ТРПВ6
| ТРПВ6 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TRPV6 , ABP/ZF, CAT1, CATL, ECAC2, HSA277909, LP6728, ZFAB, член 6 подсемейства V катионного канала переходного рецепторного потенциала, HRPTTN | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 606680 ; МГИ : 1927259 ; Гомологен : 56812 ; Генные карты : TRPV6 ; ОМА : TRPV6 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





TRPV6 представляет собой мембранный кальций (Ca 2+ ) белок канала, который особенно участвует в первом этапе Ca 2+ всасывание в кишечнике .
Классификация
[ редактировать ]Потенциал транзиторных рецепторов. Представитель 6 подсемейства ваниллоидов (TRPV6) представляет собой эпителиальный Ca. 2+ канал , который принадлежит к временного рецепторного потенциала (TRP). семейству белков [5] Семейство TRP представляет собой группу канальных белков, имеющих решающее значение для ионного гомеостаза и восприятия различных физических и химических стимулов. Каналы TRP могут определять температуру , осмотическое давление , обоняние , вкус и механические силы. [5] [6] Геном человека кодирует 28 каналов TRP, включая шесть каналов TRPV . [5] Высокий Ca 2+ -селективность TRPV5 и TRPV6 отличает эти каналы от других четырех каналов TRPV (TRPV1-TRPV4). [7] TRPV5 и TRPV6 участвуют в синтезе кальция. 2+ транспорта, тогда как TRPV1 – TRPV3 являются тепловыми сенсорами с различным температурным порогом активации, а TRPV4 участвует в определении осмолярности . [8] [9] Генетические дефекты гена TRPV6 связаны с преходящим неонатальным гиперпаратиреозом и ранним хроническим панкреатитом . Нарушение регуляции TRPV6 также участвует в гиперкальциурии , образовании камней в почках , заболеваниях костей, дефектах дифференцировки кератиноцитов , деформациях скелета, остеоартрите , мужском бесплодии , синдроме Пендреда и некоторых подтипах рака . [8] [9]
Идентификация
[ редактировать ]Пэн и др. крысы идентифицировали TRPV6 в 1999 году в двенадцатиперстной кишке в попытке найти Ca. 2+ транспортирующие белки, участвующие в Ca 2+ поглощение. [10] TRPV6 также называли белком-транспортером кальция 1 (CaT1). [10] [11] первоначально, хотя название эпителиального кальциевого канала 2 (ECaC2) [12] [13] и CaT1-подобный (CaT-L) [14] также использовались в ранних исследованиях для описания канала. [10] [12] [13] [14] TRPV6 человека и мыши Ортологи были клонированы Peng et al и Weber et al соответственно. [11] [12] Название TRPV6 было подтверждено в 2005 году. [15]
Расположение генов, расположение хромосом и филогения
[ редактировать ]человека TRPV6 Ген расположен в хромосомном локусе 7q33-34 рядом с его гомологом TRPV5 на 7q35. [16] [17] Ген TRPV6 у человека кодирует мРНК длиной 2906 п.н. [17] В отличие от большинства других белков, которые инициируют трансляцию с помощью кодона AUG , трансляция TRPV6 инициируется путем чтения, не опосредованного кодоном AUG. [18] Белок TRPV6 имеет N-концевое удлинение длиной 40 аминокислот в плаценте и в некоторых физиологических условиях по сравнению с аннотированной версией белка, используемой в биологических исследованиях. [18] Однако еще предстоит определить, является ли длинная версия белка TRPV6 доминирующей формой в разных тканях.
| Разновидность | Человек | Крыса | Мышь |
| Хромосомное расположение | 7q33-q34 | 4q22 | 6Б2 |
| Длина с аннотациями | 725 | 727 | 727 |
| In vivo длина aa а | 765 | 767 | 767 |
| Нуклеотид RefSeq | НМ_018646 | НМ_053686 | НМ_022413 |
| Белок RefSeq | НП_061116 | НП_446138 | НП_071858 |
а Подлежит проверке в различных тканях.
Была выдвинута гипотеза, что гены Trpv5 и Trpv6 возникли из одного предкового гена в результате событий дупликации генов . [16] [19] Филогенетический анализ показал, что паралоги TRPV6 у млекопитающих , зауропсидов , амфибий и хондрихтиев возникли в результате независимых событий дупликации у предков каждой группы. [19] Предполагается, что два специализированных Ca 2+ -селективные гомологи Trpv возникли как адаптация для достижения большей степени функциональной специализации для решения различных проблем с почками у наземных животных. [19]
два аллеля гена TRPV6 (первоначально обозначенные как CaT-La и CaT-Lb). У человека идентифицированы [14] Эти аллели демонстрируют спаренный полиморфизм, порождающий две версии одного и того же гена. [14] [20] Полиморфизмы приводят к появлению «предкового варианта» и «производного варианта», которые различаются пятью основаниями и тремя аминокислотами. [14] [20] Предковые аллели кодируют C197 (157, в скобках указана аннотированная нумерация аминокислот), M418 (378) и M721 (681), тогда как производные аллели кодируют R197 (157), V418 (378) и T721 (681). [20] [21] Частота предкового аллеля TRPV6 варьируется в разных группах населения. [20] [21] Предполагается, что давление отбора, которое могло изменить распределение аллелей TRPV6 , включает изменения в структуре потребления молока, одомашнивание животных, изменение воздействия ультрафиолетового света из-за трансэкваториальной миграции, геномные адаптации, обеспечивающие иммунные преимущества популяциям, сталкивающимся с новыми патогенами. [20] [21] [22]
Распределение тканей
[ редактировать ]Белок TRPV6 экспрессируется в эпителиальных тканях, таких как кишечник , почки , плацента , придаток яичка и экзокринные железы , такие как поджелудочная железа , предстательная и слюнная , потовые и молочные железы . [23] [24] Экспрессия белка TRPV6 у людей была продемонстрирована подходов в пищеводе , желудке , тонком кишечнике , толстой кишке , поджелудочной железе, молочных железах, яичниках , щитовидной железе и простате с помощью иммуногистохимических . [23] Экспрессия TRPV6 в основном ограничивается апикальной мембраной эпителиальных клеток. В кишечнике белок экспрессируется на мембране щеточной каймы энтероцитов .
Сообщалось о различиях в профиле экспрессии TRPV6, возможно, из-за различий в зависимости от анализа, таких как конструкция праймеров , гибридизационные зонды , ПЦР по сравнению с нозерн-блоттингом , полуколичественная ПЦР по сравнению с ОТ-ПЦР и антителами, используемыми для иммунодетекции. [25] На профиль экспрессии TRPV6 также влияют возраст, пол, Ca 2+ и уровни витамина D3 в пище, гормональный статус, расположение в тканях, клеточное расположение, репродуктивный статус и статус отлучения от груди (см. Раздел «Регулирование» ).
У людей транскрипты TRPV6 были обнаружены в плаценте, поджелудочной железе, раке предстательной железы, двенадцатиперстной кишке и простате с помощью нозерн-блоттинга; и в двенадцатиперстной кишке, тощей кишке, плаценте, поджелудочной железе, семенниках, почках, головном мозге и толстой кишке с помощью полуколичественной ПЦР. [13] У грызунов экспрессия TRPV6 была подтверждена в двенадцатиперстной кишке, слепой кишке, тонкой кишке, толстой кишке, плаценте, поджелудочной железе, простате и придатках яичка с помощью нозерн-блоттинга. [10] [17] [26] У мышей содержание транскриптов TRPV6, измеренное с помощью RT-PCR, следующее: простата > желудок, головной мозг > легкие > двенадцатиперстная кишка, слепая кишка, сердце, почки, кости > толстая кишка > скелетные мышцы > поджелудочная железа. [27]
Данные, полученные из Атласа белков человека и на основе секвенирования РНК, позволяют предположить, что мРНК TRPV6 имеет низкий уровень в большинстве тканей, за исключением плаценты, слюнной железы, поджелудочной железы и простаты. [24] [28] мРНК TRPV6 экспрессируется в апикальном домене мышиных остеокластов кортикальной кости. [29] [30] Кортикальные и трабекулярные остеоциты не экспрессируют мРНК TRPV6, тогда как остеобласты демонстрируют слабую экспрессию. [31]
Структура и биофизические свойства
[ редактировать ]Первичная и вторичная структура
[ редактировать ]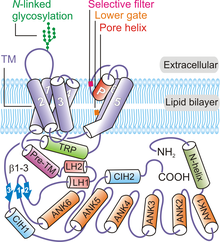
В целом четыре субъединицы TRPV6 образуют тетрамерный канал, обладающий четырехкратной симметрией . [8] [32] Начиная с N-конца и продвигаясь к С-концу белка, каждый полипептид TRPV6 содержит: N-концевую спираль, домен анкиринового повтора (ARD), содержащий шесть анкириновых повторов, линкерный домен β-шпилечной структуры, составляющий два β-конца. -нити , мотив спираль-поворот-спираль, спираль пре-SI, домен TM, состоящий из шести спиралей TM (от S1 до S6), пора-петля (также называемая P-петлей), амфипатическая спираль TRP, C-конец крючок и β-цепь из шести остатков (β3) ( рис. 1 ). [8] [32]
Третичная и четверичная структура
[ редактировать ]
Белок канала TRPV6 обладает четырехкратной симметрией и содержит два основных компартмента: трансмембранный домен высотой 30 Å с центральной порой ионного канала и внутриклеточную юбку высотой ~70 Å и шириной ~110 Å, окружающую 50 Å × 50 Å полость широкая полость под ионным каналом. [32] Кластеризация четырех субъединиц TRPV6 образует водную пору, обладающую четырехкратной симметрией ( рис. 2 ). Пре-SI-спираль связывает внутриклеточную часть белка с доменом TM через линкерный домен, состоящий из структуры β-шпильки и мотива спираль-поворот-спираль. Спирали от S1 до S4 образуют трансмембранный спиральный пучок или домен ТМ, который встраивается почти перпендикулярно плоскости плазматической мембраны. [32]
Элементы порового модуля состоят из S5, S6 и P-петли в TM-доменах. [32] Модуль пор каждого полипептида TRPV6 участвует во взаимодействиях между субъединицами, образуя центральную ионную пору ( рис. 1 ). [32] Порообразующие элементы каждой субъединицы TRPV6 также взаимодействуют с доменами S1-S4 соседнего полипептида в порядке замены доменов. [32] [33] Межсубъединичные взаимодействия также происходят между внеклеточными петлями S1-S2 и петлями S5-P и S6-P соседних субъединиц TRPV6. [32] Консервативный сайт N-связанного гликозилирования в петле S1-S2 необходим для Klotho -опосредованной активации. [34] Внутриклеточная юбка белка TRPV6 в основном состоит из анкириновых повторов. [32] Домен TRP ориентирован параллельно мембране и участвует в гидрофобных взаимодействиях с доменом TM и гидрофильных взаимодействиях во внутриклеточной юбке. N-концевая спираль, С-концевой крючок и β-листы (образованные структурой β-шпильки в линкерном домене) в канале участвуют в межсубъединичных взаимодействиях с ARD, обеспечивая каркас для удержания элементов внутриклеточной юбки. вместе. [8] [32]
Архитектура пор и места связывания катионов
