Потенциал-управляемый натриевый канал
Потенциал-управляемые натриевые каналы ( VGSC ), также известные как потенциал-зависимые натриевые каналы ( VDSC ), представляют собой группу потенциал-управляемых ионных каналов, обнаруженных в мембранах возбудимых клеток ( например , мышечных , глиальных клеток , нейронов и т. д.). с проницаемостью для ионов натрия Na + . Они являются основными каналами, участвующими в потенциале действия возбудимых клеток.
Структура
[ редактировать ]
Натриевые каналы состоят из крупных альфа-субъединиц , которые связаны с вспомогательными белками, такими как бета-субъединицы . Альфа-субъединица образует ядро канала и функционирует сама по себе. Когда белок альфа-субъединицы экспрессируется клеткой, он способен образовывать поры в клеточной мембране, которые проводят Na. + потенциалзависимым образом, даже если бета-субъединицы или другие известные модулирующие белки не экспрессируются. Когда вспомогательные белки собираются с α-субъединицами, образующийся комплекс может демонстрировать измененную зависимость от напряжения и клеточную локализацию.
Альфа-субъединица состоит из четырех повторяющихся доменов, обозначенных от I до IV, каждый из которых содержит шесть трансмембранных сегментов, обозначенных от S1 до S6. Высококонсервативный сегмент S4 действует как датчик напряжения канала. Чувствительность этого канала к напряжению обусловлена положительными аминокислотами, расположенными в каждой третьей позиции. [2] При стимуляции изменением трансмембранного напряжения этот сегмент перемещается к внеклеточной стороне клеточной мембраны, позволяя каналу стать проницаемым для ионов. Ионы проводятся через центральную полость поры, которая состоит из двух основных областей. Более внешняя (т.е. более внеклеточная) часть поры образована «P-петлями» (область между S5 и S6) четырех доменов. Эта область является самой узкой частью поры и отвечает за ее ионную селективность. Внутренняя часть (т.е. более цитоплазматическая) поры представляет собой ворота поры и образована объединенными сегментами S5 и S6 четырех доменов. Поровая область также имеет боковые туннели или фенестрации, которые проходят перпендикулярно оси поры. Предполагается, что эти фенестрации, соединяющие центральную полость с мембраной, важны для доступности лекарств. [3] [4] [5]
В натриевых каналах млекопитающих область, связывающая домены III и IV, также важна для функции канала. Этот линкер DIII-IV отвечает за закрытие ворот поры после открытия канала, инактивируя их. [6]
ворота
[ редактировать ]Напряжение-управляемый Na + каналы имеют три основных конформационных состояния: закрытое, открытое и инактивированное. Переходы вперед/назад между этими состояниями соответственно называются активацией/деактивацией (между открытым и закрытым соответственно), инактивацией/реактивацией (между неактивным и открытым соответственно) и восстановлением из инактивации/инактивацией закрытого состояния (между неактивным и закрытым состоянием). , соответственно). Закрытые и инактивированные состояния непроницаемы для ионов.
До возникновения потенциала действия аксональная мембрана находится в нормальном потенциале покоя , около -70 мВ в большинстве нейронов человека, и Na + каналы находятся в деактивированном состоянии и заблокированы на внеклеточной стороне воротами активации . В ответ на повышение мембранного потенциала примерно до -55 мВ (в данном случае вызванное потенциалом действия) активационные ворота открываются, позволяя положительно заряженному Na + ионы поступают в нейрон через каналы, вызывая повышение напряжения на мембране нейрона до +30 мВ в нейронах человека. Поскольку напряжение на мембране изначально отрицательное, а по мере того, как его напряжение увеличивается до нуля и выше (от -70 мВ в состоянии покоя до максимума +30 мВ), говорят, что оно деполяризуется. Это увеличение напряжения представляет собой фазу нарастания потенциала действия.
