клетка Пуркинье
| клетка Пуркинье | |
|---|---|
 Рисунок клеток Пуркинье голубя (А), сделанный Сантьяго Рамоном-и-Кахалем. | |
| Подробности | |
| Произношение | Часто / pɜːrˈkɪ n dʒ ; как jee KIN i / pur - - произносится [1] но чешское произношение такое ( Чешский: [ˈpurkɪɲɛ] клетки |
| Расположение | Мозжечок |
| Форма | Плоская дендритная беседка |
| Функция | Тормозной проекционный нейрон |
| Нейромедиатор | ПЕРЕДНИЙ |
| Пресинаптические связи | Параллельные волокна и лазящие волокна |
| Постсинаптические связи | Глубокие ядра мозжечка |
| Идентификаторы | |
| МеШ | D011689 |
| Нейроимена | 365 |
| НейроЛекс ID | sao471801888 |
| ТА98 | A14.1.07.404 |
| ФМА | 67969 |
| Анатомические термины нейроанатомии | |
Клетки Пуркинье или нейроны Пуркинье , названные в честь чешского физиолога Яна Евангелиста Пуркине, который идентифицировал их в 1837 году. [ нужна ссылка ] представляют собой уникальный тип крупных нейронов, расположенных в мозжечка коре головного мозга . Благодаря своим клеточным телам в форме колбы, множеству ветвящихся дендритов и одному длинному аксону эти клетки необходимы для контроля двигательной активности. Клетки Пуркинье в основном выделяют нейротрансмиттер ГАМК (гамма-аминомасляная кислота), который ингибирует некоторые нейроны, снижая передачу нервных импульсов. Клетки Пуркинье эффективно контролируют и координируют двигательные движения тела посредством этих тормозящих действий. [2] [3]
Структура
[ редактировать ]




Эти клетки являются одними из крупнейших нейронов в человеческом мозге ( клетки Бетца ). самыми крупными являются [4] со сложной дендритной древовидной структурой, характеризующейся большим количеством дендритных шипов . Клетки Пуркинье находятся в Пуркинье мозжечка слое . Клетки Пуркинье выстроены подобно костяшкам домино, сложенным друг перед другом. Их большие дендритные ветви образуют почти двумерные слои, через которые проходят параллельные волокна из более глубоких слоев. Эти параллельные волокна образуют относительно более слабые возбуждающие ( глутаматергические ) синапсы с шипиками в дендритах клеток Пуркинье, тогда как лазящие волокна, исходящие из нижнего ядра оливы в мозговом веществе, обеспечивают очень мощный возбуждающий импульс к проксимальным дендритам и соме клеток. Параллельные волокна проходят ортогонально через дендритную ветвь нейрона Пуркинье и содержат до 200 000 параллельных волокон. [5] образуя синапс гранулярная клетка-клетка Пуркинье с одной клеткой Пуркинье.
Канонически каждая взрослая клетка Пуркинье получает примерно 500 синапсов лазающих волокон, все из которых происходят из одного лазящего волокна нижней оливы. [6] Это привело к идее, что «высоко консервативное взаимно-однозначное соотношение объединяет дендриты Пуркинье в единый вычислительный отсек». [7] Однако теперь обнаружено, что множественная иннервация «происходит» у мышей среди подмножества клеток Пуркинье с множественными первичными дендритами, дендритным мотивом, который редко встречается у грызунов, но «преобладает» у людей. [7]
мозжечка Как корзинчатые, так и звездчатые клетки (обнаруженные в молекулярном слое ) обеспечивают ингибирующий (ГАМКергический) вход в клетку Пуркинье, при этом корзинчатые клетки образуют синапсы на начальном сегменте аксона клеток Пуркинье, а звездчатые клетки - на дендритах.
Клетки Пуркинье посылают тормозные проекции в глубокие ядра мозжечка и представляют собой единственный результат всей двигательной координации в коре мозжечка.