[ редактировать ]Пора TRPV6 состоит из четырех основных элементов: внеклеточного преддверия, селективного фильтра, гидрофобной полости и нижних ворот. [32] [35] [36] Обращенный к центральному просвету канала селективный фильтр с четырьмя остатками ( 538 ВРЕМЯ 541 ), содержащий четыре боковые цепи аспартата 541 (D541) (по одной от каждого протомера), имеет решающее значение для Ca 2+ избирательность и другие биофизические свойства канала. [32] [35] [36] Этот фильтр образует отрицательно заряженное кольцо, которое различает ионы в зависимости от их размера и заряда. Мутации в критическом порообразующем остатке TRPV6 блокируют Ca 2+ При поглощении была использована стратегия для создания моделей потери функции TRPV6 для изучения роли канала в физиологии животных. [35] [36] Считается, что в канале TRPV6 существуют четыре различных типа сайтов связывания катионов. [32] Сайт 1 расположен в центральной поре и находится в той же плоскости, что и ключевые селективные остатки D541. Считается, что сайт 2 присутствует примерно на 6-8 Å ниже сайта 1, за ним следует сайт 3, который расположен на центральной оси поры примерно на 6,8 Å ниже сайта 2. Считается, что сайты 2 и 3 взаимодействуют с частично гидратированными или экваториально-гидратированными. Калифорния 2+ ионы. Наконец, четыре симметричных сайта связывания катионов во внеклеточном вестибуле опосредуют рекрутирование катионов по направлению к внеклеточному вестибулу TRPV6 и называются сайтами рекрутирования. [32]
Проникновение ионов
[ редактировать ]Проводимость TRPV6 для двухвалентных катионов соответствует предпочтениям: Ca 2+ > Ба 2+ > Сэр 2+ > Мн 2 . Внутриклеточный магний 2+ ингибирует TRPV6 и способствует сильному внутреннему выпрямлению канала. [37] Активность поглощения TRPV6 ингибируется двухвалентным Pb. 2 , С 2+ , компакт-диск 2+ , Зн 2 , Ко 2+ , Фе 2+ , и трехвалентные катионы La 3+ , Фе 3+ , Б-г 3+ . Концентрация ионов для достижения ингибирования колеблется от 1 до 10 мкМ. [38] Белок TRPV6 конститутивен с одноканальной проводимостью 42-58 пс. [7] [39] При низком Са 2+ концентрации, один Ca 2+ ион связывается в селективном фильтре, образованном D541, и пропускает Na + проникновение. При высоком Са 2+ концентрация, Са 2+ проникновение происходит по механизму выбивания, который включает образование короткоживущих конформаций, включающих связывание трех Ca 2+ ионы к остатку D541. [39]
Шлюз канала
[ редактировать ]
Конформационные изменения, участвующие в открытии канала, связаны с остатком аланина 566 (A566) и происходят в спирали S6, выстилающей поры ( рис. 3 ). [39] Верхняя часть спирали S6 претерпевает спиральный переход от α к π, который заставляет нижнюю часть спирали поворачиваться на 100 градусов и отклоняться от оси поры на 11 градусов. [39] Это конформационное изменение перемещает нижнюю часть спирали, закрывающей пору, и тем самым увеличивает размер поры. Конформационное изменение изменяет остатки, обращенные к оси поры, и запускает образование новых субъединиц электростатических связей и солевых мостиков , которые компенсируют высокие энергетические затраты на неблагоприятный спиральный переход α-к-π, который происходит во время открытия канала. [39]
Регуляция с помощью фосфатидилинозитол-4,5-бисфосфата (PIP 2 ) и кальмодулина (CaM)
[ редактировать ]Приток Ca 2+ внутри клетки запускает механизмы отрицательной обратной связи для подавления активности TRPV6 и предотвращения Ca 2+ перегрузка. [9] Активность канала TRPV6 регулируется внутриклеточным уровнем фосфолипида фосфатидилинозитол-4,5-бисфосфата (PIP 2 ) и взаимодействием с Ca 2+ - кальмодулина (CaM). Комплекс [9] Истощение PIP 2 или связывания CaM инактивирует TRPV6. [40] [41] [42] [43] [44] Приток Ca 2+ в клетках, экспрессирующих TRPV6, активируется фосфолипаза C (PLC), которая, в свою очередь, гидролизует PIP 2 . Истощение уровней PIP 2 приводит к снижению активности каналов, поскольку большинству каналов TRP требуется этот липид для активации. [40] [43] [44] Липид PIP 2 может подавлять Ca 2+ -CaM-опосредованное ингибирование TRPV6. В целом, инактивация TRPV6 кальмодулином регулируется балансом внутриклеточного кальция. 2+ и PIP 2 . концентрация [40] [41] [42] [43] [44]
Взаимодействующие белки
[ редактировать ]Среди более 20 идентифицированных на данный момент взаимодействий TRPV6 функциональные последствия Ca 2+ Наиболее подробно охарактеризованы -связывающий белок кальмодулин (СаМ) и глюкуронидаза Клото [36, 37, 41, 42]. [34] [40] [41] [45] [46] Функциональные последствия активации канала TRPV6 суммированы в таблице ниже). [47]
| Интерактор | Последствие |
| БСПРЫ | Н/Д |
| Кальбиндин-Д 28к | Н/Д |
| Кальмодулин | Торможение |
| Циклофилин Б | Активация |
| НАХОДИТЬ | ПО 4- лиация |
| Я-МИД | Н/Д |
| Клото | Активация, гликозилирование (Asn-357) |
| НХЕРФ4 | Активация |
| НИПСНАП1 | Торможение |
| ОНЕМЕВШИЙ | Торможение |
| ПТЭН | Н/Д |
| ПТП1Б | ДеПО 4- лиация (Тир-161 и Тир-162) |
| РАБ11А | Активация, Увеличение уровня плазматической мембраны |
| РГС2 | Н/Д |
| РИР1 | Н/Д |
| С100А10 | Активация, Увеличение уровня плазматической мембраны |
| СРЦ | ПО 4 ляции (Тир-161, 162) |
| ТРПК1 | Сохраняется в отделении скорой помощи, торможение |
| ТРМЛ3 | Н/Д |
| ТРПВ5 | образование тетрамера, Создание нового канала |
Сокращения
Белковый интерактор
BSPRY: белок, содержащий домен B-Box и Spry; FYN: Fyn-киназа, принадлежащая к семейству киназ Src; I-MFA: ингибитор семейства Myo D; NHERF: Регуляторный фактор Na-обменника; NIPSNAP14-нитрофенилфосфатазный домен и ненейрональный гомолог 1 белка, подобного SNAP25; Онемение: мутация дрозофилы, которая удаляет большую часть сенсорных нейронов в развивающейся периферической нервной системе; PTP: протеин-тирозинфосфатаза; Rab11a: член семейства онкогенов РАН; RGS2: регулятор передачи сигналов G-белка 2; RyR1: рианодиновый рецептор 1; TRPC1: переходный рецепторный потенциал, канонический 1; TRPML3: временный рецепторный потенциал муколипина-3.
Физиологические функции
[ редактировать ]Калифорния 2+ Белки селективных каналов TRPV6 и TRPV5 взаимодействуют, поддерживая концентрацию кальция в определенных органах. [22] [48] TRPV6 действует как апикальный Ca. 2+ входные каналы, опосредующие трансклеточный транспорт этого иона в кишечник, плаценту и, возможно, в некоторые другие экзокринные органы. TRPV6 также играет важную роль в транспорте кальция от матери к плоду. [49] дифференцировка кератиноцитов, [50] и Ca 2+ гомеостаз в эндолимфатической системе вестибулярного аппарата, [51] [52] и поддержание мужской фертильности. [53] [54]
С а 2+ всасывание в кишечнике
[ редактировать ]Два маршрута Ка 2+ Распознаются абсорбция: парацеллюлярный транспорт и трансклеточный транспорт (см. рисунок 4 ). [55] с высоким содержанием Ca 2+ -диета способствует парацеллюлярному транспорту ионов по всей длине кишечника, позволяя им проходить между межклеточными плотными соединениями , соединяющими эпителиальные клетки. Напротив, в условиях, когда [Ca 2+ ] в просвете кишечника ниже по сравнению с его концентрацией в плазме (например, при низком диетическом питании Ca 2+ ), трансклеточный путь необходим для адекватного Ca 2+ поглощение. Три важных этапа трансклеточного кальция 2+ транспорт распознается: поступление Ca в клетку 2+ ион на апикальной стороне через TRPV6 (Шаг-1), связывание Ca 2+ ион с кальбиндином -D 9k (Шаг-2) и выход Ca 2+ с базолатеральной стороны через плазматическую мембрану Ca 2+ АТФаза (PMCA1b). [55] Гормон витамин D3 ( или 1,25(OH) 2D3 . кальции ) играет важную роль в опосредованном TRPV6 кишечном 2+ поглощение). [55]
Что 2+ реабсорбция в почках
[ редактировать ]В отличие от кишечника, где TRPV6 является привратником Ca 2+ Всасывание, трансклеточная реабсорбция этого иона в почках происходит через TRPV5. Хотя TRPV5 является признанным посредником в трансклеточной реабсорбции Ca 2+ В почках мыши с нокаутом TRPV6 (KO) также испытывают трудности с концентрацией мочи и проявляют гиперкальциурию. [56] Известно, что TRPV6 локализуется совместно с TRPV5 Calbindin-D 28K в апикальных доменах дистальных извитых канальцев и соединительных канальцев [20]. Мыши TRPV5 KO компенсируют Ca 2+ потеря за счет увеличения экспрессии TRPV6 в двенадцатиперстной кишке. [56] Более того, недавнее исследование, анализирующее чувствительные к витамину D гены в почках овцы , собаки и лошади , показало, что TRPV6, CalD 9k /calD 28k и PMCA могут быть основными путями, управляющими трансклеточным кальцием. 2+ транспортируется в почках овец, собак и лошадей. [57]
Материнско-плодовый Ca 2+ транспорт
[ редактировать ]TRPV6 играет незаменимую роль в плацентарном кальции. 2+ транспорт. [49] плода Пик минерализации костей приходится на поздние сроки беременности. На этом этапе кровь плода имеет более высокую концентрацию Са. 2+ по сравнению с материнской кровью, тем самым создавая условия, требующие активного трансклеточного транспорта кальция. 2+ от матери к плоду. [58] [59] Этот процесс очень важен, поскольку дефекты плацентарного транспорта кальция могут быть предшественниками кальция. 2+ синдромы дефицита и ограничения внутриутробного развития . [60] Экспрессия TRPV6 увеличивается в 14 раз в течение последних 4 дней гестационного мышей периода и совпадает с пиковой фазой минерализации костей плода. [49] Белок TRPV6 обильно экспрессируется в тканях плаценты млекопитающих. [49] [61] [62] [63] [64] Действительно, экспрессия TRPV6 примерно в 1000 раз выше по сравнению с TRPV5. В плаценте TRPV6 экспрессируется в трофобластах и синцитиотрофобластах . [14] [61] У мышей мРНК и белок TRPV6 экспрессируются во внутриплацентарном желточном мешке и висцеральном слое экстраплацентарного желточного мешка. [49] Самое главное, что плоды TRPV6 KO демонстрируют снижение на 40% 45 Что 2+ транспортная активность и резкое снижение массы золы (показатель здоровья костей плода). [49] Известно, что у людей стресс сдвига жидкости (FSS) в трофобластах индуцирует TRPV6-опосредованный Ca 2+ приток и способствует образованию микроворсинок посредством механизма, включающего Эзрин и Akt-фосфорилирование. [65]
Эпидидимальный Са 2+ регулирование и влияние на мужскую фертильность
[ редактировать ]Регуляция концентрации кальция в просвете придатка яичка имеет решающее значение для подвижности сперматозоидов . [66] TRPV6-опосредованное снижение люминального кальция 2+ концентрация в придатках яичка имеет решающее значение для мужской фертильности мышей. [53] Мыши TRPV6 KO или мыши, экспрессирующие версию канала TRPV6 с потерей функции ( Trpv6 Д541А гомозиготные мыши) имеют сильно нарушенную фертильность . [53] Мыши, экспрессирующие нефункциональный TRPV6, имеют в 10 раз более высокую концентрацию Ca. 2+ в просвете придатка яичка и Ca 2+ поглощение в этом пространстве снижается в 7–8 раз. [53] [54] Увеличение Ca 2+ концентрация ионов в просвете придатка яичка приводит к значительным нарушениям подвижности, способности к оплодотворению и жизнеспособности сперматозоидов в TRPV6. Д541А мыши. [53] [54] Похоже, что TRPV6 и трансмембранный хлоридный канал 16A ( TMEM16A ) действуют совместно, снижая люминальную концентрацию Ca. 2+ в просвете придатка яичка. [67]
Здоровье костей