| Потенциал действия | Мембранный потенциал | Целевой потенциал | Целевое состояние ворот | Целевое состояние нейрона |
|---|---|---|---|---|
| Отдых | −70 мВ | −55 мВ | Деактивировано → Активировано | поляризованный |
| Восходящий | −55 мВ | 0 мВ | Активировано | Поляризованный → Деполяризованный |
| Восходящий | 0 мВ | +30 мВ | Активировано → Деактивировано | деполяризованный |
| Падение | +30 мВ | 0 мВ | Деактивирован | Деполяризованный → Реполяризованный |
| Падение | 0 мВ | −70 мВ | Деактивирован | реполяризованный |
| Перекус | −70 мВ | −75 мВ | Деактивировано → Деактивировано | Реполяризованный → Гиперполяризованный |
| Отскок | −75 мВ | −70 мВ | Деактивирован | Гиперполяризованный → Поляризованный |
На пике потенциала действия, когда достаточно Na + проник в нейрон и потенциал мембраны стал достаточно высоким, Na + каналы инактивируются, закрывая свои ворота инактивации . Ворота инактивации можно рассматривать как «пробку», привязанную к доменам III и IV внутриклеточной альфа-субъединицы канала. Закрытие ворот инактивации вызывает Na + поток через канал прекращается, что, в свою очередь, приводит к прекращению роста мембранного потенциала. Закрытие ворот инактивации создает рефрактерный период внутри каждого отдельного Na. + канал. Этот рефрактерный период исключает возможность движения потенциала действия в противоположном направлении обратно к соме. Когда ворота инактивации закрыты, канал считается инактивированным. С На + Канал больше не вносит вклад в мембранный потенциал, потенциал снижается обратно до потенциала покоя по мере того, как нейрон реполяризуется и впоследствии гиперполяризуется, и это составляет фазу спада потенциала действия. Таким образом, рефрактерный период каждого канала жизненно важен для однонаправленного распространения потенциала действия вниз по аксону для правильной связи между нейронами.
Когда напряжение мембраны становится достаточно низким, ворота инактивации снова открываются, а ворота активации закрываются в процессе, называемом деинактивацией . Когда ворота активации закрыты и ворота инактивации открыты, Na + канал снова находится в деактивированном состоянии и готов участвовать в другом потенциале действия.
Когда какой-либо ионный канал не инактивируется, о нем говорят, что он постоянно (или тонически) активен. Некоторые виды ионных каналов по своей природе постоянно активны. Однако генетические мутации, вызывающие постоянную активность в других каналах, могут вызвать заболевание, вызывая чрезмерную активность определенных типов нейронов. Мутации, мешающие Na + инактивация каналов может способствовать сердечно-сосудистым заболеваниям или эпилептическим припадкам из-за оконных токов , что может привести к перевозбуждению мышечных и/или нервных клеток.
Моделирование поведения ворот
[ редактировать ]Временное поведение Na + каналы можно моделировать с помощью марковской схемы или формализма типа Ходжкина – Хаксли . В первой схеме каждый канал занимает отдельное состояние с дифференциальными уравнениями, описывающими переходы между состояниями; во втором случае каналы рассматриваются как совокупность , на которую влияют три независимые переменные стробирования. Каждая из этих переменных может принимать значения от 1 (полная проницаемость для ионов) до 0 (полная непроницаемость), произведение этих переменных дает процент проводящих каналов. Можно показать, что модель Ходжкина – Хаксли эквивалентна марковской модели. [ нужны дальнейшие объяснения ]
Непроницаемость для других ионов
[ редактировать ]Поры натриевых каналов содержат селективный фильтр, состоящий из отрицательно заряженных аминокислотных остатков, которые притягивают положительные Na. + ион и не допускайте отрицательно заряженных ионов, таких как хлорид . Катионы текут в более суженную часть поры шириной 0,3 на 0,5 нм , что достаточно велико, чтобы позволить одному Na + воды, ион с молекулой связанной с прохождением. Чем больше К + ион не может пройти через эту область. Ионы разных размеров также не могут хорошо взаимодействовать с отрицательно заряженными остатками глутаминовой кислоты , выстилающими поры. [ нужна ссылка ]
Разнообразие
[ редактировать ]Потенциал-управляемые натриевые каналы обычно состоят из альфа-субъединицы, которая образует пору ионной проводимости, и одной-двух бета-субъединиц, которые выполняют несколько функций, включая модуляцию шлюзования каналов. [7] Экспрессии одной альфа-субъединицы достаточно для создания функционального канала.
