Мембранный потенциал
Эта статья нуждается в дополнительных цитатах для проверки . ( август 2022 г. ) |
Мембранный потенциал (также трансмембранный потенциал или мембранное напряжение ) — это разница электрического потенциала между внутренней и внешней частью биологической клетки . Он равен внутреннему потенциалу минус внешний потенциал. Это энергия (т.е. работа ) на каждый заряд, которая необходима для перемещения (очень малого) положительного заряда с постоянной скоростью через клеточную мембрану снаружи внутрь. изменение кинетической энергии и образование излучения (Если заряду разрешено изменять скорость, необходимо учитывать .)
Типичные значения мембранного потенциала, обычно выражаемые в милливольтах и обозначаемые как мВ, находятся в диапазоне от –80 мВ до –40 мВ. Для таких типичных отрицательных мембранных потенциалов требуется положительная работа для перемещения положительного заряда изнутри наружу. Однако тепловая кинетическая энергия позволяет ионам преодолевать разность потенциалов. Для избирательно проницаемой мембраны это обеспечивает чистый поток против градиента. (Это своего рода осмос .)
Описание
[ редактировать ]Все клетки животных окружены мембраной , состоящей из липидного бислоя со встроенными в него белками . Мембрана служит одновременно изолятором и диффузионным барьером для движения ионов . Трансмембранные белки , также известные как белки -переносчики ионов или белки ионного насоса , активно проталкивают ионы через мембрану и создают градиенты концентрации через мембрану, а ионные каналы позволяют ионам перемещаться через мембрану вниз по этим градиентам концентрации. Ионные насосы и ионные каналы электрически эквивалентны набору батарей и резисторов, вставленных в мембрану, и, следовательно, создают напряжение между двумя сторонами мембраны.
Почти все плазматические мембраны имеют электрический потенциал, причем внутренняя часть обычно отрицательная по отношению к внешней. [1] Мембранный потенциал выполняет две основные функции. Во-первых, он позволяет клетке функционировать как батарея, обеспечивая питание для работы различных «молекулярных устройств», встроенных в мембрану. [2] Во-вторых, в электрически возбудимых клетках , таких как нейроны и мышечные клетки , он используется для передачи сигналов между различными частями клетки.
Сигналы в нейронах и мышечных клетках
[ редактировать ]Сигналы генерируются в возбудимых клетках путем открытия или закрытия ионных каналов в одной точке мембраны, вызывая локальное изменение мембранного потенциала. Это изменение электрического поля может быть быстро обнаружено соседними или более удаленными ионными каналами в мембране. Эти ионные каналы могут затем открываться или закрываться в результате изменения потенциала, воспроизводя сигнал.
В невозбудимых клетках и в возбудимых клетках в их исходном состоянии мембранный потенциал поддерживается на относительно стабильном значении, называемом потенциалом покоя . Для нейронов потенциал покоя определяется в пределах от –80 до –70 милливольт; то есть внутренняя часть ячейки имеет отрицательное базовое напряжение чуть меньше одной десятой вольта. Открытие и закрытие ионных каналов может вызвать отклонение от потенциала покоя. Это называется деполяризацией, если внутреннее напряжение становится менее отрицательным (скажем, от –70 мВ до –60 мВ) или гиперполяризацией, если внутреннее напряжение становится более отрицательным (скажем, от –70 мВ до –80 мВ). В возбудимых клетках достаточно большая деполяризация может вызвать потенциал действия , при котором мембранный потенциал меняется быстро и значительно в течение короткого времени (порядка от 1 до 100 миллисекунд), часто меняя свою полярность. Потенциалы действия генерируются путем активации определенных потенциалзависимых ионных каналов .
В нейронах факторы, влияющие на мембранный потенциал, разнообразны. Они включают в себя многочисленные типы ионных каналов, некоторые из которых являются химически управляемыми, а некоторые — потенциалзависимыми. Поскольку потенциалзависимые ионные каналы контролируются мембранным потенциалом, в то время как сам мембранный потенциал находится под влиянием этих же ионных каналов, возникают петли обратной связи, которые обеспечивают сложную временную динамику, включая колебания и регенеративные события, такие как потенциалы действия.
Градиенты концентрации ионов
[ редактировать ]
Большая фиолетовая структура со стрелкой представляет трансмембранный калиевый канал и направление чистого движения калия.
Различия в концентрациях ионов на противоположных сторонах клеточной мембраны приводят к возникновению напряжения, называемого мембранным потенциалом . [3]
Многие ионы имеют градиент концентрации через мембрану, в том числе ионы калия (K + ), который имеет высокую концентрацию внутри и низкую концентрацию снаружи мембраны. Натрий (Na + ) и хлорид (Cl − ) ионы находятся в высоких концентрациях во внеклеточной области и в низких концентрациях во внутриклеточных областях. Эти градиенты концентрации обеспечивают потенциальную энергию, способствующую образованию мембранного потенциала. Это напряжение устанавливается, когда мембрана проницаема для одного или нескольких ионов.
В простейшем случае, показанном здесь, если мембрана избирательно проницаема для калия, эти положительно заряженные ионы могут диффундировать по градиенту концентрации наружу клетки, оставляя после себя некомпенсированные отрицательные заряды. Такое разделение зарядов и вызывает мембранный потенциал. Система в целом электронейтральна. Некомпенсированные положительные заряды снаружи клетки и некомпенсированные отрицательные заряды внутри клетки физически выстраиваются на поверхности мембраны и притягиваются друг к другу через липидный бислой . Таким образом, мембранный потенциал физически расположен только в непосредственной близости от мембраны. Именно разделение этих зарядов на мембране лежит в основе мембранного напряжения.
Эта диаграмма представляет собой лишь приближение ионного вклада в мембранный потенциал. Другие ионы, включая натрий, хлорид, кальций и другие, играют менее важную роль, хотя у них сильный градиент концентрации, поскольку они имеют более ограниченную проницаемость, чем калий.
Физическая основа
[ редактировать ]Мембранный потенциал в клетке в конечном итоге зависит от двух факторов: электрической силы и диффузии. Электрическая сила возникает в результате взаимного притяжения частиц с противоположными электрическими зарядами (положительных и отрицательных) и взаимного отталкивания частиц с одинаковым типом заряда (положительных или обоих отрицательных). Диффузия возникает из-за статистической тенденции частиц перераспределяться из областей, где они высоко концентрированы, в области с низкой концентрацией.
Напряжение
[ редактировать ]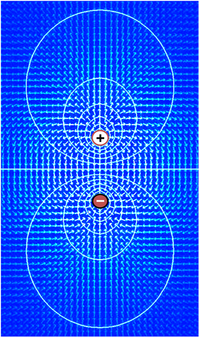
Напряжение, которое является синонимом разницы электрических потенциалов , представляет собой способность пропускать электрический ток через сопротивление. Действительно, самое простое определение напряжения дается законом Ома : V=IR, где V — напряжение, I — ток, а R — сопротивление. Если в электрическую цепь включен источник напряжения, например батарея, то чем выше напряжение источника, тем большую силу тока он будет пропускать через доступное сопротивление. Функциональное значение напряжения заключается только в разности потенциалов между двумя точками цепи. Идея напряжения в одной точке бессмысленна. В электронике принято присваивать нулевое напряжение некоторому произвольно выбранному элементу схемы, а затем присваивать напряжения для других элементов, измеряемые относительно этой нулевой точки. Не имеет значения, какой элемент выбран в качестве нулевой точки — функция цепи зависит только от разностей, а не от напряжений как таковых . Однако в большинстве случаев по соглашению нулевой уровень чаще всего присваивается той части цепи, которая находится в контакте с земля.