[ редактировать ]В условиях неоптимального диетического кальция 2+ , нормальный уровень кальция в сыворотке у мышей TRPV6 KO поддерживается за счет костей. [68] [69] TRPV6 играет важную роль в остеокластах, но не в остеобластах. [68] [69] У мышей истощение TRPV6 приводит к усилению дифференцировки остеокластов. [29] тогда как TRPV5 необходим для правильной остеокластической резорбции кости. [68]
Дифференциация кератиноцитов
[ редактировать ]Дифференцировка кератиноцитов регулируется переключением кальция — процессом, который влечет за собой приток кальция. 2+ в кератиноците, что вызывает широкие транскрипционные изменения, необходимые для образования десмосом , их стратификации и ороговения. [70] У мышей TRPV6 KO наблюдаются более тонкие слои рогового слоя , а у 20% мышей также наблюдаются алопеция и дерматит . [56] Замалчивание TRPV6 ухудшает Ca 2+ -опосредованная дифференцировка первичных кератиноцитов человека и подавляет маркеры дифференцировки, такие как инволюкрин , трансглутаминаза-1 и цитокератин-10 . Гормон 1,25-дигидроксивитамин-D3 активирует TRPV6 в кератиноцитах и запускает Ca 2+ приток. Это, в свою очередь, индуцирует экспрессию путей, специфичных для дифференцировки кератиноцитов. [50]
Роль во внутреннем ухе
[ редактировать ]Белки TRPV5 и TRPV6 экспрессируются в нескольких областях внутреннего уха , а также в первичных культурах эпителия протока полукружного канала (SCCD). [51] [52] Некоторые исследования показали, что TRPV5 и TRPV6 необходимы для снижения уровня кальция. 2+ концентрация в просвете эндолимфы млекопитающих , что необходимо для нормального слуха и равновесия. [51] [52] [71]
Маточная и плацентарная экспрессия TRPV6 и последствия при беременности
[ редактировать ]и в эндометрии матке у млекопитающих. Сообщалось об экспрессии TRPV6 [72] [73] [74] Считается, что экспрессия TRPV6 в матке у грызунов гормонально регулируется 17β-эстрадиолом и прогестероном . У грызунов мРНК TRPV6 экспрессируется в лабиринтной и губчатой зоне, а также в участках матки, не прикрепленных к плаценте. Стадия беременности является важным регулятором экспрессии TRPV6. Снижение экспрессии TRPV5/6 и, как следствие, снижение содержания Ca 2+ Считается, что транспорт меняет пролиферативный профиль трофобластов человека; процесс, который, в свою очередь, связан с развитием преэклампсии . [73] Такое сопоставление экспрессии TRPV6 и ее жесткой регуляции половыми гормонами во время беременности позволяет предположить, что этот белок может быть важен для имплантации эмбриона , однако убедительных доказательств этой связи не существует. [72] [73] [74]
Последствия для болезней человека
[ редактировать ]Транзиторный неонатальный гиперпаратиреоз
[ редактировать ]Потеря TRPV6 в плаценте мышей серьезно ухудшает содержание кальция. 2+ транспортируется через трофобласт и замедляет рост эмбриона, вызывает кальцификацию костей и ухудшает развитие костей. Считается, что у людей недостаточный транспорт от матери к плоду, вызванный патогенными геномными вариантами TRPV6, является причиной дефектов скелета, наблюдаемых в отдельных сообщениях о случаях транзиторного неонатального гиперпаратиреоза (TNHP). Считается, что эти варианты нарушают локализацию белка в плазматической мембране. Секвенирование экзома у младенца с тяжелой антенатальной . торакальной недостаточностью с сопутствующими аномалиями скелета плода указывает на критическую роль TRPV6 в транспорте от матери к плоду Исследование показало, что сложные гетерозиготные варианты TRPV6 приводят к тяжелой недостаточности минерализации и тяжелой дисплазии скелета плода. [75] [76] [77]
Хронический панкреатит
[ редактировать ]Недавние данные показывают, что естественная потеря вариантов функции TRPV6 предрасполагает определенные демографические группы к хроническому панкреатиту (ХП) из-за нарушения регуляции гомеостаза кальция в клетках поджелудочной железы. [78] [79] Исследования секвенирования среди пациентов с хроническим панкреатитом выявили наличие 33 миссенс-вариантов и 2 нонсенс -вариантов, предрасполагающих японских , немецких и французских пациентов к более высокому риску ХП. [79] В целом, эти исследования показали, что вызывающие заболевание геномные варианты TRPV6 с потерей функции чрезмерно представлены у пациентов с ХП из Германии, Франции, Китая и Японии по сравнению с контрольной группой в их соответствующих группах. [78] [79] Считается, что варианты с потерей функции нарушают транспорт кальция в поджелудочной железе, снижая общий уровень белка и/или подвергая риску Ca. 2+ активность восприятия на канале. [79]
Образование камней в почках
[ редактировать ]Роль TRPV6 в образовании камней в почках была доказана в ходе исследований секвенирования, проведенных на группе из 170 пациентов в Швейцарии . [80] Исследования показали, что частота гаплотипа усиления функции TRPV6 значительно выше у Ca. 2+- камни образующие ( нефролитиаз ) по сравнению с не образующими камни. Наблюдаемые фенотипы гиперкальциурии в исследованиях на животных и исследованиях однонуклеотидных полиморфизмов (SNP) TRPV6 позволяют предположить, что гаплотип TRPV6 может быть важным фактором риска абсорбционной и почечной гиперкальциурии (камни в почках из-за нарушения кишечной абсорбции и почечной реабсорбции соответственно). Более низкая заболеваемость мочекаменной болезнью у афроамериканцев и относительно более высокая распространенность предкового гаплотипа позволяют предположить теорию, согласно которой этот гаплотип обеспечивает преимущество повышенного содержания кальция. 2+ реабсорбция у этой группы населения и снижает частоту образования камней в почках. [14] [20] [22] [80]
Костно-резорбтивные заболевания
[ редактировать ]У мышей TRPV6 KO проявляются симптомы, подобные остеопорозу, такие как снижение минеральной плотности костей и гиперкальциурия. [56] Гормон эстроген , дефицит которого связан с постменопаузальным остеопорозом , также регулирует экспрессию TRPV6 у людей. Действительно, более низкая абсорбция кальция, наблюдаемая у пожилых женщин в постменопаузе, объясняется снижением TRPV6. [81] С-концевая часть сорицидина представляет собой препарат, ингибирующий Ca. 2+ -активность поглощения путем связывания с TRPV6. Доклинические исследования этого препарата показали большие перспективы в лечении костно-резорбтивных заболеваний. [28]
Высокая степень сходства между симптомами наследственного витамин-D-резистентного рахита (HVDRR) и наблюдаемыми фенотипами у мышей TRPV6 KO побудила некоторых экспертов постулировать патологические связи между заболеванием и дисфункцией TRPV6. [48] TRPV6 играет важную хондропротекторную роль, регулируя многочисленные аспекты функции хондроцитов , такие как секреция внеклеточного матрикса , высвобождение ферментов, разрушающих матрикс, пролиферация клеток и апоптоз . [82] Кроме того, мыши с нокаутом TRPV6 демонстрируют множественные фенотипы остеоартрита (ОА), такие как хряща фибрилляция , покраснение и потеря протеогликанов . [82]
Синдром Пендреда
[ редактировать ]Ген дисфункции Slc26a4 связан с синдромом Пендреда — генетическим заболеванием, которое приводит к синдромальной глухоте у детей. [71] [83] Заболевание вызвано мутациями, при которых нарушается функция кодируемого белка пендрина - аниона Cl. − /HCO3 − обменник выражен во внутреннем ухе. [71] [83] Считается, что потеря функции этого гена снижает значение pH эндолимфы млекопитающих и ухудшает содержание кальция. 2+ поглощение через TRPV5 и TRPV6. [83] Это, в свою очередь, может предотвратить поглощение Ca 2+ и ухудшает люминальное снижение Ca 2+ концентрация в эндолимфатической системе уха. [71] [83]
Рак
[ редактировать ]Сверхэкспрессия TRPV6 была подтверждена при толстой кишки, паращитовидной железы , поджелудочной железы и щитовидной железы. раке [23] тогда как, как сообщается, его экспрессия снижается при раке пищевода , [84] немелкоклеточный рак легкого , [85] и рак почки . [86] TRPV6 считается онкоканалом, который, как предполагается, опосредует прогрессирование рака, запуская Ca 2+ -индуцированные аберрации в молекулярных драйверах, регулирующих такие процессы, как клеточный цикл , апоптоз и миграция ; тем самым давая раковым клеткам преимущества в пролиферативной и выживаемости. [25] [28] [87] Сверхэкспрессия TRPV6 сильно коррелирует с патологической стадией , степенью опухоли , внепростатической инвазией, в лимфатические узлы метастазами и устойчивостью к андроген -таргетной терапии при раке предстательной железы . [14] [23] [87] [88] Экспрессия TRPV6 рекламировалась как прогностический маркер прогрессирующего рака простаты, поскольку ее экспрессия сильно зависит от степени опухоли. [87] [88] Экспрессия TRPV6 значительно повышена в ткани аденокарциномы молочной железы по сравнению с нормальной тканью молочной железы. [89] [90] Об экспрессии TRPV6 сообщалось во многих линиях клеток рака молочной железы и линиях клеток рака простаты. [87] [91] [92] [93] Клеточные линии рака простаты PC-3 и LnCAP сверхэкспрессируют TRPV6 по сравнению с доброкачественными эпителиальными клетками PrEC и BPH-1. [87] Замалчивание TRPV6 в клетках рака предстательной железы снижает скорость пролиферации, накопление S-фазы и экспрессию опухолевого маркера экспрессии ядерного антигена пролиферирующих клеток (PCNA). [92] Считается, что сверхэкспрессия TRPV6 индуцирует аберрантный Ca. 2+- поглощение в линии рака простаты и активация транскрипционного фактора ядерного фактора активированных Т-клеток (NFAT). [92]
Экспрессия TRPV6 усиливается эстрогеном, прогестероном и эстрадиолом в клеточной линии рака молочной железы T47D . [90] В соответствии с этими наблюдениями рецепторов эстрогена антагонист тамоксифен снижает экспрессию TRPV6 в клетках T47D и подавляет Ca. 2+ -поглощение канала как в ER-положительных , так и в ER-негативных клеточных линиях рака молочной железы. [94] Сверхэкспрессия TRPV6 связана с раком толстой кишки на ранней стадии , а ее подавление при раке толстой кишки вызывает апоптоз и ингибирует пролиферацию раковых клеток. [95] С точки зрения механизма, мутации в кальмодулин-связывающих доменах каналов TRPV6 придают инвазивные свойства клеткам аденокарциномы толстой кишки. [96] белки p38α и GADD45α усиливают передачу сигналов экспрессии TRPV6 в клетках рака толстой кишки SW480 за счет усиления передачи сигналов витамина D. Считается, что [97] Сообщалось, что TRPV6 усиливает индуцированную инсулиноподобными факторами роста (IGF) передачу сигналов PI3K - PDK1 - Akt при раке толстой кишки человека и способствует развитию рака толстой кишки. [98]