Альфа-субъединицы
[ редактировать ]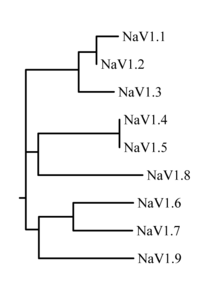
Семейство натриевых каналов включает 9 известных членов с идентичностью аминокислот >50% в трансмембранных сегментах и внеклеточных петлевых областях. В настоящее время используется стандартизированная номенклатура натриевых каналов, которая поддерживается IUPHAR . [8]
Белки этих каналов получили названия от Na v 1.1 до Na v 1.9. Названия генов обозначаются как от SCN1A до SCN5A, затем от SCN8A до SCN11A. [8] «Десятый член», Na x , не действует потенциалзависимым образом. Он имеет схожую общую структуру. О его реальной функции известно немного, кроме того, что он также связан с бета-субъединицами. [9]
Вероятная эволюционная связь между этими каналами, основанная на сходстве их аминокислотных последовательностей, показана на рисунке 1. Отдельные натриевые каналы отличаются не только различиями в их последовательностях, но также по их кинетике и профилям экспрессии. Некоторые из этих данных обобщены в таблице 1 ниже.
| Название белка | Ген | Профиль выражения | Сопутствующие каналопатии человека |
|---|---|---|---|
| В версии 1.1 | SCN1A | Центральные нейроны , [периферические нейроны] и сердечные миоциты | фебрильная эпилепсия , GEFS+ , синдром Драве (также известный как тяжелая миклоническая эпилепсия младенчества или SMEI), пограничный SMEI (SMEB), синдром Веста (также известный как инфантильные спазмы ), синдром Дуза (также известный как миоклоническая астатическая эпилепсия ), трудноизлечимая детская эпилепсия с генерализованными тонико-клоническими припадками (ICEGTC), синдромом Панайотопулоса, семейной гемиплегической мигренью (СГМ), семейным аутизмом, энцефалитом Расмуссена и синдромом Леннокса-Гасто [10] |
| На версии 1.2 | SCN2A | Центральные нейроны, периферические нейроны | наследственные фебрильные судороги , эпилепсия и расстройства аутистического спектра. |
| На версии 1.3 | SCN3A | Центральные нейроны, периферические нейроны и кардиомиоциты | эпилепсия, боль, пороки развития головного мозга [11] [12] |
| На версии 1.4 | SCN4A | Скелетные мышцы | гиперкалиемический периодический паралич , врожденная парамиотония и миотония, отягощенная калием. |
| На версии 1.5 | SCN5A | Сердечные миоциты, неиннервированные скелетные мышцы, центральные нейроны, гладкомышечные клетки желудочно-кишечного тракта и интерстициальные клетки Кахаля. | Со стороны сердца: синдром удлиненного интервала QT 3 типа, синдром Бругада , прогрессирующее заболевание сердечной проводимости , семейная фибрилляция предсердий и идиопатическая фибрилляция желудочков ; [13] Желудочно-кишечный тракт: синдром раздраженного кишечника ; [14] |
| На версии 1.6 | SCN8A | Центральные нейроны, ганглии дорсальных корешков , периферические нейроны , сердце, глиальные клетки. | Эпилепсия , [15] атаксия , дистония , тремор [16] |
| На версии 1.7 | SCN9A | Ганглии дорсальных корешков , симпатические нейроны, шванновские клетки и нейроэндокринные клетки. | эритромелалгия , ПЭПД , нечувствительность к боли, связанная с каналопатией [11] и недавно обнаружил инвалидизирующую форму фибромиалгии (полиморфизм rs6754031). [17] |
| На версии 1.8 | SCN10A | Дорсальные корешковые ганглии | боль, [11] нервно-психические расстройства |
| В версии 1.9 | SCN11A | Дорсальные корешковые ганглии | боль [11] |
| На х | SCN7A | сердце, матка, скелетные мышцы, астроциты, ганглиозные клетки дорсальных корешков | никто не известен |
Бета-субъединицы
[ редактировать ]Бета-субъединицы натриевых каналов представляют собой трансмембранные гликопротеины типа 1 с внеклеточным N-концом и цитоплазматическим С-концом. Будучи членами суперсемейства Ig, бета-субъединицы содержат прототипную петлю V-set Ig во внеклеточном домене. Они не имеют никакой гомологии со своими аналогами кальциевых и калиевых каналов. [18] Вместо этого они гомологичны молекулам адгезии нервных клеток (CAM) и большому семейству CAM L1. Существует четыре различных бета-версии, названных в порядке открытия: SCN1B, SCN2B, SCN3B, SCN4B (таблица 2). Бета 1 и бета 3 взаимодействуют с альфа-субъединицей нековалентно, тогда как бета 2 и бета 4 связываются с альфа через дисульфидную связь. [19] Натриевые каналы с большей вероятностью остаются открытыми на подпороговом мембранном потенциале при взаимодействии с бета-токсинами, что, в свою очередь, вызывает немедленное ощущение боли. [20]
Роль бета-субъединиц как молекул клеточной адгезии