Тот же принцип применим к напряжению в клеточной биологии. В электрически активной ткани разность потенциалов между любыми двумя точками можно измерить, вставив электрод в каждую точку, например, один внутри и один снаружи клетки, и подключив оба электрода к выводам того, что по сути является специализированным вольтметром. По соглашению, нулевое значение потенциала присваивается внешней стороне клетки, а знак разности потенциалов между внешней и внутренней частью определяется потенциалом внутренней части относительно внешнего нуля.
С математической точки зрения определение напряжения начинается с понятия электрического поля E , векторного поля, задающего величину и направление каждой точке пространства. Во многих ситуациях электрическое поле является консервативным полем что его можно выразить как градиент скалярной функции V , то есть E = –∇ V. , а это означает , Это скалярное поле V называется распределением напряжения. Определение допускает произвольную константу интегрирования, поэтому абсолютные значения напряжения не имеют смысла. В общем, электрические поля можно рассматривать как консервативные только в том случае, если магнитные поля не оказывают на них существенного влияния, но это условие обычно хорошо применимо и к биологическим тканям.
Поскольку электрическое поле представляет собой градиент распределения напряжения, быстрые изменения напряжения в небольшой области подразумевают сильное электрическое поле; и наоборот, если напряжение остается примерно одинаковым на большой области, электрические поля в этой области должны быть слабыми. Сильное электрическое поле, эквивалентное сильному градиенту напряжения, означает, что на любые заряженные частицы, находящиеся в этой области, действует сильная сила.
Ионы и силы, управляющие их движением
[ редактировать ]
Электрические сигналы внутри биологических организмов, как правило, управляются ионами . [5] Наиболее важными катионами для потенциала действия являются натрий ( Na + ) и калий (K + ). [6] Оба они являются одновалентными катионами, несущими один положительный заряд. Потенциалы действия также могут включать кальций (Ca 2+ ), [7] Это двухвалентный катион, несущий двойной положительный заряд. Хлорид - анион (Cl − ) играет важную роль в потенциалах действия некоторых водорослей , [8] но играет незначительную роль в потенциалах действия большинства животных. [9]
Ионы пересекают клеточную мембрану под двумя воздействиями: диффузией и электрическими полями . Простой пример, когда два раствора — A и B — разделены пористым барьером, показывает, что диффузия гарантирует, что они в конечном итоге смешаются в равные растворы. Такое смешивание происходит из-за разницы их концентраций. Область с высокой концентрацией будет диффундировать в область с низкой концентрацией. В целях расширения примера предположим, что раствор А содержит 30 ионов натрия и 30 ионов хлорида. Также пусть раствор Б содержит только 20 ионов натрия и 20 ионов хлорида. Если предположить, что барьер позволяет проходить через него ионам обоих типов, то будет достигнуто устойчивое состояние, при котором оба раствора содержат 25 ионов натрия и 25 ионов хлорида. Однако если пористый барьер избирательно пропускает ионы, то сама по себе диффузия не будет определять конечный результат. Возвращаясь к предыдущему примеру, построим теперь барьер, проницаемый только для ионов натрия. Теперь только натрию разрешено диффундировать через барьер от его более высокой концентрации в растворе A к более низкой концентрации в растворе B. Это приведет к большему накоплению ионов натрия, чем ионов хлорида в растворе B, и меньшему количеству ионов натрия, чем ионы натрия. хлорид-ионы в растворе А.
Это означает, что в растворе B имеется суммарный положительный заряд из-за более высокой концентрации положительно заряженных ионов натрия, чем отрицательно заряженных ионов хлорида. Аналогичным образом, в растворе А имеется суммарный отрицательный заряд из-за большей концентрации отрицательных ионов хлорида, чем положительных ионов натрия. Поскольку противоположные заряды притягиваются, а подобные отталкиваются, на ионы теперь также влияют электрические поля, а также силы диффузии. Следовательно, положительные ионы натрия с меньшей вероятностью попадут в теперь более положительный раствор B и останутся в теперь более отрицательном растворе A. Точка, в которой силы электрического поля полностью противодействуют силе диффузии, называется равновесным потенциалом. В этот момент чистый поток конкретного иона (в данном случае натрия) равен нулю.
Плазматические мембраны
[ редактировать ]
Каждая клетка окружена плазматической мембраной , которая имеет структуру липидного бислоя , в который встроено множество типов крупных молекул. Поскольку плазматическая мембрана состоит из молекул липидов, она по своей природе обладает высоким электрическим сопротивлением, другими словами, низкой собственной проницаемостью для ионов. Однако некоторые молекулы, внедренные в мембрану, способны либо активно транспортировать ионы с одной стороны мембраны на другую, либо обеспечивать каналы, по которым они могут перемещаться. [10]
В электрической терминологии плазматическая мембрана функционирует как комбинированный резистор и конденсатор . Сопротивление возникает из-за того, что мембрана препятствует движению зарядов через нее. Емкость возникает из-за того, что липидный бислой настолько тонкий, что скопление заряженных частиц на одной стороне приводит к возникновению электрической силы, которая притягивает противоположно заряженные частицы к другой стороне. На емкость мембраны относительно не влияют внедренные в нее молекулы, поэтому она имеет более или менее инвариантное значение, оцениваемое в 2 мкФ/см. 2 (общая емкость участка мембраны пропорциональна его площади). С другой стороны, проводимость чистого липидного бислоя настолько низка, что в биологических ситуациях в ней всегда доминирует проводимость альтернативных путей, обеспечиваемых встроенными молекулами. Таким образом, емкость мембраны более или менее постоянна, а вот сопротивление сильно варьируется.
Толщина плазматической мембраны оценивается примерно в 7-8 нанометров. Поскольку мембрана очень тонкая, для создания внутри нее сильного электрического поля не требуется очень большого трансмембранного напряжения. Типичные мембранные потенциалы в клетках животных составляют порядка 100 милливольт (то есть одна десятая вольта), но расчеты показывают, что это создает электрическое поле, близкое к максимальному, которое может выдержать мембрана. Разница, намного превышающая 200 милливольт, может вызвать пробой диэлектрика , то есть искрение на мембране.
Облегченная диффузия и транспорт
[ редактировать ]
Сопротивление чистого липидного бислоя прохождению через него ионов очень велико, но структуры, встроенные в мембрану, могут значительно усиливать движение ионов, активно или пассивно , посредством механизмов, называемых облегченным транспортом и облегченной диффузией . Два типа структур, которые играют наибольшую роль, — это ионные каналы и ионные насосы , которые обычно образуются из скоплений белковых молекул. Ионные каналы обеспечивают проходы, по которым могут перемещаться ионы. В большинстве случаев ионный канал проницаем только для определенных типов ионов (например, натрия и калия, но не для хлорида или кальция), а иногда проницаемость варьируется в зависимости от направления движения ионов. Ионные насосы, также известные как переносчики ионов или белки-переносчики, активно транспортируют определенные типы ионов с одной стороны мембраны на другую, иногда используя для этого энергию, полученную в результате метаболических процессов.