TRPV6 активируется в тканях первичного рака у пациентов с раком поджелудочной железы и способствует пролиферации нейроэндокринных опухолей поджелудочной железы по NFAT-зависимым механизмам. [99] Замалчивание TRPV6 индуцирует апоптоз и остановку клеточного цикла в клетках рака поджелудочной железы и ингибирует их инвазию, пролиферацию и миграцию. [100] Принудительная экспрессия TRPV6 в клетках рака желудка увеличивает их чувствительность к капсаицином апоптозу, индуцированному , тогда как siRNA -опосредованное замалчивание канала подавляет эту чувствительность. [101] снижение уровня TRPV6 при карциноме пищевода Было высказано предположение, что является прогностическим маркером специфической выживаемости у пациентов, страдающих раком пищевода . [102] Низкая совместная экспрессия TRPV5 и TRPV6 считается прогностическим маркером плохой безрецидивной выживаемости при немелкоклеточном раке легкого . [85]
Фармакологическое таргетирование
[ редактировать ]Известно, что несколько химических ингибиторов ингибируют TRPV6. Некоторые соединения, продемонстрировавшие ингибирующую активность в отношении TRPV6, включают TH-1177, 2-аминоэтоксидифенилборат (2-APB), производное 2-APB 22b, эконазол , миконазол , производное пиперазина Cis-22a, капсаицин , Δ 9 - тетрагидроканнабиварин , ксестоспонгин C, лидокаин , наночастицы с золотыми клетками (PTX-PP@Au NPs) и сорцидин C-13 (SOR-C13). синтетический пептид [28] Среди различных стратегий ингибирования, протестированных до сих пор, наиболее многообещающим оказался пептид SOR-C13, состоящий из 13 аминокислот. Этот пептид из 13 аминокислот, полученный из пептида из 54 аминокислот, обнаруженного в паралитическом яде северной короткохвостой бурозубки ( Blarina brevicauda ), снижает рост рака на клеточных и животных моделях. Этот противораковый препарат недавно завершил фазу I клинического исследования безопасности, в котором приняли участие 23 пациента с распространенными солидными опухолями эпителиального происхождения, не реагирующими на все стандартные методы лечения. [28]
Регулирование
[ редактировать ]Регуляцию TRPV6 можно исследовать главным образом в контексте ее физиологических, гормональных и молекулярных факторов. [22] Гормональная регуляция TRPV6 охарактеризована наиболее подробно. его регуляция гормоном витамином D3 и В связи с этим достаточно подробно изучена половыми гормонами. Исследования на грызунах показывают, что канал TRPV6 регулируется широким спектром физиологических факторов, таких как диета, возраст, пол, беременность, лактация, половые гормоны, физические упражнения, возраст и пол. Некоторые биологические и фармакологические агенты, которые, как известно, регулируют TRPV6, включают глюкокортикоиды , иммунодепрессанты и диуретики . [22]
Витамин Д
[ редактировать ]Множественные эксперименты по дозозависимому и временному изменению на грызунах и клеточных линиях рака толстой кишки убедительно показали, что мРНК TRPV6 активно индуцируется этим витамином D в чрезвычайно низких концентрациях. [103] [104] по меньшей мере пять элементов ответа на витамин D (VDRE) в положениях -1,2, -2,1, -3,5, -4,3 и -5,5 т.п.н. относительно сайта начала транскрипции (TSS). В транскриптах TRPV6 было идентифицировано [105] Среди этих пяти сайтов VDRE в положениях -1,2, -2,1 и -4,3 т.п.н. значительно более чувствительны к 1,25-(OH) 2D3 OH) 2D3 ( по сравнению с VDRE, расположенными в -3,5 и -5,5 т.п.н., которые, по-видимому, не реагируют на 1,25- . вносят существенный вклад в регуляцию транскрипции, опосредованную витамином D, в кишечнике. [105] С точки зрения механизма, транскрипция TRPV6 инициируется в ответ на передачу сигналов, опосредованную рецептором витамина D (VDR), хотя нельзя исключать и другие непрямые механизмы. [104] [106] Важные этапы регуляции транскрипции, опосредованной витамином D, включают 1) связывание витамина D с родственным ему рецептором витамина D (VDR), 2) транслокацию гетеродимерного комплекса рецептор витамина D (VDR) -ретиноид X-рецептора в ядре , 3) связывание VDR. Комплекс -RXR на промоторе гена TRPV6 , 4) рекрутирование коактиватора стероидного рецептора 1 и РНК-полимеразы II на промоторе и 5) активация транскрипции, опосредованная гистона H4 событиями ацетилирования . [107]
Диета
[ редактировать ]Уровень Са 2+ и витамин D в рационе являются наиболее важными регуляторами экспрессии TRPV6. [104] Считается, что экспрессия TRPV6 сильно модулируется для точной настройки Ca. 2+ всасывание из рациона, особенно в условиях, когда диетический Ca 2+ доступность низкая. [103] [104] У грызунов ограничение Ca 2+ наличие в рационе вызывает резкое усиление экспрессии TRPV6 в двенадцатиперстной кишке. [103] [104] Приток кальция из рациона и его последующее связывание с кальбиндином-D 9k могут быть лимитирующим этапом, который модулирует витамин D-зависимую регуляцию TRPV6. [108] Когда диетический Ca 2+ недостаточно, нормальный уровень кальция в крови у мышей TRPV6 KO поддерживается за счет костей. [68] [69] У многих линий грызунов генетические вариации мРНК TRPV6, кальбиндина-D 9k , PMCA1b влияют на абсорбцию кальция в кишечнике и его влияние на плотность костного мозга. [109]
Беременность и лактация
[ редактировать ]Дуоденальная экспрессия транскриптов TRPV6 усиливается у мышей WT и VDR KO во время беременности и лактации . [110] Гормон пролактин усиливает транскрипцию TRPV6 и способствует увеличению содержания кальция в кишечнике. 2+ абсорбция у кормящих и беременных крыс, возможно, как адаптивный механизм преодоления потери минерализации костей во время лактации. [111]
Старение
[ редактировать ]Кишечная экспрессия TRPV6 у мышей резко варьируется в зависимости от возраста и относительного расположения тканей. [112] Дуоденальная экспрессия TRPV6 не обнаруживается на P1 и увеличивается в 6 раз по мере старения мышей до P14. Аналогичным образом, экспрессия также варьируется с возрастом в тощей кишке , где уровни TRPV6 увеличиваются от P1 до P14, становятся слабыми в возрасте 1 месяца и становятся необнаружимыми у более старых мышей. [112] Экспрессия TRPV6 у старых крыс (12 месяцев) как минимум на 50% ниже по сравнению с более молодыми крысами (2 месяца). [104] У мышей WT и VDR KO возрастное снижение кишечной абсорбции Ca 2+ сопровождается снижением дуоденальной экспрессии TRPV6. [113]
Половые гормоны
[ редактировать ]Половые гормоны играют важную роль в регуляции TRPV6. По сравнению с мышами-самцами, у мышей-самок наблюдается в 2 раза более высокое увеличение экспрессии мРНК TRPV6 в двенадцатиперстной кишке после лечения витамином D. [ нужна ссылка ] Считается, что дифференциальная регуляция TRPV6, связанная с половыми гормонами, у разных полов коррелирует с различиями в относительном риске остеопороза у пожилых женщин в постменопаузе, у которых, как сообщается, наблюдается более низкая экспрессия TRPV6 и VDR по сравнению с мужчинами. [81]
Лечение эстрогенами усиливает транскрипты TRPV6 в 8 раз у мышей VDR KO и в 4 раза у мышей с удаленными яичниками . [106] KO наблюдалось снижение мРНК TRPV6 более чем на 50% У мышей с рецептором эстрогена α . [110] Считается, что эстроген может дифференциально регулировать Ca. 2+ абсорбция в двенадцатиперстной кишке за счет увеличения экспрессии TRPV6 через ERα. [114] Антипрогестероновый агент RU486 и антиэстрогенный агент ICI 182,780 подавляют экспрессию TRPV6 у грызунов за счет их соответствующего антагонистического действия на рецепторы прогестерона и эстрогена . [115] эстроген, прогестерон и дексаметазон Известно, что усиливают экспрессию TRPV6 в коре головного мозга и гипоталамусе мышей, что позволяет предположить потенциальное участие TRPV6 в абсорбции кальция в головном мозге . [116]
Глюкокортикоиды
[ редактировать ]Подкожное введение глюкокортикоидов дексаметазона индуцирует как почечную, так и кишечную экспрессию TRPV6 у мышей в течение 24 часов, тогда как пероральное применение преднизолона снижает TRPV6, что также сопровождается снижением содержания кальция. 2+ всасывание в двенадцатиперстной кишке. [117] [118] Кишечная регуляция TRPV6 в ответ на глюкокортикоиды, по-видимому, зависит от VDR. [117] [118] Фермент сыворотки и глюкокортикоид-регулируемая киназа 1 ( SKG1 ) регулируют экспрессию TRPV6 путем усиления фосфатидилинозитол-3-фосфат-5-киназы PIKfyve (PIP5K3). [119] Эта киназа имеет решающее значение для генерации вторичного мессенджера PIP 2 , известного липидного активатора TRPV6. [119]
- ТРПВ
- ТРПВ5
- кальциевые каналы
- поглощение кальция
- трансклеточный путь
- литниковый механизм
- кальмодулин
- транспорт матери и плода
- транзиторный неонатальный гиперпаратиреоз
- хронический панкреатит
- камни в почках
- рак
Примечания
[ редактировать ]Версия этой статьи 2020 года была обновлена внешним экспертом в рамках модели двойной публикации. Соответствующая академическая рецензируемая статья была опубликована в журнале Gene и может цитироваться как: Винаяк Хаттар, Линюнь Ван, Цзи-Бин Пэн (5 апреля 2022 г.). «Кальций-селективный канал TRPV6: структура, функции и значение для здоровья и болезней». Джин . Серия обзоров Gene Wiki. 817 . дои : 10.1016/J.GENE.2022.146192 . ISSN 0378-1119 . ПМИД 35031425 . Викиданные Q110874871 . |
Ссылки
[ редактировать ]- ^ Jump up to: а б с ENSG00000165125 GRCh38: выпуск Ensembl 89: ENSG00000276971, ENSG00000165125 — Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000029868 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с Венкатачалам К., Монтелл С. (2007). «Каналы ГТО» . Ежегодный обзор биохимии . 76 : 387–417. doi : 10.1146/annurev.biochem.75.103004.142819 . ПМК 4196875 . ПМИД 17579562 .
- ^ Монтелл С., Бирнбаумер Л., Флокерзи В., Биндельс Р.Дж., Бруфорд Э.А., Катерина М.Дж. и др. (февраль 2002 г.). «Единая номенклатура суперсемейства катионных каналов TRP» . Молекулярная клетка . 9 (2): 229–31. дои : 10.1016/s1097-2765(02)00448-3 . OCLC 1106536919 . ПМИД 11864597 .
- ^ Jump up to: а б Юэ Л., Пэн Дж.Б., Хедигер М.А., Клэпхэм Д.Э. (апрель 2001 г.). «CaT1 проявляет поровые свойства кальциевого канала, активируемого высвобождением кальция». Природа . 410 (6829): 705–9. Бибкод : 2001Natur.410..705Y . дои : 10.1038/35070596 . ПМИД 11287959 . S2CID 4404582 .
- ^ Jump up to: а б с д и Ельшанская М.В., Надеждин К.Д., Курникова М.Г., Соболевский А.И. (февраль 2020 г.). «Структура и функция кальций-селективного канала TRP TRPV6» . Журнал физиологии . 599 (10): 2673–2697. дои : 10.1113/JP279024 . ПМЦ 7689878 . ПМИД 32073143 .
- ^ Jump up to: а б с д Пэн Дж.Б., Сузуки Ю., Гимеси Дж., Хедигер М.А. (2018). «Кальций-селективные каналы TRPV5 и TRPV6». Каналы входа кальция в невозбудимые клетки . Методы передачи сигналов. Бока-Ратон, Флорида: CRC Press/Тейлор и Фрэнсис. стр. 241–274. дои : 10.1201/9781315152592-13 . ISBN 978-1-315-15259-2 . ПМИД 30299660 .
- ^ Jump up to: а б с д Пэн Дж.Б., Чен XZ, Бергер У.В., Васильев П.М., Цукагути Х., Браун Э.М., Хедигер М.А. (август 1999 г.). «Молекулярное клонирование и характеристика каналообразного транспортера, опосредующего всасывание кальция в кишечнике» . Журнал биологической химии . 274 (32): 22739–46. дои : 10.1074/jbc.274.32.22739 . ПМИД 10428857 . S2CID 23616713 .
