Калиевый канал внутреннего выпрямителя
| Входной калиевый канал выпрямителя | |||
|---|---|---|---|
 кристаллическая структура внутреннего калиевого канала выпрямителя | |||
| Идентификаторы | |||
| Символ | ИРК | ||
| Пфам | PF01007 | ||
| Пфам Клан | CL0030 | ||
| ИнтерПро | ИПР013521 | ||
| СКОП2 | 1н9п / СКОПе / СУПФАМ | ||
| TCDB | 1.А.2 | ||
| Суперсемейство OPM | 8 | ||
| белок OPM | 3САУ | ||
| |||
Калиевые каналы с внутренним выпрямлением ( Kir ) представляют собой , IRK специфическую липидзависимую подгруппу калиевых каналов . На сегодняшний день в различных типах клеток млекопитающих идентифицировано семь подсемейств. [1] растения, [2] и бактерии. [3] Они активируются фосфатидилинозитол-4,5-бисфосфатом ( PIP 2 ). Нарушение работы каналов связано с несколькими заболеваниями. [4] [5] Каналы IRK обладают поровым доменом, гомологичным потенциал -управляемым ионным каналам , и фланкирующими трансмембранными сегментами (TMS). Они могут существовать в мембране в виде гомо- или гетероолигомеров , и каждый мономер содержит от 2 до 4 ТМС. С точки зрения функции эти белки транспортируют калий (K + ) , с большей тенденцией к K + поглощение, чем К + экспорт. [3] Процесс внутренней выпрямления был открыт Денисом Ноблом в клетках сердечной мышцы в 1960-х годах. [6] и Ричардом Адрианом и Аланом Ходжкиным в 1970 году в клетках скелетных мышц. [7]
Обзор внутреннего исправления
[ редактировать ]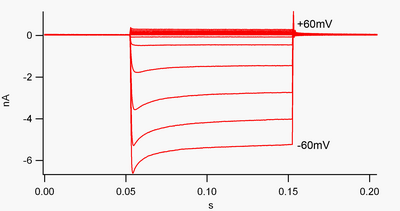
Канал, который является «внутренним выпрямлением», — это канал, который легче пропускает ток (положительный заряд) во внутреннем направлении (внутрь ячейки), чем во внешнем направлении (из ячейки). Считается, что этот ток может играть важную роль в регуляции активности нейронов, помогая стабилизировать мембранный потенциал покоя клетки.
По соглашению, входящий ток (положительный заряд, перемещающийся в ячейку) отображается в зажиме напряжения как отклонение вниз, тогда как внешний ток (положительный заряд, перемещающийся из ячейки) отображается как отклонение вверх. калия При мембранном потенциале, отрицательном по отношению к обратному потенциалу , внутрь выпрямляется К. + каналы поддерживают поток положительно заряженного K + ионы проникают в клетку, возвращая мембранный потенциал к потенциалу покоя. Это можно увидеть на рисунке 1: когда мембранный потенциал оказывается отрицательным по отношению к потенциалу покоя канала (например, -60 мВ), внутрь течет ток (т. е. в клетку течет положительный заряд). Однако, когда мембранный потенциал становится положительным по отношению к потенциалу покоя канала (например, +60 мВ), эти каналы пропускают очень небольшой ток. Проще говоря, этот канал пропускает гораздо больший ток во внутрь, чем наружу, в своем диапазоне рабочего напряжения. Эти каналы не являются идеальными выпрямителями, поскольку они могут пропускать некоторый внешний ток в диапазоне напряжений примерно на 30 мВ выше потенциала покоя.
