Разрыв соединения
Тон или стиль этой статьи могут не отражать энциклопедический тон , используемый в Википедии . ( Апрель 2024 г. ) |
| Разрыв соединения | |
|---|---|
 Щелевой переход позвоночных | |
| Идентификаторы | |
| МеШ | D017629 |
| ТД | Х1.00.01.1.02024 |
| ФМА | 67423 |
| Анатомическая терминология | |
Щелевые соединения — одна из трех широких категорий межклеточных связей , которые образуются между несколькими типами клеток животных . [1] [2] Впервые их изображения были получены с помощью электронного микроскопа примерно в 1952 году. [3] [4] Они были названы в 1969 году. [5] после разрыва в 2–4 нм они соединяют клеточные мембраны. [6]
Щелевые соединения используют белковые комплексы, известные как глобулы, для соединения одной клетки с другой и для соединения везикул внутри клетки с внешней клеточной мембраной. [7] Глобулы состоят из более мелких белковых компонентов, называемых коннексинами . [8] [9] и шесть коннексинов образуют канал, называемый коннексоном . Существует более 26 различных коннексинов и как минимум 12 неконнексинных компонентов. [10] которые образуют специализированную область мембраны, называемую комплексом щелевых соединений. Эти компоненты включают белок ZO-1 , который скрепляет мембраны. [11] натриевые каналы , [12] и аквапорин . [13] [14]
Благодаря развитию секвенирования нового поколения стало известно больше белков щелевых соединений . Было обнаружено, что коннексины структурно гомологичны у позвоночных и беспозвоночных, но различаются по последовательности. [15] В результате термин иннексин используется для дифференциации коннексинов беспозвоночных. [16] Известно более 20 иннексинов. [17] наряду с уннексинами у паразитов и виннексинами у вирусов.
Щелевые контакты способны передавать потенциалы действия между нейронами . Пары коннексонов действуют как обобщенные регулируемые ворота для ионов и более мелких молекул между клетками. Полуканальные коннексоны образуют каналы во внеклеточную среду. [18] [19] [20] [21]
Щелевой переход также можно назвать связующим звеном или коммуникативным пятном. Его не следует путать с эфапсом . Хотя эфапса, как и щелевой контакт, включает передачу электрических сигналов, они отличаются друг от друга, поскольку эфапса включает в себя электрические сигналы, внешние по отношению к клеткам. Эфапсы часто изучаются в контексте электрически индуцированных потенциалов, распространяющихся среди групп мембран нервных клеток, даже при отсутствии связи между щелевыми соединениями и при отсутствии известных дискретных субклеточных структур. [22] [23] В отличие от щелевых соединений, конкретная структура, связанная с эфапсой, еще не описана, поэтому этот процесс часто называют эфапсическим соединением, а не эфапсой.
Структура [ править ]

У позвоночных щелевых соединений полуканалы представляют собой преимущественно гомо- гетерогексамеры белков коннексина или . Гетерогексамеры в бляшках щелевых соединений помогают формировать однородное межклеточное пространство размером 2–4 нм. [24] Таким образом, полуканалы в мембране каждой клетки выравниваются друг с другом, образуя путь межклеточной коммуникации . [25]
беспозвоночных Щелевые контакты белков состоят из иннексинов семейства . Иннексины не имеют значительной гомологии последовательностей с коннексинами. [26] Хотя иннексины отличаются по последовательности от коннексинов, они достаточно похожи на коннексины, чтобы образовывать щелевые соединения in vivo так же, как это делают коннексины. [27] [28] [29]
Недавно охарактеризованное семейство паннексинов , [30] который первоначально считался образующим межклеточные каналы (с аминокислотной последовательностью, аналогичной иннексинам [31] ) фактически функционирует как одномембранный канал, который сообщается с внеклеточной средой и, как было показано, пропускает кальций и АТФ . [32] Это привело к идее, что паннексины не могут образовывать межклеточные соединения таким же образом, как коннексины и иннексины, и поэтому не должны использовать одни и те же названия полуканалов/каналов. [33] Другие представили доказательства, основанные на генетическом секвенировании и общем функционировании в тканях, что паннексины по-прежнему следует считать частью семейства белков щелевых соединений, несмотря на структурные различия. коннексина . больше групп ортологов Эти исследователи также отмечают, что еще предстоит открыть [34]
Каналы щелевого соединения, образованные из двух одинаковых полуканалов, называются гомотипическими, а каналы с разными полуканалами — гетеротипическими. В свою очередь, полуканалы с одинаковым белковым составом называются гомомерными , а с разными белками — гетеромерными . Состав каналов влияет на функцию каналов щелевых соединений, и разные коннексины не обязательно образуют гетеротипы со всеми остальными. [35]
До того, как иннексины и коннексины были хорошо охарактеризованы, гены, кодирующие каналы щелевых соединений коннексина, были классифицированы в одну из трех групп (A, B и C; например, GJA1 , GJC1 ), на основе картирования генов и сходства последовательностей . [36] [37] [38] Однако гены коннексина не кодируют непосредственно экспрессию каналов щелевых соединений; гены могут производить только белки, составляющие каналы щелевых соединений. Наиболее широко используется альтернативная система наименования, основанная на молекулярной массе белка (например, коннексин43=GJA1, коннексин30.3=GJB4).
Уровни организации [ править ]
У позвоночных две пары из шести белков коннексина образуют коннексон. У беспозвоночных шесть белков-иннексинов образуют иннексон. В остальном структуры аналогичны. Термин «полуканал», используемый в этом описании, относится к половине канала щелевого перехода.
- Гены коннексина (ДНК) транскрибируются в РНК, которая затем транслируется с образованием коннексина.
- Один белок коннексин имеет четыре трансмембранных домена. [24] [39]
- Шесть белков коннексина создают один канал коннексина (полуканал). Когда идентичные белки коннексина объединяются в один коннексон, его называют гомомерным коннексоном. Когда разные белки коннексина объединяются в один коннексон, его называют гетеромерным коннексоном.
- Два коннексона, соединенные через клеточную мембрану, образуют канал щелевого соединения.
Когда два идентичных коннексона собираются вместе, образуя канал щелевого соединения, он называется гомотипическим каналом. Когда один гомомерный коннексон и один гетеромерный коннексон соединяются, это называется гетеротипическим каналом щелевого соединения. Когда два гетеромерных коннексона соединяются, это также называется гетеротипическим каналом щелевого соединения. - От десятков до тысяч каналов щелевых соединений группируются в определенных областях, что позволяет формировать пары коннексонов. [40] Макромолекулярный комплекс называется бляшкой щелевого соединения. Молекулы, отличные от коннексинов, участвуют в бляшках щелевых соединений, включая zonula occludens-1 ( ZO-1 ) и натриевые каналы . [11] [12]
Свойства пар коннексонов [ править ]

Пара каналов коннексон или иннексон:
- Обеспечивает прямую электрическую связь между клетками, хотя разные субъединицы полуканалов могут придавать разную одноканальную проводимость , примерно от 30 до 500 пС.
- Обеспечивает химическую связь между клетками посредством передачи небольших вторичных мессенджеров , таких как инозитолтрифосфат ( IP
3 ) и кальций ( Ca 2+
), [42] хотя разные субъединицы полуканалов могут придавать разную селективность конкретным молекулам. - Обычно обеспечивает трансмембранное перемещение молекул размером менее 485 дальтон. [43] (1100 дальтон через щелевые контакты беспозвоночных [44] ), хотя разные субъединицы полуканалов могут придавать порам разные размеры и разную селективность по заряду. Крупные биомолекулы, включая нуклеиновые кислоты и белки , не могут переноситься в цитоплазме между клетками через пары полуканалов щелевого соединения.
- Обеспечивает, чтобы молекулы и ток, проходящие через щелевой переход, не просачивались в межклеточное пространство.
коннексонов полуканалов Свойства как
Непарные коннексоны или иннексоны могут действовать как полуканалы в одной мембране, позволяя клетке обмениваться молекулами непосредственно с внешней частью клетки. Было показано, что коннексоны будут доступны для этого до того, как они будут включены в бляшки щелевого соединения. [40] Некоторые свойства этих непарных коннексонов перечислены ниже:
- Размер пор или трансмембранных каналов сильно варьируется и находится в диапазоне примерно 8-20 Å в диаметре. [45]
- Они соединяют цитоплазму клетки с внешней поверхностью клетки и по умолчанию находятся в закрытом состоянии, чтобы предотвратить утечку из клетки. [46] [47]
- Некоторые коннексоны реагируют на внешние факторы открыванием. Механический сдвиг и различные заболевания могут стать причиной этого. [48]
Установление дополнительных свойств коннексонов, отличных от свойств пар коннексонов, оказывается трудным из-за экспериментального разделения их эффектов на организмах. [48]
Возникновение и распространение [ править ]
Щелевые соединения наблюдались в различных органах и тканях животных, где клетки контактируют друг с другом. С 1950-х по 1970-е годы они были обнаружены в:
- Человеческий островок Лангерганса, [49] миометрий, [50] и линза для глаз [51]
- Поджелудочная железа крысы, печень, кора надпочечников, придатки яичка, двенадцатиперстная кишка, мышцы, [52] и семенные канальцы [53]
- Роговица кролика, [54] яичник, [55] и кожа [56]
- Сетчатка обезьяны [57]
- Куриные эмбрионы [58]
- Эмбрионы лягушки [59]
- Рыбья бластодерма [60]
- Раковые нервы [61]
- миноги и Сердце оболочника [62] [63]
- Акустико-вестибулярные рецепторы золотых рыбок и хомяков, чувствительные к давлению [64]
- Дафния печеночная слепая [65]
- головоногих моллюсков Пищеварительный эпителий [66]
- Гидра мышца [67]
- Капсулы гемоцитов таракана [68]
- Реагрегированные клетки [69] [70]
Щелевые соединения по-прежнему обнаруживаются почти во всех здоровых клетках животных, которые соприкасаются друг с другом. Такие методы, как конфокальная микроскопия, позволяют более быстро обследовать большие площади тканей. Было показано, что в тканях, которые традиционно считались изолированными клетками, например, в костях, были клетки, которые все еще были связаны щелевыми соединениями, хотя и слабо. [71] Исключением являются клетки, которые обычно не контактируют с соседними клетками, например клетки крови, взвешенные в плазме крови. Скелетные мышцы взрослых являются возможным исключением из правила, хотя из-за их большого размера трудно быть в этом уверенным. Аргумент, используемый против щелевых соединений скелетных мышц, заключается в том, что, если бы они присутствовали, щелевые соединения могли бы произвольно распространять сокращения через клетки, составляющие мышцу. Однако другие типы мышц имеют щелевые соединения, которые не вызывают произвольных сокращений. [72] Иногда количество щелевых соединений уменьшено или отсутствует в больных тканях, таких как рак. [73] [74] [75] или процесс старения. [76]
С момента открытия иннексинов, паннексинов и уннексинов пробелы в наших знаниях о межклеточной коммуникации становятся все более очевидными. Иннексины выглядят и ведут себя аналогично коннексинам, и можно увидеть, что они выполняют ту же роль, что и коннексины у беспозвоночных. Паннексины также внешне похожи на коннексины, хотя, по-видимому, им нелегко образовывать щелевые контакты. Из более чем 20 групп многоклеточных животных коннексины обнаружены только у позвоночных и оболочников . Гораздо более распространены иннексины и паннексины, включая гомологи иннексина у позвоночных. [77] [78] Одноклеточные паразиты Trypanosomatidae предположительно имеют гены уннексина, способствующие заражению животных, включая человека. [79] Еще более мелкий аденовирус имеет собственный виннексин. [80] очевидно, получен из иннексина, чтобы способствовать его передаче между насекомыми-хозяевами вируса.
