Потенциально-зависимый анионный канал
| Эукариотический Порин | |||
|---|---|---|---|
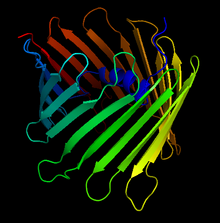 Кристаллическая структура потенциалзависимого анионного канала человека. Стрелками обозначены антипараллельные бета-листы, образующие характерный бета-бочонок. | |||
| Идентификаторы | |||
| Символ | Порин_3 | ||
| Пфам | PF01459 | ||
| ИнтерПро | ИПР001925 | ||
| PROSITE | PDOC00483 | ||
| TCDB | 1.Б.8 | ||
| Суперсемейство OPM | 189 | ||
| белок OPM | 3эмн | ||
| CDD | cd07306 | ||
| |||
Потенциал-зависимые анионные каналы , или митохондриальные порины , представляют собой класс пориновых ионных каналов, расположенных на внешней митохондриальной мембране . [1] [2] Ведутся споры относительно того, экспрессируется ли этот канал в поверхностной мембране клетки. [3] [4] [5]
Этот основной белок внешней митохондриальной мембраны эукариот образует потенциал-зависимый анион-селективный канал (VDAC), который ведет себя как общая диффузионная пора для небольших гидрофильных молекул. [6] [7] [8] [9] Канал принимает открытую конформацию при низком или нулевом мембранном потенциале и закрытую конформацию при потенциалах выше 30–40 мВ. VDAC облегчает обмен ионов и молекул между митохондриями и цитозолем и регулируется взаимодействием с другими белками и небольшими молекулами. [10]
Структура
[ редактировать ]Этот белок содержит около 280 аминокислот и образует бета-цилиндр , охватывающий внешнюю мембрану митохондрий. [11] [12]
С момента его открытия в 1976 году был проведен обширный анализ функций и структуры белков VDAC. Появилась характерная особенность пор: при восстановлении в плоские липидные бислои происходит зависимое от напряжения переключение между анион-селективным состоянием с высокой проводимостью с высоким потоком метаболитов и катион-селективным состоянием с низкой проводимостью с ограниченным прохождением метаболитов.
Спустя более чем 30 лет после его первоначального открытия, в 2008 году, были завершены три независимых структурных проекта VDAC-1. Первая была решена с помощью многомерной ЯМР-спектроскопии . Второй применил гибридный подход с использованием кристаллографических данных. Третий был для кристаллов VDAC-1 мыши, определенных методами рентгеновской кристаллографии . Три проекта 3D-структур VDAC-1 выявили множество структурных особенностей. Во-первых, VDAC-1 представляет собой новый структурный класс белков β-бочонка внешней мембраны с нечетным числом нитей. Другой аспект заключается в том, что отрицательно заряженная боковая цепь остатка Е73 ориентирована в сторону гидрофобного окружения мембраны. 19-нитевая 3D-структура, полученная в разных экспериментальных источниках в трех разных лабораториях, соответствует данным ЭМ и АСМ из нативных мембранных источников и представляет собой биологически значимое состояние VDAC-1. [10]
Механизм
[ редактировать ]При мембранном потенциале, превышающем 30 мВ (положительном или отрицательном), VDAC переходит в закрытое состояние и переходит в открытое состояние, как только напряжение падает ниже этого порога. Хотя оба состояния допускают прохождение простых солей, VDAC гораздо более строг в отношении органических анионов, категории, к которой относится большинство метаболитов . [13] Точный механизм связи изменений напряжения с конформационными изменениями внутри белка еще не разработан, но исследования Thomas et al. предполагают, что при переходе белка в закрытую форму изменения напряжения приводят к удалению большого участка белка из канала и уменьшению эффективного радиуса пор. [14] Несколько остатков лизина , а также Glu-152 считаются особенно важными сенсорными остатками внутри белка. [15]
Биологическая функция
[ редактировать ]Потенциал-зависимый ионный канал играет ключевую роль в регулировании метаболического и энергетического потока через внешнюю митохондриальную мембрану. Он участвует в транспортировке АТФ , АДФ , пирувата , малата и других метаболитов и, таким образом, активно взаимодействует с ферментами метаболических путей. [13] АТФ-зависимые цитозольные ферменты гексокиназа , глюкокиназа и глицерин-киназа , а также митохондриальный фермент креатинкиназа Было обнаружено, что связываются с VDAC. Это связывание помещает их в непосредственную близость к АТФ, высвобождаемому из митохондрий. В частности, предполагается, что связывание гексокиназы играет ключевую роль в сочетании гликолиза с окислительным фосфорилированием . [14] Кроме того, VDAC является важным регулятором Ca. 2+ транспорт в митохондрии и из митохондрий. Потому что Ка 2+ является кофактором метаболических ферментов, таких как пируватдегидрогеназа и изоцитратдегидрогеназа . На выработку энергии и гомеостаз влияет проницаемость VDAC для кальция. 2+ . [16]
Актуальность заболевания
[ редактировать ]Также было показано, что VDAC играет роль в апоптозе . [17] Во время апоптоза VDAC модифицирует переходную пору проницаемости митохондрий, вызывая высвобождение апоптогенных факторов, таких как цитохром с . Однако VDAC не являются важными компонентами переходной поры митохондриальной проницаемости. Хотя цитохром с играет важную роль в окислительном фосфорилировании внутри митохондрий. В цитозоле он активирует протеолитические ферменты, называемые каспазами , которые играют важную роль в гибели клеток. [18] Хотя механизм высвобождения цитохрома c, облегчаемого VDAC, еще полностью не выяснен, некоторые исследования показывают, что олигомеризация между отдельными субъединицами может создавать большие гибкие поры, через которые может проходить цитохром c. [19] Более важным фактором является то, что высвобождение цитохрома c также регулируется семейством белков Bcl-2 : Bax напрямую взаимодействует с VDAC, увеличивая размер пор и способствуя высвобождению цитохрома c, в то время как антиапоптотический Bcl-xL производит прямо противоположный эффект. [20] Фактически было показано, что антитела, которые ингибируют VDAC, также мешают Bax-опосредованному высвобождению цитохрома с как в изолированных митохондриях, так и в целых клетках. [21] Эта ключевая роль в апоптозе предполагает, что VDAC является потенциальной мишенью для химиотерапевтических препаратов.