Эти каналы отличаются от калиевых каналов, которые обычно отвечают за реполяризацию клетки после потенциала действия , таких как замедленное выпрямление и калиевые каналы А-типа . Эти более «типичные» калиевые каналы преимущественно проводят калиевые токи наружу (а не внутрь) при деполяризованных мембранных потенциалах, и их можно рассматривать как «внешне выпрямляющие». При первом обнаружении внутреннее выпрямление было названо «аномальным выпрямлением», чтобы отличить его от внешних калиевых токов. [8]
Внутренние выпрямители также отличаются от калиевых каналов с тандемными порами , которые в значительной степени ответственны за «утечку» K. + токи. [9] Некоторые внутренние выпрямители, называемые «слабыми внутренними выпрямителями», несут измеримый выходной K. + токи при напряжении, положительном к K + реверсивный потенциал (соответствует, но превышает малые токи выше линии 0 нА на рисунке 1). Они вместе с каналами «утечки» устанавливают мембранный потенциал покоя клетки. Другие каналы внутреннего выпрямления, называемые «сильными внутренними выпрямителями», вообще пропускают очень небольшой внешний ток и в основном активны при напряжениях, отрицательных по отношению к K. + реверсивный потенциал, при котором они переносят внутрь ток (намного большие токи ниже линии 0 нА на рисунке 1). [10]
Механизм внутреннего исправления
[ редактировать ]Феномен внутреннего выпрямления Kir - каналов является результатом блокады с высоким сродством эндогенных полиаминов , а именно спермина , а также ионов магния , которые закупоривают поры каналов при положительных потенциалах, что приводит к уменьшению внешних токов. Эта зависимая от напряжения блокировка полиаминов приводит к эффективной проводимости тока только во внутреннем направлении. Хотя основная идея блокады полиаминов понятна, конкретные механизмы все еще остаются спорными. [11]
Активация по PIP 2
[ редактировать ]K ir Все каналы требуют для активации фосфатидилинозитол-4,5-бисфосфата (PIP 2 ). [12] PIP 2 связывается с K ir 2.2 и напрямую активирует его, обладая свойствами агониста. [13] В этом отношении Kir - каналы представляют собой PIP2 ионные каналы, управляемые лигандом .
Роль
[ редактировать ]Kir - каналы обнаружены во многих типах клеток, включая макрофаги , сердца и почек клетки , лейкоциты , нейроны и эндотелиальные клетки . Опосредуя небольшой деполяризующий K + ток при отрицательных мембранных потенциалах, они помогают установить мембранный потенциал покоя, а в случае группы K ir 3 они помогают опосредовать тормозные реакции нейротрансмиттеров , но их роль в клеточной физиологии варьируется в зависимости от типа клеток:
| Расположение | Функция |
|---|---|
| кардиомиоциты | Kir - каналы закрываются при деполяризации, замедляя реполяризацию мембраны и помогая поддерживать более продолжительный потенциал сердечного действия . Этот тип канала внутреннего выпрямителя отличается от выпрямителя с задержкой K. + каналы , которые помогают реполяризовать нервные и мышечные клетки после потенциалов действия ; и каналы утечки калия , которые обеспечивают большую часть основы мембранного потенциала покоя . |
| эндотелиальные клетки | Kir - каналы участвуют в регуляции синтазы оксида азота . |
| почки | K или экспортирует избыток калия в собирательные трубочки для удаления с мочой или, альтернативно, может участвовать в обратном захвате калия обратно в организм. |
| нейроны и клетки сердца | IRK, активированные G-белком (K ir 3), являются важными регуляторами, модулируемыми нейротрансмиттерами. Мутация в канале GIRK2 приводит к мутации мыши-ткача. Мыши-мутанты «Weaver» являются атаксическими и демонстрируют опосредованную нейровоспалением дегенерацию дофаминергических нейронов. [14] По сравнению с неатаксическими контрольными мутантами Уивера наблюдаются нарушения координации движений и изменения в региональном метаболизме мозга. [15] Мышей Weaver уже более 30 лет исследуют в лабораториях, занимающихся развитием нервной системы и болезнями. |
