VDAC1
| VDAC1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | VDAC1 , PORIN, VDAC-1, потенциалзависимый анионный канал 1 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 604492 ; МГИ : 106919 ; Гомологен : 107244 ; Генные карты : VDAC1 ; OMA : VDAC1 — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||



Потенциал-зависимый анион-селективный канал 1 (VDAC-1) представляет собой белок бета-барреля , который у человека кодируется VDAC1, геном расположенным на хромосоме 5 . [4] [5] Он образует ионный канал во внешней мембране митохондрий (ОММ), а также во внешней клеточной мембране. В ОММ он позволяет АТФ диффундировать из митохондрий в цитоплазму . В клеточной мембране он участвует в регуляции объема. Во всех эукариотических клетках митохондрии ответственны за синтез АТФ среди других метаболитов, необходимых для выживания клеток. Таким образом, VDAC1 обеспечивает связь между митохондрией и клеткой, обеспечивая баланс между клеточным метаболизмом и клеточной смертью. Помимо метаболического проникновения, VDAC1 также действует как каркас для белков, таких как гексокиназа , которые, в свою очередь, могут регулировать метаболизм. [6]
Этот белок представляет собой потенциал-зависимый анионный канал и имеет высокую структурную гомологию с другими изоформами VDAC ( VDAC2 и VDAC3 ), которые участвуют в регуляции клеточного метаболизма , митохондриального апоптоза и сперматогенеза. [7] [8] [9] [10] Чрезмерная экспрессия и неправильная регуляция этой поры могут привести к апоптозу в клетке, что приводит к множеству заболеваний в организме. В частности, поскольку VDAC1 является основным каналом транспорта ионов кальция, его дисфункция связана с раком, болезнью Паркинсона (БП) и болезнью Альцгеймера . [11] [12] [13] Кроме того, недавние исследования показали, что чрезмерная экспрессия белка VDAC1 связана с диабетом 2 типа. Университет Лунда опубликовал исследование, которое продемонстрировало, что блокирование чрезмерной экспрессии VDAC1 может предотвратить распространение диабета 2 типа. [14]

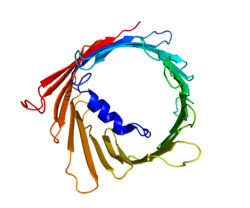
Структура
[ редактировать ]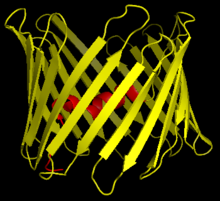
Три изоформы VDAC (VDAC1, VDAC2 и VDAC3) имеют высококонсервативные последовательности ДНК, а также трехмерные структуры, образующие широкую структуру β-цилиндра, внутри которой находится альфа-спиральный N-концевой сегмент, частично закрывающий пору. [15] Структура VDAC1 была расшифрована тремя независимыми лабораториями с помощью рентгеновской кристаллографии , спектроскопии ядерного магнитного резонанса (ЯМР) или их комбинации. Два из этих структурных исследований были использованы для определения структуры человеческого VDAC1 (hVDAC1), тогда как рентгеновская кристаллография использовалась для определения структуры мышиного VDAC1 (mVDAC1), которая отличается от hVDAC1 только двумя остатками. [16] [17] [18] Эти определенные структуры согласуются с более ранними исследованиями кругового дихроизма , которые предсказывали наличие доменов альфа-спирали и β-цепи. [19]
Структурный анализ структуры mVDAC1 показал бочкообразный канал, состоящий из 19 амфипатических β-нитей, N-конец и C-конец которого обращены к межмембранному пространству митохондрии. [20] [21] β-нити соединены петлями и расположены антипараллельно, за исключением β-цепей 1 и 19, которые параллельны. [18] Пора имеет высоту 40 Ẳ, простирается на расстояние 27 Ẳ на 20 Ẳ у отверстий и сужается до 20 Ẳ на 14 Ẳ на N-концевом сегменте α-спирали в открытом состоянии. [22] Конформация закрытого состояния еще предстоит выделить и определить. Кроме того, N-конец имеет альфа-спиральный сегмент, который удерживается на внутренней стенке поры за счет гидрофобных взаимодействий с остатками на β-листах 8–18. [18] Этот N-конец может служить основой для движения ионов или прикрепления белков. Одним из таких примеров является место стыковки для связывания HK1. [6] Важным остатком, на который следует обратить внимание, является глутамат, расположенный на 73-м остатке аминокислотной цепи (E73). Этот остаток обнаружен в VDAC1 и VDAC2, но не в VDAC3. Боковая цепь этого заряженного остатка направлена в сторону фосфолипидного бислоя , который обычно вызывает возникновение сил отталкивания. Однако E73 участвует в функции и взаимодействии VDAC1. [23]
Функция
