T сильно снижается, тогда как реколонизация специфической безпатогенной (SPF) микробиотой восстанавливает нормальное количество этих лимфоцитов в кишечнике. Механизм, с помощью которого микробиота кишечника индуцирует образование клеток RORγt+ Treg, включает выработку короткоцепочечных жирных кислот (КЦЖК), от которых зависит эта индукция. SCFAs являются побочным продуктом ферментации и переваривания пищевых волокон, поэтому мыши, свободные от микробов, имеют очень низкие концентрации как SCFAs, так и RORγt Treg-клеток. Индукция клеток RORγt Treg также зависит от присутствия дендритные клетки у взрослых, клетки Thetis у новорожденных и презентация антигена MHC II . [50] [51]
Клетки RORγt+ Treg отсутствуют в тимусе и не экспрессируют Helios или Neuropilin-1 , но имеют высокую экспрессию CD44 , IL-10 , ICOS, CTLA-4 и нуклеотидаз CD39 и CD73, что указывает на сильную регуляторную функцию. [50]
Функция регуляторных Т-лимфоцитов RORγt+
[ редактировать ]Индукция RORγt+ Treg-клеток в лимфатических узлах тонкой кишки имеет решающее значение для установления толерантности к просветному антигену кишечника . Эти клетки особенно важны для предотвращения пищевой аллергии. Одним из механизмов является выработка подавляющих молекул, таких как цитокин IL-10 . Эти клетки также подавляют популяцию клеток Th17 и ингибируют выработку IL-17 , подавляя тем самым провоспалительную реакцию. [50]
У мышей RORγt+ Treg толстой кишки отсутствуют в течение первых двух недель после рождения. Генерация RORγt+ Treg на раннем этапе после рождения необходима для предотвращения развития различных кишечных иммунопатологий в более позднем возрасте. Особенно важным является период постепенного перехода от использования исключительно материнского молока к введению твердой пищи, между 15 и 20 днями жизни, когда вводится большое количество микробных антигенов и комменсальная микробиота оседает в кишечнике. В это время защитные клетки RORγt+ Treg индуцируются микробными антигенами, и нормальный гомеостаз кишечника поддерживается за счет индукции толерантности к комменсальной микробиоте. Отсутствие индукции RORγt+ Treg-клеток привело у мышей к развитию тяжелого колита . [52] На количество RORγt+ Treg, индуцированных в раннем возрасте, влияет материнское молоко, особенно количество антител IgA , присутствующих в материнском молоке. У взрослых мышей RORγt+ Treg и IgA ингибируют друг друга. Аналогичным образом, у мышей, которых кормят приемные матери с более высокими титрами IgA в молоке, будет развиваться меньше RORγt+ Treg по сравнению с мышами, которых кормят молоком с более низкими титрами IgA. [53]
Также было показано, что RORγt+ Treg важны для пероральной толерантности и предотвращения пищевой аллергии. Младенцы с развитой пищевой аллергией имеют другой состав фекальной микробиоты по сравнению со здоровыми детьми, имеют повышенный уровень IgE, связанного с фекальной микробиотой, и пониженный уровень секреторного IgA. У мышей защита от пищевой аллергии вызывалась введением видов Clostridiales и Bacteroidales . При их введении происходит расширение кишечных клеток RORγt+ Treg в пользу GATA3+ Treg, что обеспечивает защиту от аллергии. [54]
Дефицит триптофана , незаменимой аминокислоты, изменяет метаболизм комменсальной микробиоты, что приводит к увеличению количества RORγt+ Treg-клеток и уменьшению количества Gata3+ Treg-клеток. Эта индукция, возможно, регулируется стимуляцией арильного углеводородного рецептора метаболитами, продуцируемыми комменсальными бактериями, использующими триптофан в качестве источника энергии. [55]
Меньшее количество клеток RORγt+ Treg присутствует у мышей без микробов, колонизированных микробиотой, связанной с воспалительным заболеванием кишечника, по сравнению с мышами без микробов, колонизированными здоровой микробиотой. Нарушение регуляции клеток RORγt+ Treg способствует увеличению количества клеток Th2 , а меньшее количество клеток RORγt+ Treg компенсируется увеличением количества клеток Helios+ Treg. Как именно RORγt+ Treg могут защитить от колита, пока неизвестно. [56]
Регуляторные Т-лимфоциты RORγt+ при раке
[ редактировать ]Патологическим может быть вовлечение регуляторных Т-клеток RORγt+ при колоректальном раке. Было обнаружено, что RORγt+ Treg, которые способны экспрессировать IL-17, увеличиваются при колоректальном раке и по мере развития рака теряют способность экспрессировать противовоспалительный IL-10. Аналогичным образом такие RORγt+ Treg, экспрессирующие IL-17, размножаются в слизистой оболочке пациентов с болезнью Крона. [57] [58] Истощение RORγt+ Treg у мышей с колоректальным раком вызывало усиление реактивности опухолеспецифических Т-клеток и улучшало иммунный надзор за раком. Это улучшение не вызвано потерей IL-17, поскольку было доказано, что он способствует прогрессированию рака. [58] В опухолях мышей с условным нокаутом RORγt+ Treg было подтверждено снижение экспрессии IL-6 , снижение экспрессии IL-6 дендритными клетками CD11c+ и сверхэкспрессия CTLA-4 . IL-6 опосредует активацию транскрипционного фактора STAT3 , который имеет решающее значение для пролиферации раковых клеток. [59]
Gata3+ регуляторные Т-лимфоциты
[ редактировать ]Другой важной подгруппой Treg-клеток являются Gata3+ Treg-клетки, которые реагируют на IL-33 в кишечнике и влияют на регуляцию эффекторных Т-клеток во время воспаления. В отличие от клеток RORγt+ Treg, эти клетки экспрессируют Helios и не зависят от микробиома. [51] [60]
Gata3+ T-reg являются основными иммуносупрессорами при воспалении кишечника, а T-reg используют Gata3 для ограничения воспаления тканей. Эта популяция клеток также ограничивает иммунитет Т-клеток Th17 в кишечнике, поскольку T-reg с дефицитом Gata3 экспрессируют более высокие Rorc и IL-17a . транскрипты [61]
Болезнь
[ редактировать ]Важный вопрос заключается в том, как модулируется иммуносупрессивная активность регуляторных Т-клеток в ходе продолжающегося иммунного ответа. Хотя иммуносупрессивная функция регуляторных Т-клеток предотвращает развитие аутоиммунных заболеваний, она нежелательна при иммунных ответах на инфекционные микроорганизмы.
Инфекции
[ редактировать ]При встрече с инфекционными микроорганизмами активность регуляторных Т-клеток может подавляться прямо или косвенно другими клетками, чтобы облегчить элиминацию инфекции. Экспериментальные данные, полученные на мышиных моделях, позволяют предположить, что некоторые патогены, возможно, эволюционировали, чтобы манипулировать регуляторными Т-клетками с целью подавления иммунитета хозяина и, таким образом, потенцировать собственное выживание. Например, сообщалось, что активность регуляторных Т-клеток увеличивается при некоторых инфекционных контекстах, таких как ретровирусные инфекции (наиболее известная из которых – ВИЧ), микобактериальные инфекции (например, туберкулез [62] ), а также различные паразитарные инфекции, включая лейшманию и малярию .
