Кальций-активируемый калиевый канал
Эта статья включает список общих ссылок , но в ней отсутствуют достаточные соответствующие встроенные цитаты . ( Август 2018 г. ) |
Кальций-активируемые калиевые каналы – это калиевые каналы, закрытые кальцием. [1] или которые структурно или филогенетически связаны с кальций-зависимыми каналами. Впервые они были обнаружены в 1958 году Гардосом, который увидел, что уровень кальция внутри клетки может влиять на проницаемость калия через клеточную мембрану. Затем, в 1970 году, Мич первым заметил, что внутриклеточный кальций может вызывать калиевые токи. У человека они делятся на три подтипа: большая проводимость или BK-каналы , которые имеют очень высокую проводимость в диапазоне от 100 до 300 пС, промежуточная проводимость или IK-каналы со промежуточной проводимостью от 25 до 100 пС и малая проводимость или SK-каналы. с малыми проводимостями от 2-25 пСм. [2]
Это семейство ионных каналов по большей части активируется внутриклеточным Ca. 2+ и содержит 8 членов в геноме человека. Однако некоторые из этих каналов ( каналы KCa4 и KCa5 ) вместо этого реагируют на другие внутриклеточные лиганды, такие как Na + , кл − и pH . Более того, несколько членов семейства активируются как лигандом, так и напряжением , что еще больше усложняет описание этого семейства. α-субъединицы K- Ca -канала имеют шесть или семь трансмембранных сегментов, подобно KV , каналам но иногда с дополнительной N-концевой трансмембранной спиралью. α-субъединицы образуют гомо- и гетеротетрамерные комплексы. Домен, связывающий кальций, может содержаться в последовательности субъединицы α, как в KCa1 , или может присутствовать в дополнительном кальцийсвязывающем белке, таком как кальмодулин .
Структура
[ редактировать ]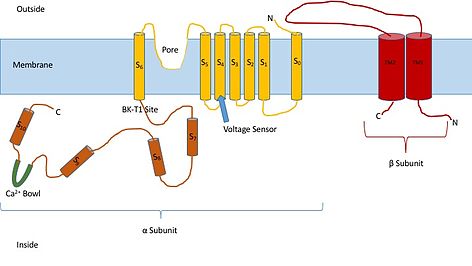
Знание структуры этих каналов может дать представление об их функции и механизме открытия. Они состоят из двух разных субъединиц: альфа и бета. Альфа-субъединица представляет собой тетрамер, который образует пору, датчик напряжения и область, чувствительную к кальцию. Эта субъединица канала состоит из семи трансмембранных единиц и большой внутриклеточной области. Датчик напряжения представляет собой трансмембранную область S4, которая имеет несколько остатков аргинина , которые действуют, «чувствуя» изменения заряда и двигаясь очень похоже на другие потенциалзависимые калиевые каналы. Когда они движутся в ответ на изменения напряжения, они открывают и закрывают ворота. Линкер между регионами S5 и S6 служит для формирования поры канала. Основная часть внутри клетки, на которую следует обратить внимание, — это кальциевая чаша. Считается, что эта чаша является местом связывания кальция. [3]
Считается, что бета-субъединица канала является регуляторной субъединицей канала. Существует четыре различных типа бета-субъединицы: 1, 2, 3 и 4. Бета-2 и 3 являются тормозящими, а бета-1 и 4 — возбуждающими, или они заставляют канал быть более открытым, чем закрытым. Возбуждающие бета-субъединицы влияют на альфа-субъединицы таким образом, что канал редко инактивируется. [4]
Классификация и описание гомологии
[ редактировать ]Человеческие K- Ca- каналы
[ редактировать ]Ниже приведен список из 8 известных кальций-активируемых калиевых каналов человека, сгруппированных в соответствии с гомологией последовательностей трансмембранных гидрофобных ядер: [5]
Хотя это не подразумевается в названии, но подразумевается структурой, эти каналы также могут активироваться напряжением. Считается, что различные способы активации этих каналов независимы друг от друга. Эта особенность канала позволяет им участвовать во многих различных физиологических функциях. Физиологические эффекты BK-каналов широко изучались с использованием нокаутных мышей. При этом было замечено, что произошли изменения в кровеносных сосудах мышей. У животных без каналов ВК наблюдалось повышение среднего артериального давления и сосудистого тонуса. Эти данные указывают на то, что каналы BK участвуют в расслаблении гладкомышечных клеток. В любой мышечной клетке повышенное внутриклеточное содержание кальция вызывает сокращение. В гладкомышечных клетках повышенный уровень внутриклеточного кальция вызывает открытие BK-каналов, которые, в свою очередь, позволяют ионам калия выходить из клетки. Это вызывает дальнейшую гиперполяризацию и закрытие потенциалзависимых кальциевых каналов, после чего может произойти расслабление. У нокаутированных мышей также наблюдался интенционный тремор, более короткая длина шага и более медленная скорость плавания. Все это симптомы атаксия , что указывает на то, что каналы BK очень важны в мозжечке. [6]
Подтипы БК-каналов
Промежуточные каналы проводимости, по-видимому, наименее изучены из всех каналов. Считается, что структурно они очень похожи на каналы BK, основными отличиями которых являются проводимость и методы модуляции. Известно, что ИК-каналы модулируются кальмодулином, а ВК-каналы — нет.