Термин «щелевое соединение» не может быть определен одним белком или семейством белков с определенной функцией. Например, структуры щелевых контактов обнаружены у губок , несмотря на отсутствие паннексинов. Хотя мы все еще находимся на ранних стадиях понимания нервной системы губки. [81] щелевые контакты губок могут пока указывать на пути межклеточной коммуникации. [82] [83]
Функции [ править ]
приписывают по крайней мере пять дискретных функций Белкам щелевых соединений :
- Электрическая и метаболическая связь между клетками
- Электрический и метаболический обмен через полуканалы
- Гены-супрессоры опухолей ( Cx43 , Cx32 и Cx36 )
- Адгезивная функция не зависит от проводящего канала щелевого соединения (нейронная миграция в неокортексе)
- Роль карбоксильного конца в сигнальных цитоплазматических путях (Cx43)
В более общем смысле можно рассматривать щелевые контакты на простейшем уровне как прямой путь от клетки к клетке для электрических токов, малых молекул и ионов. Контроль этой связи позволяет осуществлять сложные последующие воздействия на многоклеточные организмы.
Эмбриональное развитие органов и тканей [ править ]
В 1980-х годах были исследованы более тонкие роли щелевых соединений в коммуникации. Было обнаружено, что связь через щелевые соединения может быть нарушена добавлением антител против коннексина в эмбриональные клетки. [84] [85] Эмбрионы с участками заблокированных щелевых соединений не смогли нормально развиваться. Механизм, с помощью которого антитела блокируют щелевые контакты, был неясен; Для выяснения механизма были проведены систематические исследования. [86] [87] Уточнение этих исследований показало, что щелевые соединения играют ключевую роль в развитии полярности клеток. [88] и симметрия слева и справа у животных. [89] [90] Хотя передача сигналов, определяющая положение органов тела, по-видимому, зависит от щелевых соединений, то же самое происходит и с более фундаментальной дифференцировкой клеток на более поздних стадиях эмбрионального развития. [91] [92] [93] [94] [95]
Было обнаружено, что щелевые контакты отвечают за передачу сигналов, необходимых для воздействия лекарств. [96] И наоборот, было показано, что некоторые лекарства блокируют каналы щелевых соединений. [97]
Эффект свидетеля болезнь и
Гибель клеток [ править ]
Эффект свидетеля имеет оттенок убийства невиновного свидетеля. Когда клетки умирают или разрушаются из-за болезни или травмы, сообщения передаются соседним клеткам через щелевые контакты. Это может привести к гибели здоровых клеток-свидетелей. [98] Поэтому важно учитывать эффект свидетеля в больных клетках, открывая возможности для большего финансирования и расцвета исследований. [99] [100] [101] [102] [103] [104] [105] [106] [107]
Позже эффект свидетеля был исследован в отношении клеток, поврежденных радиацией или механическим повреждением и, в свою очередь, заживляющих ран. [108] [109] [110] [111] [112] Болезнь, по-видимому, влияет на способность щелевых соединений выполнять свою роль в заживлении ран. [113] [114] Пероральное введение блокаторов щелевых соединений для уменьшения симптомов заболевания в отдаленных частях тела постепенно становится реальностью. [115]
Реструктуризация тканей [ править ]
Хотя существует тенденция сосредотачиваться на эффекте свидетеля при заболевании из-за возможности терапевтических возможностей, есть свидетельства того, что существует более центральная роль в нормальном развитии тканей. Для того чтобы ткань достигла своей окончательной конфигурации, может потребоваться смерть некоторых клеток и окружающего их матрикса; щелевые соединения кажутся важными для этого процесса. [116] [117] Существуют также более сложные исследования, которые пытаются объединить наше понимание одновременной роли щелевых соединений как в заживлении ран, так и в развитии тканей. [118] [119] [120]
Болезнь [ править ]
Мутации коннексинов связаны со многими заболеваниями у человека, включая глухоту , [121] сердца мерцательная аритмия (остановка) и катаракта . Изучение этих мутаций помогло прояснить некоторые функции коннексинов. [122] [123]
Считается, что полуканалы играют основную роль в прогрессировании и тяжести многих заболеваний; Частично это связано с тем, что полуканалы представляют собой открытую дверь наружу каждой клетки. [48]
Области электрической связи [ править ]
Щелевые контакты электрически и химически соединяют клетки по всему телу большинства животных. Электрическая связь может быть относительно быстродействующей и может использоваться на коротких расстояниях внутри организма. Ткани в этом разделе имеют хорошо известные функции, которые, как наблюдают, координируются щелевыми соединениями, при этом межклеточная передача сигналов происходит в течение микросекунд или меньше.
Сердце [ править ]

Щелевые соединения особенно важны для сердечной мышцы : сигнал о сокращении эффективно передается через щелевые соединения, позволяя клеткам сердечной мышцы сокращаться в унисон. Важность подчеркивается тем, что вторичный эфаптический путь сигнала к сокращению также связан с бляшками щелевого соединения. Эта избыточность в передаче сигнала, связанная с бляшками щелевых соединений, является первой описанной и затрагивает натриевые каналы, а не коннексины. [12] [124]
Глазная линза [ править ]
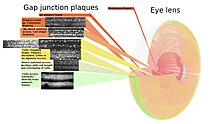
Точный контроль преломления света, структурных размеров и прозрачности являются ключевыми аспектами структуры хрусталика, позволяющими фокусировать взгляд. Прозрачности способствует отсутствие нервов и кровеносных сосудов в хрусталике, поэтому на щелевые соединения приходится большая нагрузка межклеточной коммуникации, чем в других тканях, что отражается в большом количестве щелевых соединений. Кристалличность . хрусталика также означает, что клетки и щелевые соединения хорошо упорядочены для систематического картирования того, где находятся бляшки щелевых соединений Поскольку ни одна клетка внутренней части хрусталика не теряется в течение жизни животного, возможна полная карта щелевых соединений. [125]
На соответствующем рисунке показано, как размер, форма и частота образования бляшек щелевых соединений изменяются по мере роста клеток. По мере роста клетки волокон постепенно изолируются от более прямого обмена метаболитами с водянистой влагой через капсулу и эпителий хрусталика. Изолированность коррелирует с классической круглой формой более крупных бляшек, показанных в желтой зоне и разрушающихся. Изменение морфологии волоконных клеток требует движения везикул через бляшки щелевого соединения на более высоких частотах в этой области. [126]
Нейроны [ править ]
Щелевой контакт, расположенный в нейроне, часто называют электрическим синапсом . Электрический синапс был обнаружен с помощью электрических измерений до того, как была описана структура щелевого перехода. Электрические синапсы присутствуют во всей центральной нервной системе и были изучены, в частности, в неокортексе , гиппокампе , вестибулярном ядре , ретикулярном ядре таламуса , голубом пятне , нижнем оливковом ядре , мезэнцефальном ядре тройничного нерва , вентральной области покрышки , обонятельной луковице , сетчатке и спинной позвоночных мозг . [127]
Были некоторые наблюдения взаимодействия в голубом пятне между слабыми нейронами и глиальными клетками и в мозжечке между нейронами Пуркинье и глиальными клетками Бергмана . Похоже, что астроциты соединены щелевыми контактами как с другими астроцитами, так и с олигодендроцитами . [128] Более того, мутации в генах щелевого соединения Cx43 и Cx56.6 вызывают дегенерацию белого вещества, аналогичную той, которая наблюдается при болезни Пелицеуса-Мерцбахера и рассеянном склерозе .
Белки коннексина, экспрессируемые в щелевых соединениях нейронов, включают m CX36 , mCX57 и m CX45 , при этом мРНК по меньшей мере пяти других коннексинов (m Cx26 , m Cx30.2 , m Cx32 , m Cx43 , m Cx47 ) обнаружены, но без иммуноцитохимических доказательств наличия соответствующий белок внутри ультраструктурно определенных щелевых соединений. Эти мРНК, по-видимому, подавляются или разрушаются микроинтерферирующими РНК ( миРНК ), специфичными для клеточного типа и клеточной линии.
Сетчатка [ править ]
Нейроны сетчатки демонстрируют обширную связь как внутри популяций одного типа клеток, так и между разными типами клеток. [129]
Матка [ править ]
Мышца матки ( миометрий остается в спокойном, расслабленном состоянии ) во время беременности для поддержания развития плода . Непосредственно перед родами миометрий трансформируется в активированную сократительную единицу за счет увеличения экспрессии коннексина-43 ( CX43 , также известного как белок Gap Junction Alpha-1, GJA1 ), способствующего образованию щелевого соединения (GJ) между отдельными клетками миометрия. Важно отметить, что образование ЩК способствует коммуникации между соседними миоцитами , что облегчает перенос небольших молекул, таких как вторичные мессенджеры, метаболиты и небольшие ионы, для электрического связывания. Как и у всех видов, сокращения миометрия матки распространяются от спонтанных потенциалов действия в результате внезапного изменения проницаемости плазматической мембраны . Это приводит к увеличению внутриклеточной концентрации Ca²⁺ , способствуя распространению потенциала действия через электрически связанные клетки. [130] Совсем недавно было обнаружено, что макрофаги матки непосредственно физически соединяются с миоцитами матки через CX43, передавая Ca²⁺, что способствует сокращению и возбуждению мышц матки во время начала родов. [131]
Функция полуканала [ править ]


Полуканалы вносят вклад в клеточную сеть щелевых соединений и позволяют высвобождать АТФ , глутамат , НАД+ и простагландин Е2 из клеток, которые могут действовать как посланники для клеток, в противном случае отключенных от такой передачи сообщений. [132] В этом смысле бляшка щелевого соединения образует отношения один к одному с соседней клеткой, последовательно соединяя множество клеток вместе. Полуканалы образуют связь один ко многим с окружающей тканью.
В более широком масштабе связь клеток «один ко многим» обычно осуществляется сосудистой и нервной системами. Это затрудняет обнаружение вклада полуканалов во внеклеточную связь в целых организмах. В глазном хрусталике сосудистая и нервная системы отсутствуют, что делает большую зависимость от полуканалов и облегчает их обнаружение. На границе раздела хрусталика с водянистой влагой (где хрусталик обменивается метаболитами) на сопровождающих микрофотографиях можно увидеть как бляшки щелевых соединений, так и более диффузное распределение коннексонов.