Ионные насосы
[ редактировать ]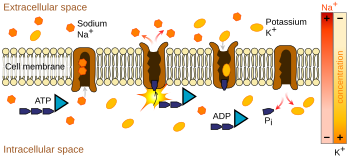
Ионные насосы представляют собой интегральные мембранные белки, которые осуществляют активный транспорт , т. е. используют клеточную энергию (АТФ) для «перекачки» ионов против градиента их концентрации. [11] Такие ионные насосы забирают ионы с одной стороны мембраны (снижая там их концентрацию) и высвобождают их с другой стороны (увеличивая там их концентрацию).
Ионный насос, наиболее отвечающий потенциалу действия, — это натриево-калиевый насос , который транспортирует три иона натрия из клетки и два иона калия внутрь. [12] [13] Как следствие, концентрация ионов калия K + внутри нейрона примерно в 30 раз больше, чем снаружи, тогда как концентрация натрия снаружи примерно в пять раз больше, чем внутри. [13] [14] [15] Аналогичным образом другие ионы имеют разные концентрации внутри и снаружи нейрона, например, кальций , хлорид и магний . [15]
Если бы количество ионов каждого типа было равным, натриево-калиевый насос был бы электрически нейтральным, но из-за обмена три к двум он обеспечивает чистое перемещение одного положительного заряда от внутриклеточного к внеклеточному за каждый цикл. тем самым способствуя положительной разности напряжений. Насос имеет три эффекта: (1) он повышает концентрацию натрия во внеклеточном пространстве и снижает во внутриклеточном пространстве; (2) повышает концентрацию калия во внутриклеточном пространстве и снижает во внеклеточном пространстве; (3) он придает внутриклеточному пространству отрицательное напряжение по отношению к внеклеточному пространству.
Натриево-калиевый насос работает относительно медленно. Если бы ячейка была инициализирована повсюду с одинаковыми концентрациями натрия и калия, насосу потребовалось бы несколько часов, чтобы установить равновесие. Насос работает постоянно, но его эффективность становится все менее эффективной по мере снижения концентрации натрия и калия, доступных для перекачки.
Ионные насосы влияют на потенциал действия только путем установления относительного соотношения внутриклеточных и внеклеточных концентраций ионов. Потенциал действия включает главным образом открытие и закрытие ионных каналов, а не ионных насосов. Если ионные насосы отключить, удалив их источник энергии или добавив ингибитор, такой как уабаин , аксон все равно может запустить сотни тысяч потенциалов действия, прежде чем их амплитуды начнут значительно уменьшаться. [11] В частности, ионные насосы не играют существенной роли в реполяризации мембраны после потенциала действия. [6]
Еще одним функционально важным ионным насосом является натрий-кальциевый обменник . Этот насос работает концептуально аналогично натрий-калиевому насосу, за исключением того, что в каждом цикле он обменивает три Na + из внеклеточного пространства за один Са ++ из внутриклеточного пространства. Поскольку чистый поток заряда направлен внутрь, этот насос фактически работает «под гору» и, следовательно, не требует какого-либо источника энергии, кроме напряжения на мембране. Его наиболее важным эффектом является выкачивание кальция наружу; он также обеспечивает приток натрия внутрь, тем самым противодействуя натрий-калиевому насосу, но, поскольку общие концентрации натрия и калия намного выше, чем концентрации кальция, этот эффект относительно неважен. Конечным результатом работы натрий-кальциевого обменника является то, что в состоянии покоя внутриклеточная концентрация кальция становится очень низкой.
Ионные каналы
[ редактировать ]
Ионные каналы представляют собой интегральные мембранные белки с порами, через которые ионы могут перемещаться между внеклеточным пространством и внутренней частью клетки. Большинство каналов специфичны (селективны) для одного иона; например, большинство калиевых каналов характеризуются коэффициентом селективности в отношении калия по сравнению с натрием 1000:1, хотя ионы калия и натрия имеют одинаковый заряд и лишь незначительно различаются по своему радиусу. Пора канала обычно настолько мала, что ионы должны проходить через нее в порядке одного файла. [17] Поры каналов могут быть открытыми или закрытыми для прохождения ионов, хотя ряд каналов демонстрируют различные уровни субпроводимости. Когда канал открыт, ионы проникают через поры канала вниз по трансмембранному градиенту концентрации для этого конкретного иона. Скорость ионного потока через канал, т.е. амплитуда одноканального тока, определяется максимальной проводимостью канала и электрохимической движущей силой для этого иона, которая представляет собой разницу между мгновенным значением мембранного потенциала и значением обратного потенциала . [18]

Канал может иметь несколько различных состояний (соответствующих разным конформациям белка), но каждое такое состояние является либо открытым, либо закрытым. В общем случае закрытые состояния соответствуют либо сжатию поры, что делает ее непроходимой для иона, либо отдельной части белка, закупоривающей пору. Например, потенциалзависимый натриевый канал подвергается инактивации , при которой часть белка попадает в пору, закрывая ее. [19] Эта инактивация отключает ток натрия и играет решающую роль в потенциале действия.
Ионные каналы можно классифицировать по тому, как они реагируют на окружающую среду. [20] Например, ионные каналы, участвующие в формировании потенциала действия, являются чувствительными к напряжению каналами ; они открываются и закрываются в ответ на напряжение на мембране. Лиганд-зависимые каналы образуют еще один важный класс; эти ионные каналы открываются и закрываются в ответ на связывание молекулы-лиганда , например нейромедиатора . Другие ионные каналы открываются и закрываются под действием механических сил. Другие ионные каналы, например каналы сенсорных нейронов , открываются и закрываются в ответ на другие стимулы, такие как свет, температура или давление.
Каналы утечки
[ редактировать ]Каналы утечки представляют собой простейший тип ионных каналов, поскольку их проницаемость более или менее постоянна. Наибольшее значение в нейронах имеют каналы утечки, калиевые и хлоридные каналы. Даже они не совсем постоянны по своим свойствам: во-первых, большинство из них зависят от напряжения в том смысле, что проводят лучше в одном направлении, чем в другом (другими словами, они являются выпрямителями ); во-вторых, некоторые из них способны блокироваться химическими лигандами, даже если для их работы лиганды не требуются.
Лиганд-зависимые каналы
[ редактировать ]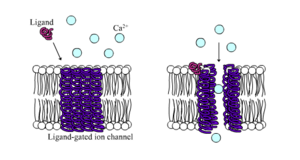
Лиганд-управляемые ионные каналы — это каналы, проницаемость которых значительно увеличивается, когда какой-либо тип химического лиганда связывается со структурой белка. Клетки животных содержат сотни, если не тысячи их типов. Большая их часть выполняет функцию рецепторов нейротрансмиттеров — они встречаются в постсинаптических участках, и химический лиганд, который их воротает, высвобождается пресинаптическим окончанием аксона . Одним из примеров этого типа является рецептор AMPA , рецептор нейромедиатора глутамата , который при активации обеспечивает прохождение ионов натрия и калия. Другим примером является ГАМК А рецептор , рецептор нейромедиатора ГАМК , который при активации позволяет проходить ионам хлорида.