Молекулярный
[ редактировать ]Слой Пуркинье мозжечка, который содержит клеточные тела клеток Пуркинье и глию Бергмана , экспрессирует большое количество уникальных генов. [8] Маркеры генов, специфичных для Пуркинье, были также предложены путем сравнения транскриптома мышей с дефицитом Пуркинье с транскриптомом мышей дикого типа. [9] Одним из показательных примеров является белок 4 клеток Пуркинье ( PCP4 ) у нокаутных мышей , у которых наблюдается нарушение локомоторного обучения и заметно измененная синаптическая пластичность нейронов Пуркинье. [10] [11] PCP4 ускоряет как ассоциацию, так и диссоциацию кальция (Ca 2+ ) с кальмодулином (СаМ) в цитоплазме клеток Пуркинье, а его отсутствие нарушает физиологию этих нейронов. [10] [11] [12] [13]
Разработка
[ редактировать ]Исследования эмбрионов млекопитающих подробно описали нейрогенное происхождение клеток Пуркинье. [14] На ранних стадиях развития клетки Пуркинье возникают в желудочковой зоне нервной трубки, предшественнике нервной системы эмбриона. Все нейроны мозжечка происходят из зародышевого нейроэпителия желудочковой зоны. [15] Клетки Пуркинье специфически генерируются из предшественников желудочкового нейроэпителия эмбрионального зачатка мозжечка. [16] Первые клетки, образующиеся из зачатка мозжечка, образуют колпачок над ромбовидной полостью развивающегося мозга, называемой четвертым желудочком, образуя два полушария мозжечка. Клетки Пуркинье, которые развиваются позже, относятся к центральной части мозжечка, называемой червем. Они развиваются в зачатке мозжечка, который покрывает четвертый желудочек, и ниже щелевидной области, называемой перешейком развивающегося мозга. Клетки Пуркинье мигрируют к внешней поверхности коры мозжечка и образуют слой клеток Пуркинье.
Клетки Пуркинье рождаются на самых ранних стадиях нейрогенеза мозжечка. Нейрогенин2 вместе с нейрогенином1 временно экспрессируются в ограниченных доменах желудочкового нейроэпителия в течение временного окна генеза клеток Пуркинье. [17] Этот паттерн пространственно-временного распределения предполагает, что нейрогенины участвуют в спецификации фенотипически гетерогенных субпопуляций клеток Пуркинье, в конечном итоге ответственных за построение каркаса топографии мозжечка.
На мышах и людях имеются доказательства того, что клетки костного мозга либо сливаются с клетками Пуркинье мозжечка, либо генерируют их, и вполне возможно, что клетки костного мозга либо путем прямого образования, либо путем слияния клеток могут играть роль в восстановлении повреждений центральной нервной системы. [18] [19] [20] [21] [22] Дальнейшие данные указывают на возможность существования общего предка стволовых клеток среди нейронов Пуркинье, B-лимфоцитов и альдостерон -продуцирующих клеток человека коры надпочечников . [21]
Функция
[ редактировать ]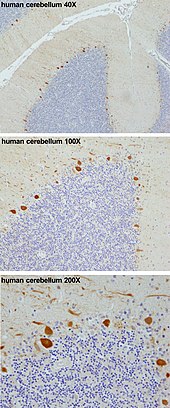
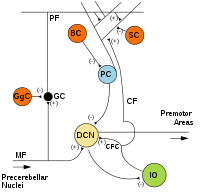
МФ: Мшистое волокно .
DCN: Глубокие ядра мозжечка .
Я: Плохие оливки .
CF: Восхождение по волокну .
GC: Гранульная клетка .
PF: Параллельное волокно .
ПК: клетка Пуркинье.
GgC: клетка Гольджи .
СК: Звездчатая клетка .
БК: Ячейка-корзина .
Клетки Пуркинье демонстрируют две различные формы электрофизиологической активности:
- Простые спайки возникают с частотой 17–150 Гц (Raman and Bean, 1999) либо спонтанно, либо когда клетки Пуркинье активируются синаптически параллельными волокнами, аксонами гранулярных клеток.