Открытие [ править ]
Форма для работы [ править ]
Задолго до демонстрации разрыва в щелевых соединениях их видели на стыке соседних нервных клеток. Непосредственная близость соседних клеточных мембран к щелевому соединению заставила исследователей предположить, что они играют роль в межклеточной коммуникации, в частности в передаче электрических сигналов. [61] [133] [134] Также было доказано, что щелевые контакты обладают электрическим выпрямлением и называются электрическими синапсами . [135] [136] Позже было обнаружено, что химические вещества также могут транспортироваться между клетками через щелевые контакты. [137]
В большинстве ранних исследований неявно или явно было указано, что область щелевого соединения отличалась по структуре от окружающих мембран, поэтому она выглядела по-другому. Было показано, что щелевое соединение создает микроокружение между двумя клетками во внеклеточном пространстве или зазоре . Эта часть внеклеточного пространства была в некоторой степени изолирована от окружающего пространства, а также соединена тем, что мы сейчас называем парами коннексонов, которые образуют еще более плотно запечатанные мостики, пересекающие щелевое соединение между двумя клетками. При просмотре в плоскости мембраны с помощью методов замораживания-перелома возможно распределение коннексонов внутри бляшки щелевого соединения с более высоким разрешением. [138]
В некоторых соединениях наблюдаются свободные островки коннексина. Наблюдение в основном не имело объяснения до тех пор, пока Peracchia с помощью тонких срезов трансмиссионной электронной микроскопии (ТЕМ) не показала, что везикулы систематически связаны с бляшками щелевых контактов. [139] Исследование Пераккиа, вероятно, было также первым исследованием, описывающим парные структуры коннексонов, которые он назвал глобулой . Исследования, показывающие, что везикулы связаны с щелевыми соединениями, и предполагающие, что содержимое везикул может перемещаться через бляшки соединения между двумя клетками, были редкими, поскольку большинство исследований были сосредоточены на коннексонах, а не на везикулах. Более позднее исследование с использованием комбинации методов микроскопии подтвердило ранние доказательства вероятной функции щелевых соединений при межклеточном переносе везикул. Области переноса везикул были связаны с островками, свободными от коннексина, внутри бляшек щелевых соединений. [126] Было показано, что коннексин 43 необходим для переноса целых митохондрий в соседние клетки, хотя не установлено, передаются ли митохондрии непосредственно через мембрану или внутри пузырька. [140]
нервные синапсы и химические Электрические
Из-за широкого распространения щелевых соединений в типах клеток, отличных от нервных клеток, термин « щелевые соединения» стал использоваться более широко, чем такие термины, как «электрический синапс» или «нексус». Другое измерение во взаимоотношениях между нервными клетками и щелевыми соединениями было выявлено при изучении образования химических синапсов и наличия щелевых соединений. Отслеживая развитие нервов у пиявок с подавленной экспрессией щелевого соединения, было показано, что двунаправленный щелевой контакт (электрический нервный синапс) должен сформироваться между двумя клетками, прежде чем они смогут вырасти и сформировать однонаправленный химический нервный синапс . [141] Химический нервный синапс — это синапс, который чаще всего называют более двусмысленным термином « нервный синапс» .
Состав [ править ]
Коннексины [ править ]
Очищение [142] [143] Из бляшек межклеточных щелевых соединений, обогащенных белком, образующим каналы ( коннексином был обнаружен белок, образующий гексагональные массивы ), при дифракции рентгеновских лучей . В связи с этим систематическое изучение и идентификация преобладающего белка щелевого соединения [144] стало возможным. Усовершенствованные ультраструктурные исследования с помощью TEM [145] [146] показали, что белок встречается комплементарным образом в обеих клетках, участвующих в бляшке щелевого соединения. Бляшка щелевого соединения представляет собой относительно большую область мембраны, наблюдаемую на тонком срезе ПЭМ и замороженном переломе (FF), заполненную трансмембранными белками как в тканях, так и в более щадящих препаратах щелевого соединения. С очевидной способностью одного только белка обеспечивать межклеточную связь, наблюдаемую в щелевых соединениях. [147] термин «щелевое соединение» имел тенденцию становиться синонимом группы собранных коннексинов, хотя это не было показано in vivo. Биохимический анализ щелевых соединений, выделенных из различных тканей, выявил семейство коннексинов. [148] [149] [150]
Ультраструктура и биохимия изолированных щелевых соединений, о которых уже упоминалось, показали, что коннексины преимущественно группируются в бляшках или доменах щелевых соединений, и коннексины являются наиболее охарактеризованным компонентом. Было отмечено, что организация белков в массивы с бляшками щелевых соединений может иметь важное значение. [59] [151] Вполне вероятно, что эта ранняя работа уже отражала присутствие не только коннексинов в щелевых соединениях. Объединение новых областей замораживания-разрыва, чтобы заглянуть внутрь мембран, и иммуноцитохимии для маркировки клеточных компонентов (иммуномаркировка реплик замораживания-разрыва или FRIL и иммуномаркировка тонких срезов) показало, что бляшки щелевых соединений in vivo содержат белок коннексин. [152] [125] Более поздние исследования с использованием иммунофлуоресцентной микроскопии больших участков ткани прояснили разнообразие более ранних результатов. Было подтверждено, что бляшки щелевых соединений имеют переменный состав, поскольку являются домом для белков коннексона и неконнексина, что делает современное использование терминов «щелевое соединение» и «бляшка щелевого соединения» невзаимозаменяемыми. [14] Подводя итог, можно сказать, что в ранней литературе термин «щелевое соединение» относился к регулярному зазору между мембранами у позвоночных и непозвоночных животных, по-видимому, соединенному «глобулами». Соединение коррелирует со способностью клетки напрямую соединяться со своими соседями через поры в их мембранах. Тогда какое-то время щелевые соединения относились только к структуре, содержащей коннексины, и не считалось, что здесь больше ничего задействовано. Позже было обнаружено, что «бляшка» щелевого соединения содержит и другие молекулы, которые помогли определить ее структуру и заставить ее функционировать.
«Мемориальная доска» или «Мемориальная доска» [ править ]
Ранние описания щелевых соединений , коннексонов или иннексонов не относились к ним как таковым; использовалось много других терминов. Вероятно, синаптические диски [153] были точной ссылкой на бляшки на щелевых соединениях. Хотя подробная структура и функции коннексона в то время были описаны ограниченно, общая структура диска была относительно большой и ее легко можно было увидеть с помощью различных методов ПЭМ. Диски позволили исследователям с помощью TEM легко найти коннексоны, содержащиеся внутри диска, например, участки in vivo и in vitro. Диск или бляшка , по-видимому, имели структурные свойства, отличные от свойств, придаваемых только коннексонами/иннексонами. [67] Считалось, что если участок мембраны в бляшке передает сигналы, то этот участок мембраны необходимо каким-то образом герметизировать, чтобы предотвратить утечку. [154] Более поздние исследования показали, что бляшки щелевого соединения являются домом для белков, не содержащих коннексины, что делает современное использование терминов «щелевое соединение» и «бляшка щелевого соединения» невзаимозаменяемыми, поскольку область бляшки щелевого соединения может содержать белки, отличные от коннексинов. [14] [126] Так как коннексины не всегда занимают всю площадь бляшки, остальные компоненты, описанные в литературе, могут быть лишь долговременными или кратковременными резидентами. [155] [35] [156]
Исследования, позволяющие увидеть внутреннюю плоскость мембраны щелевых соединений во время формирования, показали, что «формационная бляшка» образовывалась между двумя клетками до того, как коннексины проникли внутрь. Это были области, свободные от частиц - при наблюдении с помощью TEM FF это указывало на очень маленький размер или отсутствие трансмембранного образования . белки, вероятно, присутствовали. Мало что известно о том, какие структуры составляют формирующую бляшку или как структура формационной бляшки меняется, когда коннексины и другие компоненты перемещаются внутрь и наружу. Одно из более ранних исследований образования малых щелевых соединений описывает ряды частиц и гало без частиц. [157] Щелевые контакты большего размера описывались как бляшки с перемещающимися в них коннексинами. Считалось, что щелевые контакты частиц образуются через 4–6 часов после появления бляшек образования. [158] как коннексины могут транспортироваться к бляшкам с помощью тубулина . Становится все более ясным, [88] [159]
Ранним исследователям было трудно проанализировать образование бляшек и неконнексиновой части классической бляшки щелевого соединения. На TEM FF и тонком срезе он выглядит как домен липидной мембраны, который каким-то образом может образовывать сравнительно жесткий барьер для других липидов и белков. Имеются косвенные доказательства того, что определенные липиды преимущественно участвуют в формировании бляшек, однако это нельзя считать окончательным. [160] [161] Трудно представить себе разрушение мембраны для анализа мембранных бляшек без изменения их состава. Путем изучения коннексинов, находящихся в мембранах, были изучены липиды, связанные с коннексинами. [162] Было обнаружено, что специфические коннексины имеют тенденцию преимущественно ассоциироваться со специфическими фосфолипидами. Поскольку формирование бляшек предшествует коннексинам, эти результаты все еще не дают уверенности в том, что уникально в составе самих бляшек. Другие данные показывают, что коннексины связаны с белковыми каркасами, используемыми в другом соединении, zonula occludens ZO-1 . [163] Хотя это помогает нам понять, как коннексины могут перемещаться в бляшки, образующие щелевые соединения, состав самой бляшки все еще несколько схематичен. Некоторый прогресс в составлении бляшек щелевого соединения in vivo достигнут с использованием TEM FRIL. [155] [163]
См. также [ править ]
- Модуляция щелевого перехода
- Белок щелевого соединения
- Иннексин
- Виннексин
- Вставной диск
- Ионный канал
- Узловой комплекс
- Плотное соединение
Ссылки [ править ]
- ^ Келселл, Дэвид П.; Данлоп, Джон; Ходжинс, Малкольм Б. (2001). «Болезни человека: ключ к расшифровке кода коннексина?». Тенденции в клеточной биологии . 11 (1): 2–6. дои : 10.1016/S0962-8924(00)01866-3 . ПМИД 11146276 .
- ^ Виллеке, Клаус; Эйбергер, Юрген; Деген, Иоахим; Экардт, Доминик; Ромуальди, Алессандро; Гюльденагель, Мартин; Дойч, Урбан; Зёль, Горан (2002). «Структурное и функциональное разнообразие генов коннексина в геноме мыши и человека» . Биологическая химия . 383 (5): 725–37. дои : 10.1515/BC.2002.076 . ПМИД 12108537 . S2CID 22486987 .
- ^ Робертсон, доктор юридических наук (1952). Ультраструктура синапса беспозвоночных (Диссертация). Кембридж, Массачусетс: Массачусетский технологический институт.
- ^ Робертсон, доктор юридических наук (1 февраля 1953 г.). «Ультраструктура двух синапсов беспозвоночных». Экспериментальная биология и медицина . 82 (2): 219–223. дои : 10.3181/00379727-82-20071 . ПМИД 13037850 .
- ^ Брайтман М.В., Риз Т.С. (март 1969 г.). «Соединения между тесно прилегающими клеточными мембранами в мозге позвоночных» . Дж. Клеточная Биол . 40 (3): 648–677. дои : 10.1083/jcb.40.3.648 . ПМК 2107650 . ПМИД 5765759 .
- ^ Ревель, Япония; Карновский, MJ (1 июня 1967 г.). «Гексагональный набор субъединиц в межклеточных соединениях сердца и печени мыши» . Журнал клеточной биологии . 33 (3): С7–12. дои : 10.1083/jcb.33.3.C7 . ПМК 2107199 . ПМИД 6036535 .
- ↑ Перейти обратно: Перейти обратно: а б Пераккья, Камилло (1 апреля 1973 г.). «Низкоомные соединения у раков» . Журнал клеточной биологии . 57 (1): 66–76. дои : 10.1083/jcb.57.1.66 . ПМК 2108946 . ПМИД 4120611 .
- ^ Гуденаф, Дэниел А. (1 мая 1974 г.). «Объемная изоляция щелевых соединений гепатоцитов мыши» . Журнал клеточной биологии . 61 (2): 557–563. дои : 10.1083/jcb.61.2.557 . ПМК 2109294 . ПМИД 4363961 .
- ^ Бейер, ЕС; Пол, ДЛ; Гуденаф, Д.А. (1 декабря 1987 г.). «Коннексин43: белок сердца крысы, гомологичный белку щелевого соединения печени» . Журнал клеточной биологии . 105 (6): 2621–2629. дои : 10.1083/jcb.105.6.2621 . ПМК 2114703 . ПМИД 2826492 .
- ^ Эрве, Жан-Клод; Бурмейстер, Николя; Сарруй, Дени; Даффи, Хизер С. (май 2007 г.). «Комплексы щелевых соединений: от партнеров к функциям». Прогресс биофизики и молекулярной биологии . 94 (1–2): 29–65. doi : 10.1016/j.pbiomolbio.2007.03.010 . ПМИД 17507078 .