[ редактировать ]VDAC1 принадлежит к семейству митохондриальных поринов и, как ожидается, будет иметь те же биологические функции, что и другие изоформы VDAC. [24] Из трех изоформ VDAC1 является основным каналом транспорта ионов кальция в митохондриях и наиболее широко транскрибируется. [12] [25] VDAC1 участвует в клеточном метаболизме путем транспортировки АТФ и других небольших метаболитов через внешнюю митохондриальную мембрану (ОММ), что позволяет регулировать цикл ТСА и, как следствие, выработку активных форм кислорода (АФК). [11] В дрожжевых клетках АФК накапливаются в условиях окислительного стресса, что приводит к нарушению функции митохондрий и «маленькому» фенотипу. Однако мелкие дрожжевые клетки демонстрируют более длительную продолжительность жизни, чем клетки дикого типа, и указывают на защитную функцию VDAC1 в аналогичных обстоятельствах, таких как старение. [6] [25]
стробирование напряжения
[ редактировать ]VDAC1 обеспечивает проводимость молекул в митохондрии и из них. Его проницаемость зависит от конформационного состояния VDAC1, которое определяется напряжением. При низком напряжении (10 мВ) пора находится в «открытом» состоянии, где канал слабо селективен к анионам и обеспечивает больший поток метаболитов. Из-за большого размера пор метаболические ворота в условиях насыщения АТФ обнаруживают транспорт 2 000 000 АТФ в секунду и транспорт 10 000 АТФ в физиологических условиях. [26] При более высоком напряжении в положительном или отрицательном направлении (>30 мВ) пора находится в «закрытом» состоянии и слабо селективна по катионам, что позволяет транспортировать меньше метаболитов. [18] Поток метаболитов можно рассматривать как незначительный. Это изменение состояний опосредовано конформационным изменением белка, которое еще предстоит обнаружить. Поскольку альфа-спиральный N-концевой сегмент расположен в центре поры, он идеально расположен для метаболических ворот. Это заставило исследователей поверить в то, что Альфа-спираль внесла ключевой вклад в определение конформационных состояний. Однако более поздние исследования показали, что N-конец не нужен для правильного управления напряжением и, следовательно, предполагают, что гибкий бета-цилиндр является механизмом конформационных изменений. [22]
Олигомеризация
[ редактировать ]Атомно-силовая микроскопия (АСМ) выявила наличие мономеров VDAC1, а также димеров и более крупных олигомеров, демонстрирующих взаимодействие поры с самой собой, однако димеры встречаются чаще. [27] В частности, было показано, что hVDAC1 располагается в параллельных димерах, что приводит к увеличению проницаемости пор. [16] Также было показано, что глутамат, расположенный в 73-м положении VDAC1, играет роль в олигомеризации в присутствии кальция. [23] VDAC также могут олигомеризоваться, образуя часть переходной поры митохондриальной проницаемости (MPTP) и, таким образом, облегчая высвобождение цитохрома C, что приводит к апоптозу. Также было обнаружено, что VDAC взаимодействуют с про- или антиапоптотическими белками, такими как белки и киназы семейства Bcl-2, и поэтому могут способствовать апоптозу независимо от MPTP. [24]
Клиническое значение
[ редактировать ]Все потенциалозависимые анионные каналы участвуют в транспорте ионов и метаболитов, хотя их физиологические роли различны. Из-за своей роли дисфункция каналов может привести к различным заболеваниям. VDAC1 вовлечен в развитие рака посредством его взаимодействия с антиапоптотическим семейством белков, белками Bcl-2 , особенно Bcl-xl и Mcl-1 , которые сверхэкспрессируются при раке. Эти два белка Bcl-2 взаимодействуют с VDAC1, регулируя транспорт ионов кальция через OMM и, в конечном итоге, выработку АФК. В то время как высокие уровни АФК вызывают гибель клеток, нелетальные уровни мешают путям передачи сигнала, которые затем могут способствовать пролиферации, миграции и инвазии клеток в раковые клетки. [11] Более того, сверхэкспрессия VDAC1 связана с усилением апоптотического ответа, противораковыми препаратами и эффективностью лечения, что дополнительно подтверждает, что VDAC1 является терапевтической мишенью для лечения рака. [11] [28]
Функция VDAC1 в транспорте ионов кальция также связана с нейродегенеративными заболеваниями. При БП VDAC1 повышает уровень ионов кальция в митохондриях, что приводит к увеличению проницаемости митохондрий, нарушению потенциала митохондриальных мембран, повышению продукции АФК, гибели клеток и дегенерации нейронов. [12] Было показано, что VDAC1 взаимодействует с амилоидом β (Aβ), что приводит к увеличению проводимости канала и, в конечном итоге, к апоптозу клетки. [13]
Взаимодействия
[ редактировать ]VDAC1 действует как каркас для многих белков, а также обеспечивает поток ионов и метаболитов посредством взаимодействий внутри пор.