- Сложные спайки — это медленные спайки частотой 1–3 Гц, характеризующиеся начальным продолжительным спайком большой амплитуды, за которым следует высокочастотный всплеск потенциалов действия меньшей амплитуды. Они вызваны активацией карабкающихся волокон и могут включать в себя генерацию опосредованных кальцием потенциалов действия в дендритах. После сложной импульсной активности простые спайки могут быть подавлены мощным входным комплексным спайком. [23]
Клетки Пуркинье проявляют спонтанную электрофизиологическую активность в виде цепочек спайков, как натрий-зависимых, так и кальций-зависимых. Первоначально это было показано Родольфо Ллинасом (Ллинас и Хесс (1977) и Ллинас и Сугимори (1980)). Кальциевые каналы P-типа были названы в честь клеток Пуркинье, где они впервые были обнаружены (Llinas et al. 1989), которые имеют решающее значение для функции мозжечка. Активация клетки Пуркинье по лазящим волокнам может переводить ее активность из спокойного состояния в спонтанно активное и наоборот, служа своеобразным тумблером. [24] Эти результаты были оспорены исследованием, показавшим, что такое переключение с помощью входов лазающих волокон происходит преимущественно у анестезированных животных и что клетки Пуркинье у бодрствующих животных в целом функционируют почти непрерывно в северной части штата. [25] Но это последнее исследование само по себе было оспорено [26] и переключение клеток Пуркинье с тех пор наблюдалось у бодрствующих кошек. [27] Вычислительная модель клетки Пуркинье показала, что за переключение ответственны внутриклеточные вычисления кальция. [28]
Результаты показали, что дендриты клеток Пуркинье выделяют эндоканнабиноиды , которые могут временно подавлять как возбуждающие, так и тормозные синапсы. [29] Режим собственной активности клеток Пуркинье задается и контролируется натрий-калиевым насосом . [30] Это говорит о том, что насос может быть не просто гомеостатической молекулой, «хозяйственной» для ионных градиентов. Вместо этого это может быть вычислительный элемент в мозжечке и мозге. [31] , мутация Na Действительно +
- К +
насос вызывает быстрое начало дистонии, паркинсонизма; его симптомы указывают на то, что это патология вычислений мозжечка. [32] Кроме того, использование яда уабаина для блокирования Na +
- К +
насосы в мозжечке живой мыши вызывают атаксию и дистонию . [33] Численное моделирование экспериментальных данных позволяет предположить, что in vivo Na +
- К +
насос производит длинные периоды покоя (>>1 с) до срабатывания нейронов Пуркинье; они могут иметь вычислительную роль. [34] Алкоголь ингибирует Na +
- К +
насосы в мозжечке, и именно это, вероятно, нарушает работу мозжечка и координацию тела. [35] [36]
Клиническое значение
[ редактировать ]У людей клетки Пуркинье могут быть повреждены различными причинами: токсическое воздействие, например, алкоголя или лития; аутоиммунные заболевания ; генетические мутации, вызывающие спиноцеребеллярную атаксию, глютеновую атаксию , болезнь Унферрихта-Лундборга или аутизм ; и нейродегенеративные заболевания, о генетической основе которых неизвестно, такие как мозжечковый тип множественной системной атрофии или спорадические атаксии. [37] [38]
Глютеновая атаксия — аутоиммунное заболевание, вызванное употреблением глютена . [39] Гибель клеток Пуркинье в результате воздействия глютена необратима. Ранняя диагностика и лечение безглютеновой диетой могут улучшить атаксию и предотвратить ее прогрессирование. [37] [40] Менее 10% людей с глютеновой атаксией имеют какие-либо желудочно-кишечные симптомы, однако около 40% имеют поражение кишечника. [40] На его долю приходится 40% атаксий неясного происхождения и 15% всех атаксий. [40]
Нейродегенеративное заболевание спиноцеребеллярная атаксия типа 1 (SCA1) вызвано нестабильной экспансией полиглутамина в белке атаксина 1 . Этот дефект белка атаксина 1 вызывает повреждение митохондрий в клетках Пуркинье, что приводит к преждевременной дегенерации клеток Пуркинье. [41] В результате координация движений ухудшается и в конечном итоге наступает смерть.
У некоторых домашних животных может развиться состояние, при котором клетки Пуркинье начинают атрофироваться вскоре после рождения, называемое абиотрофией мозжечка . Это может привести к таким симптомам, как атаксия , интенционный тремор, гиперреактивность, отсутствие рефлекса угрозы , скованная походка или высокая походка, явное отсутствие осознания положения стопы (иногда стоя или ходьба с согнутой ногой) и общая неспособность определить пространство и расстояние. [42] Подобное состояние, известное как гипоплазия мозжечка, возникает, когда клетки Пуркинье не развиваются внутриутробно или отмирают до рождения.