[ редактировать ]Помимо регуляции открытия каналов, бета-субъединицы натриевых каналов также модулируют экспрессию каналов и образуют связи с внутриклеточным цитоскелетом через анкирин и спектрин . [7] [21] [22] Потенциал-управляемые натриевые каналы также собираются с множеством других белков, таких как белки FHF (гомологичный фактор фактора роста фибробластов), кальмодулин, цитоскелет или регуляторные киназы. [23] [7] [24] [25] [26] которые образуют комплекс с натриевыми каналами, влияя на его экспрессию и/или функцию. Несколько бета-субъединиц взаимодействуют с одной или несколькими молекулами внеклеточного матрикса (ECM). Контактин, также известный как F3 или F11, связывается с бета-1, как показано посредством коиммунопреципитации. [27] Фибронектин- подобные (FN-подобные) повторы тенасцина -C и тенасцина -R связываются с бета-2, в отличие от повторов, подобных эпидермальному фактору роста (EGF-подобных), которые отталкивают бета2. [28] Дезинтегрин и металлопротеиназа (ADAM) 10 отбрасывает эктодомен бета 2, возможно, вызывая рост нейритов. [29] Бета-3 и бета-1 связываются с нейрофасцином в узлах Ранвье в развивающихся нейронах. [30]
| Название белка | Генная ссылка | Собирается с | Профиль выражения | Сопутствующие каналопатии человека |
|---|---|---|---|---|
| На v β1 | SCN1B | от Na v 1,1 до Na v 1,7 | Центральные нейроны, периферические нейроны, скелетные мышцы, сердце, глия. | эпилепсия (GEFS+), синдром Бругада [31] |
| Na v β2 | СКН2Б | от Na v 1,1, от Na v 1,2, от Na v 1,5 до Na v 1,7 | Центральные нейроны, периферические нейроны, сердце, глия | синдром Бругада [31] |
| Na v β3 | SCN3B | от Na v 1,1 до Na v 1,3, Na v 1,5 | центральные нейроны, надпочечники, почки, периферические нейроны | синдром Бругада [31] |
| Na v β4 | SCN4B | Na v 1,1, Na v 1,2, Na v 1,5 | сердце, скелетные мышцы, центральные и периферические нейроны | никто не известен |
Эволюция
[ редактировать ]Потенциал-управляемый натриевый канал присутствует у представителей хоанофлагеллят , которые считаются ближайшими живыми одноклеточными родственниками животных. [32] [33] Это говорит о том, что предковая форма животного канала была среди многих белков, которые играют центральную роль в жизни животных, но, как полагают, развились до многоклеточности. [34] Четырехдоменный потенциалзависимый натриевый канал животных, вероятно, развился из односубъединичного ионного канала, который, вероятно, был проницаем для ионов калия, посредством последовательности двух событий дупликации. [35] Эта модель подтверждается тем фактом, что субъединицы I и III (а также II и IV) группируются по сходству, что позволяет предположить, что двухканальный промежуточный продукт, образующийся в результате первой дупликации, существовал достаточно долго, чтобы между двумя его субъединицами произошла дивергенция. После второго дублирования на канале осталось два набора одинаковых доменов. [35] Считается, что образовавшийся четырехдоменный канал был проницаем в первую очередь для кальция и несколько раз независимо достигал селективности по натрию. [36] [37] После дивергенции от беспозвоночных линия позвоночных претерпела две полногеномные дупликации (WGD), в результате чего у предковых позвоночных появился набор из четырех прологов генов натриевых каналов, все из которых были сохранены. [38] [39] После разделения четвероногих и костистых костей костистые кости, вероятно, подверглись третьему WGD, что привело к появлению восьми прологов натриевых каналов, экспрессируемых у многих современных рыб. [38] Считается, что современный набор натриевых генов млекопитающих, состоящий из десяти паралогов, возник в результате серии параллельных и вложенных дупликаций, включающих два из четырех паралогов, присутствующих у предков всех четвероногих. [39]
У электрических рыб
[ редактировать ]У некоторых рыб есть электрический орган, функционирующий с помощью натриевых каналов, управляемых напряжением, что позволяет им общаться, обездвиживать добычу или отпугивать хищников, в зависимости от рыбы. Электрические органы, используемые для общения, развивались независимо по крайней мере дважды: у Gymnotiformes Южной Америки и Mormyriformes Африки. [40] У неэлектрических рыб каналы Na v 1.4a и Na v 1.4b присутствуют в мышцах, тогда как у Gymnotiformes и Mormyriformes Na v 1.4a исчез в мышцах и экспрессируется в электрическом органе. [40]
Источники
[ редактировать ]![]() В эту статью включен текст Марьям Иман, Атефе Саадабади и Асгара Давуда, доступный по лицензии CC BY 4.0 .