Основным метаболитом, который движется по этому каналу, является АТФ. Сайт связывания с низким сродством, используемый для быстрого транспорта этой молекулы, был обнаружен с помощью подхода моделирования состояния Маркова . Было показано, что АТФ последовательно связывается с несколькими основными остатками внутри поры, по существу перемещаясь по каналу. [29]
Также было показано, что VDAC1 взаимодействует с:
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000020402 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Блакли-Дайсон Э., Бальдини А., Литт М., Маккейб Э.Р., Форте М. (март 1994 г.). «Гены человека, кодирующие потенциал-зависимый анионный канал (VDAC) внешней митохондриальной мембраны: картирование и идентификация двух новых изоформ». Геномика . 20 (1): 62–67. дои : 10.1006/geno.1994.1127 . ПМИД 7517385 .
- ^ «Ген Энтрез: потенциал-зависимый анионный канал VDAC1 1» .
- ^ Jump up to: а б с д Рейна С, Палермо В, Гуарнера А, Гуарино Ф, Мессина А, Маццони С, Де Пинто В (июль 2010 г.). «Замена N-конца VDAC1 на VDAC3 восстанавливает полную активность канала и придает клетке антивозрастные свойства». Письма ФЭБС . 584 (13): 2837–2844. Бибкод : 2010FEBSL.584.2837R . дои : 10.1016/j.febslet.2010.04.066 . hdl : 11573/126366 . ПМИД 20434446 . S2CID 22130291 .
- ^ Субеди К.П., Ким Дж.С., Кан М., Сон М.Дж., Ким Ю.С., Ву Ш. (февраль 2011 г.). «Зависимый от напряжения анионный канал 2 модулирует искры Ca²+ в покое, но не индуцированную потенциалом действия передачу сигналов Ca²+ в кардиомиоцитах». Клеточный кальций . 49 (2): 136–143. дои : 10.1016/j.ceca.2010.12.004 . ПМИД 21241999 .
- ^ Jump up to: а б Альвира С.М., Умеш А., Хустед С., Ин Л., Хоу Ю., Люу С.К. и др. (ноябрь 2012 г.). «Зависимое от напряжения взаимодействие анионного канала-2 с синтазой оксида азота усиливает выработку оксида азота эндотелиальными клетками легочной артерии» . Американский журнал респираторной клеточной и молекулярной биологии . 47 (5): 669–678. дои : 10.1165/rcmb.2011-0436OC . ПМК 3547107 . ПМИД 22842492 .
- ^ Ченг Э.Х., Шейко ТВ, Фишер Дж.К., Крейген В.Дж., Корсмейер С.Дж. (июль 2003 г.). «VDAC2 ингибирует активацию BAK и апоптоз митохондрий». Наука . 301 (5632): 513–517. Бибкод : 2003Sci...301..513C . дои : 10.1126/science.1083995 . ПМИД 12881569 . S2CID 37099525 .
- ^ Ли З, Ван Ю, Сюэ Ю, Ли Икс, Цао Х, Чжэн С.Дж. (февраль 2012 г.). «Критическая роль потенциал-зависимого анионного канала 2 в апоптозе, индуцированном вирусом инфекционной бурсальной болезни, в клетках-хозяевах посредством взаимодействия с VP5» . Журнал вирусологии . 86 (3): 1328–1338. дои : 10.1128/JVI.06104-11 . ПМК 3264341 . ПМИД 22114330 .