T- reg- клетки играют важную роль во время ВИЧ- инфекции. Они подавляют иммунную систему, тем самым ограничивая клетки-мишени и уменьшая воспаление, но это одновременно нарушает клиренс вируса посредством клеточно-опосредованного иммунного ответа и увеличивает резервуар, стимулируя CD4. + Т-клетки в состояние покоя, включая инфицированные клетки. Кроме того, T- reg- клетки могут быть инфицированы ВИЧ, что напрямую увеличивает размер резервуара ВИЧ. Таким образом, T- reg -клетки исследуются в качестве мишени для исследований по лечению ВИЧ. [63] Некоторые стратегии истощения Т- рег- клеток были протестированы на ВИО, инфицированных приматах, кроме человека , и было показано, что они вызывают реактивацию вируса и усиление специфичного для ВИО CD8. + Т-клеточные ответы. [64]
Регуляторные Т-клетки играют большую роль в патологии висцерального лейшманиоза и в предотвращении избыточного воспаления у пациентов, излеченных от висцерального лейшманиоза.
ЕСЛИ
[ редактировать ]Есть некоторые доказательства того, что T- reg -клетки могут быть дисфункциональными и вызывать нейровоспаление при боковом амиотрофическом склерозе из-за более низкой экспрессии FOXP3. [65] -клеток ex vivo Экспансия T- reg для последующей аутологичной трансплантации в настоящее время исследуется после получения многообещающих результатов в I фазе клинических испытаний. [66]
Беременность
[ редактировать ]Хотя регуляторные Т-клетки увеличиваются за счет поликлональной экспансии как системно, так и локально во время здоровой беременности, чтобы защитить плод от материнского иммунного ответа (процесс, называемый материнской иммунной толерантностью), данные свидетельствуют о том, что эта поликлональная экспансия нарушена у матерей с преэклампсией и их потомства. [67] Исследования показывают, что снижение производства и развития регуляторных Т-клеток во время преэклампсии может ухудшить иммунную толерантность матери, что приводит к гиперактивному иммунному ответу, характерному для преэклампсии. [68]
Рак
[ редактировать ]
CD4 + регуляторные Т-клетки часто связаны с солидными опухолями как у людей, так и у мышей. Увеличение количества регуляторных Т-клеток при раке молочной железы, колоректального рака и рака яичников связано с худшим прогнозом. [69]
CD70 + В-клетки неходжкинской лимфомы индуцируют экспрессию FOXP3 и регуляторную функцию во внутриопухолевых CD4 + CD25 − Т-клетки. [70]
Большинство опухолей вызывают у хозяина иммунный ответ, опосредованный опухолевыми антигенами, что отличает опухоль от других нераковых клеток. Это приводит к появлению большого количества инфильтрирующих опухоль лимфоцитов (TIL) в ТМЭ. [71] Эти лимфоциты могут нацеливаться на раковые клетки и, следовательно, замедлять или прекращать развитие опухоли. Однако этот процесс сложен, поскольку Т- рег- клетки, по-видимому, преимущественно передаются в ТМЕ. Хотя Т- рег- клетки обычно составляют лишь около 4% CD4 + Т-клетки, они могут составлять до 20–30% CD4. + Население вокруг ТМЕ. [72]
Соотношение Т- рег и эффекторных Т-клеток в ТМЕ является определяющим фактором успеха противоракового иммунного ответа. Высокие уровни Т- рег- клеток в ТМЭ связаны с плохим прогнозом при многих видах рака. [73] такие как рак яичников, молочной железы, почек и поджелудочной железы. [72] Это указывает на то, что Т- рег -клетки подавляют эффекторные Т-клетки и препятствуют иммунному ответу организма против рака. Однако при некоторых типах рака верно обратное: высокие уровни Т- рег -клеток связаны с положительным прогнозом. Эта тенденция наблюдается при таких видах рака, как колоректальная карцинома и фолликулярная лимфома . Это может быть связано со способностью T- reg- клеток подавлять общее воспаление, которое, как известно, запускает пролиферацию клеток и метастазирование. [72] Эти противоположные эффекты указывают на то, что роль Т - клеток в развитии рака сильно зависит как от типа, так и от местоположения опухоли.
Хотя до сих пор не совсем понятно, как Т- рег -клетки преимущественно доставляются в ТМЕ, хемотаксис, вероятно, обусловлен продукцией хемокинов опухолью. Инфильтрации T- reg в TME способствует связывание хемокинового рецептора CCR4, который экспрессируется на клетках T- reg , с его лигандом CCL22, который секретируется многими типами опухолевых клеток. [74] Экспансия Т- рег- клеток в месте опухоли также может объяснить повышенный уровень Т- рег- клеток. Известно, что цитокин TGF-β, который обычно продуцируется опухолевыми клетками, индуцирует дифференцировку и экспансию T- reg- клеток. [74]
Белок 3 Forkhead Box ( FOXP3 ) как фактор транскрипции является важным молекулярным маркером T- reg -клеток. Полиморфизм FOXP3 (rs3761548) может быть вовлечен в прогрессирование рака желудка , влияя на функцию T reg и секрецию иммуномодулирующих цитокинов, таких как IL-10 , IL-35 и TGF-β . [75]
T- reg- клетки, присутствующие в TME, могут быть либо индуцированными T- reg , либо естественными (тимическими) T- reg , которые развиваются из наивных предшественников. Однако ассоциированные с опухолью T- reg могут также происходить из IL-17A. + Фоксp3 + T- reg , которые развиваются из клеток Th17. [76] [77]
В целом, иммуносупрессия ТМЭ во многом способствовала неудачным результатам многих методов иммунотерапии рака. Истощение Т- рег -клеток на животных моделях показало повышенную эффективность иммунотерапевтического лечения, и поэтому многие методы иммунотерапии в настоящее время включают истощение Т- рег . [2]
Терапия рака, нацеленная на регуляторные Т-лимфоциты
[ редактировать ]T- reg в TME в большом количестве являются эффекторными T -reg , которые сверхэкспрессируют иммуносупрессивные молекулы, такие как CTLA-4. Антитела против CTLA-4 вызывают истощение T- reg и, таким образом, увеличивают CD8. + Т-клетки эффективны против опухоли. Антитело против CTLA-4 ипилимумаб было одобрено для пациентов с поздней стадией меланомы. Молекула контрольной точки иммунитета PD-1 ингибирует активацию как обычных Т-клеток, так и T -reg , а использование антител против PD-1 может привести к активации и иммуносупрессивной функции T- reg . Устойчивость к лечению анти-PD-1-mAb, вероятно, вызвана повышенной активностью Т- reg -клеток. Быстрое прогрессирование рака при блокаде PD-1 называется гиперпрогрессирующим заболеванием. Терапия, направленная на подавление T- reg, включает моноклональные антитела против CD25 и моноклональные антитела против CCR4. Агонист OX40 и агонисты GITR в настоящее время исследуются. [76] [78] Терапия, направленная на передачу сигналов TCR, также возможна путем блокирования тирозинкиназ. Например, ингибитор тирозинкиназы дазатиниб используется для лечения хронического миелолейкоза и связан с ингибированием T- reg . [79]
Молекулярная характеристика
[ редактировать ]Подобно другим Т-клеткам, в тимусе развиваются регуляторные Т-клетки . Последние исследования показывают, что регуляторные Т-клетки определяются экспрессией фактора транскрипции семейства forkhead FOXP3 (forkhead box p3). Экспрессия FOXP3 необходима для развития регуляторных Т-клеток и, по-видимому, контролирует генетическую программу, определяющую судьбу этой клетки. [80] Подавляющее большинство регуляторных Т-клеток, экспрессирующих Foxp3, обнаружено в главном комплексе гистосовместимости (MHC) класса II, ограниченном экспрессией CD 4 (CD4 + ) популяции и экспрессируют высокие уровни альфа-цепи рецептора интерлейкина-2 (CD25). В дополнение к FOXP3-экспрессирующему CD4 + CD25 + , также, по-видимому, существует незначительная популяция MHC класса I с ограниченным CD8. + Регуляторные Т-клетки, экспрессирующие FOXP3. Эти FOXP3-экспрессирующие CD8 + Т-клетки, по-видимому, не функционируют у здоровых людей, но при аутоиммунных заболеваниях они индуцируются стимуляцией рецепторов Т-клеток для подавления иммунных ответов, опосредованных IL-17. [81] В отличие от обычных Т-клеток, регуляторные Т-клетки не продуцируют IL-2 и поэтому являются анергическими на исходном уровне.