Рецепторы нейромедиаторов активируются лигандами, которые появляются во внеклеточной области, но существуют и другие типы лиганд-управляемых каналов, которые контролируются взаимодействиями на внутриклеточной стороне.
Каналы, зависящие от напряжения
[ редактировать ]Потенциал-управляемые ионные каналы , также известные как потенциал-зависимые ионные каналы , представляют собой каналы, на проницаемость которых влияет мембранный потенциал. Они образуют еще одну очень большую группу, каждый член которой имеет определенную ионную селективность и определенную зависимость от напряжения. Многие из них также зависят от времени — другими словами, они не реагируют на изменение напряжения немедленно, а только после задержки.
Одним из наиболее важных членов этой группы является тип потенциалзависимых натриевых каналов, лежащих в основе потенциалов действия. Их иногда называют натриевыми каналами Ходжкина-Хаксли, поскольку первоначально они были охарактеризованы Аланом Ллойдом Ходжкином и Эндрю Хаксли в их работе, получившей Нобелевскую премию. исследования физиологии потенциала действия. Канал закрывается при уровне напряжения покоя, но резко открывается, когда напряжение превышает определенный порог, обеспечивая большой приток ионов натрия, что приводит к очень быстрому изменению мембранного потенциала. Восстановление после потенциала действия частично зависит от типа потенциалзависимого калиевого канала, который закрывается на уровне напряжения покоя, но открывается вследствие большого изменения напряжения, возникающего во время потенциала действия.
Потенциал разворота
[ редактировать ]Обратный потенциал (или равновесный потенциал ) иона — это значение трансмембранного напряжения, при котором диффузионные и электрические силы уравновешиваются, так что нет чистого потока ионов через мембрану. Это означает, что трансмембранное напряжение точно противодействует силе диффузии иона, так что чистый ток иона через мембрану равен нулю и неизменен. Обратный потенциал важен, потому что он создает напряжение, которое действует на каналы, проницаемые для этого иона, — другими словами, он дает напряжение, которое генерирует градиент концентрации ионов, когда он действует как батарея .
Равновесный потенциал конкретного иона обычно обозначается обозначением E ion . Равновесный потенциал для любого иона можно рассчитать с помощью уравнения Нернста . [21] Например, обратимый потенциал для ионов калия будет следующим:
![{\displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}\ln {\frac {[K^{+}]_{o}}{[K^{+}] _{я}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/f898ec5e5474736ecfe73462fdcb4b4d92a21618)
где
- Е экв,К + = равновесный потенциал калия, измеряемый в вольтах
- R = универсальная газовая постоянная , равная 8,314 джоуля ·К. −1 ·моль −1
- T = абсолютная температура , измеренная в кельвинах (= K = градусы Цельсия + 273,15)
- z = количество элементарных зарядов рассматриваемого иона, участвующего в реакции
- F = постоянная Фарадея , равная 96 485 кулонов ·моль. −1 или СП −1 ·моль −1
- [К + ] o = внеклеточная концентрация калия, измеренная в моль ·м −3 или ммоль·л −1
- [К + ] i = внутриклеточная концентрация калия
Даже если два разных иона имеют одинаковый заряд (т. е. K + и На + ), они все равно могут иметь очень разные равновесные потенциалы, при условии, что их внешняя и/или внутренняя концентрации различаются. Возьмем, к примеру, равновесные потенциалы калия и натрия в нейронах. Равновесный потенциал калия E K составляет -84 мВ при 5 мМ калия снаружи и 140 мМ внутри. С другой стороны, равновесный потенциал натрия E Na составляет примерно +66 мВ с примерно 12 мМ натрия внутри и 140 мМ снаружи. [примечание 1]
Изменения мембранного потенциала в процессе развития
[ редактировать ]развития Мембранный потенциал покоя нейрона фактически меняется в ходе организма . Чтобы нейрон в конечном итоге смог полностью реализовать свою взрослую функцию, его потенциал должен жестко регулироваться во время развития. По мере развития организма мембранный потенциал покоя становится более отрицательным. [22] Глиальные клетки также дифференцируются и пролиферируют по мере развития мозга . [23] Добавление этих глиальных клеток увеличивает способность организма регулировать внеклеточный калий . Падение внеклеточного калия может привести к снижению мембранного потенциала на 35 мВ. [24]
Возбудимость клеток
[ редактировать ]Возбудимость клеток — это изменение мембранного потенциала, необходимое для клеточных реакций в различных тканях. Возбудимость клеток — это свойство, которое индуцируется во время раннего эмбриогенеза. [25] Возбудимость клетки также определяется как легкость, с которой может быть вызван ответ. [26] Потенциалы покоя и пороговые потенциалы составляют основу возбудимости клеток, и эти процессы имеют основополагающее значение для генерации градуированных потенциалов и потенциалов действия.
Важнейшими регуляторами возбудимости клеток являются концентрации внеклеточных электролитов (т.е. Na + , К + , Как 2+ , кл − , мг 2+ ) и ассоциированные белки. Важными белками, которые регулируют возбудимость клеток, являются потенциалзависимые ионные каналы , транспортеры ионов (например, Na+/K+-АТФаза , транспортеры магния , кислотно-основные транспортеры ), мембранные рецепторы и активируемые гиперполяризацией циклические нуклеотидные каналы . [27] Например, калиевые каналы и кальций-чувствительные рецепторы являются важными регуляторами возбудимости в нейронах , кардиомиоцитах и многих других возбудимых клетках, таких как астроциты . [28] Ион кальция также является наиболее важным вторичным мессенджером в передаче сигналов возбудимых клеток . Активация синаптических рецепторов инициирует длительные изменения возбудимости нейронов. [29] Гормоны щитовидной железы , надпочечников и другие гормоны также регулируют возбудимость клеток, например, прогестерон и эстроген модулируют возбудимость гладкомышечных клеток миометрия .
Считается, что многие типы клеток имеют возбудимую мембрану. Возбудимыми клетками являются нейроны, мышцы ( сердечные , скелетные , гладкие ), эндотелиальные клетки сосудов , перициты , юкстагломерулярные клетки , интерстициальные клетки Кахаля , многие типы эпителиальных клеток (например, бета-клетки , альфа-клетки , дельта-клетки , энтероэндокринные клетки , легочные нейроэндокринные клетки) . , пинеалоциты ), глиальные клетки (например, астроциты), механорецепторные клетки (например, волосковые клетки и клетки Меркеля ), хеморецепторные клетки (например, гломусные клетки , вкусовые рецепторы ), некоторые растительные клетки и, возможно, иммунные клетки . [30] Астроциты проявляют форму неэлектрической возбудимости, основанную на внутриклеточных вариациях кальция, связанных с экспрессией нескольких рецепторов, посредством которых они могут обнаруживать синаптический сигнал. В нейронах в некоторых частях клетки имеются разные свойства мембран, например, дендритная возбудимость наделяет нейроны способностью обнаруживать совпадения пространственно разделенных входных сигналов. [31]
Эквивалентная схема
[ редактировать ]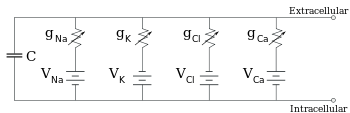
Электрофизиологи моделируют эффекты различий в концентрации ионов, ионных каналов и емкости мембраны с точки зрения эквивалентной схемы , которая предназначена для представления электрических свойств небольшого участка мембраны. Эквивалентная схема состоит из конденсатора, включенного параллельно с четырьмя дорожками, каждый из которых состоит из последовательно включенной батареи с переменной проводимостью. Емкость определяется свойствами липидного бислоя и считается постоянной. Каждый из четырех параллельных путей исходит от одного из основных ионов: натрия, калия, хлорида и кальция. Напряжение каждого ионного пути определяется концентрацией ионов на каждой стороне мембраны; см. раздел «Потенциал разворота» выше. Проводимость каждого ионного пути в любой момент времени определяется состояниями всех ионных каналов, которые потенциально проницаемы для этого иона, включая каналы утечки, лиганд-управляемые каналы и потенциал-управляемые ионные каналы.