- ↑ Перейти обратно: Перейти обратно: а б Жилерон, Жером; Каретт, Дайан; Фиорини, Селин; Бенкдане, Мерием; Сегретен, Доминик; Пуэнтис, Жорж (март 2009 г.). «Эндоцитоз бляшек щелевого соединения коннексина 43 подразумевает молекулярное ремоделирование партнеров ZO-1 и c-Src» . Коммуникативная и интегративная биология . 2 (2): 104–106. дои : 10.4161/cib.7626 . ПМК 2686357 . ПМИД 19704902 .
- ↑ Перейти обратно: Перейти обратно: а б с Иванович, Эна; Кучера, Ян П. (ноябрь 2021 г.). «Локализация кластеров каналов Na + в суженной перинексии щелевых соединений усиливает передачу сердечного импульса посредством эфаптической связи: модельное исследование» . Журнал физиологии . 599 (21): 4779–4811. дои : 10.1113/JP282105 . ПМЦ 9293295 . ПМИД 34533834 .
- ^ Ю, Сюнь Шон; Инь, Синье; Лафер, Эйлин М.; Цзян, Жан X. (июнь 2005 г.). «Регуляция развития прямого взаимодействия между внутриклеточной петлей коннексина 45.6 и С-концом основного внутреннего белка (аквапорина-0)» . Журнал биологической химии . 280 (23): 22081–22090. дои : 10.1074/jbc.M414377200 . ПМИД 15802270 .
- ↑ Перейти обратно: Перейти обратно: а б с Грюйтерс, WTM (1989). «Белок, не являющийся коннексоном (MIP), участвует в формировании щелевых соединений хрусталика» . Журнал клеточной науки . 93 (3): 509–513. дои : 10.1242/jcs.93.3.509 . ПМИД 2691517 .
- ^ Фелан, Полина; Стеббингс, Люси А.; Бейнс, Ричард А.; Бэкон, Джонатан П.; Дэвис, Джейн А.; Форд, Крис (январь 1998 г.). «Белок Shaking-B дрозофилы образует щелевые соединения в парных ооцитах Xenopus». Природа . 391 (6663): 181–184. Бибкод : 1998Natur.391..181P . дои : 10.1038/34426 . ПМИД 9428764 . S2CID 205003383 .
- ^ Фелан, Полина; Бэкон, Джонатан П.; А. Дэвис, Джейн; Стеббингс, Люси А.; Тодман, Мартин Г. (сентябрь 1998 г.). «Иннексины: семейство белков щелевых соединений беспозвоночных» . Тенденции в генетике . 14 (9): 348–349. дои : 10.1016/s0168-9525(98)01547-9 . ПМЦ 4442478 . ПМИД 9769729 .
- ^ Ортис, Дженнифер; Бобков Юрий В; ДеБиасс, Мелисса Б; Митчелл, Дороти Дж; Эдгар, Эллисон; Мартиндейл, Марк К; Мосс, Энтони Дж; Бабонис, Лесли С; Райан, Джозеф Ф. (3 февраля 2023 г.). «Независимая радиационная передача сигналов иннексина в гребневиках» . Молекулярная биология и эволюция . 40 (2): msad025. дои : 10.1093/molbev/msad025 . ПМЦ 9949713 . ПМИД 36740225 .
- ^ Фуршпан, Э.Дж.; Поттер, Д.Д. (август 1957 г.). «Механизм передачи нервного импульса в синапсах рака». Природа . 180 (4581): 342–343. Бибкод : 1957Natur.180..342F . дои : 10.1038/180342a0 . ПМИД 13464833 . S2CID 4216387 .
- ^ Лампе, Пол Д.; Лау, Алан Ф. (2004). «Влияние фосфорилирования коннексина на связь по щелевым соединениям» . Международный журнал биохимии и клеточной биологии . 36 (7): 1171–86. дои : 10.1016/S1357-2725(03)00264-4 . ПМК 2878204 . ПМИД 15109565 .
- ^ Лампе, Пол Д.; Лау, Алан Ф. (2000). «Регуляция щелевых соединений путем фосфорилирования коннексинов». Архив биохимии и биофизики . 384 (2): 205–15. дои : 10.1006/abbi.2000.2131 . ПМИД 11368307 .
- ^ Сцемес, Элиана; Спрей, Дэвид С.; Меда, Паоло (апрель 2009 г.). «Коннексины, паннексины, иннексины: новые роли «полуканалов» » . Архив Pflügers: Европейский журнал физиологии . 457 (6): 1207–1226. дои : 10.1007/s00424-008-0591-5 . ПМК 2656403 . ПМИД 18853183 .
- ^ Мартинес-Банаклоча, Маркос (13 февраля 2020 г.). «Астроглиальная изопотенциальность и влияние биомагнитного поля, связанного с кальцием, на соединение кортикальных нейронов» . Клетки . 9 (2): 439. doi : 10.3390/cells9020439 . ПМК 7073214 . ПМИД 32069981 .
- ^ Паркер, Дэвид (22 декабря 2022 г.). «Нейробиологическая редукция: от клеточных объяснений поведения к вмешательствам» . Границы в психологии . 13 : 987101. doi : 10.3389/fpsyg.2022.987101 . ПМЦ 9815460 . ПМИД 36619115 .
- ↑ Перейти обратно: Перейти обратно: а б Осима, Ацунори; Цукихара, Томитаке (2009) . ; Маэда, Соджи; Суга , Мичихиро 7238): 597–602. Бибкод : 2009Natur.458..597M . doi : nature07869 . PMID 19340074. 10.1038 / S2CID 4431769 .
- ^ Перкинс, Гай А.; Гуденаф, Дэниел А.; Сосинский, Джина Э. (1998). «Формирование межклеточного канала щелевого соединения требует поворота на 30 градусов для соединения двух противоположных коннексонов». Журнал молекулярной биологии . 277 (2): 171–7. дои : 10.1006/jmbi.1997.1580 . ПМИД 9514740 .
- ^ Консорциум по секвенированию C. elegans (11 декабря 1998 г.). «Последовательность генома нематоды C. elegans: платформа для изучения биологии». Наука . 282 (5396): 2012–2018. Бибкод : 1998Наука...282.2012. . дои : 10.1126/science.282.5396.2012 . ПМИД 9851916 .
- ^ Ганфорнина, доктор медицинских наук; Санчес, Д; Эррера, М; Бастиани, MJ (1999). «Экспрессия развития и молекулярная характеристика двух белков каналов щелевых соединений, экспрессируемых во время эмбриогенеза у кузнечика Schistocerca americana». Генетика развития . 24 (1–2): 137–150. doi : 10.1002/(SICI)1520-6408(1999)24:1/2<137::AID-DVG13>3.0.CO;2-7 . hdl : 10261/122956 . ПМИД 10079517 .
- ^ Старич, Т.А. (1996). «eat-5 и unc-7 представляют собой мультигенное семейство Caenorhabditis elegans, участвующее в межклеточном соединении» . Дж. Клеточная Биол . 134 (2): 537–548. дои : 10.1083/jcb.134.2.537 . ПМК 2120886 . ПМИД 8707836 .
- ^ Симонсен, Карина Т.; Моерман, Дональд Г.; Наус, Кристиан К. (2014). «Щелевые контакты у C. elegans» . Границы в физиологии . 5 : 40. дои : 10.3389/fphys.2014.00040 . ПМК 3920094 . ПМИД 24575048 .
- ^ Барбе, Монтана (1 апреля 2006 г.). «Клеточно-клеточная связь за пределами коннексинов: каналы паннексина». Физиология . 21 (2): 103–114. дои : 10.1152/физиол.00048.2005 . ПМИД 16565476 .
- ^ Панчина, Юрий; Кельмансон, Илья; Мац, Михаил; Лукьянов Константин; Усман, Наталья; Лукьянов, Сергей (июнь 2000 г.). «Вездесущее семейство предполагаемых молекул щелевых соединений» . Современная биология . 10 (13): Р473–Р474. Бибкод : 2000CBio...10.R473P . дои : 10.1016/S0960-9822(00)00576-5 . ПМИД 10898987 . S2CID 20001454 .
- ^ Ломан, Александр В.; Исаксон, Брант Э. (2014). «Дифференциация полуканалов коннексина и каналов паннексина при высвобождении клеточного АТФ» . Письма ФЭБС . 588 (8): 1379–1388. Бибкод : 2014FEBSL.588.1379L . дои : 10.1016/j.febslet.2014.02.004 . ПМЦ 3996918 . ПМИД 24548565 .
- ^ Сосинский Г.Е., Боасса Д., Дермицель Р., Даффи Х.С., Лэрд Д.В., Маквикар Б., Наус CC, Пенуэла С., Сцемс Е, Спрей Д.С., Томпсон Р.Дж., Чжао Х., Даль Г. (01.05.2011). «Каналы паннексина не являются полуканалами щелевого соединения» . Каналы . 5 (3): 193–197. дои : 10.4161/chan.5.3.15765 . ПМК 3704572 . ПМИД 21532340 .
- ^ Сливко-Колчик, Георгий А. (26 февраля 2019 г.). «Существуют ли щелевые соединения без коннексинов или паннексинов?» . БМК Экол. Эвол . 19 (Приложение 1). 46. Бибкод : 2019BMCEE..19S..46S . дои : 10.1186/s12862-019-1369-4 . ПМК 6391747 . ПМИД 30813901 .
- ↑ Перейти обратно: Перейти обратно: а б Эрве, JC; Бурмейстер, Н; Сарруй, Д; Даффи, HS (май 2007 г.). «Комплексы щелевых соединений: от партнеров к функциям». Прога Биофиз Мол Биол . 94 (1–2): 29–65. doi : 10.1016/j.pbiomolbio.2007.03.010 . ПМИД 17507078 .
- ^ Се, CL; Кумар, Нью-Мексико; Гилула, Северная Каролина; Франке, Ю (март 1991 г.). «Распределение генов белков мембранных каналов щелевых соединений на хромосомах человека и мыши». Соматическая клетка и молекулярная генетика . 17 (2): 191–200. дои : 10.1007/bf01232976 . ПМИД 1849321 . S2CID 44622463 .
- ^ Кумар, Нью-Мексико; Гилула, Северная Каролина (февраль 1992 г.). «Молекулярная биология и генетика каналов щелевых соединений». Семинары по клеточной биологии . 3 (1): 3–16. дои : 10.1016/s1043-4682(10)80003-0 . ПМИД 1320430 .
- ^ Крен, БТ; Кумар, Нью-Мексико; Ван, SQ; Гилула, Северная Каролина; Стир, CJ (ноябрь 1993 г.). «Дифференциальная регуляция транскриптов и белков множественных щелевых соединений во время регенерации печени крыс» . Журнал клеточной биологии . 123 (3): 707–18. дои : 10.1083/jcb.123.3.707 . ПМК 2200133 . ПМИД 8227133 .
- ^ Осима, А; Тани, К; Фудзиеси, Ю. (1 декабря 2016 г.). «Атомная структура канала щелевого соединения иннексина-6, определенная с помощью криоЭМ» . Нат. Коммун . 7 : 13681. Бибкод : 2016NatCo...713681O . дои : 10.1038/ncomms13681 . ПМК 5146279 . ПМИД 27905396 .
- ↑ Перейти обратно: Перейти обратно: а б Лауф, Ундина; Гипманс, Бен Н.Г.; Лопес, Патрисия; Браконно, Себастьен; Чен, Шу-Чи; Фальк, Матиас М. (6 августа 2002 г.). «Динамический транспорт и доставка коннексонов к плазматической мембране и прирастание к щелевым контактам в живых клетках» . Труды Национальной академии наук . 99 (16): 10446–10451. Бибкод : 2002PNAS...9910446L . дои : 10.1073/pnas.162055899 . ПМК 124935 . ПМИД 12149451 .