- ^ Jump up to: а б с д и Хуан Х., Шах К., Брэдбери Н.А., Ли С., Уайт С. (октябрь 2014 г.). «Mcl-1 способствует миграции клеток рака легких путем прямого взаимодействия с VDAC, увеличивая поглощение митохондриями Ca2+ и образование активных форм кислорода» . Смерть клеток и болезни . 5 (10): e1482. дои : 10.1038/cddis.2014.419 . ПМЦ 4237246 . ПМИД 25341036 .
- ^ Jump up to: а б с Чу Ю., Голдман Дж. Г., Келли Л., Хе Ю., Валичек Т., Кордауэр Дж. Х. (сентябрь 2014 г.). «Аномальный альфа-синуклеин уменьшает потенциал-зависимый анионный канал 1 ниграла при спорадической и экспериментальной болезни Паркинсона». Нейробиология болезней . 69 : 1–14. дои : 10.1016/j.nbd.2014.05.003 . ПМИД 24825319 . S2CID 22722682 .
- ^ Jump up to: а б Смилански А., Дангур Л., Накдимон И., Бен-Хайль Д., Мизрахи Д., Шошан-Бармац В. (декабрь 2015 г.). «Зависимый от напряжения анионный канал 1 опосредует токсичность β-амилоида и представляет собой потенциальную мишень для терапии болезни Альцгеймера» . Журнал биологической химии . 290 (52): 30670–30683. дои : 10.1074/jbc.M115.691493 . ПМЦ 4692199 . ПМИД 26542804 .
- ^ Чжан Э., Мохаммед Аль-Амили I, Мохаммед С., Луан С., Асплунд О., Ахмед М. и др. (январь 2019 г.). «Сохранение секреции инсулина при диабете путем ингибирования сверхэкспрессии VDAC1 и поверхностной транслокации в β-клетках» . Клеточный метаболизм . 29 (1): 64–77.e6. дои : 10.1016/j.cmet.2018.09.008 . ПМК 6331340 . ПМИД 30293774 .
- ^ Амодео Г.Ф., Скорчапино М.А., Мессина А., Де Пинто В., Чеккарелли М. (2014). «Распределение заряженных остатков модулирует селективность открытого состояния человеческих изоформ потенциалзависимого анион-селективного канала» . ПЛОС ОДИН . 9 (8): e103879. Бибкод : 2014PLoSO...9j3879A . дои : 10.1371/journal.pone.0103879 . ПМК 4146382 . ПМИД 25084457 .
- ^ Jump up to: а б Байрубер М., Майнс Т., Хабек М., Беккер С., Гиллер К., Виллингер С. и др. (октябрь 2008 г.). «Структура потенциал-зависимого анионного канала человека» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (40): 15370–15375. Бибкод : 2008PNAS..10515370B . дои : 10.1073/pnas.0808115105 . ПМК 2557026 . ПМИД 18832158 .
- ^ Хиллер С., Гарсес Р.Г., Малия Т.Дж., Орехов В.Я., Коломбини М., Вагнер Г. (август 2008 г.). «Структура раствора интегрального мембранного белка человека VDAC-1 в мицеллах детергента» . Наука . 321 (5893): 1206–1210. Бибкод : 2008Sci...321.1206H . дои : 10.1126/science.1161302 . ПМЦ 2579273 . ПМИД 18755977 .
- ^ Jump up to: а б с д Уджвал Р., Касио Д., Коллетье Дж.П., Фахам С., Чжан Дж., Торо Л. и др. (ноябрь 2008 г.). «Кристаллическая структура мышиного VDAC1 при разрешении 2,3 А раскрывает механизмы механизма пропускания метаболитов» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (46): 17742–17747. Бибкод : 2008PNAS..10517742U . дои : 10.1073/pnas.0809634105 . ПМЦ 2584669 . ПМИД 18988731 .
- ^ Шанмугавадиву Б., Апелл Х.Дж., Майнс Т., Зет К., Кляйншмидт Дж.Х. (апрель 2007 г.). «Правильное сворачивание бета-цилиндра мембранного белка человека VDAC требует наличия липидного бислоя» . Журнал молекулярной биологии . 368 (1): 66–78. дои : 10.1016/j.jmb.2007.01.066 . ПМИД 17336328 .