- ^ Jump up to: а б Пэн Дж.Б., Чен XZ, Бергер У.В., Веремович С., Мортон CC, Василев П.М. и др. (ноябрь 2000 г.). «Человеческий белок-переносчик кальция CaT1». Связь с биохимическими и биофизическими исследованиями . 278 (2): 326–32. дои : 10.1006/bbrc.2000.3716 . ПМИД 11097838 .
- ^ Jump up to: а б с Вебер К., Эрбен Р.Г., Рамп А., Адамски Дж. (декабрь 2001 г.). «Геновая структура и регуляция мышиных эпителиальных кальциевых каналов ECaC1 и 2». Связь с биохимическими и биофизическими исследованиями . 289 (5): 1287–94. дои : 10.1006/bbrc.2001.6121 . ПМИД 11741335 .
- ^ Jump up to: а б с Хендероп Дж.Г., Веннекенс Р., Мюллер Д., Пренен Дж., Другманс Г., Биндельс Р.Дж., Нилиус Б. (декабрь 2001 г.). «Функция и экспрессия семейства эпителиальных Ca (2+) каналов: сравнение ECaC1 и 2 млекопитающих» . Журнал физиологии . 537 (Часть 3): 747–61. дои : 10.1113/jphysicalol.2001.012917 . ПМК 2278984 . ПМИД 11744752 .
- ^ Jump up to: а б с д и ж г час Виссенбах У., Нимейер Б.А., Фиксемер Т., Шнайдевинд А., Трост С., Кавали А. и др. (июнь 2001 г.). «Экспрессия CaT-подобного, нового кальций-селективного канала, коррелирует со злокачественностью рака простаты» . Журнал биологической химии . 276 (22): 19461–8. дои : 10.1074/jbc.m009895200 . ПМИД 11278579 . S2CID 25833991 .
- ^ Клэпхэм Д.Э., Джулиус Д., Монтелл С., Шульц Г. (декабрь 2005 г.). «Международный союз фармакологии. XLIX. Номенклатура и структурно-функциональные связи временных рецепторных потенциальных каналов». Фармакологические обзоры . 57 (4): 427–50. дои : 10.1124/пр.57.4.6 . ПМИД 16382100 . S2CID 17936350 .
- ^ Jump up to: а б Мюллер Д., Хендероп Дж.Г., Мерккс Г.Ф., ван Ос Ч., Биндельс Р.Дж. (август 2000 г.). «Геновая структура и хромосомное картирование эпителиальных кальциевых каналов человека». Связь с биохимическими и биофизическими исследованиями . 275 (1): 47–52. дои : 10.1006/bbrc.2000.3227 . ПМИД 10944439 .
- ^ Jump up to: а б с Фехер-Трост К., Вайсгербер П., Виссенбах У. (2014). «Каналы TRPV6». Катионные каналы переходного рецепторного потенциала млекопитающих (TRP) . Справочник по экспериментальной фармакологии. Том. 222. Берлин, Гейдельберг: Springer Berlin Heidelberg. стр. 359–84. дои : 10.1007/978-3-642-54215-2_14 . ISBN 978-3-642-54214-5 . ПМИД 24756713 .
- ^ Jump up to: а б Фехер-Трост С., Виссенбах Ю., Бек А., Шалковски П., Стергер С., Дорр Дж. и др. (июнь 2013 г.). «Белок TRPV6 in vivo начинается с триплета, отличного от AUG, декодируемого как метионин, перед канонической инициацией в AUG» . Журнал биологической химии . 288 (23): 16629–44. дои : 10.1074/jbc.M113.469726 . ПМЦ 3675598 . ПМИД 23612980 .
- ^ Jump up to: а б с Флорес-Альдама Л., Вандевеге М.В., Завала К., Коленсо К.К., Гонсалес В., Браучи С.Е., Опасо Х.К. (май 2020 г.). «Эволюционный анализ выявляет независимое происхождение репертуара генов и структурных мотивов, связанных с быстрой инактивацией кальций-селективных каналов TRPV» . Научные отчеты . 10 (1): 8684. Бибкод : 2020NatSR..10.8684F . дои : 10.1038/s41598-020-65679-6 . ПМК 7250927 . ПМИД 32457384 .
- ^ Jump up to: а б с д и ж Эйки Дж.М., Суонсон В.Дж., Мадой Дж., Эберл М., Шрайвер, доктор медицины (июль 2006 г.). «TRPV6 демонстрирует необычные модели полиморфизма и дивергенции в популяциях по всему миру» . Молекулярная генетика человека . 15 (13): 2106–13. дои : 10.1093/hmg/ddl134 . ПМИД 16717058 .
- ^ Jump up to: а б с Соэдзима М., Тачида Х., Кода Ю. (февраль 2009 г.). «Анализ последовательности человеческого TRPV6 предполагает положительный отбор за пределами Африки». Биохимическая генетика . 47 (1–2): 147–53. дои : 10.1007/s10528-009-9222-x . ПМИД 19169858 . S2CID 39897282 .
- ^ Jump up to: а б с д и Пэн Дж.Б. (2011). «TRPV5 и TRPV6 в трансклеточном транспорте Ca2+: регуляция, дупликация генов и полиморфизмы в африканских популяциях». Потенциальные каналы временных рецепторов . Достижения экспериментальной медицины и биологии. Том. 704. стр. 239–75. дои : 10.1007/978-94-007-0265-3_14 . ISBN 978-94-007-0264-6 . ПМИД 21290300 .
- ^ Jump up to: а б с д Чжуан Л., Пэн Дж.Б., Тоу Л., Таканага Х., Адам Р.М., Хедигер М.А., Фриман М.Р. (декабрь 2002 г.). «Кальций-селективный ионный канал, CaT1, локализован апикально в эпителии желудочно-кишечного тракта и аберрантно экспрессируется при злокачественных новообразованиях человека» . Лабораторные исследования; Журнал технических методов и патологии . 82 (12): 1755–64. дои : 10.1097/01.lab.0000043910.41414.e7 . ПМИД 12480925 . S2CID 6053178 .
- ^ Jump up to: а б «Краткая информация об экспрессии белка TRPV6» . Атлас белков человека . Проверено 1 августа 2020 г.
- ^ Jump up to: а б Легенький В, Рафаэль М, Преварская Н (март 2012 г.). «Роль канала TRPV6 при раке» . Журнал физиологии . 590 (6): 1369–76. дои : 10.1113/jphysicalol.2011.225862 . ПМЦ 3382328 . ПМИД 22331416 .
- ^ Хирнет Д., Олауссон Дж., Фехер-Трост С., Бёддинг М., Настаинчик В., Виссенбах У. и др. (май 2003 г.). «Ген TRPV6, кДНК и белок». Клеточный кальций . 33 (5–6): 509–18. дои : 10.1016/s0143-4160(03)00066-6 . ПМИД 12765696 .
- ^ Нидженхейс Т., Хендероп Дж.Г., ван дер Кемп А.В., Биндельс Р.Дж. (ноябрь 2003 г.). «Локализация и регуляция эпителиального Са2+-канала TRPV6 в почках» . Журнал Американского общества нефрологов . 14 (11): 2731–40. doi : 10.1097/01.asn.0000094081.78893.e8 . ПМИД 14569082 .
- ^ Jump up to: а б с д и Стюарт Дж. М. (2020). «TRPV6 как мишень для терапии рака» . Журнал рака . 11 (2): 374–387. дои : 10.7150/jca.31640 . ПМК 6930427 . ПМИД 31897233 .
- ^ Jump up to: а б Чен Ф, Ни Б, Ян Ё, Йе Т, Чен А (2014). «Нокаут TRPV6 вызывает остеопению у мышей за счет увеличения дифференцировки и активности остеокластов» . Клеточная физиология и биохимия . 33 (3): 796–809. дои : 10.1159/000358653 . ПМИД 24686448 . S2CID 19539099 .
- ^ ван дер Эрден BC, Вайсгербер П., Фратцль-Зельман Н., Олауссон Дж., Хоэндероп Дж.Г., Шредерс-Кедам М. и др. (май 2012 г.). «Канал временного рецепторного потенциала TRPV6 динамически экспрессируется в костных клетках, но не имеет решающего значения для минерализации костей у мышей». Журнал клеточной физиологии . 227 (5): 1951–9. дои : 10.1002/jcp.22923 . ПМИД 21732366 . S2CID 7094759 .
- ^ Литтл Р., Муймо Р., Робсон Л., Харрис К., Грабовски П.С. (29 ноября 2011 г.). «Ионный канал транзиторного рецепторного потенциала TRPV6 экспрессируется на низких уровнях в остеобластах и играет небольшую роль в поглощении кальция остеобластами» . ПЛОС ОДИН . 6 (11): e28166. Бибкод : 2011PLoSO...628166L . дои : 10.1371/journal.pone.0028166 . ПМЦ 3226639 . ПМИД 22163264 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Саотоме К., Сингх А.К., Ельшанская М.В., Соболевский А.И. (июнь 2016 г.). «Кристаллическая структура эпителиального кальциевого канала TRPV6» . Природа . 534 (7608): 506–11. Бибкод : 2016Natur.534..506S . дои : 10.1038/nature17975 . ПМК 4919205 . ПМИД 27296226 .
- ^ Сингх А.К., Саотоме К., Соболевский А.И. (сентябрь 2017 г.). «Обмен трансмембранных доменов в эпителиальном кальциевом канале TRPV6» . Научные отчеты . 7 (1): 10669. Бибкод : 2017NatSR...710669S . дои : 10.1038/s41598-017-10993-9 . ПМЦ 5587609 . ПМИД 28878326 .
- ^ Jump up to: а б Лу П., Борос С., Чанг К., Бинделс Р.Дж., Хендероп Дж.Г. (ноябрь 2008 г.). «Бета-глюкуронидаза клото исключительно активирует эпителиальные Ca2+-каналы TRPV5 и TRPV6» . Нефрология, Диализ, Трансплантация . 23 (11): 3397–402. дои : 10.1093/ndt/gfn291 . hdl : 2066/70822 . ПМИД 18495742 .
- ^ Jump up to: а б с Овсяник Г., Талавера К., Воетс Т., Нилиус Б. (январь 2006 г.). «Проницаемость и избирательность каналов ГТО». Ежегодный обзор физиологии . 68 (1): 685–717. doi : 10.1146/annurev.phyol.68.040204.101406 . ПМИД 16460288 .
- ^ Jump up to: а б с Воетс Т., Янссенс А., Другманс Г., Нилиус Б. (апрель 2004 г.). «Архитектура внешних пор Ca2+-селективного канала TRP» . Журнал биологической химии . 279 (15): 15223–30. дои : 10.1074/jbc.m312076200 . ПМИД 14736889 . S2CID 23827272 .
- ^ Воетс Т., Янссенс А., Пренен Дж., Дрогманс Г., Нилиус Б. (март 2003 г.). «Mg2+-зависимое ворота и сильная внутренняя выпрямление катионного канала TRPV6» . Журнал общей физиологии . 121 (3): 245–60. дои : 10.1085/jgp.20028752 . ПМК 2217333 . ПМИД 12601087 .
- ^ Ковач Г., Данко Т., Бержерон М.Дж., Балаж Б., Сузуки Ю., Жембери А., Хедигер М.А. (январь 2011 г.). «Катионы тяжелых металлов проникают в эпителиальный катионный канал TRPV6». Клеточный кальций . 49 (1): 43–55. дои : 10.1016/j.ceca.2010.11.007 . ПМИД 21146870 . S2CID 41086787 .
- ^ Jump up to: а б с д и Сингх А.К., Саотоме К., Соболевский А.И. (сентябрь 2017 г.). «Обмен трансмембранных доменов в эпителиальном кальциевом канале TRPV6» . Научные отчеты . 7 (1): 10669. Бибкод : 2017NatSR...710669S . bioRxiv 10.1101/141523 . дои : 10.1038/s41598-017-10993-9 . ПМЦ 5587609 . ПМИД 28878326 .