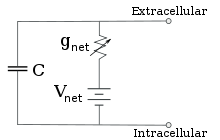
Для фиксированных концентраций ионов и фиксированных значений проводимости ионного канала эквивалентную схему можно свести, используя уравнение Гольдмана , как описано ниже, к схеме, содержащей емкость параллельно с батареей и проводимостью. В электрическом плане это разновидность RC-цепи (резистивно-емкостной цепи), и ее электрические свойства очень просты. Начиная с любого начального состояния, ток, текущий через проводимость или емкость, затухает с экспоненциальным ходом времени, с постоянной времени τ = RC , где C — емкость мембранного участка, а = 1/g net R чистое сопротивление. В реальных ситуациях постоянная времени обычно находится в диапазоне 1–100 миллисекунд. В большинстве случаев изменения проводимости ионных каналов происходят в более быстром временном масштабе, поэтому RC-цепь не является хорошим приближением; однако дифференциальное уравнение, используемое для моделирования мембранного участка, обычно представляет собой модифицированную версию уравнения RC-цепи.
Потенциал покоя
[ редактировать ]Когда мембранный потенциал клетки в течение длительного периода времени существенно не меняется, его называют потенциалом покоя или напряжением покоя. Этот термин используется для обозначения мембранного потенциала невозбудимых клеток, а также мембранного потенциала возбудимых клеток в отсутствие возбуждения. В возбудимых клетках другими возможными состояниями являются градуированные мембранные потенциалы (переменной амплитуды) и большие потенциалы действия, повышающие мембранный потенциал по принципу «все или ничего», которые обычно следуют фиксированному временному графику. Возбудимые клетки включают нейроны , мышечные клетки и некоторые секреторные клетки желез . Однако даже в других типах клеток мембранное напряжение может претерпевать изменения в ответ на внешние или внутриклеточные стимулы. Например, деполяризация плазматической мембраны, по-видимому, является важным шагом в запрограммированной гибели клеток . [32]
Взаимодействия, генерирующие потенциал покоя, моделируются уравнением Гольдмана . [33] По форме оно похоже на уравнение Нернста, показанное выше, поскольку оно основано на зарядах рассматриваемых ионов, а также на разнице между их внутренней и внешней концентрацией. Однако он также учитывает относительную проницаемость плазматической мембраны для каждого рассматриваемого иона.
![{\displaystyle E_{m}={\frac {RT}{F}}\ln {\left({\frac {P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\ mathrm {out} }+P_{\mathrm {Na} }[\mathrm {Na} ^{+}]_{\mathrm {out} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{ -}]_{\mathrm {in} }}{P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\mathrm {in} }+P_{\mathrm {Na} }[ \mathrm {Na} ^{+}]_{\mathrm {in} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{-}]_{\mathrm {out} }}}\right )}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/4b29151571b6fb807674bf342d01f178908a1a98)
Три иона, которые появляются в этом уравнении, — это калий (K + ), sodium (Na + ) и хлорид (Cl − ). Кальций не используется, но его можно добавить в ситуациях, в которых он играет значительную роль. [34] Поскольку хлоридные термины являются анионами, их трактуют иначе, чем катионные; внутриклеточная концентрация указана в числителе, а внеклеточная концентрация - в знаменателе, обратном катионным членам. Pi означает относительную проницаемость иона типа i.
По сути, формула Гольдмана выражает мембранный потенциал как средневзвешенное значение реверсивных потенциалов для отдельных типов ионов, взвешенных по проницаемости. (Хотя мембранный потенциал изменяется примерно на 100 мВ во время потенциала действия, концентрации ионов внутри и снаружи клетки изменяются незначительно. Они остаются близкими к соответствующим концентрациям, когда мембрана находится в состоянии покоя.) В большинстве животных клеток проницаемость для калия в состоянии покоя значительно выше, чем проницаемость для натрия. Как следствие, потенциал покоя обычно близок к потенциалу обращения калия. [35] [36] Проницаемость для хлоридов может быть достаточно высокой, чтобы быть значительной, но, в отличие от других ионов, хлорид не перекачивается активно и поэтому уравновешивается при обратимом потенциале, очень близком к потенциалу покоя, определяемому другими ионами.
Значения мембранного потенциала покоя в большинстве клеток животных обычно варьируются от потенциала обращения калия (обычно около -80 мВ) до около -40 мВ. Потенциал покоя в возбудимых клетках (способных создавать потенциалы действия) обычно составляет около -60 мВ - более деполяризованные напряжения могут привести к спонтанной генерации потенциалов действия. Незрелые или недифференцированные клетки демонстрируют сильно варьирующие значения напряжения покоя, обычно значительно более положительные, чем в дифференцированных клетках. [37] В таких клетках величина потенциала покоя коррелирует со степенью дифференцировки: недифференцированные клетки в ряде случаев могут вообще не обнаруживать разницы трансмембранных напряжений.
Поддержание потенциала покоя может быть метаболически дорогостоящим для клетки из-за необходимости активной перекачки ионов для противодействия потерям из-за каналов утечки. Цена самая высокая, когда функция клетки требует особенно деполяризованного значения мембранного напряжения. Например, потенциал покоя адаптированных к дневному свету мясной мухи ( Calliphora vicina ) фоторецепторов может достигать -30 мВ. [38] Этот повышенный мембранный потенциал позволяет клеткам очень быстро реагировать на зрительные сигналы; Цена заключается в том, что поддержание потенциала покоя может потреблять более 20% общего клеточного АТФ . [39]
С другой стороны, высокий потенциал покоя в недифференцированных клетках не обязательно влечет за собой высокие метаболические затраты. Этот очевидный парадокс разрешается путем изучения происхождения потенциала покоя. Малодифференцированные клетки характеризуются чрезвычайно высоким входным сопротивлением, [37] это означает, что на этой стадии жизни клетки присутствует мало каналов утечки. В результате проницаемость для калия становится аналогичной проницаемости для ионов натрия, что помещает потенциал покоя между обратными потенциалами для натрия и калия, как обсуждалось выше. Снижение токов утечки также означает, что для компенсации не требуется активной накачки, а значит, и низкие метаболические затраты.