клеток используется ряд различных методов В исследованиях для идентификации и мониторинга Т- рег- . Первоначально использовалась высокая экспрессия поверхностных маркеров CD25 и CD4 (CD4 + CD25 + клетки). Это проблематично, поскольку CD25 также экспрессируется на нерегуляторных Т-клетках в условиях иммунной активации, например, во время иммунного ответа на патоген. Согласно экспрессии CD4 и CD25, регуляторные Т-клетки составляют около 5–10% зрелых CD4. + около 1–2% Т- рег Субпопуляция Т-клеток у мышей и людей, тогда как в цельной крови можно измерить . Дополнительное измерение клеточной экспрессии белка FOXP3 позволило провести более специфичный анализ Т- рег- клеток (CD4 + CD25 + ФОКС3 + клетки). Однако FOXP3 также временно экспрессируется в активированных эффекторных Т-клетках человека, что усложняет правильный анализ T reg с использованием CD4, CD25 и FOXP3 в качестве маркеров у людей. Таким образом, комбинация поверхностных маркеров золотого стандарта с определенными T- reg в неактивированном CD3 + CD4 + Т-клетки характеризуются высокой экспрессией CD25 в сочетании с отсутствием или низким уровнем экспрессии поверхностного белка CD127 (IL-7RA). Если жизнеспособные клетки не требуются, то добавление FOXP3 к комбинации CD25 и CD127 обеспечит дополнительную строгость. Описано несколько дополнительных маркеров, например, высокие уровни CTLA-4 (цитотоксичная молекула-4, связанная с Т-лимфоцитами) и GITR (глюкокортикоид-индуцированный рецептор TNF) также экспрессируются на регуляторных Т-клетках, однако функциональное значение этой экспрессии остается. быть определены. Существует большой интерес к выявлению маркеров клеточной поверхности, которые уникально и специфично экспрессируются на всех регуляторных Т-клетках, экспрессирующих FOXP3. Однако до настоящего времени такая молекула не была идентифицирована.
Идентификация T- reg после активации клеток является сложной задачей, поскольку обычные Т-клетки будут экспрессировать CD25, временно экспрессировать FOXP3 и терять экспрессию CD127 после активации. Было показано, что T reg можно обнаружить с помощью анализа маркеров, индуцированных активацией, по экспрессии CD39. [82] в сочетании с совместной экспрессией CD25 и OX40 (CD134), которые определяют антигенспецифические клетки после 24-48-часовой стимуляции антигеном. [83] [84]
Помимо поиска новых белковых маркеров, анализа и мониторинга Т- рег- в литературе описан другой метод более точного клеток. Этот метод основан на анализе метилирования ДНК . Только в клетках T reg , но не в клетках любого другого типа, включая активированные эффекторные T-клетки, определенная область гена FOXP3 (TSDR, T reg -специфическая деметилированная область) обнаруживается деметилированной, что позволяет контролировать T reg клетки посредством реакция ПЦР или другие методы анализа на основе ДНК. [85] Взаимодействие между клетками Th17 и регуляторными Т-клетками важно при многих заболеваниях, таких как респираторные заболевания. [86]
Недавние данные свидетельствуют о том, что тучные клетки могут быть важными медиаторами Т- рег -зависимой периферической толерантности. [87]
Эпитопы
[ редактировать ]Регуляторные эпитопы Т-клеток («Трегитопы») были открыты в 2008 году и состоят из линейных последовательностей аминокислот, содержащихся в моноклональных антителах и иммуноглобулине G (IgG). С момента их открытия данные показали, что трегитопы могут иметь решающее значение для активации естественных регуляторных Т-клеток. [88] [89] [90]
Были выдвинуты гипотезы о потенциальных применениях регуляторных эпитопов Т-клеток: толерантность к трансплантатам, белковым препаратам, терапии с помощью переливания крови и диабету I типа , а также снижение иммунного ответа для лечения аллергии . [91] [92] [93] [94] [95] [96] [90]
Генетический дефицит
[ редактировать ]Генетические мутации в гене, кодирующем FOXP3, были выявлены как у людей, так и у мышей на основании наследственного заболевания, вызванного этими мутациями. Это заболевание представляет собой наиболее яркое доказательство того, что регуляторные Т-клетки играют решающую роль в поддержании нормальной функции иммунной системы. У людей с мутациями FOXP3 развивается тяжелое и быстро фатальное аутоиммунное заболевание, известное как дисрегуляция , полиэндокринопатия , -сцепленной энтеропатии иммунная синдром Х ( IPEX ). [97] [98]
Синдром IPEX характеризуется развитием подавляющего системного аутоиммунитета на первом году жизни, что приводит к часто наблюдаемой триаде: водянистой диарее, экзематозному дерматиту и эндокринопатии, чаще всего наблюдаемой как инсулинозависимый сахарный диабет . У большинства людей наблюдаются другие аутоиммунные явления, включая Кумбс-положительную гемолитическую анемию, аутоиммунную тромбоцитопению, аутоиммунную нейтропению и тубулярную нефропатию. Большинство больных мужчин умирают в течение первого года жизни либо от метаболических нарушений, либо от сепсиса. Аналогичное заболевание также наблюдается у спонтанных мышей с мутантом FOXP3, известных как «назальные».
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Беттелли Э., Кэрриер Ю., Гао В., Корн Т., Стром Т.Б., Оукка М. и др. (май 2006 г.). «Реципрокные пути развития для генерации патогенных эффекторных TH17 и регуляторных Т-клеток». Природа . 441 (7090): 235–238. Бибкод : 2006Natur.441..235B . дои : 10.1038/nature04753 . ПМИД 16648838 . S2CID 4391497 .
- ^ Перейти обратно: а б Куриэль Т.Дж. (май 2007 г.). «Трегс и переосмысление иммунотерапии рака» . Журнал клинических исследований . 117 (5): 1167–1174. дои : 10.1172/JCI31202 . ПМК 1857250 . ПМИД 17476346 .
- ^ Чен В (август 2011 г.). «Трегс в иммунотерапии: возможности и проблемы» . Иммунотерапия . 3 (8): 911–914. дои : 10.2217/imt.11.79 . ПМИД 21843075 .
- ^ Мияра М, Горохов Г, Эренштейн М, Мюссе Л, Сакагути С, Амура З (октябрь 2011 г.). «Регуляторные Т-клетки FoxP3+ человека при системных аутоиммунных заболеваниях». Обзоры аутоиммунитета . 10 (12): 744–755. дои : 10.1016/j.autrev.2011.05.004 . ПМИД 21621000 .
- ^ Носбаум А., Превель Н., Труонг Х.А., Мехта П., Эттингер М., Шаршмидт Т.С. и др. (март 2016 г.). «Авангард: регуляторные Т-клетки способствуют заживлению кожных ран» . Журнал иммунологии . 196 (5): 2010–2014. doi : 10.4049/jimmunol.1502139 . ПМЦ 4761457 . ПМИД 26826250 .