В эту статью включен текст Марьям Иман, Атефе Саадабади и Асгара Давуда, доступный по лицензии CC BY 4.0 .
Ссылки
[ редактировать ]- ^ Ю Ф.Х., Каттералл, Вашингтон (2003). «Обзор семейства потенциал-управляемых натриевых каналов» . Геномная биология . 4 (3): 207. doi : 10.1186/gb-2003-4-3-207 . ПМК 153452 . ПМИД 12620097 .
- ^ Николлс, Мартин, Фукс, Браун, Даймонд, Вайсблат. (2012) «От нейрона к мозгу», 5-е изд. стр. 86
- ^ Хилле, Б. (1977) Местные анестетики: гидрофильные и гидрофобные пути реакции лекарственного средства на рецептор. Журнал общей физиологии, 69, 497–515. http://dx.doi.org/10.1085/jgp.69.4.497
- ^ Гамаль Эль-Дин, Тамер М. и др. «Фенестрации контролируют блокировку потенциалзависимого натриевого канала в состоянии покоя». Труды Национальной академии наук 115.51 (2018): 13111-13116. https://doi.org/10.1073/pnas.1814928115
- ^ Тао, Элейн и Бен Корри. «Характеристика размера фенестрации подтипов натриевых каналов и их доступности для ингибиторов». Биофизический журнал 121.2 (2022): 193-206. https://doi.org/10.1016/j.bpj.2021.12.025
- ^ Ян, Чжэнь и др. «Структура комплекса Nav1.4-β1 электрического угря». Ячейка 170.3 (2017): 470–482. https://doi.org/10.1016/j.cell.2017.06.039
- ^ Jump up to: а б с Исом Л.Л. (февраль 2001 г.). «Бета-субъединицы натриевых каналов: что угодно, кроме вспомогательных». Нейробиолог . 7 (1): 42–54. дои : 10.1177/107385840100700108 . ПМИД 11486343 . S2CID 86422657 .
- ^ Jump up to: а б Каттералл В.А., Голдин А.Л., Ваксман С.Г. (декабрь 2005 г.). «Международный союз фармакологии. XLVII. Номенклатура и структурно-функциональные связи потенциалзависимых натриевых каналов» . Фармакологические обзоры . 57 (4): 397–409. дои : 10.1124/пр.57.4.4 . ПМИД 16382098 . S2CID 7332624 .
- ^ Ноланд, Кэмерон Л.; Чуа, Хан Чоу; Кшонсак, Марк; Хойссер, Стефани Андреа; Браун, Нина; Чанг, Тимоти; Тэм, Кристина; Тан, Цзя; Артур, Кристофер П.; Чиферри, Клаудио; Плесс, Стефан Александр; Паянде, Цзян (17 марта 2022 г.). «Структурно-ориентированное разблокирование NaX обнаруживает неселективный, чувствительный к тетродотоксину катионный канал» . Природные коммуникации . 13 (1): 1416. doi : 10.1038/s41467-022-28984-4 . ПМЦ 8931054 . ПМИД 35301303 .