- ^ Маккоммис К.С., Бейнс С.П. (июнь 2012 г.). «Роль VDAC в гибели клеток: друг или враг?» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1818 (6): 1444–1450. дои : 10.1016/j.bbamem.2011.10.025 . ПМЦ 3288473 . ПМИД 22062421 .
- ^ Томаселло М.Ф., Гуарино Ф., Рейна С., Мессина А., Де Пинто В. (декабрь 2013 г.). «Топография потенциал-зависимого анион-селективного канала 1 (VDAC1) во внешней мембране митохондрий, обнаруженная в интактной клетке» . ПЛОС ОДИН . 8 (12): е81522. Бибкод : 2013PLoSO...881522T . дои : 10.1371/journal.pone.0081522 . ПМЦ 3855671 . ПМИД 24324700 .
- ^ Jump up to: а б Тейджидо О., Уджвал Р., Хиллердал К.О., Куллман Л., Ростовцева Т.К., Абрамсон Дж. (март 2012 г.). «Прикрепление N-концевой α-спирали к стенке потенциал-зависимого анионного канала не предотвращает его потенциал-регулирование» . Журнал биологической химии . 287 (14): 11437–11445. дои : 10.1074/jbc.M111.314229 . ПМЦ 3322836 . ПМИД 22275367 .
- ^ Jump up to: а б Кейнан Н., Пахима Х., Бен-Хайль Д., Шошан-Бармац В. (июль 2013 г.). «Роль кальция в олигомеризации VDAC1 и митохондриально-опосредованном апоптозе» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1833 (7): 1745–1754. дои : 10.1016/j.bbamcr.2013.03.017 . ПМИД 23542128 .
- ^ Jump up to: а б Ли MJ, Ким JY, Сук К., Пак JH (май 2004 г.). «Идентификация индуцируемого гипоксией фактора 1 альфа-чувствительного гена HGTD-P в качестве медиатора в митохондриальном апоптотическом пути» . Молекулярная и клеточная биология . 24 (9): 3918–3927. дои : 10.1128/mcb.24.9.3918-3927.2004 . ПМЦ 387743 . ПМИД 15082785 .
- ^ Jump up to: а б Де Пинто В., Гуарино Ф., Гуарнера А., Мессина А., Рейна С., Томаселло Ф.М. и др. (2010). «Характеристика изоформ VDAC человека: особая функция VDAC3?» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1797 (6–7): 1268–1275. дои : 10.1016/j.bbabio.2010.01.031 . ПМИД 20138821 .
- ^ Ростовцева Т., Коломбини М. (май 1997 г.). «Каналы VDAC опосредуют и контролируют поток АТФ: значение для регуляции функции митохондрий» . Биофизический журнал . 72 (5): 1954–1962. Бибкод : 1997BpJ....72.1954R . дои : 10.1016/s0006-3495(97)78841-6 . ПМЦ 1184392 . ПМИД 9129800 .
- ^ Хугенбум Б.В., Суда К., Энгель А., Фотиадис Д. (июль 2007 г.). «Супрамолекулярные ансамбли потенциалзависимых анионных каналов в нативной мембране». Журнал молекулярной биологии . 370 (2): 246–255. дои : 10.1016/j.jmb.2007.04.073 . ПМИД 17524423 .
- ^ Jump up to: а б Вейстхал С., Кейнан Н., Бен-Хайль Д., Ариф Т., Шошан-Бармац В. (октябрь 2014 г.). «Ca(2+)-опосредованная регуляция уровней экспрессии VDAC1 связана с индукцией гибели клеток». Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1843 (10): 2270–2281. дои : 10.1016/j.bbamcr.2014.03.021 . ПМИД 24704533 .
- ^ Чоудхари О.П., Пас А., Адельман Дж.Л., Коллетье Дж.П., Абрамсон Дж., Грабе М. (июль 2014 г.). «Структурное моделирование проливает свет на механизм транспорта АТФ через VDAC1» . Структурная и молекулярная биология природы . 21 (7): 626–632. дои : 10.1038/nsmb.2841 . ПМЦ 4157756 . ПМИД 24908397 .