Генетические состояния атаксия-телеангиэктазия и болезнь Нимана-Пика типа C, а также эссенциальный тремор мозжечка приводят к прогрессирующей потере клеток Пуркинье.При болезни Альцгеймера иногда наблюдают патологию позвоночника, а также потерю дендритных ветвей клеток Пуркинье. [43] Клетки Пуркинье также могут быть повреждены вирусом бешенства при его миграции из места заражения на периферии в центральную нервную систему. [44]
Этимология
[ редактировать ]Клетки Пуркинье названы в честь чешского ученого Яна Евангелиста Пуркине, открывшего их в 1839 году. [ нужна ссылка ]
См. также
[ редактировать ]Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Джонс, Дэниел (2011). Роуч, Питер ; Сеттер, Джейн ; Эслинг, Джон (ред.). Кембриджский словарь английского произношения (18-е изд.). Издательство Кембриджского университета. ISBN 978-0-521-15255-6 .
- ^ «Клетка Пуркинье | Гранулярные клетки, мозжечок и нейроны | Британника» . www.britanica.com . 05.01.2024 . Проверено 16 января 2024 г.
- ^ Пол, Маника С.; Лимаим, Фатен (2023), «Гистология, клетки Пуркинье» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 31424738 , получено 16 января 2024 г.
- ^ Первс Д., Августин Г.Дж., Фицпатрик Д., Холл У.К., ЛаМантия А., Макнамара Дж.О. и Уайт Л.Е. (2008). Нейронаука. 4-е изд . Синауэр Ассошиэйтс. стр. 432–4. ISBN 978-0-87893-697-7 .
- ^ Тиррелл, Т; Уиллшоу, Д. (29 мая 1992 г.). «Кора мозжечка: ее моделирование и актуальность теории Марра». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 336 (1277): 239–57. Бибкод : 1992РСТБ.336..239Т . дои : 10.1098/rstb.1992.0059 . ПМИД 1353267 .
- ^ Вадиче, Дж.И.; Яр, CE (25 октября 2001 г.). «Мультивезикулярное высвобождение при восхождении на синапсы волокон-клеток Пуркинье» . Нейрон . 32 (2): 301–13. дои : 10.1016/S0896-6273(01)00488-3 . ПМИД 11683999 .
- ^ Перейти обратно: а б Буш, Сайлас Э.; Гензель, Кристиан (2023). «Мультииннервация лазающих волокон дендритов Пуркинье мыши с ветвлением, свойственным человеку» . Наука . 381 (6656): 420–427. дои : 10.1126/science.adi1024 . ISSN 0036-8075 . ПМЦ 10962609 . ПМИД 37499000 .
- ^ Кирш, Л; Лискович, Н; Чечик, Г (декабрь 2012 г.). Олер, Уве (ред.). «Локализация генов в слоях мозжечка путем классификации изображений ISH» . PLOS Вычислительная биология . 8 (12): e1002790. Бибкод : 2012PLSCB...8E2790K . дои : 10.1371/journal.pcbi.1002790 . ПМЦ 3527225 . ПМИД 23284274 .
- ^ Ронг, Ю; Ван Т; Морган Дж (2004). «Идентификация кандидатных маркеров, специфичных для клеток Пуркинье, путем анализа экспрессии генов у мышей дикого типа и pcd3j». Молекулярные исследования мозга . 13 (2): 128–145. doi : 10.1016/j.molbrainres.2004.10.015 . ПМИД 15582153 .
- ^ Перейти обратно: а б с Фелисола С.Дж., Накамура Й., Оно Й., Китамура К., Кикучи К., Онодера Й., Исе К., Такасе К., Сугавара А., Хаттангади Н., Рейни В.Е., Сато Ф., Сасано Х. (апрель 2014 г.). «PCP4: регулятор синтеза альдостерона в тканях надпочечников человека» . Журнал молекулярной эндокринологии . 52 (2): 159–167. дои : 10.1530/JME-13-0248 . ПМК 4103644 . ПМИД 24403568 .
- ^ Перейти обратно: а б Вэй П., Бландон Дж.А., Ронг Ю., Захаренко С.С., Морган Дж.И. (2011). «Нарушение локомоторного обучения и изменение синаптической пластичности мозжечка у мышей с нулевым содержанием pep-19 / PCP4» . Мол. Клетка. Биол . 31 (14): 2838–44. дои : 10.1128/MCB.05208-11 . ПМК 3133400 . ПМИД 21576365 .
- ^ Путки Дж.А., Клирекопер К., Гертнер Т.Р., Ваксхэм М.Н. (2004). «Новая роль белков мотива IQ в регуляции функции кальмодулина» . Ж. Биол. Хим . 278 (50): 49667–70. дои : 10.1074/jbc.C300372200 . ПМИД 14551202 .