ИК-каналы показали тесную связь с кальцификацией сосудов, поскольку ингибирование канала вызывает уменьшение кальцификации сосудов. Чрезмерная экспрессия этих каналов оказывает на организм совершенно иной эффект. Исследования показали, что такое лечение вызывает пролиферацию гладкомышечных клеток сосудов. Это открытие послужило толчком к дальнейшему исследованию этих каналов, и исследователи обнаружили, что IK-каналы регулируют клеточный цикл в раковых клетках, В- и Т-лимфоцитах и стволовых клетках. Эти открытия обещают будущее лечение каналов IK.
Подтипы или ИК-каналы
Кальций с малой проводимостью активирует калиевые каналы, существенно отличаясь от своих родственников с большей проводимостью. Основное и наиболее интригующее отличие каналов SK заключается в том, что они нечувствительны к напряжению. Эти каналы могут быть открыты только при повышении уровня внутриклеточного кальция. Эта особенность каналов SK позволяет предположить, что они имеют несколько иную структуру, чем каналы BK и IK.
Как и другие калиевые каналы, они участвуют в гиперполяризации клеток после потенциала действия. Свойство этих каналов активировать кальций позволяет им участвовать в вазорегуляции, слуховой настройке волосковых клеток, а также в циркадном ритме. Исследователи пытались выяснить, какие каналы отвечают за реполяризацию и постгиперполяризацию потенциалов действия. Они сделали это, фиксируя напряжение на клетках, обрабатывая их различными блокаторами каналов BK и SK, а затем стимулируя клетку создавать ток. Исследователи обнаружили, что реполяризация клеток происходит из-за каналов BK и что часть постгиперполяризации происходит из-за тока через каналы SK. Они также обнаружили, что при блокировке SK-каналов ток во время постгиперполяризации все же возникал. Был сделан вывод, что существует другой неизвестный тип калиевых каналов, обеспечивающих эти токи. [7]
Понятно, что в МАИ участвуют каналы СК. Неясно, как именно это происходит. Есть три разные идеи о том, как это сделать. 1) Простая диффузия кальция объясняет медленную кинетику этих токов, 2) Медленная кинетика обусловлена другими каналами с медленной активацией, или 3) Кальций просто активирует систему вторичных мессенджеров для активации SK-каналов. Было показано, что простая диффузия является маловероятным механизмом, поскольку ток чувствителен к температуре, а диффузионный механизм не будет чувствителен к температуре. Это также маловероятно, поскольку с концентрацией кальция меняется только амплитуда тока, а не кинетика активации канала.