- ^ Адигбе Д.О., Нисикава Х. (2013). «Естественные и индуцированные Т-регуляторные клетки при раке» . Границы в иммунологии . 4 : 190. дои : 10.3389/fimmu.2013.00190 . ПМК 3708155 . ПМИД 23874336 .
- ^ Куриэль Т.Дж. (апрель 2008 г.). «Регуляторные Т-клетки и лечение рака» . Современное мнение в иммунологии . 20 (2): 241–246. дои : 10.1016/j.coi.2008.04.008 . ПМК 3319305 . ПМИД 18508251 .
- ^ Хори С., Номура Т., Сакагути С. (февраль 2003 г.). «Контроль развития регуляторных Т-клеток с помощью транскрипционного фактора Foxp3». Наука . 299 (5609): 1057–1061. Бибкод : 2003Sci...299.1057H . дои : 10.1126/science.1079490 . ПМИД 12522256 . S2CID 9697928 .
- ^ Сингх Б., Шварц Дж.А., Сэндрок С., Беллемор С.М., Никопур Э. (ноябрь 2013 г.). «Модуляция аутоиммунных заболеваний интерлейкином (IL)-17, продуцирующим регуляторные Т-хелперные клетки (Th17)» . Индийский журнал медицинских исследований . 138 (5): 591–594. ПМЦ 3928692 . ПМИД 24434314 .
- ^ Шевач Э.М. (2000). «Регуляторные Т-клетки при аутоиммунитете*» . Ежегодный обзор иммунологии . 18 : 423–449. doi : 10.1146/annurev.immunol.18.1.423 . ПМИД 10837065 . S2CID 15160752 .
- ^ Шметтерер К.Г., Нойнкирхнер А., Пикль В.Ф. (июнь 2012 г.). «Естественные регуляторные Т-клетки: маркеры, механизмы и манипуляции» . Журнал ФАСЭБ . 26 (6): 2253–2276. дои : 10.1096/fj.11-193672 . ПМИД 22362896 . S2CID 36277557 .
- ^ Сакагути С (2004). «Естественно возникающие Т-клетки, регуляторные CD4+, для иммунологической аутотолерантности и негативного контроля иммунных ответов». Ежегодный обзор иммунологии . 22 : 531–562. doi : 10.1146/annurev.immunol.21.120601.141122 . ПМИД 15032588 .
- ^ Ли МО, Руденский А.Ю. (апрель 2016). «Передача сигналов Т-клеточных рецепторов в контроле дифференцировки и функции регуляторных Т-клеток» . Обзоры природы. Иммунология . 16 (4): 220–233. дои : 10.1038/nri.2016.26 . ПМЦ 4968889 . ПМИД 27026074 .
- ^ Перейти обратно: а б с д Сантамария Х.К., Борелли А., Ирла М. (11 февраля 2021 г.). «Регуляторная гетерогенность Т-клеток в тимусе: влияние на их функциональную активность» . Границы в иммунологии . 12 : 643153. дои : 10.3389/fimmu.2021.643153 . ПМЦ 7904894 . ПМИД 33643324 .
- ^ Оуэн Д.Л., Сьяастад Л.Е., Фаррар М.А. (октябрь 2019 г.). «Регуляторное развитие Т-клеток в тимусе» . Журнал иммунологии . 203 (8): 2031–2041. дои : 10.4049/gymmunol.1900662 . ПМК 6910132 . ПМИД 31591259 .
- ^ Оуэн Д.Л., Махмуд С.А., Сьяастад Л.Е., Уильямс Дж.Б., Спэньер Дж.А., Симеонов Д.Р. и др. (февраль 2019 г.). «Регуляторные Т-клетки тимуса возникают посредством двух различных программ развития» . Природная иммунология . 20 (2): 195–205. дои : 10.1038/s41590-018-0289-6 . ПМК 6650268 . ПМИД 30643267 .
- ^ Перейти обратно: а б с д Тио Н., Дарриг Ж., Аду В., Грос М., Бине Б., Перальс С. и др. (июнь 2015 г.). «Периферические регуляторные Т-лимфоциты, рециркулирующие в тимус, подавляют развитие своих предшественников». Природная иммунология . 16 (6): 628–634. дои : 10.1038/ni.3150 . ПМИД 25939024 . S2CID 7670443 .
- ^ Перейти обратно: а б Пандиян П., Чжэн Л., Исихара С., Рид Дж., Ленардо М.Дж. (декабрь 2007 г.). «Регуляторные Т-клетки CD4 + CD25 + Foxp3 + индуцируют опосредованный депривацией цитокинов апоптоз эффекторных CD4 + Т-клеток». Природная иммунология . 8 (12): 1353–1362. дои : 10.1038/ni1536 . ПМИД 17982458 . S2CID 8925488 .
- ^ Ченг Г., Ю А, Малек Т.Р. (май 2011 г.). «Толерантность Т-клеток и многофункциональная роль передачи сигналов IL-2R в Т-регуляторных клетках» . Иммунологические обзоры . 241 (1): 63–76. дои : 10.1111/j.1600-065X.2011.01004.x . ПМК 3101713 . ПМИД 21488890 .
- ^ Киммиг С., Пшибильски Г.К., Шмидт К.А., Лауриш К., Мёвес Б., Радбрух А., Тиль А. (март 2002 г.). «Две подгруппы наивных Т-хелперных клеток с различным содержанием круга вырезания Т-клеточных рецепторов в периферической крови взрослого человека» . Журнал экспериментальной медицины . 195 (6): 789–794. дои : 10.1084/jem.20011756 . ПМК 2193736 . ПМИД 11901204 .
- ^ Токер А., Энгельберт Д., Гарг Г., Полански Дж.К., Флосс С., Мияо Т. и др. (апрель 2013 г.). «Активное деметилирование локуса Foxp3 приводит к образованию стабильных регуляторных Т-клеток в тимусе» . Журнал иммунологии . 190 (7): 3180–3188. doi : 10.4049/jimmunol.1203473 . ПМИД 23420886 .
- ^ Перейти обратно: а б с д Николоули Е., Эльфаки Ю., Херппих С., Шельмбауэр С., Делачер М., Фальк С. и др. (январь 2021 г.). «Рециркуляционный ИЛ-1Р2 + тонкая настройка внутритимная разработка Treg в воспалительных условиях » и иммунология . 18 1) DOI : 10.1038 . 182–193 . Трегс : / S41423-019-0352-8 . ( молекулярная Клеточная S2CID 210913733 .
- ^ Петерс В.А., Йостинг Дж.Дж., Фройнд Г.Г. (август 2013 г.). «Рецептор 2 IL-1 (IL-1R2) и его роль в иммунной регуляции» . Мозг, поведение и иммунитет . 32 : 1–8. дои : 10.1016/j.bbi.2012.11.006 . ПМЦ 3610842 . ПМИД 23195532 .
- ^ Прочтите С., Мальмстрем В., Паури Ф. (июль 2000 г.). «Антиген 4, ассоциированный с цитотоксическим Т-лимфоцитом, играет важную роль в функции регуляторных клеток CD25(+)CD4(+), которые контролируют воспаление кишечника» . Журнал экспериментальной медицины . 192 (2): 295–302. дои : 10.1084/jem.192.2.295 . ПМК 2193261 . ПМИД 10899916 .
- ^ Коллисон Л.В., Уоркман С.Дж., Куо Т.Т., Бойд К., Ван Ю., Виньяли К.М. и др. (ноябрь 2007 г.). «Ингибирующий цитокин IL-35 способствует регуляторной функции Т-клеток». Природа . 450 (7169): 566–569. Бибкод : 2007Natur.450..566C . дои : 10.1038/nature06306 . ПМИД 18033300 . S2CID 4425281 .