Примеры
[ редактировать ]Дрожжи содержат двух членов этого семейства (гены POR1 и POR2); у позвоночных есть как минимум три члена (гены VDAC1, VDAC2 и VDAC3). [11]
Люди, как и большинство высших эукариот, кодируют три разных VDAC; VDAC1 , VDAC2 и VDAC3 . Вместе с TOMM40 и TOMM40L они представляют собой семейство эволюционно родственных β-бочек. [22]
Растения имеют наибольшее количество VDAC. Arabidopsis кодирует четыре разных VDAC, но у других видов это число может быть больше. [23]
Ссылки
[ редактировать ]- ^ Хугенбум Б.В., Суда К., Энгель А., Фотиадис Д. (2007). «Супрамолекулярные ансамбли потенциалзависимых анионных каналов в нативной мембране». Дж. Мол. Биол . 370 (2): 246–55. дои : 10.1016/j.jmb.2007.04.073 . ПМИД 17524423 .
- ^ Блакли-Дайсон, Э; Форте, М. (сентябрь 2001 г.). «Каналы VDAC» . ИУБМБ Жизнь . 52 (3–5): 113–8. дои : 10.1080/15216540152845902 . ПМИД 11798022 . S2CID 38314888 .
- ^ Сабиров Р.З., Мерзляк П.Г. (июнь 2012 г.). «Споры о плазмалемме VDAC и загадка макси-анионных каналов» . Биохим. Биофиз. Акта . 1818 (6): 1570–80. дои : 10.1016/j.bbamem.2011.09.024 . ПМИД 21986486 .
- ^ Де Пинто, В.; Мессина, А.; Лейн, DJR; Лоуэн, А. (2010). «Потенциал-зависимый анион-селективный канал (VDAC) в плазматической мембране» . Письма ФЭБС . 584 (9): 1793–1799. Бибкод : 2010FEBSL.584.1793D . дои : 10.1016/j.febslet.2010.02.049 . ПМИД 20184885 . S2CID 3391282 .
- ^ Нихаге, К.; Стинблок, К.; Пурше, Т.; Борнхойзер, М.; Корбей, Д.; Хофлак, Б. (2011). Борлонган, Цезарио V (ред.). «Протеом клеточной поверхности мезенхимальных стромальных клеток человека» . ПЛОС ОДИН . 6 (5): е20399. Бибкод : 2011PLoSO...620399N . дои : 10.1371/journal.pone.0020399 . ПМК 3102717 . ПМИД 21637820 .
- ^ Бенц Р. (1994). «Проникновение гидрофильных растворов через внешние мембраны митохондрий: обзор митохондриальных поринов». Биохим. Биофиз. Акта . 1197 (2): 167–196. дои : 10.1016/0304-4157(94)90004-3 . ПМИД 8031826 .
- ^ Маннелла, Калифорния (1992). «Входы и выходы митохондриальных мембранных каналов». Тенденции биохимии. Наука . 17 (8): 315–320. дои : 10.1016/0968-0004(92)90444-E . ПМИД 1384178 .
- ^ Диханич М (1990). «Биогенез и функция эукариотических поринов». Эксперименты . 46 (2): 146–153. дои : 10.1007/BF02027310 . ПМИД 1689252 . S2CID 2199583 .
- ^ Форте М., Гай Х.Р., Маннелла, Калифорния (1987). «Молекулярная генетика ионного канала VDAC: структурная модель и анализ последовательности» . Дж. Биоэнергетика. Биомембр . 19 (4): 341–350. дои : 10.1007/BF00768537 . ПМИД 2442148 . S2CID 10219032 .