- ^ Jump up to: а б с д Цао С, Захарян Э, Борбиро И, Рохач Т (февраль 2013 г.). «Взаимодействие между кальмодулином и фосфатидилинозитол-4,5-бисфосфатом в Ca2+-индуцированной инактивации временных рецепторных потенциальных ваниллоидных 6-каналов» . Журнал биологической химии . 288 (8): 5278–90. дои : 10.1074/jbc.M112.409482 . ПМК 3581402 . ПМИД 23300090 .
- ^ Jump up to: а б с Бёддинг М., Флокерци В. (август 2004 г.). «Са2+-зависимость Са2+-селективного канала TRPV6» . Журнал биологической химии . 279 (35): 36546–52. дои : 10.1074/jbc.m404679200 . ПМИД 15184369 . S2CID 22842694 .
- ^ Jump up to: а б Ламберс Т.Т., Вайдема А.Ф., Нилиус Б., Хендероп Дж.Г., Биндельс Р.Дж. (июль 2004 г.). «Регуляция эпителиального Ca2(+)-канала TRPV6 мыши с помощью Ca(2+)-сенсора кальмодулина» . Журнал биологической химии . 279 (28): 28855–61. дои : 10.1074/jbc.m313637200 . hdl : 2066/57723 . ПМИД 15123711 . S2CID 23453339 .
- ^ Jump up to: а б с Захарян Э., Цао С., Рохач Т. (ноябрь 2011 г.). «Внутриклеточный АТФ поддерживает активность TRPV6 посредством липидкиназ и генерации PtdIns(4,5) P2» . Журнал ФАСЭБ . 25 (11): 3915–28. дои : 10.1096/fj.11-184630 . ПМЦ 3205842 . ПМИД 21810903 .
- ^ Jump up to: а б с Тьягараджан Б., Бенн Б.С., Кристакос С., Рохач Т. (март 2009 г.). «Фосфолипаза С-опосредованная регуляция временного рецепторного потенциала ваниллоидных 6-каналов: значение для активного транспорта Ca2+ в кишечнике» . Молекулярная фармакология . 75 (3): 608–16. дои : 10.1124/моль.108.052449 . ПМК 2684912 . ПМИД 19073818 .
- ^ Дерлер И., Хофбауэр М., Кар Х., Фрич Р., Муйк М., Кепплингер К. и др. (ноябрь 2006 г.). «Динамическая, но не конститутивная ассоциация кальмодулина с крысиными каналами TRPV6 обеспечивает точную настройку Ca2+-зависимой инактивации» . Журнал физиологии . 577 (Часть 1): 31–44. дои : 10.1113/jphysicalol.2006.118661 . ПМК 2000671 . ПМИД 16959851 .
- ^ Нимейер Б.А., Бергс К., Виссенбах У., Флокерци В., Трост К. (март 2001 г.). «Конкурентная регуляция CaT-подобного входа Ca2+ с помощью протеинкиназы C и кальмодулина» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3600–5. Бибкод : 2001PNAS...98.3600N . дои : 10.1073/pnas.051511398 . ПМК 30699 . ПМИД 11248124 .
- ^ Шин Ю.К., Шин С.Ю., Со И, Квон Д., Чон Дж.Х. (январь 2011 г.). «База данных TRIP: созданная вручную база данных межбелковых взаимодействий для каналов TRP млекопитающих» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D356-61. дои : 10.1093/nar/gkq814 . ПМК 3013757 . ПМИД 20851834 . S2CID 16278877 .
- ^ Jump up to: а б Сузуки Ю., Ландовски К.П., Хедигер М.А. (март 2008 г.). «Механизмы и регуляция эпителиальной абсорбции Ca2+ в норме и при заболеваниях». Ежегодный обзор физиологии . 70 (1): 257–71. doi : 10.1146/annurev.phyol.69.031905.161003 . ПМИД 17850211 .
- ^ Jump up to: а б с д и ж Сузуки Ю., Ковач К.С., Таканага Х., Пэн Дж.Б., Ландовски К.П., Хедигер М.А. (август 2008 г.). «Кальциевый канал TRPV6 участвует в транспорте кальция от матери к плоду у мышей» . Журнал исследований костей и минералов . 23 (8): 1249–56. дои : 10.1359/jbmr.080314 . ПМК 2680174 . ПМИД 18348695 .
- ^ Jump up to: а б Легенький В, Бек Б, Полаковска Р, Чарверон М, Бордат П, Скрыма Р, Преварская Н (август 2007 г.). «TRPV6 представляет собой канал входа Ca2+, необходимый для Ca2+-индуцированной дифференцировки кератиноцитов человека» . Журнал биологической химии . 282 (31): 22582–91. дои : 10.1074/jbc.m611398200 . ПМИД 17550901 . S2CID 22082147 .
- ^ Jump up to: а б с Ямаути Д., Накая К., Равендран Н.Н., Харбидж Д.Г., Сингх Р., Вангеманн П., Маркус Д.С. (январь 2010 г.). «Экспрессия эпителиальной системы транспорта кальция в улитке и вестибулярном лабиринте крысы» . Физиология БМК . 10 (1): 1. дои : 10.1186/1472-6793-10-1 . ПМЦ 2825184 . ПМИД 20113508 . S2CID 5773117 .
- ^ Jump up to: а б с Ямаути Д., Равендран Н.Н., Пондугула С.Р., Кампалли С.Б., Саннеман Дж.Д., Харбидж Д.Г., Маркус Д.С. (июнь 2005 г.). «Витамин D усиливает экспрессию мРНК ECaC1 в полукружном канале». Связь с биохимическими и биофизическими исследованиями . 331 (4): 1353–7. дои : 10.1016/j.bbrc.2005.04.053 . ПМИД 15883024 .
- ^ Jump up to: а б с д и Вайсгербер П., Крибс У., Цвиловский В., Олауссон Дж., Кретц О., Стергер С. и др. (май 2011 г.). «Мужская фертильность зависит от поглощения Ca²+ TRPV6 в эпидидимальном эпителии». Научная сигнализация . 4 (171): ра27. дои : 10.1126/scisignal.2001791 . ПМИД 21540454 . S2CID 206670887 .
- ^ Jump up to: а б с Вайсгербер П., Крибс У., Цвиловский В., Олауссон Дж., Кретц О., Стергер С. и др. (май 2012 г.). «Удаление гена Trpv6 приводит к серьезным нарушениям абсорбции Ca2+ в придатках яичка и мужской фертильности, подобно одиночной мутации в порах D541A» . Журнал биологической химии . 287 (22): 17930–41. дои : 10.1074/jbc.m111.328286 . ПМЦ 3365704 . ПМИД 22427671 .
- ^ Jump up to: а б с Диас де Барбоса Дж., Гуиззарди С., Толоса де Таламони Н. (июнь 2015 г.). «Молекулярные аспекты всасывания кальция в кишечнике» . Всемирный журнал гастроэнтерологии . 21 (23): 7142–54. дои : 10.3748/wjg.v21.i23.7142 . ПМЦ 4476875 . ПМИД 26109800 .
- ^ Jump up to: а б с д Бьянко С.Д., Пэн Дж.Б., Таканага Х., Сузуки Й., Кресченци А., Кос СН и др. (февраль 2007 г.). «Выраженное нарушение гомеостаза кальция у мышей с целенаправленным нарушением гена кальциевого канала Trpv6» . Журнал исследований костей и минералов . 22 (2): 274–85. дои : 10.1359/jbmr.061110 . ПМЦ 4548943 . ПМИД 17129178 .
- ^ Азарпейкан С., Диттмер К.Э., Маршалл Дж.К., Перера К.К., Джи Е.К., Акке Э., Томпсон К.Г. (15 сентября 2016 г.). «Оценка и сравнение экспрессии генов, чувствительных к витамину D, в почках овец, собак и лошадей» . ПЛОС ОДИН . 11 (9): e0162598. Бибкод : 2016PLoSO..1162598A . дои : 10.1371/journal.pone.0162598 . ПМК 5025205 . ПМИД 27632366 .
- ^ Стулк Дж. (июль 1997 г.). «Плацентарный перенос неорганических ионов и воды». Физиологические обзоры . 77 (3): 805–36. дои : 10.1152/physrev.1997.77.3.805 . ПМИД 9234966 .
- ^ Ковач CS, Кроненберг HM (декабрь 1997 г.). «Метаболизм кальция и костей у матери и плода во время беременности, послеродового периода и лактации» . Эндокринные обзоры . 18 (6): 832–72. дои : 10.1210/edrv.18.6.0319 . ПМИД 9408745 .
- ^ Шарма Д., Шастри С., Шарма П. (январь 2016 г.). «Ограничение внутриутробного развития: антенатальные и послеродовые аспекты» . Клиническая медицина. Педиатрия . 10 : 67–83. дои : 10.4137/cmped.s40070 . ПМЦ 4946587 . ПМИД 27441006 .
- ^ Jump up to: а б Бернуччи Л., Энрикес М., Диас П., Рикельме Г. (ноябрь 2006 г.). «В базальной мембране синцитиотрофобласта плаценты человека присутствуют различные типы кальциевых каналов». Плацента . 27 (11–12): 1082–95. дои : 10.1016/j.placenta.2005.12.007 . hdl : 10533/178038 . ПМИД 16564089 .
- ^ Ли Г.С., Чон Э.Б. (июль 2007 г.). «Экспрессия TRPV6 в матке во время эстрального цикла и беременности на мышиной модели». Американский журнал физиологии. Эндокринология и обмен веществ . 293 (1): E132-8. дои : 10.1152/ajpendo.00666.2006 . ПМИД 17374692 .
- ^ Моро Р., Хамель А., Дауд Г., Симоно Л., Лафон Дж. (ноябрь 2002 г.). «Экспрессия кальциевых каналов при дифференцировке культивируемых клеток трофобласта из плаценты человека» . Биология размножения . 67 (5): 1473–9. дои : 10.1095/biolreprod.102.005397 . ПМИД 12390878 . S2CID 23847059 .
- ^ Ян Х, Ким Т.Х., Ан Б.С., Чхве К.С., Ли Х.Х., Ким Дж.М., Джунг Э.Б. (март 2013 г.). «Дифференциальная экспрессия каналов транспорта кальция в первичных клетках и тканях плаценты, полученных из плаценты при преэклампсии». Молекулярная и клеточная эндокринология . 367 (1–2): 21–30. дои : 10.1016/j.mce.2012.12.012 . ПМИД 23267838 . S2CID 5276054 .
- ^ Миура С., Сато К., Като-Негиши М., Тешима Т., Такеучи С. (ноябрь 2015 г.). «Сдвиг жидкости запускает образование микроворсинок посредством механочувствительной активации TRPV6» . Природные коммуникации . 6 (1): 8871. Бибкод : 2015NatCo...6.8871M . дои : 10.1038/ncomms9871 . ПМК 4660203 . ПМИД 26563429 .
- ^ Экройд Х., Асквит К.Л., Джонс Р.К., Эйткен Р.Дж. (апрель 2004 г.). «Развитие путей передачи сигнала во время созревания придатков яичка зависит от кальция» . Биология развития . 268 (1): 53–63. дои : 10.1016/j.ydbio.2003.12.015 . ПМИД 15031104 .
- ^ Гао Д., Чжан Б.Л., Люн М.К., Ау СК, Вонг П.Ю., Шум В.В. (август 2016 г.). «Сопряжение TRPV6 и TMEM16A в главных эпителиальных клетках придатка яичка крысы» . Журнал общей физиологии . 148 (2): 161–82. дои : 10.1085/jgp.201611626 . ПМЦ 4969799 . ПМИД 27481714 .
- ^ Jump up to: а б с д Либен Л., Бенн Б.С., Аджибаде Д., Стокманс И., Моерманс К., Хедигер М.А. и др. (август 2010 г.). «Trpv6 опосредует кишечную абсорбцию кальция во время ограничения кальция и способствует гомеостазу костей» . Кость . 47 (2): 301–8. дои : 10.1016/j.bone.2010.04.595 . ПМК 2902603 . ПМИД 20399919 .