- ^ Клеерекопер К.К., Путкей Дж.А. (2009). «PEP-19, внутренне неупорядоченный регулятор передачи сигналов кальмодулина» . Ж. Биол. Хим . 284 (12): 7455–64. дои : 10.1074/jbc.M808067200 . ПМК 2658041 . ПМИД 19106096 .
- ^ Сотело С, Росси Ф (2013). «Миграция и дифференцировка клеток Пуркинье». Справочник по мозжечку и мозжечковым расстройствам . стр. 147–178. дои : 10.1007/978-94-007-1333-8_9 . ISBN 978-94-007-1332-1 . S2CID 80927298 .
- ^ Хосино М (2006). «Молекулярный механизм, регулирующий спецификацию ГАМКергических нейронов в мозжечке». Мозжечок . 5 (3): 193–198. дои : 10.1080/14734220600589202 . ПМИД 16997750 . S2CID 20937713 .
- ^ Карлетти Б., Росси Ф. (2008). «Нейрогенез в мозжечке». Нейробиолог . 14 (1): 91–100. дои : 10.1177/1073858407304629 . ПМИД 17911211 . S2CID 34889988 .
- ^ Зордан П., Крочи Л., Хоукс Р., Консалес Г.Г. (2008). «Сравнительный анализ экспрессии пронейральных генов в эмбриональном мозжечке» . Дев Дин . 237 (6): 726–735. дои : 10.1002/dvdy.21571 . ПМИД 18498101 .
- ^ Хесс, округ Колумбия, Хилл В.Д., Кэрролл Дж.Э., Борлонган К.В. (2004). «Генерируют ли клетки костного мозга нейроны?» . Архив неврологии . 61 (4): 483–485. дои : 10.1001/archneur.61.4.483 . ПМИД 15096394 .
- ^ Вейманн Дж. М., Йоханссон CB, Трехо А., Блау Х. М. (2003). «Стабильные перепрограммированные гетерокарионы спонтанно образуются в нейронах Пуркинье после трансплантации костного мозга» . Природная клеточная биология . 5 (11): 959–966. дои : 10.1038/ncb1053 . ПМИД 14562057 . S2CID 33685652 .
- ^ Альварес-Доладо М., Пардал Р., Гарсия-Вердуго Х.М., Фике Дж.Р., Ли Х.О., Пфеффер К., Лоис С., Моррисон С.Дж., Альварес-Буйя А. (2003). «Слияние клеток костномозгового происхождения с нейронами Пуркинье, кардиомиоцитами и гепатоцитами» . Природа . 425 (6961): 968–973. Бибкод : 2003Natur.425..968A . дои : 10.1038/nature02069 . hdl : 2027.42/62789 . ПМИД 14555960 . S2CID 4394453 .
- ^ Перейти обратно: а б Фелисола С.Дж., Катсу К., Исе К., Накамура Ю., Арай Ю., Сато Ф., Сасано Х. (2015). «Экспрессия белка 3 Pre-B-лимфоцитов (VPREB3) в коре надпочечников: прецедент неиммунологической роли в нормальных и неопластических тканях человека» . Эндокринная патология . 26 (2): 119–28. дои : 10.1007/s12022-015-9366-7 . ПМИД 25861052 . S2CID 27271366 .
- ^ Кемп К., Уилкинс А., Сколдинг Н. (2014). «Слияние клеток в мозге: две клетки вперед, одна клетка назад» . Акта Нейропатологика . 128 (5): 629–638. дои : 10.1007/s00401-014-1303-1 . ПМК 4201757 . ПМИД 24899142 .
- ^ Эрик Р. Кандел, Джеймс Х. Шварц, Томас М. Джесселл (2000). Принципы нейронауки. 4/е. МакГроу-Хилл. стр.837-40.
- ^ Левенштейн Ю., Махон С., Чаддертон П., Китамура К., Сомполинский Х., Яром Ю. и др. (2005). «Бистабильность клеток Пуркинье мозжечка, модулируемая сенсорной стимуляцией». Природная неврология . 8 (2): 202–211. дои : 10.1038/nn1393 . PMID 15665875 . S2CID 5543355 .