Подтипы каналов SK
Другие подсемейства
[ редактировать ]Прокариотические K- Ca- каналы
[ редактировать ]ряд прокариотических KCa - Описан каналов как структурно, так и функционально. Все они либо закрываются кальцием, либо другими лигандами и гомологичны человека каналам KCa , в частности воротному кольцу KCa1.1 . Эти структуры послужили матрицами для лигандного гейтирования.
| Белок | Разновидность | Лиганд | Функция | Ссылка |
|---|---|---|---|---|
| Кч | кишечная палочка | Неизвестный | Канал | [8] [9] |
| МтК | Метанотермобактер термоавтотрофный | Кальций , Кадмий , Барий , pH | Канал | [10] [11] [12] [13] [14] |
| ТркА/ТркХ | Вибрион парагемолитический | АТФ , АДФ | Канал | [15] [16] |
| КтрАБ | Бацилла субтилис | АТФ, АДФ | Транспортер | [17] |
| ГсуК | Геобактер серыредуценс | Кальций, АДФ, НАД | Канал | [18] |
| ТМ1088 | Термотога приморская | Неизвестный | Неизвестный | [19] |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Вергара, К.; Латорре, Р.; Маррион, Невада; Адельман, JP (1998). «Кальций-активируемые калиевые каналы». Современное мнение в нейробиологии . 8 (3): 321–329. дои : 10.1016/s0959-4388(98)80056-1 . ISSN 0959-4388 . ПМИД 9687354 . S2CID 40840564 .
- ^ УИВЕР, ЭМИ К.; БОМБЕН, ВАЛЕРИ К.; СОНТАЙМЕР, ХАРАЛЬД (15 августа 2006 г.). «Экспрессия и функция кальций-активируемых калиевых каналов в клетках глиомы человека» . Глия . 54 (3): 223–233. дои : 10.1002/glia.20364 . ISSN 0894-1491 . ПМК 2562223 . ПМИД 16817201 .
- ^ Гхатта, Шринивас; Ниммагадда, Дипти; Сюй, Сяопин; О'Рурк, Стивен Т. (1 апреля 2006 г.). «Калиевые каналы, активируемые кальцием, с большой проводимостью: структурные и функциональные последствия». Фармакология и терапия . 110 (1): 103–116. doi : 10.1016/j.pharmthera.2005.10.007 . ПМИД 16356551 .
- ^ «Кальций- и натрий-активируемые калиевые каналы | Введение | Руководство BPS/IUPHAR по ФАРМАКОЛОГИИ» . www.guidetopharmacology.org . Проверено 22 апреля 2019 г.
- ^ Вэй А.Д., Гутман Г.А., Олдрич Р., Чанди К.Г., Гриссмер С., Вульф Х. (декабрь 2005 г.). «Международный союз фармакологии. ЛИИ. Номенклатура и молекулярные взаимоотношения кальций-активируемых калиевых каналов». Фармакологические обзоры . 57 (4): 463–72. дои : 10.1124/пр.57.4.9 . ПМИД 16382103 . S2CID 8290401 .
- ^ Бреннер, Р. (2000). «Клонирование и функциональная характеристика новых бета-субъединиц кальций-активируемых калиевых каналов с большой проводимостью, hKCNMB3 и hKCNMB4» . J Биол Хим . 275 (9): 6453–6461. дои : 10.1074/jbc.275.9.6453 . ПМИД 10692449 .
- ^ Сах, Панкадж (1996). «Ca2+-активированные K+-токи в нейронах: типы, физиологическая роль и модуляция». Тенденции в нейронауках . 19 (4): 150–154. дои : 10.1016/s0166-2236(96)80026-9 . ПМИД 8658599 . S2CID 9504595 .
- ^ Молочник Р. (апрель 1994 г.). «Гомолог эукариотических белков калиевых каналов Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (9): 3510–4. Бибкод : 1994PNAS...91.3510M . дои : 10.1073/pnas.91.9.3510 . ПМК 43609 . ПМИД 8170937 .
- ^ Цзян Ю, Пико А., Каден М., Чайт Б.Т., Маккиннон Р. (март 2001 г.). «Структура домена RCK из K+-канала E. coli и демонстрация его присутствия в канале BK человека» . Нейрон . 29 (3): 593–601. дои : 10.1016/s0896-6273(01)00236-7 . ПМИД 11301020 .
- ^ Цзян Ю, Ли А, Чен Дж, Каден М, Чайт БТ, Маккиннон Р (май 2002 г.). «Кристаллическая структура и механизм кальций-управляемого калиевого канала». Природа . 417 (6888): 515–22. Бибкод : 2002Natur.417..515J . дои : 10.1038/417515a . ПМИД 12037559 . S2CID 205029269 .