| поджелудочной железы бета-клетки | K- АТФ- каналы (состоящие из Kir 6.2 субъединиц и SUR1 ) контролируют высвобождение инсулина. |
Регулирование
[ редактировать ]Зависимость напряжения может регулироваться внешним K + , внутренним Mg 2+ , внутренним АТФ и/или G-белками . P-домены каналов IRK демонстрируют ограниченное сходство последовательностей с доменами семейства VIC. Внутренние выпрямители играют роль в установлении потенциалов клеточной мембраны, а закрытие этих каналов при деполяризации позволяет возникать потенциалы действия большой продолжительности с фазой плато. Во внутренних выпрямителях отсутствуют внутренние спирали, чувствительные к напряжению, которые есть во многих каналах семейства VIC. В некоторых случаях, например, для Kir1.1a, Kir6.1 и Kir6.2, было высказано предположение, что прямое взаимодействие с членом суперсемейства ABC придает гетеромерному комплексу уникальные функциональные и регуляторные свойства, включая чувствительность к АТФ. . Эти АТФ-чувствительные каналы обнаружены во многих тканях организма. Они обеспечивают реакцию активности канала на цитоплазматическое соотношение АТФ/АДФ (повышение уровня АТФ/АДФ закрывает канал). Человеческие рецепторы сульфонилмочевины SUR1 и SUR2 (spQ09428 и Q15527 соответственно) представляют собой белки ABC, которые регулируют каналы Kir6.1 и Kir6.2 в ответ на АТФ и CFTR ( ТК №3.А.1.208.4 ) может регулировать Кир1.1а. [16]
Структура
[ редактировать ]Кристаллическая структура [17] и функция [18] бактерий семейства IRK-C. KirBac1.1 из Burkholderia pseudomallei имеет длину 333 аминокислотных остатка (aas) с двумя N-концевыми TMS, фланкирующими P-петлю (остатки 1–150), а C-концевая половина белка является гидрофильной. Он переносит одновалентные катионы с селективностью: K ≈ Rb ≈ Cs ≫ Li ≈ Na ≈ NMGM (протонированный N -метил- D -глюкамин ). Активность ингибируется Ba 2+ , Как 2+ и низкий уровень pH. [18]
Классификация
[ редактировать ]Существует семь подсемейств каналов Kir , обозначенных как Kir 1 – Kir 7 . [1] Каждое подсемейство имеет несколько членов (т.е. Kir 2.1 , Kir 2.2 , Kir 2.3 и т. д.), которые имеют почти идентичные аминокислотные последовательности у известных видов млекопитающих.
Kir - каналы образуются из гомотетрамерных мембранных белков. Каждая из четырех идентичных белковых субъединиц состоит из двух трансмембранных альфа-спиралей (М1 и М2). Гететеротетрамеры могут образовываться между членами одного и того же подсемейства (т.е. K ir 2.1 и K ir 2.3), когда каналы сверхэкспрессируются.
Разнообразие
[ редактировать ]| Ген | Белок | Псевдонимы | Связанные субъединицы |
|---|---|---|---|
| KCNJ1 | Кир 1.1 | РОМК1 | НХЕРФ2 |
| KCNJ2 | Глава 2.1 | ИРК1 | Кир 2.2 , Кир 4.1 , ПСД-95 , САП97 , АКАП79 |
| KCNJ12 | Глава 2.2 | ИРК2 | K ir 2.1 и K ir 2.3 образуют гетеромерный канал, вспомогательную субъединицу: SAP97, Вели-1 , Вели-3 , PSD-95. |
| KCNJ4 | Глава 2.3 | ИРК3 | K ir 2.1 и K ir 2.3 образуют гетеромерный канал, PSD-95, Чапсин-110 /PSD-93 |
| KCNJ14 | Глава 2.4 | IRK4 | K ir 2.1 с образованием гетеромерного канала |
| KCNJ3 | Статья 3.1 | ГИРК1, КГА | К ir 3.2, K ir 3.4, K ir 3.5, K ir 3.1 сам по себе не функционален |
| KCNJ6 | Статья 3.2 | ГИРК2 | K ir 3.1, K ir 3.3, K ir 3.4 с образованием гетеромерного канала |
| KCNJ9 | Статья 3.3 | ГРЕЧЕСКИЙ3 | K ir 3.1, K ir 3.2 с образованием гетеромерного канала |
| KCNJ5 | Статья 3.4 | ГИРК4 | Глава 3.1 , Глава 3.2 , Глава 3.3 |
| KCNJ10 | К ир 4.1 | К ир 1,2 | K ir 4.2, K ir 5.1 и K ir 2.1 с образованием гетеромерных каналов. |
| KCNJ15 | К ир 4.2 | К ир 1,3 | |
| KCNJ16 | Глава 5.1 | ОДИН 9 | |
| KCNJ8 | Статья 6.1 | К АТФ | СУР2Б |
| KCNJ11 | Статья 6.2 | К АТФ | SUR1 , SUR2A и SUR2B |
| KCNJ13 | грязь 7.1 | К ир 1,4 |
Заболевания, связанные с Кир - каналами
[ редактировать ]- Стойкая гиперинсулинемическая гипогликемия младенчества связана с аутосомно-рецессивными мутациями K ir 6.2. Определенные мутации этого гена снижают способность канала регулировать секрецию инсулина, что приводит к гипогликемии .