- ^ Jump up to: а б Вэн С., Ли Ю, Сюй Д, Ши Ю, Тан Х (март 2005 г.). «Специфическое расщепление Mcl-1 каспазой-3 в апоптозе, индуцированном лигандом, индуцирующим апоптоз, связанным с фактором некроза опухоли (TRAIL), в Т-клетках лейкоза Jurkat» . Журнал биологической химии . 280 (11): 10491–10500. дои : 10.1074/jbc.M412819200 . ПМИД 15637055 .
- ^ Jump up to: а б Ши Ю, Чен Дж, Вэн С, Чен Р, Чжэн Ю, Чен Ц, Тан Х (июнь 2003 г.). «Идентификация места белок-белкового контакта и способа взаимодействия VDAC1 человека с белками семейства Bcl-2». Связь с биохимическими и биофизическими исследованиями . 305 (4): 989–996. дои : 10.1016/s0006-291x(03)00871-4 . ПМИД 12767928 .
- ^ Симидзу С., Кониси А., Кодама Т., Цудзимото Ю. (март 2000 г.). «Домен BH4 антиапоптотических членов семейства Bcl-2 закрывает потенциал-зависимый анионный канал и ингибирует апоптотические митохондриальные изменения и гибель клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (7): 3100–3105. Бибкод : 2000PNAS...97.3100S . дои : 10.1073/pnas.97.7.3100 . ПМК 16199 . ПМИД 10737788 .
- ^ Симидзу С., Нарита М., Цудзимото Ю. (июнь 1999 г.). «Белки семейства Bcl-2 регулируют высвобождение апоптогенного цитохрома с митохондриальным каналом VDAC». Природа . 399 (6735): 483–487. Бибкод : 1999Natur.399..483S . дои : 10.1038/20959 . ПМИД 10365962 . S2CID 4423304 .
- ^ Шварцер К., Барникол-Ватанабе С., Тиннес Ф.П., Хильшманн Н. (сентябрь 2002 г.). «Вольтаж-зависимый анион-селективный канал (VDAC) взаимодействует с легкой цепью динеина Tctex1 и белком теплового шока PBP74». Международный журнал биохимии и клеточной биологии . 34 (9): 1059–1070. дои : 10.1016/s1357-2725(02)00026-2 . ПМИД 12009301 . S2CID 11632830 .
- ^ Кусано Х., Симидзу С., Коя Р.К., Фудзита Х., Камада С., Кузумаки Н., Цудзимото Ю. (октябрь 2000 г.). «Человеческий гельсолин предотвращает апоптоз, ингибируя апоптотические изменения митохондрий посредством закрытия VDAC». Онкоген . 19 (42): 4807–4814. дои : 10.1038/sj.onc.1203868 . ПМИД 11039896 . S2CID 22482896 .
- ^ Бейнс С.П., Сонг С.Х., Чжэн Ю.Т., Ван Г.В., Чжан Дж., Ван О.Л. и др. (май 2003 г.). «Протеинкиназа Цепсилон взаимодействует с порами перехода проницаемости в сердечных митохондриях и ингибирует их» . Исследование кровообращения . 92 (8): 873–880. дои : 10.1161/01.RES.0000069215.36389.8D . ПМЦ 3691672 . ПМИД 12663490 .
- ^ Сунь Ю., Вашишт А.А., Чиу Дж., Вольшлегель Дж.А., Драйер Л. (ноябрь 2012 г.). «Вольтаж-зависимые анионные каналы (VDAC) привлекают Паркин к дефектным митохондриям, чтобы способствовать митохондриальной аутофагии» . Журнал биологической химии . 287 (48): 40652–40660. дои : 10.1074/jbc.M112.419721 . ПМЦ 3504778 . ПМИД 23060438 .
- ^ Носков С.Ю., Ростовцева Т.К., Безруков С.М. (декабрь 2013). «Транспорт АТФ через VDAC и комплекс VDAC-тубулин, исследованный с помощью равновесного и неравновесного моделирования МД» . Биохимия . 52 (51): 9246–9256. дои : 10.1021/bi4011495 . ПМЦ 7259721 . ПМИД 24245503 .
- ^ Мартель С., Аллуш М., Эспости Д.Д., Фанелли Е., Бурсье С., Генри С. и др. (январь 2013 г.). «Гликогенсинтаза-киназа 3-опосредованное потенциалзависимое фосфорилирование анионных каналов контролирует проницаемость внешней митохондриальной мембраны во время накопления липидов» . Гепатология . 57 (1): 93–102. дои : 10.1002/hep.25967 . ПМИД 22814966 . S2CID 21809602 .