- ^ Шоневилле М., Хосровани С., Винкельман Б.Х., Хобик Ф.Е., ДеДжеу М.Т., Ларсен И.М. и др. (2006). «Клетки Пуркинье у бодрствующих животных работают с повышенным мембранным потенциалом» . Природная неврология . 9 (4): 459–461. дои : 10.1038/nn0406-459 . ПМИД 16568098 .
- ^ Левенштейн Ю., Махон С., Чаддертон П., Китамура К., Сомполинский Х., Яром Ю. и др. (2006). «Клетки Пуркинье у бодрствующих животных работают с повышенным мембранным потенциалом – Ответ» . Природная неврология . 9 : 461. doi : 10.1038/nn0406-461 . S2CID 28713325 .
- ^ Ярцев М.М., Гивон-Майо Р., Маллер М., Дончин О. (2009). «Приостановка клеток Пуркинье в мозжечке бодрствующей кошки» . Границы системной нейронауки . 3 :2. doi : 10.3389/neuro.06.002.2009 . ПМК 2671936 . ПМИД 19390639 .
- ^ Форрест, доктор медицины (2014). «Динамика внутриклеточного кальция позволяет модели нейронов Пуркинье выполнять переключения и получать результаты вычислений на своих входных данных» . Границы вычислительной нейронауки . 8 : 86. дои : 10.3389/fncom.2014.00086 . ПМЦ 4138505 . ПМИД 25191262 .
- ^ Крейцер AC, Regehr WG (март 2001 г.). «Ретроградное ингибирование пресинаптического притока кальция эндогенными каннабиноидами в возбуждающих синапсах на клетки Пуркинье» . Нейрон . 29 (3): 717–27. дои : 10.1016/S0896-6273(01)00246-X . ПМИД 11301030 .
- ^ Форрест, доктор медицинских наук, Уолл, MJ, Press DA, Фэн Дж (декабрь 2012 г.). Цымбалюк Г (ред.). «Натрий-калиевый насос контролирует внутреннюю активность нейрона Пуркинье мозжечка» . ПЛОС ОДИН . 7 (12): e51169. Бибкод : 2012PLoSO...751169F . дои : 10.1371/journal.pone.0051169 . ПМЦ 3527461 . ПМИД 23284664 .
- ^ Форрест, доктор медицины (декабрь 2014 г.). «Натриево-калиевый насос — это элемент обработки информации в вычислениях мозга» . Границы в физиологии . 5 (472): 472. doi : 10.3389/fphys.2014.00472 . ПМЦ 4274886 . ПМИД 25566080 .
- ^ Пушка С (июль 2004 г.). «Расплата за насос: дистония из-за мутаций в Na +/K +-АТФазе» . Нейрон . 43 (2): 153–154. дои : 10.1016/j.neuron.2004.07.002 . ПМИД 15260948 .
- ^ Кальдерон Д.П., Фремонт Р., Краенцлин Ф., Ходаха К. (март 2011 г.). «Нейральные субстраты быстро возникающей дистонии-паркинсонизма» . Природная неврология . 14 (3): 357–65. дои : 10.1038/nn.2753 . ПМК 3430603 . ПМИД 21297628 .
- ^ Форрест, доктор медицины (2014). «Динамика внутриклеточного кальция позволяет модели нейронов Пуркинье выполнять переключения и получать результаты вычислений на своих входных данных» . Границы вычислительной нейронауки . 8 : 86. дои : 10.3389/fncom.2014.00086 . ПМЦ 4138505 . ПМИД 25191262 .
- ^ Форрест, доктор медицины (апрель 2015 г.). «Моделирование действия алкоголя на подробной модели нейронов Пуркинье и более простой суррогатной модели, которая работает более чем в 400 раз быстрее» . BMC Нейронаука . 16 (27): 27. дои : 10.1186/s12868-015-0162-6 . ПМЦ 4417229 . ПМИД 25928094 .
- ^ Форрест, Майкл (апрель 2015 г.). "the_neuroscience_reason_we_fall_over_when_drunk" . Наука 2.0 .
- ^ Перейти обратно: а б Митома Х., Адхикари К., Эшлиманн Д., Чаттопадьяй П., Хадживассилиу М., Хампе К.С. и др. (2016). «Консенсусный документ: Нейроиммунные механизмы мозжечковых атаксий» . Мозжечок (Обзор). 15 (2): 213–32. дои : 10.1007/s12311-015-0664-x . ПМЦ 4591117 . ПМИД 25823827 .