- ^ Аннакер О., Ассеман С., Рид С., Паури Ф. (июнь 2003 г.). «Интерлейкин-10 в регуляции колита, индуцированного Т-клетками». Журнал аутоиммунитета . 20 (4): 277–279. дои : 10.1016/s0896-8411(03)00045-3 . ПМИД 12791312 .
- ^ Кирли Дж., Баркер Дж.Э., Робинсон Д.С., Ллойд К.М. (декабрь 2005 г.). «Разрешение воспаления дыхательных путей и гиперреактивности после переноса in vivo регуляторных Т-клеток CD4+CD25+ зависит от интерлейкина 10» . Журнал экспериментальной медицины . 202 (11): 1539–1547. дои : 10.1084/jem.20051166 . ПМК 1350743 . ПМИД 16314435 .
- ^ Гондек, округ Колумбия, Лу Л.Ф., Кесада С.А., Сакагути С., Ноэль Р.Дж. (февраль 2005 г.). «Авангард: контактно-опосредованное подавление регуляторными клетками CD4+CD25+ включает гранзим B-зависимый и независимый от перфорина механизм» . Журнал иммунологии . 174 (4): 1783–1786. doi : 10.4049/jimmunol.174.4.1783 . ПМИД 15699103 .
- ^ Пуччетти П., Громанн Ю (октябрь 2007 г.). «IDO и регуляторные Т-клетки: роль в обратной передаче сигналов и неканонической активации NF-kappaB». Обзоры природы. Иммунология . 7 (10): 817–823. дои : 10.1038/nri2163 . ПМИД 17767193 . S2CID 5544429 .
- ^ Борселлино Г., Кляйневитфельд М., Ди Митри Д., Стерньяк А., Диамантини А., Джометто Р. и др. (август 2007 г.). «Экспрессия эктонуклеотидазы CD39 клетками Foxp3+ Treg: гидролиз внеклеточного АТФ и иммуносупрессия» . Кровь . 110 (4): 1225–1232. doi : 10.1182/blood-2006-12-064527 . ПМИД 17449799 .
- ^ Коби Дж.Дж., Шах П.Р., Ян Л., Ребхан Дж.А., Фауэлл DJ, Мосманн Т.Р. (ноябрь 2006 г.). «Т-регуляторные и праймированные некоммитированные Т-клетки CD4 экспрессируют CD73, который подавляет эффекторные Т-клетки CD4 путем преобразования 5'-аденозинмонофосфата в аденозин» . Журнал иммунологии . 177 (10): 6780–6786. дои : 10.4049/jimmunol.177.10.6780 . ПМИД 17082591 .
- ^ Хуан К.Т., Уоркман С.Дж., Флис Д., Пан Х., Марсон А.Л., Чжоу Г. и др. (октябрь 2004 г.). «Роль LAG-3 в регуляторных Т-клетках» . Иммунитет . 21 (4): 503–513. doi : 10.1016/j.immuni.2004.08.010 . ПМИД 15485628 .
- ^ Ю Х, Харден К., Гонсалес Л.С., Франческо М., Чианг Э., Ирвинг Б. и др. (январь 2009 г.). «Поверхностный белок TIGIT подавляет активацию Т-клеток, способствуя образованию зрелых иммунорегуляторных дендритных клеток». Природная иммунология . 10 (1): 48–57. дои : 10.1038/ni.1674 . ПМИД 19011627 . S2CID 205361984 .
- ^ Уорделл К.М., Макдональд К.Н., Левингс М.К., Кук Л. (январь 2021 г.). «Перекрестный разговор между регуляторными Т-клетками человека и антигенпрезентирующими клетками: уроки для клинического применения» . Европейский журнал иммунологии . 51 (1): 27–38. дои : 10.1002/eji.202048746 . hdl : 11343/276776 . ПМИД 33301176 .
- ^ Сакагути С., Ямагути Т., Номура Т., Оно М. (май 2008 г.). «Регуляторные Т-клетки и иммунная толерантность» . Клетка . 133 (5): 775–787. дои : 10.1016/j.cell.2008.05.009 . ПМИД 18510923 .
- ^ Уокер Л.С., Сансом DM (ноябрь 2011 г.). «Новая роль CTLA4 как внешнего регулятора Т-клеточных ответов». Обзоры природы. Иммунология . 11 (12): 852–863. дои : 10.1038/nri3108 . ПМИД 22116087 . S2CID 9617595 .
- ^ Перейти обратно: а б с д Шевырев Д, Терещенко В (2020). «Трег-гетерогенность, функция и гомеостаз» . Границы в иммунологии . 10 : 3100. дои : 10.3389/fimmu.2019.03100 . ПМК 6971100 . ПМИД 31993063 .
- ^ Харибхай Д., Уильямс Дж.Б., Цзя С., Никерсон Д., Шмитт Э.Г., Эдвардс Б. и др. (июль 2011 г.). «Необходимая роль индуцированных регуляторных Т-клеток в толерантности, основанная на расширении разнообразия антигенных рецепторов» . Иммунитет . 35 (1): 109–122. doi : 10.1016/j.immuni.2011.03.029 . ПМЦ 3295638 . ПМИД 21723159 .
- ^ Перейти обратно: а б Сан СМ, Холл Дж.А., Бланк Р.Б., Буладу Н., Оукка М., Мора Дж.Р., Белкаид Ю. (август 2007 г.). «Дендритные клетки собственной пластинки тонкой кишки способствуют образованию de novo T-reg-клеток Foxp3 посредством ретиноевой кислоты» . Журнал экспериментальной медицины . 204 (8): 1775–1785. дои : 10.1084/jem.20070602 . ПМК 2118682 . ПМИД 17620362 .
- ^ Муцида Д., Пак Ю., Ким Г., Туровская О., Скотт И., Кроненберг М. , Шерутр Х. (июль 2007 г.). «Взаимная дифференцировка TH17 и регуляторных Т-клеток, опосредованная ретиноевой кислотой» . Наука . 317 (5835): 256–260. Бибкод : 2007Sci...317..256M . дои : 10.1126/science.1145697 . ПМИД 17569825 . S2CID 24736012 .
- ^ Эркеленс М.Н., Мебиус Р.Е. (март 2017 г.). «Ретиноевая кислота и иммунный гомеостаз: баланс». Тенденции в иммунологии . 38 (3): 168–180. дои : 10.1016/j.it.2016.12.006 . ПМИД 28094101 .
- ^ Зиглер С.Ф., Бакнер Дж.Х. (апрель 2009 г.). «FOXP3 и регуляция дифференцировки Treg/Th17» . Микробы и инфекции . 11 (5): 594–598. дои : 10.1016/j.micinf.2009.04.002 . ПМЦ 2728495 . ПМИД 19371792 .
- ^ Поволери Г.А., Нова-Ламперти Е, Скотта С, Фанелли Г, Чен ЮК, Беккер П.Д. и др. (декабрь 2018 г.). «CD161, регулируемый ретиноевой кислотой человека. + регуляторные Т-клетки поддерживают заживление ран слизистой оболочки кишечника» . Nature Immunology . 19 (12): 1403–1414. : 10.1038 /s41590-018-0230-z . PMC 6474659. . PMID 30397350 doi
- ^ Кумбс Дж.Л., Сиддики К.Р., Арансибия-Каркамо К.В., Холл Дж., Сан К.М., Белкаид Ю., Паури Ф. (август 2007 г.). «Функционально специализированная популяция CD103+ DC слизистой оболочки индуцирует регуляторные Т-клетки Foxp3+ посредством механизма, зависимого от TGF-бета и ретиноевой кислоты» . Журнал экспериментальной медицины . 204 (8): 1757–1764. дои : 10.1084/jem.20070590 . ПМК 2118683 . ПМИД 17620361 .