- ^ Чанг, Цин; Тан, Вэньсюэ; Ахмад, Шуб; Чжоу, Биньфэй; Линь, Си (2008). Шиффманн, Рафаэль (ред.). «Межклеточный перенос метаболитов в улитке, опосредованный щелевыми соединениями, нарушен у мышей с нулевым содержанием коннексина 30» . ПЛОС ОДИН . 3 (12). е4088. Бибкод : 2008PLoSO...3.4088C . дои : 10.1371/journal.pone.0004088 . ПМК 2605248 . ПМИД 19116647 .
- ^ Альбертс, Брюс (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . [ нужна страница ]
- ^ Ху Х, Даль Г (1999). «Обмен проводимостью и воротными свойствами между полуканалами щелевого перехода» . ФЭБС Летт . 451 (2): 113–117. Бибкод : 1999FEBSL.451..113H . дои : 10.1016/S0014-5793(99)00558-X . ПМИД 10371149 . S2CID 19289550 .
- ^ Левенштейн В.Р. (июль 1966 г.). «Проницаемость мембранных соединений». Энн. Н-Й акад. Наука . 137 (2): 441–472. Бибкод : 1966NYASA.137..441L . дои : 10.1111/j.1749-6632.1966.tb50175.x . ПМИД 5229810 . S2CID 22820528 .
- ^ Хан, Али К.; Ягельницкий, Мацей; Беннетт, Брэд К.; Перди, Майкл Д.; Йегер, Марк (сентябрь 2021 г.). «Крио-ЭМ структура открытой конформации полуканала щелевого соединения в липидных бислойных нанодисках» . Структура . 29 (9): 1040–1047.e3. дои : 10.1016/j.str.2021.05.010 . ПМЦ 9616683 . ПМИД 34129834 .
- ^ Гуденаф, Дэниел А.; Пол, Дэвид Л. (апрель 2003 г.). «За разрывом: функции непарных каналов связи». Nature Reviews Молекулярно-клеточная биология . 4 (4): 285–295. дои : 10.1038/nrm1072 . ПМИД 12671651 . S2CID 18103080 .
- ^ Лэрд, Дейл В.; Лампе, Пол Д. (декабрь 2018 г.). «Терапевтические стратегии, нацеленные на коннексины» . Nature Reviews Открытие лекарств . 17 (12): 905–921. дои : 10.1038/nrd.2018.138 . ПМК 6461534 . ПМИД 30310236 .
- ↑ Перейти обратно: Перейти обратно: а б с Ван Кампенхаут, Раф; Гомес, Ана Рита; Де Грооф, Тимо ВМ; Мюлдерманс, Серж; Девогдт, Ник; Винкен, Матье (28 марта 2021 г.). «Механизмы, лежащие в основе активации полуканалов коннексина при заболеваниях» . Международный журнал молекулярных наук . 22 (7):3503 дои : 10.3390/ijms22073503 . ПМК 8036530 . ПМИД 33800706 .
- ^ Орси Л., Малаисс-Лаге Ф., Амхердт М. и др. (ноябрь 1975 г.). «Клеточные контакты в человеческих островках Лангерганса» . Дж. Клин. Эндокринол. Метаб . 41 (5): 841–4. doi : 10.1210/jcem-41-5-841 . ПМИД 1102552 . [ постоянная мертвая ссылка ]
- ^ Гарфилд, RE; Симс, С.М.; Каннан, MS; Дэниел, Э.Э. (ноябрь 1978 г.). «Возможная роль щелевых контактов в активации миометрия во время родов». Являюсь. Дж. Физиол . 235 (5): C168–79. дои : 10.1152/ajpcell.1978.235.5.C168 . ПМИД 727239 . S2CID 31610495 .
- ^ Гуденаф, Д.А. (ноябрь 1979 г.). «Щелевые соединения хрусталика: структурная гипотеза нерегулируемых межклеточных путей с низким сопротивлением». Инвестируйте. Офтальмол. Вис. Наука . 18 (11): 1104–22. ПМИД 511455 .
- ^ Friend DS, Gilula NB (июнь 1972 г.). «Вариации плотных и щелевых соединений в тканях млекопитающих» . Дж. Клеточная Биол . 53 (3): 758–76. дои : 10.1083/jcb.53.3.758 . ПМК 2108762 . ПМИД 4337577 .
- ^ МакГинли Д., Посалаки З., Провазник М. (октябрь 1977 г.). «Межклеточные соединительные комплексы семенных канальцев крысы: исследование замораживания-перелома». Анат. Рек . 189 (2): 211–31. дои : 10.1002/ar.1091890208 . ПМИД 911045 . S2CID 19611753 .
- ^ Крейцигер ГО (сентябрь 1976 г.). «Морфология латеральной мембраны и структура щелевых соединений в эндотелии роговицы кролика». Эксп. Глазное разрешение . 23 (3): 285–93. дои : 10.1016/0014-4835(76)90129-9 . ПМИД 976372 .
- ^ Альбертини, DF; Андерсон, Э. (октябрь 1974 г.). «Внешний вид и структура межклеточных связей в онтогенезе фолликула яичника кролика с особым учетом щелевых соединений» . J Клеточная Биол . 63 (1): 234–50. дои : 10.1083/jcb.63.1.234 . ПМК 2109337 . ПМИД 4417791 .
- ^ Пруткин Л. (февраль 1975 г.). «Слизистая метаплазия и щелевые контакты в опухоли кожи, обработанной кислотой витамина А, кератоакантоме» . Рак Рез . 35 (2): 364–9. ПМИД 1109802 .
- ^ Равиола, Э; Гилула, Северная Каролина (июнь 1973 г.). «Щелевые контакты между фоторецепторными клетками сетчатки позвоночных» . Proc Natl Acad Sci США . 70 (6): 1677–81. Бибкод : 1973PNAS...70.1677R . дои : 10.1073/pnas.70.6.1677 . ПМЦ 433571 . ПМИД 4198274 .
- ^ Беллэрс, Р; Бретнах, А.С.; Гросс, М. (сентябрь 1975 г.). «Репликация соединительных комплексов замораживанием-переломом в неинкубированных и инкубированных куриных эмбрионах». Ресурсы клеточных тканей . 162 (2): 235–52. дои : 10.1007/BF00209209 . ПМИД 1237352 . S2CID 38441429 .
- ↑ Перейти обратно: Перейти обратно: а б Дж. Клеточная Биол. Июль 1974;62(1) 32-47. Сборка щелевых контактов во время нейруляции амфибий. Декер Р.С., Френд Д.С.
- ^ Ленц Т.Л., Тринкаус Дж.П. (март 1971 г.). «Дифференциация соединительного комплекса поверхностных клеток в развивающейся бластодерме Fundulus» . Дж. Клеточная Биол . 48 (3): 455–72. дои : 10.1083/jcb.48.3.455 . ПМК 2108114 . ПМИД 5545331 .
- ↑ Перейти обратно: Перейти обратно: а б Робертсон, доктор юридических наук (февраль 1953 г.). «Ультраструктура двух синапсов беспозвоночных». Труды Общества экспериментальной биологии и медицины . 82 (2): 219–23. дои : 10.3181/00379727-82-20071 . ПМИД 13037850 . S2CID 39294652 .
- ^ Сибата, Ю; Ямамото, Т. (март 1977 г.). «Щелевые контакты в клетках сердечной мышцы миноги». Ресурсы клеточных тканей . 178 (4): 477–82. дои : 10.1007/BF00219569 . ПМИД 870202 . S2CID 21426059 .
- ^ Лорбер, В; Рейнс, Д.Г. (апрель 1977 г.). «Тонкая структура щелевого перехода в оболочнике сердца». Ресурсы клеточных тканей . 179 (2): 169–75. дои : 10.1007/BF00219794 . ПМИД 858161 . S2CID 21604678 .
- ^ Хама К., Сайто К. (февраль 1977 г.). «Щелевые соединения между поддерживающими клетками в некоторых акусто-вестибулярных рецепторах». Дж. Нейроцитол . 6 (1): 1–12. дои : 10.1007/BF01175410 . ПМИД 839246 . S2CID 30090247 .
- ^ Хадспет, Эй Джей; Ревель, JP (июль 1971 г.). «Сосуществование щелей и перегородок в эпителии беспозвоночных» . Дж. Клеточная Биол . 50 (1): 92–101. дои : 10.1083/jcb.50.1.92 . ПМК 2108432 . ПМИД 5563454 .
- ^ Буко-Каму, Ева (1980). «Соединительные структуры пищеварительного эпителия головоногих моллюсков». Тканевая клетка . 12 (2): 395–404. дои : 10.1016/0040-8166(80)90013-0 . ПМИД 7414602 .
- ↑ Перейти обратно: Перейти обратно: а б Хэнд, АР; Гобель, С. (февраль 1972 г.). «Структурная организация перегородок и щелевых соединений Гидры» . Дж. Клеточная Биол . 52 (2): 397–408. дои : 10.1083/jcb.52.2.397 . ПМК 2108629 . ПМИД 4109925 .
- ^ Бервальд Р.Дж. (1975). «Перевернутая щель и другие клеточные соединения в капсулах гемоцитов таракана: исследование тонкого среза и замораживания-перелома». Тканевая клетка . 7 (3): 575–85. дои : 10.1016/0040-8166(75)90027-0 . ПМИД 1179417 .
- ^ Джонсон Р., Хаммер М., Шеридан Дж., Ревел Дж. П. (ноябрь 1974 г.). «Формирование щелевого соединения между реагрегированными клетками гепатомы Новикова» . Учеб. Натл. акад. наук. США . 71 (11): 4536–40. Бибкод : 1974PNAS...71.4536J . дои : 10.1073/pnas.71.11.4536 . ПМК 433922 . ПМИД 4373716 .
- ^ Кнудсен, Калифорния; Хорвиц, А.Ф. (1978). «К механизму слияния миобластов». Прог Клин Биол Рес . 23 : 563–8. ПМИД 96453 .
- ^ Джонс С.Дж., Грей С., Сакамаки Х. и др. (апрель 1993 г.). «Частота и размер щелевых соединений между костными клетками черепа крысы». Анат. Эмбриол . 187 (4): 343–52. дои : 10.1007/BF00185892 . ПМИД 8390141 . S2CID 33191311 .
- ^ Сперелакис, Николас; Рамасами, Лакшминараянан (2005). «Каналы щелевого соединения тормозят поперечное распространение в сердечной мышце» . Биомед Рус Онлайн . 4 (1): 7. дои : 10.1186/1475-925X-4-7 . ПМК 549032 . ПМИД 15679888 .
- ^ Ларсен В.Дж., Азарния Р., Левенштейн В.Р. (июнь 1977 г.). «Межклеточная коммуникация и рост тканей: IX. Соединительная мембранная структура гибридов коммуникационно-компетентных и коммуникативно-некомпетентных клеток». Дж. Член. Биол . 34 (1): 39–54. дои : 10.1007/BF01870292 . ПМИД 561191 . S2CID 2831462 .
- ^ Корсаро CM, Migeon BR (октябрь 1977 г.). «Сравнение контактно-опосредованной коммуникации в нормальных и трансформированных клетках человека в культуре» . Учеб. Натл. акад. наук. США . 74 (10): 4476–80. Бибкод : 1977PNAS...74.4476C . дои : 10.1073/pnas.74.10.4476 . ПМК 431966 . ПМИД 270694 .