- ^ Лоссин С. «Информационная база SCN1A» . Архивировано из оригинала 21 июля 2011 г. Проверено 30 октября 2009 г.
компиляция генетических вариаций гена SCN1A, которые изменяют экспрессию или функцию Nav1.1
- ^ Jump up to: а б с д Беннетт Д.Л., Кларк А.Дж., Хуан Дж., Ваксман С.Г., Диб-Хадж С.Д. (апрель 2019 г.). «Роль потенциалзависимых натриевых каналов в передаче сигналов боли» . Физиологические обзоры . 99 (2): 1079–1151. дои : 10.1152/physrev.00052.2017 . ПМИД 30672368 .
- ^ Смит Р.С., Кенни С.Дж., Ганеш В., Джанг А., Борхес-Монрой Р., Партлоу Дж.Н. и др. (сентябрь 2018 г.). «V1.3) Регуляция складок коры головного мозга человека и развитие моторики полости рта» . Нейрон . 99 (5): 905–913.e7. дои : 10.1016/j.neuron.2018.07.052 . ПМК 6226006 . ПМИД 30146301 .
- ^ Чокалингам П., Уайлд А. (сентябрь 2012 г.). «Многогранный сердечный натриевый канал и его клиническое значение». Сердце . 98 (17): 1318–24. doi : 10.1136/heartjnl-2012-301784 . ПМИД 22875823 . S2CID 44433455 .
- ^ Бейдер А., Маццоне А., Стреге П.Р., Тестер DJ, Сайто Я.А., Бернард CE, Эндерс Ф.Т., Эк ВЕ, Шмидт П.Т., Длугош А., Линдберг Г., Карлинг П., Олссон Б., Газули М., Нардоне Г., Куомо Р., Усай- Сатта П., Галеацци Ф., Нери М., Портинкаса П., Беллини М., Барбара Дж., Камиллери М., Локк Г.Р., Талли Нью-Джерси, Д'Амато М., Акерман М.Дж., Фарруджа Дж. (июнь 2014 г.). «Потеря функции потенциалзависимого натриевого канала NaV1,5 (каналопатии) у больных с синдромом раздраженного кишечника» . Гастроэнтерология . 146 (7): 1659–1668. дои : 10.1053/j.gastro.2014.02.054 . ПМК 4096335 . ПМИД 24613995 .
- ^ Батлер К.М., да Силва С., Шафир Ю., Вайсфельд-Адамс Дж.Д., Александр Дж.Дж., Хегде М., Эскайг А. (январь 2017 г.). «De novo и унаследованные мутации эпилепсии SCN8A, обнаруженные с помощью анализа генной панели» . Исследования эпилепсии . 129 : 17–25. doi : 10.1016/j.eplepsyres.2016.11.002 . ПМК 5321682 . ПМИД 27875746 .
- ^ Мейслер М.Х., Кирни Дж.А. (август 2005 г.). «Мутации натриевых каналов при эпилепсии и других неврологических расстройствах» . Журнал клинических исследований . 115 (8): 2010–7. дои : 10.1172/JCI25466 . ПМК 1180547 . ПМИД 16075041 .
- ^ Варгас-Аларкон Дж., Альварес-Леон Э., Фрагосо Х.М., Варгас А., Мартинес А., Вальехо М., Мартинес-Лавин М. (февраль 2012 г.). «Полиморфизм натриевых каналов дорсальных корешковых ганглиев, кодируемый геном SCN9A, связанный с тяжелой фибромиалгией» . BMC Заболевания опорно-двигательного аппарата . 13 (1): 23. дои : 10.1186/1471-2474-13-23 . ПМК 3310736 . ПМИД 22348792 .
- ^ Каттералл, Вашингтон (апрель 2000 г.). «От ионных токов к молекулярным механизмам: структура и функции потенциалзависимых натриевых каналов» . Нейрон . 26 (1): 13–25. дои : 10.1016/S0896-6273(00)81133-2 . ПМИД 10798388 . S2CID 17928749 .
- ^ Исом Л.Л., Де Йонг К.С., Паттон Д.Е., Ребер Б.Ф., Оффорд Дж., Шарбонно Х., Уолш К., Голдин А.Л., Каттералл В.А. (май 1992 г.). «Первичная структура и функциональное выражение субъединицы бета 1 натриевого канала мозга крысы». Наука . 256 (5058): 839–42. Бибкод : 1992Sci...256..839I . дои : 10.1126/science.1375395 . ПМИД 1375395 .