Градуированные потенциалы
[ редактировать ]Как объяснялось выше, потенциал в любой точке клеточной мембраны определяется разницей в концентрации ионов между внутриклеточными и внеклеточными областями, а также проницаемостью мембраны для каждого типа ионов. Концентрации ионов обычно изменяются не очень быстро (за исключением Ca 2+ , где базовая внутриклеточная концентрация настолько низка, что даже небольшой приток может увеличить ее на порядки), но проницаемость ионов может измениться за доли миллисекунды в результате активации лиганд-управляемых ионных каналов. Изменение мембранного потенциала может быть большим или малым, в зависимости от того, сколько ионных каналов активировано и какого они типа, а также может быть длительным или коротким, в зависимости от продолжительности времени, в течение которого каналы остаются открытыми. Изменения этого типа называются градуированными потенциалами , в отличие от потенциалов действия, которые имеют фиксированную амплитуду и временной ход.
Как можно вывести из уравнения Гольдмана, показанного выше, эффект увеличения проницаемости мембраны для иона определенного типа смещает мембранный потенциал в сторону обратимого потенциала для этого иона. Таким образом, открытие Na + каналов смещает мембранный потенциал в сторону Na + реверсивный потенциал, который обычно составляет около +100 мВ. Аналогично, открытие K + каналов смещает мембранный потенциал примерно в сторону –90 мВ, и открытие Cl − каналов смещает его примерно в сторону –70 мВ (потенциал покоя большинства мембран). Таким образом, На + каналы смещают мембранный потенциал в положительную сторону, К + каналы смещают его в отрицательном направлении (кроме случаев, когда мембрана гиперполяризована до значения, более отрицательного, чем K + реверсивный потенциал) и Cl − каналы имеют тенденцию смещать его в сторону потенциала покоя.
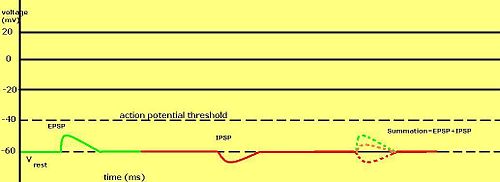
Градуированные мембранные потенциалы особенно важны в нейронах , где они производятся синапсами — временное изменение мембранного потенциала, вызванное активацией синапса одним градуированным потенциалом или потенциалом действия, называется постсинаптическим потенциалом . Нейромедиаторы , открывающие Na + каналы обычно заставляют мембранный потенциал становиться более положительным, в то время как нейротрансмиттеры, активирующие K + каналы обычно делают его более негативным; те, кто блокирует эти каналы, имеют тенденцию иметь противоположный эффект.
Считается ли постсинаптический потенциал возбуждающим или тормозящим, зависит от потенциала разворота ионов этого тока и порога срабатывания клеткой потенциала действия (около –50 мВ). Постсинаптический ток с реверсивным потенциалом выше порога, такой как типичный Na + ток считается возбуждающим. Ток с реверсивным потенциалом ниже порогового, например, типичный K + ток считается тормозным. Ток с реверсивным потенциалом выше потенциала покоя, но ниже порогового, сам по себе не вызывает потенциалы действия, но вызывает подпороговые колебания мембранного потенциала . Таким образом, нейротрансмиттеры, открывающие Na + каналы производят возбуждающие постсинаптические потенциалы , или ВПСП, тогда как нейротрансмиттеры, которые действуют, открывая K + или Cl − каналы обычно производят тормозные постсинаптические потенциалы или ТПСП. Когда в течение одного и того же периода времени открыты несколько типов каналов, их постсинаптические потенциалы суммируются (суммируются).
Другие значения
[ редактировать ]С точки зрения биофизики мембранный потенциал покоя — это просто мембранный потенциал, возникающий в результате проницаемости мембраны, которая преобладает, когда клетка отдыхает. Приведенное выше уравнение средневзвешенных значений применимо всегда, но следующий подход легче визуализировать.В любой момент времени для иона существуют два фактора, которые определяют, какое влияние этот ион окажет на мембранный потенциал клетки:
- Движущая сила этого иона
- Проницаемость этого иона
Если движущая сила велика, ион «проталкивается» через мембрану. Если проницаемость высока, иону будет легче диффундировать через мембрану.
- Движущая сила — это чистая электрическая сила, способная переместить ион через мембрану. Он рассчитывается как разница между напряжением, при котором ион «хочет» находиться (его равновесным потенциалом), и действительным мембранным потенциалом ( E m ). Итак, формально движущая сила для иона = E m – E ion
- Например, при рассчитанном нами ранее потенциале покоя, равном -73 мВ, движущая сила калия составляет 7 мВ: (-73 мВ) - (-80 мВ) = 7 мВ. Движущая сила натрия будет (-73 мВ) - (60 мВ) = -133 мВ.
- Проницаемость — это мера того, насколько легко ион может пересекать мембрану. Обычно она измеряется как (электрическая) проводимость, а единица измерения — сименс — соответствует 1 К·с. −1 ·V −1 , то есть один кулон в секунду на вольт потенциала.
Итак, в покоящейся мембране, хотя движущая сила для калия невелика, ее проницаемость очень высока. Натрий обладает огромной движущей силой, но почти не обладает проницаемостью в состоянии покоя. В этом случае калий проводит примерно в 20 раз больший ток, чем натрий, и, таким образом, оказывает в 20 раз большее влияние на Эм , чем натрий.
Однако рассмотрим другой случай — пик потенциала действия. Здесь проницаемость для Na высокая, а проницаемость для K относительно низкая. образом, мембрана перемещается в сторону E Na и вдаль от EK Таким .
Чем больше ионов проникает, тем сложнее становится предсказать мембранный потенциал. Однако это можно сделать с помощью уравнения Гольдмана-Ходжкина-Каца или уравнения взвешенных средних. Подставив градиенты концентрации и проницаемости ионов в любой момент времени, можно определить мембранный потенциал в этот момент. Уравнения GHK означают, что в любой момент значение мембранного потенциала будет представлять собой средневзвешенное значение равновесных потенциалов всех проникающих ионов. «Вес» представляет собой относительную проницаемость ионов через мембрану.
Эффекты и последствия
[ редактировать ]В то время как клетки тратят энергию на транспортировку ионов и создание трансмембранного потенциала, они, в свою очередь, используют этот потенциал для транспортировки других ионов и метаболитов, таких как сахар. Трансмембранный потенциал митохондрий стимулирует выработку АТФ , которая является общей валютой биологической энергии.
Клетки могут использовать энергию, которую они хранят в потенциале покоя, для запуска потенциалов действия или других форм возбуждения. Эти изменения мембранного потенциала обеспечивают связь с другими клетками (как и потенциалы действия) или инициируют изменения внутри клетки, что происходит в яйцеклетке , она оплодотворяется сперматозоидом когда .
Изменения диэлектрических свойств плазматической мембраны могут служить признаком таких заболеваний, как диабет и дислипидемия. [40]
В нейрональных клетках потенциал действия начинается с притока ионов натрия в клетку через натриевые каналы, что приводит к деполяризации, тогда как восстановление включает выброс калия наружу через калиевые каналы. Оба эти потока возникают путем пассивной диффузии .