Дальнейшее чтение
[ редактировать ]- Верье Ф., Миньотт Б., Ян Г., Бреннер С. (декабрь 2003 г.). «Исследование состава PTPC во время апоптоза для идентификации вирусного белка-мишени» (PDF) . Анналы Нью-Йоркской академии наук . 1010 (1): 126–142. Бибкод : 2003NYASA1010..126V . дои : 10.1196/анналы.1299.022 . ПМИД 15033708 . S2CID 22660199 .
- МакЭнери М.В., Снеговик А.М., Трифилетти Р.Р., Снайдер Ш. (апрель 1992 г.). «Выделение митохондриального бензодиазепинового рецептора: ассоциация с потенциал-зависимым анионным каналом и переносчиком аденинового нуклеотида» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (8): 3170–3174. Бибкод : 1992PNAS...89.3170M . дои : 10.1073/pnas.89.8.3170 . ПМК 48827 . ПМИД 1373486 .
- Доусон С.Дж., Уайт, Лос-Анджелес (май 1992 г.). «Лечение эндокардита, вызванного Haemophilus aphrophilus, ципрофлоксацином». Журнал инфекции . 24 (3): 317–320. дои : 10.1016/S0163-4453(05)80037-4 . ПМИД 1602151 .
- Юргенс Л., Ильсеманн П., Крацин Х.Д., Гессен Д., Эккарт К., Тиннес Ф.П., Хильшманн Н. (июль 1991 г.). «Исследование порина человека. IV. Первичные структуры «Порина 31НМ», выделенного из мембран скелетных мышц человека, и «Порина 31HL», полученного из мембран В-лимфоцитов человека, идентичны». Биологическая химия Хоппе-Сейлера . 372 (7): 455–463. дои : 10.1515/bchm3.1991.372.2.455 . ПМИД 1657034 .
- Кайзер Х., Крацин Х.Д., Тиннес Ф.П., Гётц Х., Шмидт В.Е., Эккарт К., Хильшманн Н. (декабрь 1989 г.). «[Идентификация поринов человека. II. Характеристика и первичная структура порина 31-lDa из B-лимфоцитов человека (Porin 31HL)]». Биологическая химия Хоппе-Сейлера . 370 (12): 1265–1278. ПМИД 2559745 .
- Брдичка Д., Калдис П., Валлиманн Т. (ноябрь 1994 г.). «Образование комплекса in vitro между октамером митохондриальной креатинкиназы и порином» . Журнал биологической химии . 269 (44): 27640–27644. дои : 10.1016/S0021-9258(18)47033-0 . ПМИД 7525559 .
- Ю В.Г., Вольфганг В., Форте М. (июнь 1995 г.). «Субклеточная локализация изоформ потенциал-зависимых анионных каналов человека» . Журнал биологической химии . 270 (23): 13998–14006. дои : 10.1074/jbc.270.23.13998 . ПМИД 7539795 .
- Томас Л., Блакли-Дайсон Э., Коломбини М., Форте М. (июнь 1993 г.). «Картирование остатков, образующих потенциал-сенсор потенциалзависимого анион-селективного канала» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (12): 5446–5449. Бибкод : 1993PNAS...90.5446T . дои : 10.1073/pnas.90.12.5446 . ПМК 46737 . ПМИД 7685903 .
- Блахли-Дайсон Э., Замбронич Э.Б., Ю.Х., Адамс В., Маккейб Э.Р., Адельман Дж. и др. (январь 1993 г.). «Клонирование и функциональная экспрессия в дрожжах двух человеческих изоформ внешнего мембранного канала митохондрий, потенциалзависимого анионного канала» . Журнал биологической химии . 268 (3): 1835–1841. дои : 10.1016/S0021-9258(18)53930-2 . ПМИД 8420959 .
- Шляйфф Э., Шор Г.К., Гопинг И.С. (март 1997 г.). «Человеческий митохондриальный рецептор импорта, Tom20p. Использование глутатиона для выявления специфических взаимодействий между Tom20-глутатион S-трансферазой и митохондриальными белками-предшественниками» . Письма ФЭБС . 404 (2–3): 314–318. Бибкод : 1997FEBSL.404..314S . дои : 10.1016/S0014-5793(97)00145-2 . ПМИД 9119086 . S2CID 29177508 .