- ^ Jump up to: а б с Либен Л., Кармели Г (2012). «Участие каналов TRP в костном гомеостазе» . Границы эндокринологии . 3 : 99. дои : 10.3389/fendo.2012.00099 . ПМЦ 3422722 . ПМИД 22934090 .
- ^ Ту Ц.Л., Бикле Д.Д. (июнь 2013 г.). «Роль кальций-чувствительного рецептора в регуляции кальциевой дифференцировки и функции эпидермиса» . Лучшие практики и исследования. Клиническая эндокринология и обмен веществ . 27 (3): 415–27. дои : 10.1016/j.beem.2013.03.002 . ПМЦ 3713412 . ПМИД 23856269 .
- ^ Jump up to: а б с д Накая К., Харбидж Д.Г., Вангеманн П., Шульц Б.Д., Грин Э.Д., Уолл С.М., Маркус Д.С. (май 2007 г.). «Недостаток транспорта пендрина HCO3 повышает вестибулярный эндолимфатический [Ca2+] за счет ингибирования кислоточувствительных каналов TRPV5 и TRPV6» . Американский журнал физиологии. Почечная физиология . 292 (5): Ф1314-21. дои : 10.1152/ajprenal.00432.2006 . ПМК 2515270 . ПМИД 17200157 .
- ^ Jump up to: а б Де Клерк К., Хелд К., Ван Бри Р., Меулеман С., Пирер К., Томассетти С. и др. (июнь 2015 г.). «Функциональная экспрессия временных рецепторных потенциальных каналов в стромальных клетках эндометрия человека во время лютеиновой фазы менструального цикла» . Репродукция человека . 30 (6): 1421–36. дои : 10.1093/humrep/dev068 . ПМИД 25820697 .
- ^ Jump up to: а б с Ян Х, Чхве К.С., Хён Ш., Чон Э.Б. (апрель 2011 г.). «Коэкспрессия и эстроген-опосредованная регуляция TRPV6 и PMCA1 в эндометрии человека во время менструального цикла». Молекулярное воспроизводство и развитие . 78 (4): 274–82. дои : 10.1002/mrd.21303 . ПМИД 21400627 . S2CID 21140465 .
- ^ Jump up to: а б Де Клерк К., Ван ден Эйнде С., Хеннес А., Ван Бри Р., Воетс Т., Вриенс Дж. (март 2017 г.). «Функциональная экспрессия временных рецепторных потенциальных каналов в эндометрии мыши» . Репродукция человека . 32 (3): 615–630. дои : 10.1093/humrep/dew344 . ПМИД 28077439 . S2CID 3409475 .
- ^ Сузуки Ю, Читаят Д, Савада Х, Дирдорф М.А., Маклафлин Х.М., Бегтруп А., Миллар К., Харрингтон Дж., Чонг К., Ройфман М., Гранд К., Томинага М., Такада Ф., Шустер С., Обара М., Муто Х., Кушима Р. , Нисимура Дж. (июнь 2018 г.). «Варианты TRPV6 мешают транспорту кальция между матерью и плодом через плаценту и вызывают транзиторный неонатальный гиперпаратиреоз» . Американский журнал генетики человека . 102 (6): 1104–1114. дои : 10.1016/j.ajhg.2018.04.006 . ПМЦ 5992228 . ПМИД 29861107 .
- ^ Ямасита С., Мизумото Х., Савада Х., Сузуки Ю., Хата Д. (март 2019 г.). «Мутация гена TRPV6 у дизиготного близнеца с транзиторным неонатальным гиперпаратиреозом» . Журнал Эндокринного общества . 3 (3): 602–606. дои : 10.1210/js.2018-00374 . ПМК 6389352 . ПМИД 30820485 .
- ^ Беррен С. П., Касвелл Р., Касл Б, Уэлч С. Р., Хиллиард Т. Н., Смитсон С. Ф., Эллард С. (сентябрь 2018 г.). «Гетерозиготные варианты соединения TRPV6 приводят к нарушению плацентарного транспорта кальция и тяжелой недостаточной минерализации и дисплазии скелета плода» . Американский журнал медицинской генетики. Часть А. 176 (9): 1950–1955. дои : 10.1002/ajmg.a.40484 . ПМК 6563443 . ПМИД 30144375 .
- ^ Jump up to: а б Цзоу В.Б., Ван Ю.К., Рен XL, Ван Л., Дэн С.Дж., Мао XT и др. (август 2020 г.). «Варианты TRPV6 придают китайскому населению предрасположенность к хроническому панкреатиту» . Человеческая мутация . 41 (8): 1351–1357. дои : 10.1002/humu.24032 . ПМИД 32383311 .
- ^ Jump up to: а б с д Масамунэ А., Котани Х., Сёргель Ф.Л., Чен Дж.М., Хамада С., Сакагути Р. и др. (май 2020 г.). «Варианты, влияющие на функцию кальциевого канала TRPV6, связаны с хроническим панкреатитом с ранним началом» . Гастроэнтерология . 158 (6): 1626–1641.e8. doi : 10.1053/j.gastro.2020.01.005 . ПМИД 31930989 .
- ^ Jump up to: а б Сузуки, Йоширо Паш, Андреас Бонни, Оливье Мохаупт, Маркус Г. Хедигер, Маттиас А. Фрей, Феликс Дж. (2 августа 2017 г.). Гаплотип усиления функции эпителиального кальциевого канала TRPV6 является фактором риска образования кальциевых камней в почках . Том. 17. стр. 1613–8. дои : 10.1093/hmg/ddn048 . OCLC 1156692319 . ПМИД 18276610 .
{{cite book}}:|journal=игнорируется ( справка ) CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Уолтерс Дж.Р., Балесария С., Чавеле К.М., Тейлор В., Берри Дж.Л., Хайр У. и др. (ноябрь 2006 г.). «Экспрессия TRPV6 кальциевых каналов в двенадцатиперстной кишке человека: различные взаимоотношения с системой витамина D и старением у мужчин и женщин» . Журнал исследований костей и минералов . 21 (11): 1770–7. дои : 10.1359/jbmr.060721 . ПМИД 17002582 . S2CID 22847166 .
- ^ Jump up to: а б Шервуд Дж., Бертран Дж., Зайдеманн М., Делл'Аччо Ф., Пап Т. (апрель 2016 г.). «Активация катионного канала временного рецепторного потенциала TRPC6 необходима для фенотипической стабильности хондроцитов» . Остеоартрит и хрящ . 24 : С152–С153. дои : 10.1016/j.joca.2016.01.298 . ISSN 1063-4584 .
- ^ Jump up to: а б с д Дрор А.А., Браунштейн З., Авраам К.Б. (2011). «Интеграция генетики человека и мыши раскрывает функцию пендрина при слухе и глухоте» . Клеточная физиология и биохимия . 28 (3): 535–44. дои : 10.1159/000335163 . ПМК 3709173 . ПМИД 22116368 .
- ^ Сунь Ф., Сяо Л., Чан XX, Сюн Ю, Ли Q, Юэ XJ и др. (октябрь 2016 г.). «TRPV6 является прогностическим маркером плоскоклеточного рака шейки матки на ранней стадии». Биология опухолей . 37 (12): 15743–15751. дои : 10.1007/s13277-016-5368-4 . ПМИД 27747588 . S2CID 6445506 .
- ^ Jump up to: а б Фань Х, Шен YX, Юань ЮФ (30 марта 2014 г.). «Экспрессия и прогностическая роль TRPV5 и TRPV6 при немелкоклеточном раке легкого после радикальной резекции» . Азиатско-Тихоокеанский журнал профилактики рака . 15 (6): 2559–63. дои : 10.7314/apjcp.2014.15.6.2559 . ПМИД 24761864 .
- ^ Ву Ю, Миямото Т, Ли К, Накагоми Х, Савада Н, Кира С и др. (декабрь 2011 г.). «Снижение экспрессии эпителиальных Ca2+-каналов TRPV5 и TRPV6 при почечно-клеточной карциноме человека, связанной с рецептором витамина D». Журнал урологии . 186 (6): 2419–25. дои : 10.1016/j.juro.2011.07.086 . ПМИД 22019165 .
- ^ Jump up to: а б с д и Пэн Дж.Б., Чжуан Л., Бергер У.В., Адам Р.М., Уильямс Б.Дж., Браун Э.М. и др. (апрель 2001 г.). «Экспрессия CaT1 коррелирует со степенью опухоли при раке простаты». Связь с биохимическими и биофизическими исследованиями . 282 (3): 729–34. дои : 10.1006/bbrc.2001.4638 . ПМИД 11401523 .
- ^ Jump up to: а б Фиксемер Т., Виссенбах У., Флокерци В., Бонкхофф Х. (октябрь 2003 г.). «Экспрессия Ca2+-селективного катионного канала TRPV6 при раке предстательной железы человека: новый прогностический маркер прогрессирования опухоли» . Онкоген . 22 (49): 7858–61. дои : 10.1038/sj.onc.1206895 . ПМИД 14586412 . S2CID 23626142 .
- ^ Деннин-Дютиль И., Готье М., Фаузи М., Гильбер А., Бреве М., Водри Д. и др. (2011). «Высокая экспрессия каналов транзиторного рецепторного потенциала в эпителиальных клетках и тканях рака молочной железы человека: корреляция с патологическими параметрами» . Клеточная физиология и биохимия . 28 (5): 813–22. дои : 10.1159/000335795 . ПМИД 22178934 . S2CID 45740662 .
- ^ Jump up to: а б Боланц К.А., Хедигер М.А., Ландовски К.П. (февраль 2008 г.). «Роль TRPV6 в канцерогенезе молочной железы» . Молекулярная терапия рака . 7 (2): 271–9. дои : 10.1158/1535-7163.mct-07-0478 . ПМИД 18245667 . S2CID 17158946 .
- ^ Питерс А.А., Симпсон П.Т., Бассетт Дж.Дж., Ли Дж.М., Да Силва Л., Рид Л.Е. и др. (октябрь 2012 г.). «Кальциевый канал TRPV6 как потенциальная терапевтическая мишень при раке молочной железы, отрицательном по рецепторам эстрогена» . Молекулярная терапия рака . 11 (10): 2158–68. дои : 10.1158/1535-7163.mct-11-0965 . ПМИД 22807578 . S2CID 207614323 .
- ^ Jump up to: а б с Легенький В, Флуракис М, Скрыма Р, Преварская Н (ноябрь 2007 г.). «Канал TRPV6 контролирует пролиферацию клеток рака простаты посредством Ca(2+)/NFAT-зависимых путей» . Онкоген . 26 (52): 7380–5. дои : 10.1038/sj.onc.1210545 . ПМИД 17533368 . S2CID 482760 .
- ^ Легенький В, Рафаэль М, Улиди А, Флуракис М, Халимончик С, Кондрацкий А и др. (февраль 2011 г.). «TRPV6 определяет влияние витамина D3 на рост клеток рака простаты» . ПЛОС ОДИН . 6 (2): e16856. Бибкод : 2011PLoSO...616856L . дои : 10.1371/journal.pone.0016856 . ПМК 3037935 . ПМИД 21347289 .
- ^ Боланц К.А., Ковач Г.Г., Ландовски К.П., Хедигер М.А. (декабрь 2009 г.). «Тамоксифен ингибирует активность TRPV6 через независимые от рецептора эстрогена пути в клетках рака молочной железы MCF-7, экспрессирующих TRPV6» . Молекулярные исследования рака . 7 (12): 2000–10. дои : 10.1158/1541-7786.mcr-09-0188 . ПМИД 19996302 . S2CID 26337876 .
- ^ Пелег С., Селлин Дж. Х., Ван Ю., Фриман М. Р., Умар С. (сентябрь 2010 г.). «Подавление аберрантного переходного рецепторного потенциала катионного канала, подсемейство V, экспрессия члена 6 в гиперпролиферативных криптах толстой кишки с помощью пищевого кальция» . Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 299 (3): G593-601. дои : 10.1152/ajpgi.00193.2010 . ПМЦ 2950683 . ПМИД 20508153 .