- ^ «Сервер Blackboard недоступен» (PDF) . blackboard.jhu.edu . Проверено 16 июля 2020 г.
- ^ Малхотра Дж.Д., Казен-Гиллеспи К., Хорч М., Исом Л.Л. (апрель 2000 г.). «Бета-субъединицы натриевых каналов опосредуют гомофильную клеточную адгезию и рекрутируют анкирин в точки межклеточного контакта» . Журнал биологической химии . 275 (15): 11383–8. дои : 10.1074/jbc.275.15.11383 . ПМИД 10753953 .
- ^ Малхотра Дж.Д., Купманн М.К., Казен-Гиллеспи К.А., Феттман Н., Хорч М., Исом Л.Л. (июль 2002 г.). «Структурные требования для взаимодействия бета-1-субъединиц натриевого канала с анкирином» . Журнал биологической химии . 277 (29): 26681–8. дои : 10.1074/jbc.M202354200 . ПМИД 11997395 .
- ^ Кантрелл А.Р., Каттералл Вашингтон (июнь 2001 г.). «Нейромодуляция каналов Na+: неожиданная форма клеточной пластичности». Обзоры природы. Нейронаука . 2 (6): 397–407. дои : 10.1038/35077553 . ПМИД 11389473 . S2CID 22885909 .
- ^ Шах Б.С., Раш А.М., Лю С., Тиррел Л., Блэк Дж.А., Диб-Хадж С.Д., Ваксман С.Г. (август 2004 г.). «Контактин связывается с натриевым каналом Nav1.3 в нативных тканях и увеличивает плотность каналов на поверхности клетки» . Журнал неврологии . 24 (33): 7387–99. doi : 10.1523/JNEUROSCI.0322-04.2004 . ПМК 6729770 . ПМИД 15317864 .
- ^ Виттмак Э.К., Раш А.М., Крэнер М.Дж., Гольдфарб М., Ваксман С.Г., Диб-Хадж С.Д. (июль 2004 г.). «Гомологический фактор 2B фактора роста фибробластов: ассоциация с Nav1.6 и селективная колокализация в узлах Ранвье аксонов дорсальных корешков» . Журнал неврологии . 24 (30): 6765–75. doi : 10.1523/JNEUROSCI.1628-04.2004 . ПМК 6729706 . ПМИД 15282281 .
- ^ Раш А.М., Виттмак Э.К., Тиррелл Л., Блэк Дж.А., Диб-Хадж С.Д., Ваксман С.Г. (май 2006 г.). «Дифференциальная модуляция натриевого канала Na (v) 1,6 двумя членами подсемейства гомологичного фактора 2 фактора роста фибробластов». Европейский журнал неврологии . 23 (10): 2551–62. дои : 10.1111/j.1460-9568.2006.04789.x . ПМИД 16817858 . S2CID 21411801 .
- ^ Казаринова-Нойес К., Малхотра Дж.Д., МакИвен Д.П., Маттей Л.Н., Берглунд Э.О., Раншт Б., Левинсон С.Р., Шахнер М., Шрагер П., Исом Л.Л., Сяо З.К. (октябрь 2001 г.). «Контактин связывается с каналами Na+ и увеличивает их функциональную экспрессию» . Журнал неврологии . 21 (19): 7517–25. doi : 10.1523/JNEUROSCI.21-19-07517.2001 . ПМК 6762905 . ПМИД 11567041 .
- ^ Шринивасан Дж., Шахнер М., Каттералл В.А. (декабрь 1998 г.). «Взаимодействие потенциалзависимых натриевых каналов с молекулами внеклеточного матрикса тенасцин-С и тенасцин-R» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (26): 15753–7. Бибкод : 1998PNAS...9515753S . дои : 10.1073/pnas.95.26.15753 . ПМК 28116 . ПМИД 9861042 .