- ^ Хаберманн, Х; Чанг, Вайоминг; Береза, Л; Мехта, П; Принс, Г.С. (январь 2001 г.). «Воздействие эстрогенов на развитие изменяет адгезию эпителиальных клеток и белки щелевых соединений в предстательной железе взрослых крыс» . Эндокринология . 142 (1): 359–69. дои : 10.1210/endo.142.1.7893 . ПМИД 11145599 .
- ^ Келли, Роберт О.; Фогель, Кэтрин Г.; Криссман, Гарри А.; Лухан, Кристофер Дж.; Шкипер, Бетти Э. (март 1979 г.). «Развитие стареющей клеточной поверхности. Уменьшение метаболического сотрудничества, опосредованного щелевыми соединениями, с прогрессивным субкультивированием фибробластов эмбриона человека (IMR-90)». Эксп. Сотовый Res . 119 (1): 127–43. дои : 10.1016/0014-4827(79)90342-2 . ПМИД 761600 .
- ^ Фелан, Полина (июнь 2005 г.). «Иннексины: члены эволюционно консервативного семейства белков щелевых соединений» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1711 (2): 225–245. дои : 10.1016/j.bbamem.2004.10.004 . ПМИД 15921654 .
- ^ Эрве, Жан-Клод; Фелан, Полина; Бруззоне, Роберто; Уайт, Томас В. (декабрь 2005 г.). «Коннексины, иннексины и паннексины: преодоление коммуникационного разрыва» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1719 (1–2): 3–5. дои : 10.1016/j.bbamem.2005.11.013 . ПМИД 16359939 .
- ^ Гуиса, Хуан; Гарсия, Анибал; Арриагада, Хавьера; Гутьеррес, Камила; Гонсалес, Хорхе; Маркес-Миранда, Валерия; Алегрия-Аркос, Мелисса; Дуарте, Йорли; Рохас, Максимилиано; Гонсалес-Нило, Фернандо; Саес, Хуан К.; Вега, Хосе Л. (февраль 2022 г.). «Уннексины: гомологи белков-иннексинов у паразитов Trypanosomatidae». Журнал клеточной физиологии . 237 (2): 1547–1560. дои : 10.1002/jcp.30626 . ПМИД 34779505 . S2CID 244116450 .
- ^ Тернбулл, Мэтью В.; Волков, Анн-Натали; Уэбб, Брюс А.; Фелан, Полина (июль 2005 г.). «Гены функциональных щелевых соединений кодируются вирусами насекомых» . Современная биология . 15 (13): Р491–Р492. Бибкод : 2005CBio...15.R491T . дои : 10.1016/j.cub.2005.06.052 . ПМИД 16005277 .
- ^ Мороз Леонид Л.; Романова, Дарья Ю. (23 декабря 2022 г.). «Альтернативные нейронные системы: Что такое нейрон? (гребневики, губки и плакозойи)» . Границы клеточной биологии и биологии развития . 10 : 1071961. дои : 10.3389/fcell.2022.1071961 . ПМЦ 9816575 . ПМИД 36619868 .
- ^ Бергквист, PR; Грин, ЧР (1977). «Ультраструктурное исследование расселения и метаморфоза личинок губок». Да. Биол. Мар . 18 : 289–302.
- ^ Грин, ЧР; Бергквист, PR (1979). «Специализации клеточных мембран Porifera». Coll Int Cent Natn Res Scient . 291 : 153–158.
- ^ Уорнер, Энн Э.; Гатри, Сара С.; Джилула, Нортон Б. (1984). «Антитела к белку щелевых соединений избирательно нарушают связь между соединениями у ранних эмбрионов амфибий». Природа . 311 (5982): 127–31. Бибкод : 1984Natur.311..127W . дои : 10.1038/311127a0 . ПМИД 6088995 . S2CID 2620476 .
- ^ Уорнер, А.Е. (2007). «Использование антител к белку с щелевыми соединениями для изучения роли связи через щелевые соединения во время развития». Симпозиум 125 Фонда Ciba - Соединительные комплексы эпителиальных клеток . Симпозиумы Фонда Новартис. Том. 125. стр. 154–67. дои : 10.1002/9780470513408.ch10 . ISBN 9780470513408 . ПМИД 3030673 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Бастид, Б; Жарри-Гишар, Т; Бриан, JP; Делез, Дж; Грос, Д. (апрель 1996 г.). «Влияние антипептидных антител, направленных против трех доменов коннексина 43, на проницаемость щелевых соединений культивируемых клеток сердца». Дж. Член. Биол . 150 (3): 243–53. дои : 10.1007/s002329900048 . ПМИД 8661989 . S2CID 20408672 .
- ^ Хофер, А; Дермицель, Р. (сентябрь 1998 г.). «Визуализация и функциональная блокировка полуканалов щелевых соединений (коннексонов) с помощью антител против доменов внешней петли в астроцитах». Глия . 24 (1): 141–54. doi : 10.1002/(SICI)1098-1136(199809)24:1<141::AID-GLIA13>3.0.CO;2-R . ПМИД 9700496 . S2CID 23234120 .
- ↑ Перейти обратно: Перейти обратно: а б с Фрэнсис Р., Сюй Х, Парк Х и др. (2011). Бранднер Дж. М. (ред.). «Коннексин43 модулирует полярность клеток и направленную миграцию клеток, регулируя динамику микротрубочек» . ПЛОС ОДИН . 6 (10): e26379. Бибкод : 2011PLoSO...626379F . дои : 10.1371/journal.pone.0026379 . ПМК 3194834 . ПМИД 22022608 .
- ^ Левин, Майкл; Меркола, Марк (ноябрь 1998 г.). «Щелевые соединения участвуют в раннем возникновении лево-правой асимметрии». Дев. Биол . 203 (1): 90–105. CiteSeerX 10.1.1.137.4340 . дои : 10.1006/dbio.1998.9024 . ПМИД 9806775 .
- ^ Левин, М; Меркола, М. (ноябрь 1999 г.). «Передача сигналов формирования левого и правого паттернов, опосредованная щелевыми соединениями, в ранней бластодерме цыпленка находится выше асимметрии Shh в узле». Разработка . 126 (21): 4703–14. дои : 10.1242/dev.126.21.4703 . ПМИД 10518488 .
- ^ Бани-Ягуб, Махмуд; Андерхилл, Т. Майкл; Наус, Кристиан К.Г. (1999). «Блокировка щелевого соединения препятствует нейрональной и астроглиальной дифференцировке клеток эмбриональной карциномы мыши P19». Дев. Жене . 24 (1–2): 69–81. doi : 10.1002/(SICI)1520-6408(1999)24:1/2<69::AID-DVG8>3.0.CO;2-M . ПМИД 10079512 .
- ^ Бани-Ягуб, Махмуд; Бехбергер, Джон Ф.; Андерхилл, Т. Майкл; Наус, Кристиан К.Г. (март 1999 г.). «Влияние блокировки щелевых соединений на дифференцировку нейронов клеток NTera2/клона D1 человека». Эксп. Нейрол . 156 (1): 16–32. doi : 10.1006/exnr.1998.6950 . ПМИД 10192774 . S2CID 41420671 .
- ^ Донахью, HJ; Ли, З; Чжоу, Z; Йеллоули, CE (февраль 2000 г.). «Дифференциация остеобластических клеток плода человека и межклеточная связь по щелевым соединениям». Являюсь. J. Physiol., Cell Physiol . 278 (2): C315–22. дои : 10.1152/ajpcell.2000.278.2.C315 . ПМИД 10666026 . S2CID 9894657 .
- ^ Кронье, Л; Френдо, JL; Клевета, Н; Пиду, Г; Бертен, Дж; Гибурденш, Ж; Пуэнтис, Дж; Малассин, А. (ноябрь 2003 г.). «Необходимость межклеточной коммуникации через щелевые соединения для дифференцировки ворсинчатого трофобласта человека» . Биол. Репродукция . 69 (5): 1472–80. дои : 10.1095/biolreprod.103.016360 . ПМИД 12826585 .
- ^ Эль-Саббан, Мэн; Сфейр, Эй Джей; Дахер, Миннесота; Калаани, штат Нью-Йорк; Басам, РА; Талхук, Р.С. (сентябрь 2003 г.). «Коммуникация через щелевые соединения, индуцированная ЕСМ, усиливает дифференцировку эпителиальных клеток молочной железы». Дж. Клеточная наука . 116 (Часть 17): 3531–41. дои : 10.1242/jcs.00656 . ПМИД 12893812 . S2CID 5057466 .
- ^ Чайтор, АТ; Мартин, ЧП; Эванс, Вашингтон; Рэндалл, доктор медицины; Гриффит, ТМ (октябрь 1999 г.). «Эндотелиальный компонент вызванного каннабиноидами расслабления в брыжеечной артерии кролика зависит от связи в щелевых соединениях» . Дж. Физиол . 520 (2): 539–50. дои : 10.1111/j.1469-7793.1999.00539.x . ПМК 2269589 . ПМИД 10523421 .
- ^ Шринивас, М; Хопперстад, Миннесота; Спрей, округ Колумбия (сентябрь 2001 г.). «Хинин блокирует определенные подтипы каналов щелевого соединения» . Учеб. Натл. акад. наук. США . 98 (19): 10942–7. Бибкод : 2001PNAS...9810942S . дои : 10.1073/pnas.191206198 . ПМЦ 58578 . ПМИД 11535816 .
- ^ Ли Би, Ван; Парысек, Линда М.; Варник, Рональд; Стэмбрук, Питер Дж. (декабрь 1993 г.). «Доказательства in vitro, что метаболическое сотрудничество ответственно за эффект свидетеля, наблюдаемый при ретровирусной генной терапии ВПГ». Хм. Джин Тер . 4 (6): 725–31. дои : 10.1089/hum.1993.4.6-725 . ПМИД 8186287 .
- ^ Питтс, JD (ноябрь 1994 г.). «Генная терапия рака: эффект свидетеля с использованием пути щелевых соединений». Мол. Карциног . 11 (3): 127–30. дои : 10.1002/mc.2940110302 . ПМИД 7945800 . S2CID 41136083 .
- ^ Коломбо, Бруно М.; Бенедетти, Сара; Оттоленги, Серджио; Мора, Марина; Полло, Бьянка; Поли, Джорджио; Финоккьяро, Гаэтано (июнь 1995 г.). «Эффект свидетеля»: связь гибели клеток U-87 с опосредованным ганцикловиром апоптозом близлежащих клеток и отсутствием эффекта у атимических мышей». Хм. Джин Тер . 6 (6): 763–72. дои : 10.1089/hum.1995.6.6-763 . ПМИД 7548276 .
- ^ Фик, Джеймс; Баркер, Фред Г.; Дазин, Пол; Вестфаль, Эйлин М.; Бейер, Эрик С.; Израиль, Марк А. (ноябрь 1995 г.). «Степень гетероклеточной коммуникации, опосредованной щелевыми соединениями, позволяет предсказать цитотоксичность опухоли-свидетеля in vitro» . Учеб. Натл. акад. наук. США . 92 (24): 11071–5. Бибкод : 1995PNAS...9211071F . дои : 10.1073/pnas.92.24.11071 . ПМК 40573 . ПМИД 7479939 .
- ^ Эльшами, А.А.; Сааведра, А; Чжан, Х; Кухарчук, Ю.К.; Спрей, постоянный ток; Фишман, Дж.И.; Амин, К.М.; Кайзер, ЛР; Альбельда, С.М. (январь 1996 г.). «Щелевые соединения играют роль в« эффекте свидетеля »системы тимидинкиназы/ганцикловира вируса простого герпеса in vitro». Джин Тер . 3 (1): 85–92. ПМИД 8929915 .