- ^ Перейти обратно: а б Кук Л., Рид К.Т., Хаккинен Э., де Би Б., Танака С., Смит DJ и др. (сентябрь 2021 г.). «Индукция стабильного человеческого FOXP3 + Treg с помощью миметика TGF-β, полученного из паразита» . Immunology and Cell Biology . 99 (8): 833–847. : 10.1111 /IMCB.12475 . PMC 8453874. . PMID 33929751 doi
- ^ Джонстон С.Дж., Смит Дж., Кодали Р.Б., Уайт MP, Харкус Ю., Филби К.Дж. и др. (ноябрь 2017 г.). «Структурно отличный от TGF-β имитатор кишечного гельминта эффективно индуцирует регуляторные Т-клетки» . Природные коммуникации . 8 (1): 1741. Бибкод : 2017NatCo...8.1741J . дои : 10.1038/s41467-017-01886-6 . ПМК 5701006 . ПМИД 29170498 .
- ^ Смит Д.Д., Харкус Ю., Уайт MP, Грегори В.Ф., Налер Дж., Стивенс И. и др. (апрель 2018 г.). «Белки, мимические TGF-β, образуют расширенное семейство генов мышиного паразита Heligmosomoides polygyrus» . Международный журнал паразитологии . 48 (5): 379–385. дои : 10.1016/j.ijpara.2017.12.004 . ПМЦ 5904571 . ПМИД 29510118 .
- ^ Уайт MP, Смит DJ, Кук Л., Зиглер С.Ф., Левингс МК, Майзелс Р.М. (сентябрь 2021 г.). «Паразитный цитокин, имитирующий Hp-TGM, мощно воспроизводит регуляторные эффекты TGF-β на мышиный CD4. + Т-клетки» . Иммунология и клеточная биология . 99 (8): 848–864. : 10.1111 /IMCB.12479 . PMC 9214624. . PMID 33988885 doi
- ^ Онмахт С., Парк Дж.Х., Кординг С., Винг Дж.Б., Атараши К., Обата Ю. и др. (август 2015 г.). «ИММУНОЛОГИЯ слизистых оболочек. Микробиота регулирует иммунитет 2 типа через Т-клетки RORγt⁺». Наука . 349 (6251): 989–993. Бибкод : 2015Sci...349..989O . дои : 10.1126/science.aac4263 . ПМИД 26160380 . S2CID 2663636 .
- ^ Перейти обратно: а б с Нин X, Лей З, Руй Б, Ли Ю, Ли М (05 декабря 2022 г.). «Микробиота кишечника способствует иммунной толерантности путем регулирования RORγt+ Treg-клеток при пищевой аллергии» . Расширенные исследования кишечника и микробиома . 2022 : e8529578. дои : 10.1155/2022/8529578 .
- ^ Перейти обратно: а б Онмахт С., Парк Дж.Х., Кординг С., Винг Дж.Б., Атараши К., Обата Ю. и др. (август 2015 г.). «ИММУНОЛОГИЯ слизистых оболочек. Микробиота регулирует иммунитет 2 типа через Т-клетки RORγt⁺». Наука . 349 (6251): 989–993. Бибкод : 2015Sci...349..989O . дои : 10.1126/science.aac4263 . ПМИД 26160380 . S2CID 2663636 .
- ^ Аль Набхани З., Дюлаурой С., Маркес Р., Кусу С., Аль Бонни С., Дежарден Ф. и др. (май 2019 г.). «Для устойчивости к иммунопатологиям у взрослых необходимо снижение реакции на микробиоту». Иммунитет . 50 (5): 1276–1288.e5. doi : 10.1016/j.immuni.2019.02.014 . ПМИД 30902637 .
- ^ Раманан Д., Сефик Э., Гальван-Пенья С., Ву М., Ян Л., Ян З. и др. (июнь 2020 г.). «Иммунологический способ передачи от нескольких поколений регулирует уставку кишечного Treg» . Клетка . 181 (6): 1276–1290.e13. дои : 10.1016/j.cell.2020.04.030 . ПМЦ 7393667 . ПМИД 32402238 .
- ^ Абдель-Гадир А., Стивен-Виктор Е., Гербер Г.К., Новаль Ривас М., Ван С., Харб Х. и др. (июль 2019 г.). «Микробиотная терапия действует через регуляторный путь Т-клеток MyD88/RORγt, подавляя пищевую аллергию» . Природная медицина . 25 (7): 1164–1174. дои : 10.1038/s41591-019-0461-z . ПМК 6677395 . ПМИД 31235962 .
- ^ Рэнкин Л.К., Кайзер К.А., де Лос Сантос-Алексис К., Парк Х., Улеманн А.С., Грей Д.Х., Арпая Н. (март 2023 г.). «Дефицит триптофана в пище способствует повышению RORγt в кишечнике. + Treg-клетки за счет Gata3 + Treg-клетки и изменяет метаболизм комменсальной микробиоты» . Cell Reports . 42 (3): 112135. doi : 10.1016/ . PMC 10150404. . PMID 36840944 j.celrep.2023.112135
- ^ Бриттон Г.Дж., Контиджох Э.Дж., Могно И., Веннаро О.Г., Ллевелин С.Р., Нг Р. и др. (январь 2019 г.). «Микробиота людей с воспалительным заболеванием кишечника изменяет баланс Th17 и RORγt в кишечнике» + Регуляторные Т-клетки и обострение колита у мышей» . Иммунитет . 50 (1): 212–224.e4. : 10.1016 /j.immuni.2018.12.015 . PMC 6512335. . PMID 30650377 doi
- ^ Оганесян З., Тритман Дж., Литтман Д.Р., Майер Л. (март 2011 г.). «Характеристика регуляторных Т-клеток, продуцирующих интерлейкин-17, в воспаленной слизистой оболочке кишечника у пациентов с воспалительными заболеваниями кишечника» . Гастроэнтерология . 140 (3): 957–965. дои : 10.1053/j.gastro.2010.12.002 . ПМК 3049831 . ПМИД 21147109 .
- ^ Перейти обратно: а б Блатнер Н.Р., Малкахи М.Ф., Деннис К.Л., Шолтенс Д., Бентрем Дж.Д., Филлипс Дж.Д. и др. (декабрь 2012 г.). «Экспрессия RORγt отмечает подмножество патогенных регуляторных Т-клеток при раке толстой кишки человека» . Наука трансляционной медицины . 4 (164): 164ра159. doi : 10.1126/scitranslmed.3004566 . ПМЦ 3762575 . ПМИД 23241743 .
- ^ Риццо А., Ди Джованджулио М., Столфи С., Франце Э., Фелинг Х.Дж., Карсетти Р. и др. (сентябрь 2018 г.). «RORγt-экспрессирующие Treg стимулируют рост колоректального рака, связанного с колитом, путем контроля IL6 в дендритных клетках». Исследования в области иммунологии рака . 6 (9): 1082–1092. doi : 10.1158/2326-6066.CIR-17-0698 . ПМИД 29991500 .
- ^ Якобс Дж., Ли Дж., Рингс Э.Х., Самсом Дж.Н., Геттель Дж.А. (2021). «Кишечные регуляторные Т-клетки как специализированные тканеограниченные иммунные клетки в кишечном иммунном гомеостазе и заболеваниях» . Границы в иммунологии . 12 : 716499. дои : 10.3389/fimmu.2021.716499 . ПМК 8371910 . ПМИД 34421921 .