Доза соли может заставить все еще работающие нейроны свежего куска мяса активироваться, вызывая мышечные спазмы. [41] [42] [43] [44] [45]
См. также
[ редактировать ]- Биоэлектрохимия
- Хемиосмотический потенциал
- Электрохимический потенциал
- Уравнение Гольдмана
- Мембранная биофизика
- Микроэлектродная матрица
- Сальтаторная проводимость
- Поверхностный потенциал
- Эффект Гиббса – Доннана
- Синаптический потенциал
Примечания
[ редактировать ]- ^ Знаки E Na и E K противоположны. Это связано с тем, что градиент концентрации калия направлен наружу клетки, а градиент концентрации натрия направлен внутрь клетки. Мембранные потенциалы определяются относительно внешней части клетки; таким образом, потенциал -70 мВ означает, что внутренняя часть клетки отрицательна по отношению к внешней.
Ссылки
[ редактировать ]- ^ Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Морган, Дэвид; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк: WW Norton & Company . ISBN 9780815344322 . OCLC 887605755 .
- ^ Абдул Кадир, Лина; Стейси, Майкл; Барретт-Джолли, Ричард (2018). «Новые роли мембранного потенциала: действие за пределами потенциала действия» . Границы в физиологии . 9 : 1661. дои : 10.3389/fphys.2018.01661 . ISSN 1664-042X . ПМК 6258788 . ПМИД 30519193 .
- ^ Яффе, Лайонел Ф.; Нуччителли, Ричард (1977). «Электрическое управление развитием» (PDF) . Ежегодный обзор биофизики и биоинженерии . 6 (1): 445–476. дои : 10.1146/annurev.bb.06.060177.002305 . ПМИД 326151 . Проверено 1 июня 2024 г.
- ^ Биология Кэмпбелла, 6-е издание
- ^ Джонстон и Ву, с. 9.
- ^ Перейти обратно: а б Буллок , Орканд и Гриннелл, стр. 140–41.
- ^ Буллок , Орканд и Гриннелл, стр. 153–54.
- ^ Муммерт Х., Градманн Д. (1991). «Потенциалы действия в вертлужной впадине: измерение и моделирование потенциалзависимых потоков». Журнал мембранной биологии . 124 (3): 265–73. дои : 10.1007/BF01994359 . ПМИД 1664861 . S2CID 22063907 .
- ^ Шмидт-Нильсен , стр. 483.
- ^ Либ В.Р., Штейн В.Д. (1986). «Глава 2. Простая диффузия через мембранный барьер». Транспорт и диффузия через клеточные мембраны . Сан-Диего: Академическая пресса. стр. 69–112. ISBN 978-0-12-664661-0 .
- ^ Перейти обратно: а б Ходжкин А.Л. , Кейнс Р.Д. (1955). «Активный транспорт катионов в гигантских аксонах Sepia и Loligo » . Дж. Физиол . 128 (1): 28–60. doi : 10.1113/jphysicalol.1955.sp005290 . ПМЦ 1365754 . PMID 14368574 .
- ^ Колдуэлл ПК, Ходжкин А.Л. , Кейнс Р.Д. , Шоу Т.И. (1960). «Влияние инъекции богатых энергией фосфатных соединений на активный транспорт ионов в гигантских аксонах Лолиго » . Дж. Физиол . 152 (3): 561–90. дои : 10.1113/jphysicalol.1960.sp006509 . ПМЦ 1363339 . ПМИД 13806926 .
- ^ Перейти обратно: а б Ганьон КБ, Дельпир Э (2021). «Переносчики натрия в здоровье и болезнях человека (рис. 2)» . Границы в физиологии . 11 : 588664. doi : 10.3389/fphys.2020.588664 . ПМЦ 7947867 . ПМИД 33716756 .
- ^ Штайнбах Х.Б., Шпигельман С (1943). «Баланс натрия и калия в аксоплазме нервов кальмара». Дж. Селл. Комп. Физиол . 22 (2): 187–96. дои : 10.1002/jcp.1030220209 .
- ^ Перейти обратно: а б Ходжкин А.Л. (1951). «Ионная основа электрической активности нервов и мышц». Биол. Преподобный . 26 (4): 339–409. дои : 10.1111/j.1469-185X.1951.tb01204.x . S2CID 86282580 .
- ^ Справочник CRC по химии и физике , 83-е издание, ISBN 0-8493-0483-0 , стр. 12–14–12–16.
- ^ Эйзенман Г (1961). «Об элементарном атомном происхождении равновесной ионной специфичности». В Кляйнцеллере; Котык (ред.). Симпозиум по мембранному транспорту и метаболизму . Нью-Йорк: Академическая пресса. стр. 163–79. Эйзенман Г (1965). «Некоторые элементарные факторы, участвующие в специфическом проникновении ионов». Учеб. 23-й Международный. Конгресс Физиол. наук, Токио . Амстердам: Excerta Med. Найденный. стр. 489–506.
* Даймонд Дж. М., Райт Э. М. (1969). «Биологические мембраны: физические основы селективности ионов и неэктролитов». Ежегодный обзор физиологии . 31 : 581–646. дои : 10.1146/annurev.ph.31.030169.003053 . ПМИД 4885777 . - ^ Мальчик, стр. 33–37.
- ^ Цай С.К., Ли В., Сести Ф (2007). «Множественные режимы регулирования калиевого тока а-типа». Курс. Фарм. Дес . 13 (31): 3178–84. дои : 10.2174/138161207782341286 . ПМИД 18045167 .
- ^ Голдин А.Л. (2007). «Нейрональные каналы и рецепторы». В Ваксмане С.Г. (ред.). Молекулярная неврология . Берлингтон, Массачусетс: Elsevier Academic Press. стр. 43–58. ISBN 978-0-12-369509-3 .
- ^ Первес и др. , стр. 28–32; Буллок , Орканд и Гриннелл, стр. 133–134; Шмидт-Нильсен, стр. 478–480, 596–597; Янг, с. 33–35
- ^ Санес, Дэн Х.; Такач, Кэтрин (1 июня 1993 г.). «Активнозависимое уточнение тормозных связей». Европейский журнал неврологии . 5 (6): 570–574. дои : 10.1111/j.1460-9568.1993.tb00522.x . ISSN 1460-9568 . ПМИД 8261131 . S2CID 30714579 .
- ^ КОФУДЗИ, П.; НЬЮМАН, Э.А. (1 января 2004 г.). «Буферизация калия в центральной нервной системе» . Нейронаука . 129 (4): 1045–1056. doi : 10.1016/j.neuroscience.2004.06.008 . ISSN 0306-4522 . ПМК 2322935 . ПМИД 15561419 .
- ^ Санес, Дэн Х.; Рех, Томас А. (1 января 2012 г.). Развитие нервной системы (Третье изд.). Эльзевир Академик Пресс. стр. 211–214. ISBN 9780080923208 . OCLC 762720374 .
- ^ Тости, Элизабетта (28 июня 2010 г.). «Динамическая роль ионных токов в раннем развитии» . Молекулярное воспроизводство и развитие . 77 (10): 856–867. дои : 10.1002/mrd.21215 . ISSN 1040-452X . ПМИД 20586098 . S2CID 38314187 .
- ^ Бойе, MR; Джуэлл, БР (1981). «Анализ влияния изменений скорости и ритма на электрическую активность сердца» . Прогресс биофизики и молекулярной биологии . 36 (1): 1–52. дои : 10.1016/0079-6107(81)90003-1 . ISSN 0079-6107 . ПМИД 7001542 .
