Ионный канал, чувствительный к кислоте
| Кислоточувствительный натриевый канал | |||
|---|---|---|---|
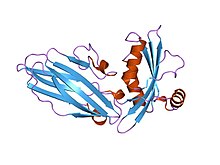 Структура кислоточувствительного ионного канала 1. [1] | |||
| Идентификаторы | |||
| Символ | АСЦ | ||
| Пфам | PF00858 | ||
| ИнтерПро | ИПР001873 | ||
| PROSITE | PDOC00926 | ||
| СКОП2 | 2 кварты / SCOPe / СУПФАМ | ||
| TCDB | 1.А.6 | ||
| Суперсемейство OPM | 181 | ||
| белок OPM | 4fz1 | ||
| |||
Кислоточувствительные ионные каналы ( ASIC ) представляют собой нейронные нечувствительные к напряжению натриевые каналы, активируемые внеклеточными протонами, проницаемыми для Na. + . ASIC1 также показывает низкий уровень Ca 2+ проницаемость. [2] Белки ASIC представляют собой подсемейство ENaC /Deg суперсемейства ионных каналов . Эти гены имеют варианты сплайсинга , которые кодируют несколько изоформ , отмеченных суффиксом. У млекопитающих кислоточувствительные ионные каналы (ASIC) кодируются пятью генами , которые продуцируют субъединицы белка ASIC: ASIC1, ASIC2, ASIC3, ASIC4 и ASIC5 . [3] Три из этих белковых субъединиц собираются в ASIC, которые могут объединяться как в гомотримерные, так и в гетеротримерные каналы, обычно встречающиеся как в центральной, так и в периферической нервной системе . [2] Однако наиболее распространенными ASIC являются ASIC1a, ASIC1a/2a и ASIC3. ASIC2b сам по себе нефункционален, но модулирует активность канала при участии в гетеромультимерах, а ASIC4 не имеет известной функции. В широком смысле ASIC являются потенциальными мишенями для лекарств из-за их участия в патологических состояниях, таких как повреждение сетчатки, судороги и ишемическое повреждение головного мозга. [4] [5]
Структура
[ редактировать ]Каждый кислоточувствительный ионный канал состоит из аминокислотной последовательности из 500–560, которая состоит из шести трансмембранных сегментов — по два на субъединицу (TMD1 и TMD2), цитоплазматических аминокарбоксильных концов и большого внеклеточного домена. [3] Внутриклеточные амино-карбоксильные концевые домены жизненно важны для внутриклеточных белковых взаимодействий и модуляций каналов, ионной проницаемости и шлюзования. Однако ворота и механика каждого кислотно-чувствительного ионного канала определяются комбинацией субъединиц ASIC, которые образуют его структуру. [3]
пора
[ редактировать ]Механика функции пор имеет фундаментальное значение для структуры канала. Между тремя субъединицами ASIC1 туннель простирается от вершины внеклеточных доменов к цитоплазме клетки. Центральный туннель проходит непосредственно между тримерной единицей, где имеет большие суженные участки, меняющие размер и форму в зависимости от состояния канала. [3]
Два трансмембранных домена (TMD1 и TMD2) каждой из трех субъединиц ASIC отвечают за пору канала. TMD2 в первую очередь участвует в выстилке просвета поры и воротах инактивации канала, тогда как TMD1 удерживает белок внутри липидного бислоя клетки . [6] TMD1 связан с β-листами внеклеточного домена, которые сгибаются, расширяя внеклеточный домен и обеспечивая прохождение ионов через канал. [3] Между сегментами TMD2 расположен селективный фильтр, который образует самую узкую часть поры, которая отвечает за доступность ASIC в основном для Na. + . Для ASIC1 девять аминокислотных остатков , по три от каждой субъединицы ASIC (Gly443, Ala444, Ser445), образуют селективный фильтр. Названный «газовым поясом», все три карбонильных атома кислорода выстилают пору, создавая отрицательный потенциал, который способствует проводимости катионов. [3] Специфический аминокислотный остаток аспартата во внеклеточном боковом просвете TMD2 в ASIC1 связан с низким содержанием кальция в канале. 2+ проводимость. Кроме того, n-концевые остатки трансмембранной области также продемонстрировали селективность в отношении Na. + , поскольку мутации в этой области изменили функцию Na + проводимость. [3]
Внеклеточная область
[ редактировать ]ASIC имеют большую внеклеточную область, похожую на кулак, которая поглощает большую часть структуры белка. В его «кулакообразной» структуре имеются домены запястья, ладони, пальца, сустава пальца, большого пальца и β-шарика. «Пальма» составляет большую часть внеклеточного домена, образованного семью β-листами, тогда как остальные вторичные структурные домены состоят из α-спиральных сегментов. [3] Внеклеточная область, отличающаяся своими специфическими аминокислотными конфигурациями, имеет основополагающее значение для индукции активации/инактивации наряду с регулированием pH . Специфическая область петли β-листа между доменами «ладонь» и «большой палец» участвует в передаче сигнала из внеклеточного домена в трансмембранные области, что приводит к конформационному изменению ASIC в открытое состояние. [3] Однако остается неясным, какие именно остатки взаимодействуют с протонами, активируя канал. В 2009 году исследования, возможно, установили связь между ароматическими остатками Tyr72, Pro287 и Trp288 и блокированием протонов ASIC. [3] Эти остатки образуют кислотный карман, который выражает электростатические потенциалы, которые отвечают за зависимость от pH при активации и модуляции каналов. [7] Этот карман во внеклеточном домене действует как резерв для концентрации катионов для дальнейшего содействия расщеплению Na. + приток. Гликозилирование также наблюдается во внеклеточной области, играя важную роль в доставке канала к поверхности мембраны, а также в установлении чувствительности ASIC к уровням pH. Дальнейшие экспериментальные данные показали, что Ca 2+ может также играть ключевую роль в модуляции сродства к протонам ASIC-гейта как внутри поры, так и во внеклеточном домене. [3]