- ^ Мениль, Марк; Пикколи, Колетт; Тираби, Жерар; Виллеке, Клаус; Ямасаки, Хироши (март 1996 г.). «Свидетельное убийство раковых клеток геном тимидинкиназы вируса простого герпеса опосредовано коннексинами» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (5): 1831–5. Бибкод : 1996ПНАС...93.1831М . дои : 10.1073/pnas.93.5.1831 . ПМК 39867 . ПМИД 8700844 .
- ^ Синура, Нобусада; Чен, Линь; Вани, Максуд А.; Ким, Ён Гю; Ларсон, Джеффри Дж.; Уорник, Рональд Э.; Саймон, Матиас; Менон, Анил Г.; и др. (май 1996 г.). «Экспрессия белка и информационной РНК коннексина 43 в астроцитомах: значение генной терапии опухолей головного мозга». Дж. Нейрохирургия . 84 (5): 839–45, обсуждение 846. doi : 10.3171/jns.1996.84.5.0839 . ПМИД 8622159 .
- ^ Хамель, В; Магнелли, Л; Кьяруги, вице-президент; Израиль, Массачусетс (июнь 1996 г.). «Апоптозная гибель клеток-свидетелей, опосредованная тимидинкиназой/ганцикловиром вируса простого герпеса». Рак Рез . 56 (12): 2697–702. ПМИД 8665496 .
- ^ Сакко, Миннесота; Бенедетти, С; Дюфло-Танцовщик, А; Мениль, М; Баньяско, Л; Стрина, Д; Фазоло, В; Вилла, А; и др. (декабрь 1996 г.). «Частичная регрессия, но неполное уничтожение опухолей молочной железы у трансгенных мышей путем ретровирусно-опосредованной передачи HSVtk 'in vivo' ». Джин Тер . 3 (12): 1151–6. ПМИД 8986442 .
- ^ Риппс, Харрис (март 2002 г.). «Гибель клеток при пигментном ретините: щелевые контакты и эффект «свидетеля»». Эксп. Глазное разрешение . 74 (3): 327–36. дои : 10.1006/exer.2002.1155 . ПМИД 12014914 .
- ^ Литтл, Джей Би; Аззам, Э.И.; Де Толедо, СМ; Нагасава, Х (2002). «Эффекты свидетеля: межклеточная передача сигналов радиационного повреждения». Дозиметрия радиационной защиты . 99 (1–4): 159–62. doi : 10.1093/oxfordjournals.rpd.a006751 . ПМИД 12194273 .
- ^ Чжоу, Х; Рандерс-Персон, Г; Сузуки, М; Уолдрен, Калифорния; Привет, ТК (2002). «Генотоксическое повреждение необлученных клеток: вклад эффекта свидетеля». Дозиметрия радиационной защиты . 99 (1–4): 227–32. doi : 10.1093/oxfordjournals.rpd.a006769 . ПМИД 12194291 .
- ^ Лоримор, ЮАР; Райт, Э.Г. (январь 2003 г.). «Радиационная нестабильность генома и эффекты свидетеля: связанные реакции воспалительного типа на стресс и травмы, вызванные радиацией? Обзор». Межд. Дж. Радиат. Биол . 79 (1): 15–25. дои : 10.1080/0955300021000045664 . ПМИД 12556327 . S2CID 44821116 .
- ^ Эрлих, HP; Диз, Т. (2003). «Роль межклеточных коммуникаций щелевых соединений в заживлении ран». Регенерация восстановления ран . 11 (6): 481–9. дои : 10.1046/j.1524-475X.2003.11616.x . ПМИД 14617290 . S2CID 25113646 .
- ^ Коутиньо, П.; Цю, К.; Франк, С.; Ван, СМ; Браун, Т.; Грин, ЧР; Беккер, Д.Л. (июль 2005 г.). «Ограничение распространения ожога за счет временного ингибирования экспрессии коннексина 43 в месте повреждения» . Br J Plast Surg . 58 (5): 658–67. дои : 10.1016/j.bjps.2004.12.022 . ПМИД 15927148 .
- ^ Ван, СМ; Линкольн, Дж.; Кук, Дж. Э.; Беккер, Д.Л. (ноябрь 2007 г.). «Аномальная экспрессия коннексина лежит в основе замедленного заживления ран на диабетической коже» . Диабет . 56 (11): 2809–17. дои : 10.2337/db07-0613 . ПМИД 17717278 .
- ^ Ривера, ЕМ; Варгас, М; Рикс-Уильямсон, Л. (1997). «Соображения по эстетической реставрации эндодонтически леченных передних зубов после внутрикоронкового отбеливания». Практика пародонтологии Эстет Дент . 9 (1): 117–28. ПМИД 9550065 .
- ^ Мугишо, Одунайо О.; Арьял, Джьоти; Шорн, Авик; Лион, Хизер; Акоста, Моника Л.; Грин, Колин Р.; Рупенталь, Ильва Д. (15 февраля 2023 г.). «Блокатор полуканалов коннексина 43, принимаемый перорально, Тонаберсат, ингибирует сосудистый распад и активацию воспаления на мышиной модели диабетической ретинопатии» . Международный журнал молекулярных наук . 24 (4): 3876. doi : 10.3390/ijms24043876 . ПМЦ 9961562 . ПМИД 36835288 .
- ^ Кусато, К; Боско, А; Розенталь, Р; Гимарайнш, Калифорния; Риз, Бельгия; Линден, Р; Спрей, округ Колумбия (июль 2003 г.). «Щелевые соединения опосредуют гибель клеток-свидетелей в развивающейся сетчатке» . Дж. Нейроски . 23 (16): 6413–22. doi : 10.1523/JNEUROSCI.23-16-06413.2003 . ПМК 6740641 . ПМИД 12878681 .
- ^ Мойер, Куртис Э.; Саггерс, Грегори К.; Эрлих, Х. Пол (2004). «Тучные клетки способствуют сокращению решетки коллагена, населенного фибробластами, посредством межклеточной связи щелевых соединений». Регенерация восстановления ран . 12 (3): 269–75. дои : 10.1111/j.1067-1927.2004.012310.x . ПМИД 15225205 . S2CID 24363587 .
- ^ Джалилян, Арканзас; Макгоги, Д; Патель, С; Со, EY; Ян, С; Ченг, Дж; Томич, М; Синха, С; и др. (май 2006 г.). «Коннексин 26 регулирует эпидермальный барьер и ремоделирование ран, а также способствует псориазоформной реакции» . Дж. Клин. Инвестируйте . 116 (5): 1243–53. дои : 10.1172/JCI27186 . ПМК 1440704 . ПМИД 16628254 .
- ^ Чжан, Ю.; Ван, Х.; Ковач, А.; Кантер, Э.М.; Ямада, штат Калифорния (февраль 2010 г.). «Снижение экспрессии Cx43 ослабляет ремоделирование желудочков после инфаркта миокарда за счет нарушения передачи сигналов TGF-бета» . Являюсь. Дж. Физиол. Сердечный цирк. Физиол . 298 (2): H477–87. дои : 10.1152/ajpheart.00806.2009 . ПМЦ 2822575 . ПМИД 19966054 .
- ^ Эй Б., Эйкинг А., Геркен Г., Подольский Д.К., Карио Э. (август 2009 г.). «TLR2 опосредует межклеточную связь через щелевые соединения через коннексин-43 при повреждении эпителиального барьера кишечника» . Ж. Биол. Хим . 284 (33): 22332–43. дои : 10.1074/jbc.M901619200 . ПМК 2755956 . ПМИД 19528242 .
- ^ Сюй, Цзи; Николсон, Брюс Дж. (январь 2013 г.). «Роль коннексинов в физиологии ушей и кожи — функциональные выводы из мутаций, связанных с болезнями» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1828 (1): 167–178. дои : 10.1016/j.bbamem.2012.06.024 . ПМЦ 3521577 .
- ^ Шринивас, Мидутуру; Верселис, Витас К.; Уайт, Томас В. (1 января 2018 г.). «Заболевания человека, связанные с мутациями коннексина» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1860 (1): 192–201. дои : 10.1016/j.bbamem.2017.04.024 . ПМЦ 5659969 . ПМИД 28457858 .
- ^ Уайт, Томас В.; Пол, Дэвид Л. (1999). «Генетические заболевания и нокауты генов раскрывают разнообразные функции коннексина». Ежегодный обзор физиологии . 61 (1): 283–310. doi : 10.1146/annurev.phyol.61.1.283 . ПМИД 10099690 .
- ^ Иванович, Эна; Кучера, Ян П. (2 ноября 2022 г.). «Извитые сердечные вставочные диски модулируют эфаптическую связь» . Клетки . 11 (21): 3477. doi : 10.3390/cells11213477 . ПМЦ 9655400 . ПМИД 36359872 .
- ↑ Перейти обратно: Перейти обратно: а б Грюйтерс, WT; Кистлер, Дж.; Булливант, С. (1 октября 1987 г.). «Формирование, распределение и диссоциация межклеточных соединений в хрусталике». Журнал клеточной науки . 88 (3): 351–359. дои : 10.1242/jcs.88.3.351 . ПМИД 3448099 .
- ↑ Перейти обратно: Перейти обратно: а б с Грюйтерс, В. (2003). «Участвуют ли мембранные бляшки щелевых соединений в межклеточном переносе везикул?» . Международная клеточная биология . 27 (9): 711–717. Бибкод : 2019BMCEE..19S..46S . дои : 10.1186/s12862-019-1369-4 . ПМК 6391747 . ПМИД 30813901 .
- ^ Коннорс; Лонг (2004). «Электрические синапсы в мозгу млекопитающих» . Анну преподобный Neurosci . 27 : 393–418. дои : 10.1146/annurev.neuro.26.041002.131128 . ПМИД 15217338 .
- ^ Ортманн-Мерфи, Дженнифер Л.; Абрамс, Чарльз К.; Шерер, Стивен С. (май 2008 г.). «Щелевые соединения пары астроцитов и олигодендроцитов» . Журнал молекулярной нейронауки . 35 (1): 101–116. дои : 10.1007/s12031-007-9027-5 . ПМК 2650399 . ПМИД 18236012 .
- ^ Бела Вёлги; Стюарт А. Блумфилд (февраль 2009 г.). «Разнообразные функциональные роли и регуляция щелевых соединений нейронов сетчатки» . Обзоры природы Неврология . 10 (7): 495–506. дои : 10.1016/S0165-0173(99)00070-3 . ПМЦ 3381350 . ПМИД 19491906 .
- ^ Гарфилд, RE; Симс, С.М.; Каннан, MS; Дэниел, Э.Э. (ноябрь 1978 г.). «Возможная роль щелевых контактов в активации миометрия во время родов». Являюсь. Дж. Физиол . 235 (5): C168–79. doi:10.1152/ajpcell.1978.235.5.C168 . PMID 727239. S2CID [https://www.semanticscholar.org/paper/Possible-role-of-gap-junctions-in-activation-of-Garfield-Sims/ce78aa441efb2f15ff4749d082197ed4db1b2584 31610495
- ^ Борос-Рауш А., Шинлова О. и Лай С.Дж. (2021). «Ингибитор хемокинов широкого спектра действия блокирует индуцированные воспалением перекрестные помехи миоцитов и макрофагов миометрия и сокращение миометрия». Клетки . 11 (1): 128. doi: 10.3390/cells11010128 PMID 35011690
- ^ Эванс, В. Ховард; Де Вюйст, Эльке; Лейберт, Люк (1 июля 2006 г.). «Сотовый Интернет через щелевые соединения: полуканалы коннексина выходят в центр внимания» . Биохимический журнал . 397 (1): 1–14. дои : 10.1042/BJ20060175 . ПМЦ 1479757 . ПМИД 16761954 .