- ^ Арбабиан А., Ифтинка М., Алтье С., Сингх П.П., Исамберт Х., Коской С. (декабрь 2020 г.). «Мутации в кальмодулин-связывающих доменах каналов TRPV4/6 придают инвазивные свойства клеткам аденокарциномы толстой кишки» . Каналы . 14 (1): 101–109. дои : 10.1080/19336950.2020.1740506 . ПМЦ 7153789 . ПМИД 32186440 .
- ^ Исидзава М., Акаги Д., Ямамото Дж., Макишима М. (сентябрь 2017 г.). «3 усиливает транскрипцию TRPV6 за счет активации p38 MAPK и экспрессии GADD45». Журнал биохимии стероидов и молекулярной биологии . 172 : 55–61. дои : 10.1016/j.jsbmb.2017.05.013 . ПМИД 28578001 . S2CID 206502344 .
- ^ Дай В., Бай Ю, Хебда Л., Чжун Икс, Лю Дж., Као Дж., Дуань С. (апрель 2014 г.). «Вызванная дефицитом кальция и регулируемая TRP-каналом передача сигналов IGF1R-PI3K-Akt регулирует аномальную пролиферацию эпителиальных клеток» . Смерть клеток и дифференцировка . 21 (4): 568–81. дои : 10.1038/cdd.2013.177 . ПМЦ 3950320 . ПМИД 24336047 .
- ^ Скшипский М., Колодзейский П.А., Мерглер С., Хаяви Н., Новак К.В., Стровски М.З. (август 2016 г.). «TRPV6 модулирует пролиферацию нейроэндокринных опухолевых клеток BON-1 поджелудочной железы человека» . Отчеты по биологическим наукам . 36 (4). дои : 10.1042/bsr20160106 . ПМЦ 4995500 . ПМИД 27450545 .
- ^ Сун Х, Донг М, Чжоу Дж, Шэн В, Ли Х, Гао В (март 2018 г.). «Экспрессия и прогностическое значение TRPV6 в развитии и прогрессировании рака поджелудочной железы» . Отчеты онкологии . 39 (3): 1432–1440. дои : 10.3892/или.2018.6216 . ПМИД 29344675 .
- ^ Чоу Дж., Норг М., Чжан Дж., Чай Дж. (апрель 2007 г.). «TRPV6 опосредует индуцированный капсаицином апоптоз в клетках рака желудка — механизмы, лежащие в основе возможного нового «горячего» лечения рака» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1773 (4): 565–76. дои : 10.1016/j.bbamcr.2007.01.001 . ПМИД 17292493 .
- ^ Чжан С.С., Се X, Вэнь Дж., Луо К.Дж., Лю К.В., Ян Х. и др. (январь 2016 г.). «TRPV6 играет новую роль в прогнозировании выживаемости пациентов с плоскоклеточным раком пищевода» . Диагностическая патология . 11 (1): 14. дои : 10.1186/s13000-016-0457-7 . ПМК 4730645 . ПМИД 26818094 .
- ^ Jump up to: а б с Сонг Ю, Пэн Х, Порта А, Таканага Х, Пэн Дж.Б., Хедигер М.А. и др. (сентябрь 2003 г.). «Переносчик кальция 1 и рибонуклеиновая кислота эпителиального кальциевого канала по-разному регулируются 1,25-дигидроксивитамином D3 в кишечнике и почках мышей» . Эндокринология . 144 (9): 3885–94. дои : 10.1210/en.2003-0314 . ПМИД 12933662 .
- ^ Jump up to: а б с д и ж Браун AJ, Критс I, Армбрехт HJ (май 2005 г.). «Влияние возраста, витамина D и кальция на регуляцию кальциевых каналов эпителия кишечника крыс». Архив биохимии и биофизики . 437 (1): 51–8. дои : 10.1016/j.abb.2005.02.007 . ПМИД 15820216 .
- ^ Jump up to: а б Мейер М.Б., Ватануки М., Ким С., Шевде Н.К., Пайк Дж.В. (июнь 2006 г.). «Дистальный промотор ваниллоидного типа 6 с потенциалом временного рецептора человека содержит множество сайтов связывания рецептора витамина D, которые опосредуют активацию 1,25-дигидроксивитамином D3 в клетках кишечника» . Молекулярная эндокринология . 20 (6): 1447–61. дои : 10.1210/me.2006-0031 . ПМИД 16574738 .
- ^ Jump up to: а б Сонг Ю, Като С, Fleet JC (февраль 2003 г.). «Мыши с нокаутом рецептора витамина D (VDR) обнаруживают VDR-независимую регуляцию кишечной абсорбции кальция и мРНК ECaC2 и кальбиндина D9k» . Журнал питания . 133 (2): 374–80. дои : 10.1093/jn/133.2.374 . ПМИД 12566470 .
- ^ Кристакос С., Дхаван П., Верстюф А., Верлинден Л., Кармелиет Дж. (январь 2016 г.). «Витамин D: метаболизм, молекулярный механизм действия и плейотропные эффекты» . Физиологические обзоры . 96 (1): 365–408. doi : 10.1152/physrev.00014.2015 . ПМЦ 4839493 . ПМИД 26681795 .
- ^ Буйон Р., Ван Кромфо С., Кармели Дж. (февраль 2003 г.). «Кишечная абсорбция кальция: молекулярные механизмы, опосредованные витамином D». Журнал клеточной биохимии . 88 (2): 332–9. дои : 10.1002/jcb.10360 . ПМИД 12520535 . S2CID 9853381 .
- ^ Реплогл РА, Ли Q, Ван Л, Чжан М, Fleet JC (март 2014 г.). «Взаимодействие генов и диеты влияет на усвоение кальция и плотность костей у мышей» . Журнал исследований костей и минералов . 29 (3): 657–65. дои : 10.1002/jbmr.2065 . ПМЦ 10591522 . ПМИД 23955923 . S2CID 1546230 .
- ^ Jump up to: а б Ван Кромфаут С.Дж., Рамменс К., Стокманс И., Ван Херк Э., Дейкс Ф.А., Эдервин АГ и др. (октябрь 2003 г.). «Гены кишечного переносчика кальция активируются эстрогенами и репродуктивным циклом посредством механизмов, независимых от рецептора витамина D» . Журнал исследований костей и минералов . 18 (10): 1725–36. дои : 10.1359/jbmr.2003.18.10.1725 . ПМИД 14584880 . S2CID 25346500 .
- ^ Чароенпхандху Н., Наккрасае Л.И., Крайдит К., Тирапорнпунтакит Дж., Тонгчоте К., Тонгон Н., Кришнамра Н. (сентябрь 2009 г.). «Двухступенчатая стимуляция кишечной абсорбции Ca(2+) во время лактации путем длительного воздействия пролактина и всплеска пролактина, вызванного грудным вскармливанием». Американский журнал физиологии. Эндокринология и обмен веществ . 297 (3): E609-19. дои : 10.1152/ajpendo.00347.2009 . ПМИД 19567804 .
- ^ Jump up to: а б Беггс М.Р., Ли Дж.Дж., Буш К., Раза А., Димке Х., Вайсгербер П. и др. (2019). «v1.3 Опосредует абсорбцию кальция в дистальном отделе тонкой кишки перед отлучением от груди» . Клеточная и молекулярная гастроэнтерология и гепатология . 8 (4): 625–642. дои : 10.1016/j.jcmgh.2019.07.005 . ПМК 6889763 . ПМИД 31398491 .
- ^ ван Абель М., Хайберс С., Хендероп Дж.Г., ван дер Кемп А.В., ван Леувен Дж.П., Бинделс Р.Дж. (декабрь 2006 г.). «Возрастные изменения в гомеостазе Ca2+: роль TRPV5 и TRPV6». Американский журнал физиологии. Почечная физиология . 291 (6): F1177-83. дои : 10.1152/ajprenal.00038.2006 . ПМИД 16705151 .
- ^ Не Икс, Цзинь Х, Вэнь Г, Сюй Дж, Ан Дж, Лю Икс и др. (январь 2020 г.). «Эстроген регулирует абсорбцию кальция в двенадцатиперстной кишке посредством дифференциальной роли рецептора эстрогена в белках-переносчиках кальция». Пищеварительные заболевания и науки . 65 (12): 3502–3513. дои : 10.1007/s10620-020-06076-x . ПМИД 31974908 . S2CID 210862678 .
- ^ Ли Б.М., Ли Г.С., Юнг Э.М., Чхве К.К., Джунг Э.Б. (май 2009 г.). «Маточная и плацентарная экспрессия гена TRPV6 регулируется посредством путей, опосредованных рецептором прогестерона или рецептором эстрогена, во время беременности у грызунов» . Репродуктивная биология и эндокринология . 7 (1): 49. дои : 10.1186/1477-7827-7-49 . ПМК 2694200 . ПМИД 19457270 .
- ^ Пак С.Ю., Ю Ю.М., Чон Э.М., Чон Э.Б. (апрель 2020 г.). «Влияние стероидного гормона на экспрессию белков, перерабатывающих кальций, в незрелом мозге самок крыс». Журнал химической нейроанатомии . 105 : 101767. doi : 10.1016/j.jchemneu.2020.101767 . ПМИД 32061997 . S2CID 211105316 .
- ^ Jump up to: а б Ким М.Х., Ли Г.С., Юнг Э.М., Чхве К.К., Джунг Э.Б. (июль 2009 г.). «Отрицательное влияние дексаметазона на экспрессию генов, перерабатывающих кальций, связано с расстройством абсорбции кальция, вызванным глюкокортикоидами». Науки о жизни . 85 (3–4): 146–52. дои : 10.1016/j.lfs.2009.05.013 . ПМИД 19490920 .
- ^ Jump up to: а б Ку Т.Х., Ян Х., Юнг Э.Б. (1 июля 2011 г.). «Экспрессия генов транспорта кальция в плаценте мышей с нокаутом кальбиндина-D9k и -D28k». Биология размножения . 85 (Suppl_1): 449. doi : 10.1093/biolreprod/85.s1.449 . ISSN 0006-3363 .
- ^ Jump up to: а б Сопьяни М., Кунерт А., Чарковский К., Клаус Ф., Лауфер Дж., Фёллер М., Ланг Ф. (февраль 2010 г.). «Регуляция Ca (2+)-канала TRPV6 киназами SGK1, PKB / Akt и PIKfyve». Журнал мембранной биологии . 233 (1–3): 35–41. дои : 10.1007/s00232-009-9222-0 . ПМИД 20041238 . S2CID 25349984 .
Дальнейшее чтение
[ редактировать ]- Хайнер И., Эйсфельд Дж., Люкхофф А. (2004). «Роль и регуляция каналов TRP в нейтрофильных гранулоцитах». Клеточный кальций . 33 (5–6): 533–40. дои : 10.1016/S0143-4160(03)00058-7 . ПМИД 12765698 .
- Клэпхэм Д.Э., Джулиус Д., Монтелл С., Шульц Г. (декабрь 2005 г.). «Международный союз фармакологии. XLIX. Номенклатура и структурно-функциональные связи временных рецепторных потенциальных каналов». Фармакологические обзоры . 57 (4): 427–50. дои : 10.1124/пр.57.4.6 . ПМИД 16382100 . S2CID 17936350 .
- Виссенбах Ю, Нимейер Б.А. (2007). «ТРПВ6». Каналы переходного рецепторного потенциала (TRP) . Справочник по экспериментальной фармакологии. Том. 179. стр. 221–34. дои : 10.1007/978-3-540-34891-7_13 . ISBN 978-3-540-34889-4 . ПМИД 17217060 .
- Шобер Дж. П., Хендероп Дж. Г., Биндельс Р. Дж. (февраль 2007 г.). «Совместное действие связанных белков в регуляции TRPV5 и TRPV6». Труды Биохимического общества . 35 (Часть 1): 115–9. дои : 10.1042/BST0350115 . ПМИД 17233615 .
Внешние ссылки
[ редактировать ]- TRPV+катион+каналы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- TRPV6 + белок, + человек Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)