- ^ Луи П.П., Чо И, Али Н. (сентябрь 2020 г.). «Тканевые регуляторные Т-клетки» . Иммунология . 161 (1): 4–17. дои : 10.1111/imm.13208 . ПМК 7450170 . ПМИД 32463116 .
- ^ Стрингари Л.Л., Ковре Л.П., да Силва Ф.Д., де Оливейра В.Л., Кампана М.С., Хадад DJ и др. (июль 2021 г.). «Повышение количества клеток CD4+CD25highFoxP3+ ухудшает микробицидную активность человека in vitro против микобактерий туберкулеза при латентном и остром туберкулезе легких» . PLOS Забытые тропические болезни . 15 (7): e0009605. дои : 10.1371/journal.pntd.0009605 . ПМЦ 8321116 . ПМИД 34324509 .
- ^ Кляйнман А.Дж., Сиванандхам Р., Пандреа И., Шугнет К.А., Апетрей С. (2018). «Регуляторные Т-клетки как потенциальные мишени для исследований по лечению ВИЧ» . Границы в иммунологии . 9 : 734. дои : 10.3389/fimmu.2018.00734 . ПМЦ 5908895 . ПМИД 29706961 .
- ^ Сиванандхам Р., Кляйнман А.Дж., Сетте П., Брокка-Кофано Э., Килапандал Венкатраман С.М., Поличиччио Б.Б. и др. (сентябрь 2020 г.). «Тестирование на приматах, не являющихся людьми, влияния различных стратегий истощения регуляторных Т-клеток на реактивацию и выведение латентного вируса иммунодефицита обезьян» . Журнал вирусологии . 94 (19): JVI.00533–20, jvi, JVI.00533–20в1. дои : 10.1128/JVI.00533-20 . ПМЦ 7495362 . ПМИД 32669326 . S2CID 220579402 .
- ^ Бирс Д.Р., Чжао В., Ван Дж., Чжан Х., Вэнь С., Нил Д. и др. (март 2017 г.). «Регуляторные Т-лимфоциты пациентов с БАС дисфункциональны и коррелируют со скоростью и тяжестью прогрессирования заболевания» . JCI-инсайт . 2 (5): е89530. doi : 10.1172/jci.insight.89530 . ПМЦ 5333967 . ПМИД 28289705 .
- ^ Тонхофф Дж.Р., Бирс Д.Р., Чжао В., Плейтес М., Симпсон Э.П., Берри Дж.Д. и др. (июль 2018 г.). «Расширенное введение аутологичных регуляторных Т-лимфоцитов при БАС: фаза I, первое исследование на людях» . Неврология . 5 (4): е465. дои : 10.1212/NXI.0000000000000465 . ПМЦ 5961523 . ПМИД 29845093 .
- ^ Цуда С., Накашима А., Сима Т., Сайто С. (2019). «Новая парадигма роли регуляторных Т-клеток во время беременности» . Границы в иммунологии . 10 :573. дои : 10.3389/fimmu.2019.00573 . ПМК 6443934 . ПМИД 30972068 .
- ^ Ху М., Эвистон Д., Сюй П., Мариньо Э., Чиджи А., Сантнер-Нанан Б. и др. (июль 2019 г.). «Снижение уровня ацетата в сыворотке матери и нарушение развития тимуса плода и регуляторных Т-клеток при преэклампсии» . Природные коммуникации . 10 (1): 3031. Бибкод : 2019NatCo..10.3031H . дои : 10.1038/s41467-019-10703-1 . ПМК 6620275 . ПМИД 31292453 .
- ^ Дранофф Г. (декабрь 2005 г.). «Терапевтическое значение внутриопухолевых регуляторных Т-клеток». Клинические исследования рака . 11 (23): 8226–8229. doi : 10.1158/1078-0432.CCR-05-2035 . ПМИД 16322278 . S2CID 18794337 .
- ^ Ян З.З., Новак А.Дж., Зисмер С.С., Витциг Т.Е., Анселл С.М. (октябрь 2007 г.). «CD70+ В-клетки неходжкинской лимфомы индуцируют экспрессию Foxp3 и регуляторную функцию во внутриопухолевых CD4+CD25 Т-клетках» . Кровь . 110 (7): 2537–2544. doi : 10.1182/blood-2007-03-082578 . ЧВК 1988926 . ПМИД 17615291 .
- ^ Гуден М.Дж., де Бок Г.Х., Лефферс Н., Дэмен Т., Найман Х.В. (июнь 2011 г.). «Прогностическое влияние инфильтрирующих опухоль лимфоцитов при раке: систематический обзор с метаанализом» . Британский журнал рака . 105 (1): 93–103. дои : 10.1038/bjc.2011.189 . ПМК 3137407 . ПМИД 21629244 .
- ^ Перейти обратно: а б с Олейника К., Ниббс Р.Дж., Грэм Г.Дж., Фрейзер А.Р. (январь 2013 г.). «Подавление, подрывная деятельность и побег: роль регуляторных Т-клеток в прогрессировании рака» . Клиническая и экспериментальная иммунология . 171 (1): 36–45. дои : 10.1111/j.1365-2249.2012.04657.x . ПМК 3530093 . ПМИД 23199321 .
- ^ Плитас Г., Руденский А.Ю. (09.03.2020). «Регуляторные Т-клетки при раке» . Ежегодный обзор биологии рака . 4 (1): 459–477. doi : 10.1146/annurev-cancerbio-030419-033428 . ISSN 2472-3428 .
- ^ Перейти обратно: а б Липпиц Б.Е. (май 2013 г.). «Цитокиновые закономерности у больных раком: систематический обзор». «Ланцет». Онкология . 14 (6): с218–е228. дои : 10.1016/s1470-2045(12)70582-x . ПМИД 23639322 .
- ^ Эззеддини Р., Соми М.Х., Тагихани М., Моаддаб С.Ю., Маснади Ширази К., Ширмохаммади М. и др. (февраль 2021 г.). «Связь полиморфизма Foxp3 rs3761548 с концентрацией цитокинов у пациентов с аденокарциномой желудка». Цитокин . 138 : 155351. doi : 10.1016/j.cyto.2020.155351 . ПМИД 33127257 . S2CID 226218796 .
- ^ Перейти обратно: а б Ли С, Цзян П, Вэй С, Сюй С, Ван Дж (июль 2020 г.). «Регуляторные Т-клетки в микроокружении опухоли: новые механизмы, потенциальные терапевтические стратегии и перспективы» . Молекулярный рак . 19 (1): 116. дои : 10.1186/s12943-020-01234-1 . ПМЦ 7367382 . ПМИД 32680511 .
- ^ Даунс-Каннер С., Берки С., Дельгофф Г.М., Эдвардс Р.П., Куриэль Т., Одунси К. и др. (март 2017 г.). «Подавляющий Ил-17А + Фоксp3 + и бывший Th17 Ил-17А нег Фоксp3 + T- reg- клетки являются источником опухолевых T -reg -клеток» . Nature Communications . 8 (1): 14649. Bibcode : ...814649D . doi : 10.1038/ncomms14649 . PMC 5355894. . PMID 28290453 2017NatCo
- ^ Тогаши Ю, Ситара К, Нисикава Х (июнь 2019 г.). «Регуляторные Т-клетки при раковой иммуносупрессии - значение для противораковой терапии». Обзоры природы. Клиническая онкология . 16 (6): 356–371. дои : 10.1038/s41571-019-0175-7 . ПМИД 30705439 . S2CID 59526013 .