- ^ Робертсон, доктор юридических наук (1963). Локк, Майкл (ред.). Клеточные мембраны в развитии . Нью-Йорк: Академическая пресса. OCLC 261587041 . [ нужна страница ]
- ^ Робертсон (1981). «Мембранная структура» . Журнал клеточной биологии . 91 (3): 189–204. дои : 10.1083/jcb.91.3.189s . JSTOR 1609517 . ПМК 2112820 . ПМИД 7033238 .
- ^ Фуршпан Э.Дж., Поттер Д.Д. (1957). «Механизм передачи нервного импульса в синапсах рака». Природа . 180 (4581): 342–343. Бибкод : 1957Natur.180..342F . дои : 10.1038/180342a0 . ПМИД 13464833 . S2CID 4216387 .
- ^ Фуршпан Э.Дж., Поттер Д.Д. (1959). «Передача в гигантских моторных синапсах рака» . Журнал физиологии . 145 (2): 289–325. дои : 10.1113/jphysicalol.1959.sp006143 . ПМК 1356828 . ПМИД 13642302 .
- ^ Пэйтон, BW; Беннетт, МВЛ; Паппас, Джорджия (декабрь 1969 г.). «Проницаемость и структура соединительных мембран электротонического синапса». Наука . 166 (3913): 1641–1643. Бибкод : 1969Sci...166.1641P . дои : 10.1126/science.166.3913.1641 . ПМИД 5360587 . S2CID 24701801 .
- ^ Чалкрофт, JP; Булливант, С. (октябрь 1970 г.). «Интерпретация клеточной мембраны печени и структуры соединения на основе наблюдения копий замораживания-перелома обеих сторон перелома» . Дж. Клеточная Биол . 47 (1): 49–60. дои : 10.1083/jcb.47.1.49 . ПМК 2108397 . ПМИД 4935338 .
- ^ Пераккья, C (апрель 1973 г.). «Соединения с низким сопротивлением у раков. II. Структурные детали и дополнительные доказательства существования межклеточных каналов с помощью замораживания-перелома и отрицательного окрашивания» . Дж. Клеточная Биол . 57 (1): 54–65. дои : 10.1083/jcb.57.1.54 . ПМК 2108965 . ПМИД 4120610 .
- ^ Ислам, М.; Дас, С.; Эмин, М.; и др. (2012). «Перенос митохондрий из стромальных клеток костномозгового происхождения в легочные альвеолы защищает от острого повреждения легких» . Нат Мед . 18 (5): 759–765. дои : 10.1038/нм.2736 . ПМЦ 3727429 . ПМИД 22504485 .
- ^ Тодд К.Л., Кристан ВБ, французский КА (ноябрь 2010 г.). «Экспрессия щелевых соединений необходима для нормального образования химических синапсов» . Дж. Нейроски . 30 (45): 15277–85. doi : 10.1523/JNEUROSCI.2331-10.2010 . ПМЦ 3478946 . ПМИД 21068332 .
- ^ Гуденаф, окружной прокурор; Стокениус, В. (1972). «Выделение щелевых соединений гепатоцитов мыши: предварительная химическая характеристика и дифракция рентгеновских лучей» . Журнал клеточной биологии . 54 (3): 646–56. дои : 10.1083/jcb.54.3.646 . ПМК 2200277 . ПМИД 4339819 .
- ^ Гуденаф, Д.А. (1974). «Объемная изоляция щелевых соединений гепатоцитов мыши: характеристика основного белка, коннексина» . Журнал клеточной биологии . 61 (2): 557–63. дои : 10.1083/jcb.61.2.557 . ПМК 2109294 . ПМИД 4363961 .
- ^ Кумар, Нью-Мексико; Гилула, НБ (1986). «Клонирование и характеристика кДНК печени человека и крысы, кодирующих белок щелевого соединения» . Журнал клеточной биологии . 103 (3): 767–76. дои : 10.1083/jcb.103.3.767 . ПМК 2114303 . ПМИД 2875078 .
- ^ МакНатт Н.С., Вайнштейн Р.С. (декабрь 1970 г.). «Ультраструктура связи. Коррелированное исследование тонкого среза и замораживания-раскола» . Дж. Клеточная Биол . 47 (3): 666–88. дои : 10.1083/jcb.47.3.666 . ПМК 2108148 . ПМИД 5531667 .
- ^ Чалкрофт, JP; Булливант, С. (1970). «Интерпретация клеточной мембраны печени и структуры соединения на основе наблюдения копий замораживания-перелома обеих сторон перелома» . Журнал клеточной биологии . 47 (1): 49–60. дои : 10.1083/jcb.47.1.49 . ПМК 2108397 . ПМИД 4935338 .
- ^ Молодой; Кон, З.А.; Гилула, НБ (1987). «Функциональная сборка проводимости щелевого соединения в липидных бислоях: демонстрация того, что основной белок массой 27 кД образует соединительный канал». Клетка . 48 (5): 733–43. дои : 10.1016/0092-8674(87)90071-7 . ПМИД 3815522 . S2CID 39342230 .
- ^ Николсон Б.Дж., Грос Д.Б., Кент С.Б., Худ Л.Е., Ревель Дж.П. (1985). «Белки щелевых соединений Mr 28 000 из сердца и печени крысы разные, но родственные» . Журнал биологической химии . 260 (11): 6514–6517. дои : 10.1016/S0021-9258(18)88810-X . ПМИД 2987225 .
- ^ Бейер ЕС, Пол Д.Л., Гуденаф Д.А. (1987). «Коннексин43: белок сердца крысы, гомологичный белку щелевого соединения печени» . Журнал клеточной биологии . 105 (6, ч. 1): 2621–2629. дои : 10.1083/jcb.105.6.2621 . ПМК 2114703 . ПМИД 2826492 .
- ^ Кистлер Дж., Киркланд Б., Булливант С. (1985). «Идентификация белка массой 70 000 D в соединительных доменах мембраны хрусталика» . Журнал клеточной биологии . 101 (1): 28–35. дои : 10.1083/jcb.101.1.28 . ПМК 2113615 . ПМИД 3891760 .
- ^ Стехелин Л.А. (май 1972 г.). «Три типа щелевых соединений, соединяющих эпителиальные клетки кишечника, визуализированные методом лиофилизации» . Учеб. Натл. акад. наук. США . 69 (5): 1318–21. Бибкод : 1972PNAS...69.1318S . дои : 10.1073/pnas.69.5.1318 . ПМК 426690 . ПМИД 4504340 .
- ^ Грюйтерс, WTM; Кистлер, Дж; Булливант, С; Гуденаф, Д.А. (1987). «Иммунолокализация MP70 в межклеточных соединениях волокон хрусталика длиной 16-17 нм» . Журнал клеточной биологии . 104 (3): 565–72. дои : 10.1083/jcb.104.3.565 . ПМК 2114558 . ПМИД 3818793 .
- ^ Робертсон, доктор юридических наук (октябрь 1963 г.). «Возникновение структуры субъединиц в мембранах единиц клубных окончаний в синапсах клеток Маутнера в мозге золотых рыбок» . Дж. Клеточная Биол . 19 (1): 201–21. дои : 10.1083/jcb.19.1.201 . ПМК 2106854 . ПМИД 14069795 .
- ^ Левенштейн В.Р., Канно Ю. (сентябрь 1964 г.). «Исследование эпителиальных (железистых) клеток. I. Изменения проницаемости поверхностных мембран» . Дж. Клеточная Биол . 22 (3): 565–86. дои : 10.1083/jcb.22.3.565 . ПМК 2106478 . ПМИД 14206423 .
- ↑ Перейти обратно: Перейти обратно: а б Озато-Сакурай Н., Фудзита А., Фудзимото Т. (2011). Вонг Н.С. (ред.). «Распределение фосфатидилинозитол-4,5-бисфосфата в ацинарных клетках поджелудочной железы крыс, выявленное с помощью метода мечения реплик замораживания-перелома» . ПЛОС ОДИН . 6 (8): e23567. Бибкод : 2011PLoSO...623567O . дои : 10.1371/journal.pone.0023567 . ПМК 3156236 . ПМИД 21858170 .
- ^ Штраус, Р.Э.; Гурди, Р.Г. (декабрь 2020 г.). «Cx43 и актиновый цитоскелет: новые роли и последствия для регуляции барьерной функции на основе межклеточных соединений» . Биомолекулы . 10 (12): 1656. doi : 10.3390/biom10121656 . ПМЦ 7764618 . ПМИД 33321985 .
- ^ Декер, Р.С.; Френд, DS (июль 1974 г.). «Сборка щелевых контактов во время нейруляции амфибий» . Дж. Клеточная Биол . 62 (1): 32–47. дои : 10.1083/jcb.62.1.32 . ПМК 2109180 . ПМИД 4135001 .
- ^ Декер, Р.С. (июнь 1976 г.). «Гормональная регуляция дифференцировки щелевых соединений» . Дж. Клеточная Биол . 69 (3): 669–85. дои : 10.1083/jcb.69.3.669 . ПМК 2109697 . ПМИД 1083855 .
- ^ Лауф У., Гипманс Б.Н., Лопес П., Браконнот С., Чен С.К., Фальк М.М. (август 2002 г.). «Динамический транспорт и доставка коннексонов к плазматической мембране и прирастание к щелевым контактам в живых клетках» . Учеб. Натл. акад. наук. США . 99 (16): 10446–51. Бибкод : 2002PNAS...9910446L . дои : 10.1073/pnas.162055899 . ПМК 124935 . ПМИД 12149451 .
- ^ Мейер, Р; Малевич, Б; Бауманн, WJ; Джонсон, Р.Г. (июнь 1990 г.). «Увеличение сборки щелевых соединений между культивируемыми клетками при добавлении холестерина». Дж. Клеточная наука . 96 (2): 231–8. дои : 10.1242/jcs.96.2.231 . ПМИД 1698798 .
- ^ Джонсон, Р.Г.; Рейнхаут, Дж. К.; Тенбрук, EM; Куэйд, Би Джей; Ясумура, Т.; Дэвидсон, КГВ; Шеридан, доктор медицинских наук; Раш, Дж. Э. (январь 2012 г.). «Сборка щелевых соединений: роль в формировании бляшек и регуляция С-конца коннексина 43» . Мол. Биол. Клетка . 23 (1): 71–86. дои : 10.1091/mbc.E11-02-0141 . ПМК 3248906 . ПМИД 22049024 .
- ^ Локк, Даррен; Харрис, Эндрю Л. (2009). «Каналы коннексина и фосфолипиды: ассоциация и модуляция» . БМК Биол . 7 (1): 52. дои : 10.1186/1741-7007-7-52 . ПМЦ 2733891 . ПМИД 19686581 .
- ↑ Перейти обратно: Перейти обратно: а б Ли X, Камасава Н., Чилофан С. и др. (сентябрь 2008 г.). «Содержащие коннексин45 щелевые контакты нейронов в сетчатке грызунов также содержат коннексин36 в обеих прилегающих гемибляшках, образуя бигомотипические щелевые соединения, а каркас обеспечивает zonula occludens-1» . Дж. Нейроски . 28 (39): 9769–89. doi : 10.1523/JNEUROSCI.2137-08.2008 . ПМЦ 2638127 . ПМИД 18815262 .
Дальнейшее чтение [ править ]
- Харрис, Эндрю; Локк, Даррен, ред. (2009). Коннексины . Нью-Йорк: Спрингер. дои : 10.1007/978-1-59745-489-6 . ISBN 978-1-934115-46-6 .
Внешние ссылки [ править ]
- Gap+Junctions в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|