- ^ Jump up to: а б Хиллер С., Абрамсон Дж., Маннелла С., Вагнер Г., Зет К. (сентябрь 2010 г.). «Трехмерные структуры VDAC представляют собой нативную конформацию» . Тенденции биохимии. Наука . 35 (9): 514–21. дои : 10.1016/j.tibs.2010.03.005 . ПМЦ 2933295 . ПМИД 20708406 .
- ^ Jump up to: а б Сэмпсон М.Дж., Ловелл Р.С., Дэвисон Д.Б., Крейген У.Дж. (1996). «Новый ген митохондриального потенциал-зависимого анионного канала мыши локализуется на хромосоме 8». Геномика . 36 (1): 192–196. дои : 10.1006/geno.1996.0445 . ПМИД 8812436 .
- ^ Зет К (2010). «Структура и эволюция белков внешней мембраны митохондрий топологии бета-цилиндра» . Биохим. Биофиз. Акта . 1797 (6–7): 1292–9. дои : 10.1016/j.bbabio.2010.04.019 . ПМИД 20450883 .
- ^ Jump up to: а б Блакли-Дайсон Э. и Форте М. (2001). «Каналы VDAC» . ИУБМБ Жизнь . 52 (3–5): 113–18. дои : 10.1080/15216540152845902 . ПМИД 11798022 . S2CID 38314888 .
- ^ Jump up to: а б Коломбини М., Блакли-Дайсон Э., Форте М. (1996). «VDAC, канал во внешней митохондриальной мембране». Ионные каналы . Том. 4. С. 169–202. дои : 10.1007/978-1-4899-1775-1_5 . ISBN 978-1-4899-1777-5 . ПМИД 8744209 .
- ^ Томас Л., Блакли-Дайсон Э., Коломбини М., Форте М. (июнь 1993 г.). «Картирование остатков, образующих потенциал-сенсор потенциалзависимого анион-селективного канала» . Учеб. Натл. акад. наук. США . 90 (12): 5446–9. Бибкод : 1993PNAS...90.5446T . дои : 10.1073/pnas.90.12.5446 . ПМК 46737 . ПМИД 7685903 .
- ^ Шошан-Бармац V; Гинцель Д. (2003). «Напряженно-зависимый анионный канал: характеристика, модуляция и роль в функции митохондрий в жизни и смерти клеток». Клеточная биохимия. Биофиз . 39 (3): 279–92. дои : 10.1385/CBB:39:3:279 . ПМИД 14716081 . S2CID 33628015 .
- ^ Лемастерс Джей Джей; Холмухамедов Э. (2006). «Потенциально-зависимый анионный канал (VDAC) как регулятор митохондрий — нестандартное мышление» . Биохим. Биофиз. Акта . 1762 (2): 181–90. дои : 10.1016/j.bbadis.2005.10.006 . ПМИД 16307870 .
- ^ Цудзимото Ю, Симидзу С (2002). «Анионный канал, зависимый от напряжения: важный игрок в апоптозе». Биохимия . 84 (2–3): 187–93. дои : 10.1016/S0300-9084(02)01370-6 . ПМИД 12022949 .
- ^ Залк Р; Исраэльсон А; Гарти Э.С.; Азулай-Зохар Х; Шошан-Бармац В. (2005). «Олигомерные состояния потенциал-зависимого анионного канала и высвобождение цитохрома с из митохондрий» . Биохим. Дж . 386 (1): 73–83. дои : 10.1042/BJ20041356 . ПМЦ 1134768 . ПМИД 15456403 .
- ^ Симидзу С; Нарита М; Цудзимото Ю. (1999). «Белки семейства Bcl-2 регулируют высвобождение апоптогенного цитохрома с митохондриальным каналом VDAC». Природа . 399 (6735): 483–7. Бибкод : 1999Natur.399..483S . дои : 10.1038/20959 . ПМИД 10365962 . S2CID 4423304 .
- ^ Симидзу С; Мацуока Ю; Синохара Ю; Йонеда Ю; Цудзимото Ю. (2001). «Основная роль потенциалзависимых анионных каналов в различных формах апоптоза в клетках млекопитающих» . Дж. Клеточная Биол . 152 (2): 237–50. дои : 10.1083/jcb.152.2.237 . ПМК 2199613 . ПМИД 11266442 .
- ^ Бэй, округ Колумбия, Хафез М., Янг М.Дж., Суд окружного прокурора (июнь 2012 г.). «Филогенетический и коэволюционный анализ семейства белков β-бочонка, состоящего из митохондриального порина (VDAC) и Tom40» . Биохим. Биофиз. Акта . 1818 (6): 1502–19. дои : 10.1016/j.bbamem.2011.11.027 . ПМИД 22178864 .
- ^ Омбле Ф., Краммер Э., Прево М. (июнь 2012 г.). «Завод ВДАК: факты и предположения» . Биохим. Биофиз. Акта . 1818 (6): 1486–501. дои : 10.1016/j.bbamem.2011.11.028 . ПМИД 22155681 .
Внешние ссылки
[ редактировать ]- Потенциально-зависимые + анионные + каналы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)