- ^ Джабер М (2017). «Мозжечок как главный игрок в двигательных нарушениях, связанных с расстройствами аутистического синдрома». Энцефале (обзор). 43 (2): 170–175. дои : 10.1016/j.encep.2016.03.018 . ПМИД 27616580 .
- ^ Сапоне А, Бай Дж.К., Чаччи С., Долинсек Дж., Грин П.Х., Хадживассилиу М., Каукинен К., Ростами К., Сандерс Д.С., Шуман М., Ульрих Р., Виллалта Д., Вольта У., Катасси С., Фазано А. (2012). «Спектр расстройств, связанных с глютеном: консенсус по новой номенклатуре и классификации» . BMC Medicine (обзор). 10:13 . дои : 10.1186/1741-7015-10-13 . ПМЦ 3292448 . ПМИД 22313950 .
- ^ Перейти обратно: а б с Хадживасилиу М., Сандерс Д.Д., Эшлиманн Д.П. (2015). «Расстройства, связанные с глютеном: атаксия глютена». Диг Дис (обзор). 33 (2): 264–8. дои : 10.1159/000369509 . ПМИД 25925933 . S2CID 207673823 .
- ^ Стуки Д.М., Рюгсеггер С., Штайнер С., Радеке Дж., Мерфи М.П., Зубер Б., Саксена С. (август 2016 г.). «Митохондриальные нарушения способствуют прогрессированию спиноцеребеллярной атаксии 1 типа и могут быть улучшены с помощью митохондриально-направленного антиоксиданта MitoQ» (PDF) . Свободный Радик. Биол. Мед . 97 : 427–440. doi : 10.1016/j.freeradbiomed.2016.07.005 . ПМИД 27394174 .
- ^ Ссылки см. в обширных ссылках и библиографии в статье об абиотрофии мозжечка , ссылка на которую приведена в начале этого абзаца.
- ^ Маврудис, Айова; Фотиу, DF; Адипепе, LF; Манани, МГ; Нджау, SD; Псарулис, Д; Коста, В.Г.; Балояннис, SJ (ноябрь 2010 г.). «Морфологические изменения клеток Пуркинье человека и отложение нейритных бляшек и нейрофибриллярных клубков на коре мозжечка при болезни Альцгеймера» . Американский журнал болезни Альцгеймера и других деменций . 25 (7): 585–91. дои : 10.1177/1533317510382892 . ПМЦ 10845446 . ПМИД 20870670 . S2CID 30688657 .
- ^ Фекаду, Маконнен (27 марта 2009 г.). «Бешенственный энцефалит, тельца Негри в цитоплазме нейронов клеток Пуркинье мозжечка» . CDC/Frontal Cortex Inc. Проверено 21 июня 2013 г. Примечание: не рецензировано.
Дальнейшее чтение
[ редактировать ]- Ллинас Р., Хесс Р. (июль 1976 г.). «Устойчивые к тетродотоксину дендритные шипы в птичьих клетках Пуркинье» . Учеб. Натл. акад. наук. США . 73 (7): 2520–3. Бибкод : 1976PNAS...73.2520L . дои : 10.1073/pnas.73.7.2520 . ПМК 430632 . ПМИД 1065905 .
- Ллинас Р., Сугимори М. (август 1980 г.). «Электрофизиологические свойства сомат клеток Пуркинье in vitro в срезах мозжечка млекопитающих» . Дж. Физиол . 305 : 171–95. дои : 10.1113/jphysicalol.1980.sp013357 . ПМЦ 1282966 . ПМИД 7441552 .
- Ллинас Р.Р., Сугимори М., Черксей Б. (1989). «Потенциал-зависимая кальциевая проводимость в нейронах млекопитающих. P-канал». Энн. Н-Й Акад. Наука . 560 (1 Кальций Чанн): 103–11. дои : 10.1111/j.1749-6632.1989.tb24084.x . ПМИД 2545128 . S2CID 84107834 .
- Форрест, Майкл (октябрь 2014 г.). Биофизика и расчеты нейрона Пуркинье мозжечка . CreateSpace. ISBN 978-1502454546 .
Внешние ссылки
[ редактировать ]- Библиотека изображений клеток — Пуркинье
- Заболевания мозжечка. Архивировано 26 августа 2009 г. в Wayback Machine.
- Поиск NIF - Клетка Пуркинье. Архивировано 8 июля 2013 г. в Wayback Machine через Информационную структуру нейронауки.