- Синдром Барттера может быть вызван мутациями Kir - каналов. Это состояние характеризуется неспособностью почек перерабатывать калий, что приводит к низкому уровню калия в организме.
- Синдром Андерсена — редкое заболевание, вызванное множественными мутациями K ir 2.1. В зависимости от мутации она может быть доминантной или рецессивной. Характеризуется периодическими параличами , нарушениями сердечного ритма и дисморфическими особенностями. (См. также KCNJ2 )
- Отравление барием, вероятно, связано с его способностью блокировать Kir - каналы.
- Атеросклероз (болезнь сердца) может быть связан с Kir - каналами. Потеря K ir токов в эндотелиальных клетках является одним из первых известных индикаторов атерогенеза (начала заболеваний сердца).
- Тиреотоксический гипокалиемический периодический паралич связан с изменением функции K ir 2.6. [19]
- Синдром EAST/SeSAME вызван мутациями KCNJ10. [20]
См. также
[ редактировать ]- Связанный с G-белком внутренний выпрямляющий калиевый канал
- хЭРГ
- База данных классификации транспортеров
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Кубо Ю., Адельман Дж.П., Клэпхэм Д.Э., Ян Л.И., Каршин А., Курачи Ю. и др. (декабрь 2005 г.). «Международный союз фармакологии. LIV. Номенклатура и молекулярные связи внутренних выпрямляющих калиевых каналов». Фармакологические обзоры . 57 (4): 509–26. дои : 10.1124/пр.57.4.11 . ПМИД 16382105 . S2CID 11588492 .
- ^ Хедрих Р., Моран О., Конти Ф., Буш Х., Беккер Д., Гамбале Ф. и др. (1995). «Калиевые каналы внутреннего выпрямителя у растений отличаются от своих аналогов у животных реакцией на напряжение и модуляторы каналов». Европейский биофизический журнал . 24 (2): 107–15. дои : 10.1007/BF00211406 . ПМИД 8582318 . S2CID 12718513 .
- ^ Перейти обратно: а б «1.A.2 Семейство K-каналов внутреннего выпрямителя (IRK-C)» . TCDB . Проверено 9 апреля 2016 г.
- ^ Хансен С.Б. (май 2015 г.). «Липидный агонизм: парадигма PIP2 лиганд-управляемых ионных каналов» . Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1851 (5): 620–8. дои : 10.1016/j.bbalip.2015.01.011 . ПМК 4540326 . ПМИД 25633344 .
- ^ Авраам М.Р., Джахангир А., Алексеев А.Е., Терзич А. (ноябрь 1999 г.). «Каналопатии внутренних выпрямляющих калиевых каналов» . Журнал ФАСЭБ . 13 (14): 1901–10. дои : 10.1096/fasebj.14.13.1901 . ПМИД 10544173 . S2CID 22205168 .
- ^ Ноубл, Денис (декабрь 1965 г.). «Электрические свойства сердечной мышцы, обусловленные внутренним (аномальным) выпрямлением» . Журнал клеточной и сравнительной физиологии . 66 (С2): 127–135. дои : 10.1002/jcp.1030660520 . ISSN 0095-9898 .
- ^ Адриан Р.Х., Чендлер В.К., Ходжкин А.Л. (июль 1970 г.). «Медленные изменения проницаемости калия в скелетных мышцах» . Журнал физиологии . 208 (3): 645–68. doi : 10.1113/jphysicalol.1970.sp009140 . ПМЦ 1348790 . ПМИД 5499788 .
- ^ Бертиль Хилле (2001). Ионные каналы возбудимых мембран . 3-е изд. (Зинауер: Сандерленд, Массачусетс), с. 151. ISBN 0-87893-321-2 .
- ^ Хилле, стр. 155.
- ^ Хилле, стр. 153.
- ^ Лопатин А.Н., Махина Е.Н., Николс К.Г. (ноябрь 1995 г.). «Механизм внутренней выпрямления калиевых каналов: «закупорка длинных пор» цитоплазматическими полиаминами» . Журнал общей физиологии . 106 (5): 923–55. дои : 10.1085/jgp.106.5.923 . ПМК 2229292 . ПМИД 8648298 .