- ^ Ким Д.Ю., Ингано Л.А., Кэри Б.В., Петтингелл В.Х., Ковач Д.М. (июнь 2005 г.). «Пресенилин/гамма-секретаза-опосредованное расщепление бета2-субъединицы потенциалзависимого натриевого канала регулирует клеточную адгезию и миграцию» . Журнал биологической химии . 280 (24): 23251–61. дои : 10.1074/jbc.M412938200 . ПМИД 15833746 .
- ^ Рэтклифф К.Ф., Вестенбрук Р.Э., Кертис Р., Каттералл В.А. (июль 2001 г.). «Субъединицы бета1 и бета3 натриевых каналов связываются с нейрофасцином через свой внеклеточный иммуноглобулинподобный домен» . Журнал клеточной биологии . 154 (2): 427–34. дои : 10.1083/jcb.200102086 . ПМК 2150779 . ПМИД 11470829 .
- ^ Jump up to: а б с Анцелевич С, Паточкай Б (январь 2016 г.). «Синдром Бругада: клинические, генетические, молекулярные, клеточные и ионные аспекты» . Современные проблемы кардиологии . 41 (1): 7–57. doi : 10.1016/j.cpcardiol.2015.06.002 . ПМЦ 4737702 . ПМИД 26671757 .
- ^ Моран Й., Барзилай М.Г., Либескинд Б.Дж., Закон Х.Х. (февраль 2015 г.). «Эволюция потенциалзависимых ионных каналов при появлении Metazoa» . Журнал экспериментальной биологии . 218 (Часть 4): 515–25. дои : 10.1242/jeb.110270 . ПМИД 25696815 .
- ^ Либескинд Б.Дж., Хиллис Д.М., Закон Х.Х. (май 2011 г.). «Эволюция натриевых каналов предшествует возникновению нервной системы у животных» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (22): 9154–9. Бибкод : 2011PNAS..108.9154L . дои : 10.1073/pnas.1106363108 . ПМК 3107268 . ПМИД 21576472 .
- ^ Кинг Н., Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. и др. (февраль 2008 г.). «Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных животных» . Природа . 451 (7180): 783–8. Бибкод : 2008Natur.451..783K . дои : 10.1038/nature06617 . ПМК 2562698 . ПМИД 18273011 .
- ^ Jump up to: а б Стронг М., Чанди К.Г., Гутман Г.А. (январь 1993 г.). «Молекулярная эволюция генов потенциал-чувствительных ионных каналов: о происхождении электрической возбудимости» . Молекулярная биология и эволюция . 10 (1): 221–42. doi : 10.1093/oxfordjournals.molbev.a039986 . ПМИД 7680747 .
- ^ Либескинд Б.Дж., Хиллис Д.М., Закон Х.Х. (ноябрь 2013 г.). «Независимое приобретение селективности натрия в натриевых каналах бактерий и животных» . Современная биология . 23 (21): R948–9. дои : 10.1016/j.cub.2013.09.025 . ПМИД 24200318 .
- ^ Касимова М.А., Граната Д, Карневале В (2016). «Натриевые каналы, управляемые напряжением: эволюционная история и отличительные особенности последовательности». Актуальные темы мембран . Том. 78. стр. 261–86. дои : 10.1016/bs.ctm.2016.05.002 . ISBN 9780128053867 . ПМИД 27586287 .
- ^ Jump up to: а б Видмарк Дж., Сундстрем Г., Окампо Даса Д., Лархаммар Д. (январь 2011 г.). «Дифференциальная эволюция потенциалзависимых натриевых каналов у четвероногих и костистых рыб» . Молекулярная биология и эволюция . 28 (1): 859–71. дои : 10.1093/molbev/msq257 . ПМИД 20924084 .
- ^ Jump up to: а б Закон Х.Х., Йост MC, Лу Ю (апрель 2011 г.). «Расширение семейства генов потенциал-зависимых каналов Na+ у ранних четвероногих совпало с появлением наземного мира и увеличением сложности мозга» . Молекулярная биология и эволюция . 28 (4): 1415–24. дои : 10.1093/molbev/msq325 . ПМК 3058772 . ПМИД 21148285 .
- ^ Jump up to: а б Закон Х, Йост М, Цвикл Д, Лу Ю, Хиллис Д (2009). «Молекулярная эволюция Na+-каналов у костистых рыб». Интегративная зоология . 4 (1): 64–74. дои : 10.1111/j.1749-4877.2008.00136.x . ПМИД 21392277 .