- ^ Оуэ Ю., Нисикава Х. (июль 2019 г.). «Регуляторные Т-(Treg)-клетки при раке: могут ли Treg-клетки стать новой терапевтической мишенью?» . Раковая наука . 110 (7): 2080–2089. дои : 10.1111/cas.14069 . ПМК 6609813 . ПМИД 31102428 .
- ^ Марсон А., Кречмер К., Фрэмптон Г.М., Якобсен Е.С., Полански Дж.К., МакИсаак К.Д. и др. (февраль 2007 г.). «Заселение Foxp3 и регуляция ключевых генов-мишеней во время стимуляции Т-клеток» . Природа . 445 (7130): 931–935. Бибкод : 2007Natur.445..931M . дои : 10.1038/nature05478 . ПМК 3008159 . ПМИД 17237765 .
- ^ Эллис С.Д., Макговерн Дж.Л., ван Маурик А., Хоу Д., Эренштейн М.Р., Нотли Калифорния (октябрь 2014 г.). «Индуцированные CD8 + FoxP3 + Treg-клетки при ревматоидном артрите модулируются фосфорилированием p38 и моноцитами, экспрессирующими мембранный фактор некроза опухоли α и CD86». Артрит и ревматология . 66 (10): 2694–2705. дои : 10.1002/арт.38761 . ПМИД 24980778 . S2CID 39984435 .
- ^ Седдики Н., Кук Л., Сюй Д.С., Петсупхан С., Браун К., Сюй Ю. и др. (июнь 2014 г.). «Человеческие антигенспецифичные CD4⁺ CD25⁺ CD134⁺ CD39⁺ Т-клетки обогащены регуляторными Т-клетками и составляют значительную часть ответных реакций». Европейский журнал иммунологии . 44 (6): 1644–1661. дои : 10.1002/eji.201344102 . ПМИД 24752698 . S2CID 24012204 .
- ^ Заундерс Дж., Мунье М.Л., Седдики Н., Петт С., Ип С., Бейли М. и др. (август 2009 г.). «Высокие уровни человеческих антиген-специфичных CD4+ Т-клеток в периферической крови выявлены путем стимулированной совместной экспрессии CD25 и CD134 (OX40)» . Журнал иммунологии . 183 (4): 2827–2836. doi : 10.4049/jimmunol.0803548 . ПМИД 19635903 .
- ^ Полони С., Шонхофер С., Ивисон С., Левингс М.К., Штайнер Т.С., Кук Л. (июль 2023 г.). «Анализ маркеров, индуцированных активацией Т-клеток, в здоровье и болезни» . Иммунология и клеточная биология . 101 (6): 491–503. дои : 10.1111/imcb.12636 . ПМЦ 10952637 . ПМИД 36825901 . S2CID 257152898 .
- ^ Вечорек Г., Асемиссен А., Модель F, Турбачева И., Флосс С., Либенберг В. и др. (январь 2009 г.). «Количественный анализ метилирования ДНК FOXP3 как новый метод подсчета регуляторных Т-клеток в периферической крови и твердых тканях». Исследования рака . 69 (2): 599–608. дои : 10.1158/0008-5472.CAN-08-2361 . ПМИД 19147574 .
- ^ Агарвал А., Сингх М., Чаттерджи Б.П., Чаухан А., Чакраборти А. (2014). «Взаимодействие клеток Т-хелперов 17 с Treg CD4(+)CD25(high) FOXP3(+) в регуляции аллергической астмы у педиатрических пациентов» . Международный журнал педиатрии . 2014 : 636238. doi : 10.1155/2014/636238 . ПМК 4065696 . ПМИД 24995020 .
- ^ Лу Л.Ф., Линд Э.Ф., Гондек Д.С., Беннетт К.А., Глисон М.В., Пино-Лагос К. и др. (август 2006 г.). «Тучные клетки являются важными посредниками в регуляторной толерантности Т-клеток». Природа . 442 (7106): 997–1002. Бибкод : 2006Natur.442..997L . дои : 10.1038/nature05010 . ПМИД 16921386 . S2CID 686654 .
- ^ «Трегитоп: электроинструмент для иммуномодуляции» . ЭпиВакс . 2 августа 2016 г.
- ^ Хуэй Дж., Баснер-Чакарян Э., Чен Ю., Дэвидсон Р.Дж., Бухлис Г., Язичиоглу М. и др. (сентябрь 2013 г.). «Модуляция ответов CD8+ Т-клеток на векторы AAV с помощью эпитопов MHC класса II, полученных из IgG» . Молекулярная терапия . 21 (9): 1727–1737. дои : 10.1038/mt.2013.166 . ПМЦ 3776637 . ПМИД 23857231 .
- ^ Перейти обратно: а б Де Гроот А.С., Мойзе Л., Макмерри Дж.А., Вамбре Э., Ван Овертвелт Л., Мойнгеон П. и др. (октябрь 2008 г.). «Активация естественных регуляторных Т-клеток с помощью IgG Fc-производного пептида «Трегитопы» » . Кровь . 112 (8): 3303–3311. дои : 10.1182/blood-2008-02-138073 . ПМК 2569179 . ПМИД 18660382 .
- ^ «Новое вливание средств НИЗ на сумму 2,25 миллиона долларов для лечения Трегитопа EpiVax, предложенного для лечения, меняющего парадигму» . Жестокие биотехнологические исследования.
- ^ Су Ю, Росси Р., Де Гроот А.С., Скотт Д.В. (август 2013 г.). «Регуляторные Т-клеточные эпитопы (трегитопы) в IgG вызывают толерантность in vivo и лишены иммуногенности как таковой» . Журнал биологии лейкоцитов . 94 (2): 377–383. дои : 10.1189/jlb.0912441 . ПМЦ 3714563 . ПМИД 23729499 .
- ^ Казенс Л.П., Су Ю., Макклейн Э., Ли Х., Терри Ф., Смит Р. и др. (2013). «Применение природных эпитопов Treg, полученных из IgG (IgG Tregitopes), для индукции антиген-специфической толерантности на мышиной модели диабета 1 типа» . Журнал исследований диабета . 2013 : 621693. doi : 10.1155/2013/621693 . ПМЦ 3655598 . ПМИД 23710469 .
- ^ Казенс Л.П., Мингоцци Ф., ван дер Марель С., Су Ю., Гарман Р., Феррейра В. и др. (октябрь 2012 г.). «Обучение толерантности: новые подходы к ферментозаместительной терапии болезни Помпе» . Человеческие вакцины и иммунотерапия . 8 (10): 1459–1464. дои : 10.4161/hv.21405 . ПМЦ 3660767 . ПМИД 23095864 .
- ^ Казенс Л.П., Наджафян Н., Мингоцци Ф., Эльяман В., Мазер Б., Мойс Л. и др. (январь 2013 г.). «Исследование in vitro и in vivo эпитопов Treg, полученных из IgG (Tregitopes): новый многообещающий инструмент для индукции толерантности и лечения аутоиммунитета» . Журнал клинической иммунологии . 33 (Приложение 1): S43–S49. дои : 10.1007/s10875-012-9762-4 . ПМЦ 3538121 . ПМИД 22941509 .
- ^ Эльяман В., Хури С.Дж., Скотт Д.В., Де Гроот А.С. (2011). «Возможное применение трегитопов в качестве иммуномодулирующих средств при рассеянном склерозе» . Международное исследование неврологии . 2011 : 256460. doi : 10.1155/2011/256460 . ПМК 3175387 . ПМИД 21941651 .
- ^ Интернет-менделевское наследование в человеке IPEX
- ^ ipex в NIH / UW GeneTests
Внешние ссылки
[ редактировать ]- Регуляторные + Т-клетки в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)