- Рейманн С., Хаасе В., Крик В., Буркхардт Г., Тиннес Ф.П. (август 1998 г.). «Эндосомы: еще одно внемитохондриальное расположение пориновых / зависимых от напряжения анион-селективных каналов 1-го типа». Архив Пфлюгерса . 436 (3): 478–480. дои : 10.1007/s004240050659 . ПМИД 9644232 . S2CID 670898 .
- Нарита М., Симидзу С., Ито Т., Читтенден Т., Лутц Р.Дж., Мацуда Х., Цудзимото Ю. (декабрь 1998 г.). «Bax взаимодействует с порой перехода проницаемости, вызывая переход проницаемости и высвобождение цитохрома с в изолированных митохондриях» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (25): 14681–14686. Бибкод : 1998PNAS...9514681N . дои : 10.1073/pnas.95.25.14681 . ПМК 24509 . ПМИД 9843949 .
- Кромптон М., Вирджи С., Уорд Дж. М. (декабрь 1998 г.). «Циклофилин-D прочно связывается с комплексами потенциал-зависимого анионного канала и адениннуклеотид-транслоказы, образуя пору перехода проницаемости» . Европейский журнал биохимии . 258 (2): 729–735. дои : 10.1046/j.1432-1327.1998.2580729.x . ПМИД 9874241 .
- Мессина А., Олива М., Розато С., Хейзинг М., Руитенбек В., ван ден Хеувел Л.П. и др. (февраль 1999 г.). «Пересмотр изоформ 1 и 2 потенциал-зависимых анионных каналов человека». Связь с биохимическими и биофизическими исследованиями . 255 (3): 707–710. дои : 10.1006/bbrc.1998.0136 . ПМИД 10049775 .
- Симидзу С., Нарита М., Цудзимото Ю. (июнь 1999 г.). «Белки семейства Bcl-2 регулируют высвобождение апоптогенного цитохрома с митохондриальным каналом VDAC». Природа . 399 (6735): 483–487. Бибкод : 1999Natur.399..483S . дои : 10.1038/20959 . ПМИД 10365962 . S2CID 4423304 .
- Декер В.К., Боулз К.Р., Шатте Э.К., Таубин Дж.А., Крейген В.Дж. (октябрь 1999 г.). «Пересмотренное точное картирование локусов потенциал-зависимых анионных каналов человека с помощью радиационного гибридного анализа». Геном млекопитающих . 10 (10): 1041–1042. дои : 10.1007/s003359901158 . ПМИД 10501981 . S2CID 27663120 .
- Жакото Э., Равагнан Л., Леффлер М., Ферри К.Ф., Виейра Х.Л., Замзами Н. и др. (январь 2000 г.). «Вирусный белок R ВИЧ-1 индуцирует апоптоз посредством прямого воздействия на переходную пору проницаемости митохондрий» . Журнал экспериментальной медицины . 191 (1): 33–46. дои : 10.1084/jem.191.1.33 . ПМК 2195797 . ПМИД 10620603 .
- Симидзу С., Цудзимото Ю. (январь 2000 г.). «Проапоптотические члены семейства Bcl-2, содержащие только BH3, индуцируют высвобождение цитохрома c, но не потерю потенциала митохондриальной мембраны, и не модулируют напрямую активность потенциал-зависимых анионных каналов» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (2): 577–582. Бибкод : 2000PNAS...97..577S . дои : 10.1073/pnas.97.2.577 . ПМК 15372 . ПМИД 10639121 .
- Штадтмюллер Ю., Эбен-Бруннен Дж., Шмид А., Гессен Д., Клеберт С., Крацин Х.Д. и др. (декабрь 1999 г.). «Порины человека 1-го типа, полученные из митохондрий и внемитохондриальные, идентичны, как выяснилось с помощью секвенирования аминокислот и электрофизиологических характеристик». Биологическая химия . 380 (12): 1461–1466. дои : 10.1515/BC.1999.189 . ПМИД 10661876 . S2CID 1566781 .
Внешние ссылки
[ редактировать ]- VDAC1+белок,+человек Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : P21796 (белок 1 вольт-зависимого анион-селективного канала) на PDBe-KB .