- ^ Такер С.Дж., Баукровиц Т. (май 2008 г.). «Как сильно заряженные анионные липиды связывают и регулируют ионные каналы» . Журнал общей физиологии . 131 (5): 431–8. дои : 10.1085/jgp.200709936 . ПМК 2346576 . ПМИД 18411329 .
- ^ Хансен С.Б., Тао X, Маккиннон Р. (август 2011 г.). «Структурная основа активации PIP2 классического внутреннего выпрямителя K+ канала Kir2.2» . Природа . 477 (7365): 495–8. Бибкод : 2011Natur.477..495H . дои : 10.1038/nature10370 . ПМК 3324908 . ПМИД 21874019 .
- ^ Пэн Дж., Се Л., Стивенсон Ф.Ф., Мелов С., Ди Монте Д.А., Андерсен Дж.К. (ноябрь 2006 г.). «Нигростриарная дофаминергическая нейродегенерация у мышей-ткачей опосредуется нейровоспалением и облегчается введением миноциклина» . Журнал неврологии . 26 (45): 11644–51. doi : 10.1523/JNEUROSCI.3447-06.2006 . ПМЦ 6674792 . ПМИД 17093086 .
- ^ Стразиэль С., Дайс В., Нодон Л., Райсман-Возари Р., Лалонд Р. (октябрь 2006 г.). «Региональные изменения в мозге активности цитохромоксидазы и координации движений у мышей-мутантов Girk2 (Wv) (Weaver)». Нейронаука . 142 (2): 437–49. doi : 10.1016/j.neuroscience.2006.06.011 . ПМИД 16844307 . S2CID 33064439 .
- ^ Заявка WO 0190360 , Wei MH, Chaturvedi K, Guegler K, Webster M, Ketchum KA, Di Francesco V, Beasley E, «Изолированные человеческие белки-переносчики, молекулы нуклеиновой кислоты, кодирующие человеческие белки-переносчики, и их применение», опубликованная 29 ноября 2001 г., передан корпорации Apperla
- ^ Куо А., Гулбис Дж.М., Антклифф Дж.Ф., Рахман Т., Лоу Э.Д., Циммер Дж. и др. (июнь 2003 г.). «Кристаллическая структура калиевого канала KirBac1.1 в закрытом состоянии» . Наука . 300 (5627): 1922–6. Бибкод : 2003Sci...300.1922K . дои : 10.1126/science.1085028 . ПМИД 12738871 . S2CID 2703162 .
- ^ Перейти обратно: а б Энкветчакул Д., Бхаттачария Дж., Желязкова И., Гросбек Д.К., Кукрас К.А., Николс К.Г. (ноябрь 2004 г.). «Функциональная характеристика прокариотического Кир-канала» . Журнал биологической химии . 279 (45): 47076–80. дои : 10.1074/jbc.C400417200 . ПМЦ 8629170 . ПМИД 15448150 .
- ^ Райан Д.П., да Силва М.Р., Сунг Т.В., Фонтейн Б., Дональдсон М.Р., Кунг А.В. и др. (январь 2010 г.). «Мутации в калиевом канале Kir2.6 вызывают предрасположенность к тиреотоксическому гипокалиемическому периодическому параличу» . Клетка . 140 (1): 88–98. дои : 10.1016/j.cell.2009.12.024 . ПМЦ 2885139 . ПМИД 20074522 .
- ^ Бокенхауэр Д., Физер С., Станеску Х.К., Бандулик С., Здебик А.А., Райхольд М. и др. (май 2009 г.). «Эпилепсия, атаксия, нейросенсорная глухота, тубулопатия и мутации KCNJ10» . Медицинский журнал Новой Англии . 360 (19): 1960–70. дои : 10.1056/NEJMoa0810276 . ПМЦ 3398803 . ПМИД 19420365 .
Дальнейшее чтение
[ редактировать ]- Хилле Б (2001). Ионные каналы возбудимых мембран (3-е изд.). Сандерленд, Массачусетс: Синауэр. стр. 149–154. ISBN 0-87893-321-2 .
Внешние ссылки
[ редактировать ]- Внутренние + выпрямители + калий + каналы в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH).
- «Внутреннее восстановление калиевых каналов» . База данных IUPHAR по рецепторам и ионным каналам . Международный союз фундаментальной и клинической фармакологии.
- UMich Ориентация белков в мембранах семейства/семейства-85 - Пространственное положение внутренних выпрямительных калиевых каналов в мембранах.