- ^ Смит Ф.Дж., Пау В.П., Чинголани Дж., Ротберг Б.С. (2013). «Структурные основы аллостерических взаимодействий между сайтами связывания Са2+ в домене RCK K+-канала» . Природные коммуникации . 4 : 2621. Бибкод : 2013NatCo...4.2621S . дои : 10.1038/ncomms3621 . ПМИД 24126388 .
- ^ Е С., Ли Ю, Чен Л, Цзян Ю (сентябрь 2006 г.). «Кристаллические структуры безлигандного воротного кольца MthK: понимание лигандного механизма управления K+-каналами» . Клетка . 126 (6): 1161–73. дои : 10.1016/j.cell.2006.08.029 . ПМИД 16990139 .
- ^ Двир Х., Валера Э., Чой С. (август 2010 г.). «Структура МТК РЦК в комплексе с кадмием» . Журнал структурной биологии . 171 (2): 231–7. дои : 10.1016/j.jsb.2010.03.020 . ПМЦ 2956275 . ПМИД 20371380 .
- ^ Смит Ф.Дж., Пау Вице-президент, Чинголани Дж., Ротберг Б.С. (декабрь 2012 г.). «Кристаллическая структура воротного кольца, связанного с Ba(2+), раскрывает элементарные этапы активации домена RCK» . Структура . 20 (12): 2038–47. дои : 10.1016/j.str.2012.09.014 . ПМЦ 3518701 . ПМИД 23085076 .
- ^ Цао Ю, Цзинь Х, Хуан Х, Деребе М.Г., Левин Э.Дж., Кабалиесваран В., Пан Ю., Пунта М., Лав Дж., Венг Дж., Квик М., Йе С., Клосс Б., Бруни Р., Мартинес-Хакерт Е., Хендриксон В.А., Рост Б., Джавич Дж.А., Раджашанкар К.Р., Цзян Ю., Чжоу М. (март 2011 г.). «Кристаллическая структура переносчика ионов калия ТркХ» . Природа . 471 (7338): 336–40. Бибкод : 2011Natur.471..336C . дои : 10.1038/nature09731 . ПМК 3077569 . ПМИД 21317882 .
- ^ Цао Ю, Пан Ю, Хуан Х, Цзинь Икс, Левин Э.Дж., Клосс Б., Чжоу М. (апрель 2013 г.). «Закрытие ионного канала TrkH связанным с ним белком RCK TrkA» . Природа . 496 (7445): 317–22. Бибкод : 2013Natur.496..317C . дои : 10.1038/nature12056 . ПМЦ 3726529 . ПМИД 23598339 .
- ^ Виейра-Пиреш Р.С., Соллоши А., Мораиш-Кабрал Ж.Х. (апрель 2013 г.). «Структура переносчика калия КтрАБ». Природа . 496 (7445): 323–8. Бибкод : 2013Природа.496..323В . дои : 10.1038/nature12055 . hdl : 10216/110345 . ПМИД 23598340 . S2CID 205233489 .
- ^ Конг С., Цзэн В., Йе С., Чен Л., Зауэр Д.Б., Лам Ю., Деребе М.Г., Цзян Ю. (2012). «Различные механизмы шлюзования, выявленные структурами мультилигандного закрытого канала K (+)» . электронная жизнь . 1 : е00184. doi : 10.7554/eLife.00184 . ПМК 3510474 . ПМИД 23240087 .
- ^ Деллер MC, Джонсон HA, Миллер MD, Спраггон Дж., Элслигер М.А., Уилсон И.А., Лесли С.А. (2015). «Кристаллическая структура двухсубъединичного октамерного литникового кольца TrkA» . ПЛОС ОДИН . 10 (3): e0122512. Бибкод : 2015PLoSO..1022512D . дои : 10.1371/journal.pone.0122512 . ПМЦ 4380455 . ПМИД 25826626 .
Внешние ссылки
[ редактировать ]- Активированные кальцием+калием+каналы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- «Кальций-активируемые калиевые каналы» . База данных IUPHAR по рецепторам и ионным каналам . Международный союз фундаментальной и клинической фармакологии.