- ^ Спинелли, Валентина; Сартиани, Лаура; Муджелли, Алессандро; Романелли, Мария Новелла; Чербай, Элизабетта (2018). «Каналы, управляемые циклическими нуклеотидами, активируемые гиперполяризацией: патофизиологические, онтогенетические и фармакологические данные об их функции в клеточной возбудимости». Канадский журнал физиологии и фармакологии . 96 (10): 977–984. дои : 10.1139/cjpp-2018-0115 . hdl : 1807/90084 . ISSN 0008-4212 . ПМИД 29969572 . S2CID 49679747 .
- ^ Джонс, Брайан Л.; Смит, Стивен М. (30 марта 2016 г.). «Кальций-чувствительный рецептор: ключевая мишень для внеклеточной передачи сигналов кальция в нейронах» . Границы в физиологии . 7 : 116. doi : 10.3389/fphys.2016.00116 . ISSN 1664-042X . ПМК 4811949 . ПМИД 27065884 .
- ^ Дебанн, Доминик; Инглеберт, Янис; Руссье, Микаэль (2019). «Пластичность внутренней возбудимости нейронов» (PDF) . Современное мнение в нейробиологии . 54 : 73–82. дои : 10.1016/j.conb.2018.09.001 . ПМИД 30243042 . S2CID 52812190 .
- ^ Давенпорт, Беннетт; Ли, Юань; Хейзер, Джастин В.; Шмитц, Карстен; Перро, Анн-Лор (23 июля 2015 г.). «Сигнатурных каналов возбудимости больше нет: каналы L-типа в иммунных клетках» . Границы в иммунологии . 6 : 375. дои : 10.3389/fimmu.2015.00375 . ISSN 1664-3224 . ПМЦ 4512153 . ПМИД 26257741 .
- ^ Сакманн, Берт (21 апреля 2017 г.). «От одиночных клеток и отдельных столбцов к корковым сетям: дендритная возбудимость, обнаружение совпадений и синаптическая передача в срезах мозга и головном мозге» . Экспериментальная физиология . 102 (5): 489–521. дои : 10.1113/ep085776 . ISSN 0958-0670 . ПМЦ 5435930 . ПМИД 28139019 .
- ^ Франко Р., Бортнер К.Д., Цидловски Дж.А. (январь 2006 г.). «Потенциальная роль электрогенного ионного транспорта и деполяризации плазматической мембраны в апоптозе» . Дж. Член. Биол . 209 (1): 43–58. дои : 10.1007/s00232-005-0837-5 . ПМИД 16685600 . S2CID 849895 .
- ^ Первес и др. , стр. 32–33; Буллок , Орканд и Гриннелл, стр. 138–140; Шмидт-Нильсен, стр. 480; Янг, с. 35–37
- ^ Спенглер С.Г. (1972). «Расширение уравнения постоянного поля, включив в него как двухвалентные, так и одновалентные ионы». Алабамский журнал медицинских наук . 9 (2): 218–23. ПМИД 5045041 .
- ^ Первес и др. , с. 34; Буллок , Орканд и Гриннелл, с. 134; Шмидт-Нильсен , стр. 478–480.
- ^ Первес и др. , стр. 33–36; Буллок , Орканд и Гриннелл, с. 131.
- ^ Перейти обратно: а б Магнусон Д.С., Морасутти DJ, Стейнс В.А., Макберни М.В., Маршалл К.С. (14 января 1995 г.). «Электрофизиологическое созревание in vivo нейронов, полученных из линии мультипотентных клеток-предшественников (эмбриональной карциномы)». Исследования развития мозга . 84 (1): 130–41. дои : 10.1016/0165-3806(94)00166-W . ПМИД 7720212 .
- ^ Юусола М., Коувалайнен Э., Ярвилехто М., Векстрем М. (сентябрь 1994 г.). «Усиление контрастности, соотношение сигнал/шум и линейность в светоадаптированных фоторецепторах мясных мух» . J Gen Physiol . 104 (3): 593–621. дои : 10.1085/jgp.104.3.593 . ПМК 2229225 . ПМИД 7807062 .
- ^ Лафлин С.Б., де Рюйтер ван Стивенинк Р.Р., Андерсон Дж.К. (май 1998 г.). «Метаболическая стоимость нейронной информации». Нат. Нейроски . 1 (1): 36–41. дои : 10.1038/236 . ПМИД 10195106 . S2CID 204995437 .
- ^ Гошал К. и др. (декабрь 2017 г.). «Диэлектрические свойства плазматической мембраны: признак дислипидемии при сахарном диабете». Арх Биохим Биофиз . 635 : 27–36. дои : 10.1016/j.abb.2017.10.002 . ПМИД 29029878 .
- ^ Уилкокс, Кристи (28 июля 2011 г.). «Мгновенный зомби — просто добавь соли» . Scientific American Сеть блогов . Проверено 30 января 2023 г.
- ^ Экипаж, Бек (3 июля 2015 г.). «Смотреть: кусок сырой говядины начинает пульсировать после того, как его принесли домой от мясника» . НаукаАлерт . Проверено 30 января 2023 г.
- ^ «女子切» . Legal.people.com.cn . Архивировано из оригинала 3 июля 2015 года . Проверено 30 января 2023 г.
- ^ Гибсон, Грей (11 января 2023 г.). «Обычная приправа заставляет свеженарезанное мясо пульсировать, что приводит в ужас Твиттер» . Ньюшуб . Проверено 30 января 2023 г.
- ^ Кост, Бен (10 января 2023 г.). «Говядина тверки: Интернет в ужасе от видео, где свежее мясо «спазмируется» » . nypost.com . Проверено 30 января 2023 г.
Дальнейшее чтение
[ редактировать ]- Альбертс и др. Молекулярная биология клетки . Издательство «Гирлянда»; 4-е издание Bk&Cdr (март 2002 г.). ISBN 0-8153-3218-1 . Уровень бакалавриата.
- Гайтон, Артур К., Джон Э. Холл. Учебник медицинской физиологии . Компания ВБ Сондерс; 10-е издание (15 августа 2000 г.). ISBN 0-7216-8677-X . Уровень бакалавриата.
- Хилле, Б. Ионный канал возбудимых мембран Sinauer Associates, Сандерленд, Массачусетс, США; 1-е издание, 1984 г. ISBN 0-87893-322-0
- Николлс Дж.Г., Мартин А.Р. и Уоллес Б.Г. От нейрона к мозгу Sinauer Associates, Inc. Сандерленд, Массачусетс, США, 3-е издание, 1992 г. ISBN 0-87893-580-0
- Ове-Стен Кнудсен. Биологические мембраны: теория транспорта, потенциалов и электрических импульсов . Издательство Кембриджского университета (26 сентября 2002 г.). ISBN 0-521-81018-3 . Выпускной уровень.
- Национальная медицинская серия для независимого изучения. Физиология . Липпинкотт Уильямс и Уилкинс. Филадельфия, Пенсильвания, США, 4-е издание, 2001 г. ISBN 0-683-30603-0
Внешние ссылки
[ редактировать ]- Функции клеточной мембраны
- Симулятор уравнений Нернста/Гольдмана
- Калькулятор уравнений Нернста
- Калькулятор уравнения Гольдмана-Ходжкина-Каца
- Калькулятор электрохимической движущей силы
- Происхождение потенциала мембран покоя - интерактивное онлайн-руководство (Flash)
| Базы данных органов управления : Национальные |
|---|