Функция
[ редактировать ]Роль ASIC заключается в том, чтобы обнаружить снижение уровня внеклеточного pH и привести к ответу или сигналу от нейрона. Долгое время считалось, что лигандом, который связывается с сайтом активации, являются исключительно протоны; однако недавние исследования показали, что ASIC4 и ASIC1 могут активироваться при нормальном уровне pH, что указывает на другие типы связывающих лиганды. [8] В повышенных кислых условиях протон связывается с каналом во внеклеточной области, активируя ионный канал для прохождения конформационных изменений, тем самым открывая трансмембранный домен 2 (TMD2). Это приводит к притоку ионов натрия через просвет TMD2. Все ASIC обладают особой проницаемостью для ионов натрия. Единственный вариант — ASIC1a, который также имеет низкую проницаемость для ионов кальция. Приток этих катионов приводит к деполяризации мембраны. Напряжение-управляемый Ca 2+ каналы активируются, что приводит к притоку кальция в клетку. Это вызывает деполяризацию нейрона и высвобождение возбуждающего ответа. В ASIC1a, Ca 2+ увеличение внутри клетки является результатом притока кальция непосредственно через канал. [8]
После активации ASIC может запускать множество различных эффекторных белков и сигнальных молекул, что приводит к различным реакциям клетки. А именно, α-актинин приводит к повышению чувствительности к pH и восстановлению десенсибилизации. Они также могут увеличить плотность тока, проходящего через канал. [8] Существует также множество протеинкиназ , которые регулируют функцию ASIC посредством фосфорилирования . К ним относятся протеинкиназа А (PKA) и протеинкиназа C (PKC). Считается, что существует гораздо больше регуляторов, однако их действие еще не подтверждено экспериментально. [8]
Есть и другие факторы, которые могут сыграть роль в регулировании ASIC. Говорят, что присутствие зрелых N-связанных гликанов на поверхности канала позволяет каналу преимущественно передавать трафик для ASIC1a. Это результат увеличения сайтов N-гликозилирования на ASIC1a и ASIC2a. [8] Высокие уровни глицерина (известно, что он ускоряет созревание белков) на поверхности ASIC2 также помогает предположить, что регуляция функции этих каналов зависит от созревания белков. Также предполагается, что окисление играет роль в незаконном обороте. [8]
Расположение
[ редактировать ]Большинство ASIC экспрессируются в нервной системе. ASIC1, ASIC2, ASIC2b и ASIC4 обычно экспрессируются как в центральной, так и в периферической нервной системе, тогда как ASIC1b и ASIC3 обычно локализуются только в периферической нервной системе.
В периферической нервной системе ASIC расположены в телах клеток постсинаптических мембран и окончаниях сенсорных нервов. Кроме того, ASIC обычно обнаруживаются в афферентных нервных волокнах кожи, мышц, суставов и внутренних органов, где было обнаружено, что они связаны с болью, вкусом и функциями желудочно-кишечного тракта. [6]
В центральной нервной системе ASIC обычно располагаются в задних рогах спинного мозга . [4] ASIC1 конкретно сконцентрирован в миндалевидном теле , что иллюстрирует его роль в тревожном поведении, а ASIC3 был обнаружен в кортиевом органе и спиральном ганглии, что иллюстрирует роль этого специфического канала в слуховом и зрительном восприятии. [6] Субъединицы ASIC1a, ASIC2a и ASIC2b также были обнаружены в гиппокампе. [9]
Физиология
[ редактировать ]ASIC являются потенциальными мишенями для лечения широкого спектра заболеваний, связанных как с ЦНС, так и с ПНС. [4] [5] Особый интерес для поля боли представляет рецептор подтипа ASIC3, который специфически экспрессируется в ноцицепторах . Этот подтип демонстрирует двухфазный ток при активации протона, при котором начальный входящий внутрь Na + за током вскоре следует устойчивый катионный ток.
ASIC важны для функции сетчатки и обеспечивают защиту в ответ на яркий свет. Восприимчивость к повреждению сетчатки увеличивается после делеции гена ASIC2. Усиление апоптоза происходило в ответ на яркий свет в гене ASIC2 -/- по сравнению с сетчаткой дикого типа. [8]
Каналы ASIC1a также играют роль в защите от судорожной активности. Судороги вызывают повышенную неконтролируемую активность нейронов в головном мозге, которая выделяет большое количество кислых везикул. [5] Каналы ASIC1a открываются в ответ и защищают от судорог, замедляя их прогрессирование. Исследования, изучающие этот феномен, показали, что удаление гена ASIC1a привело к усилению судорожной активности. [8]
Каналы ASIC1a специфически открываются в ответ на pH 5,0-6,9 и способствуют патологии ишемического повреждения головного мозга, поскольку их активация вызывает небольшое увеличение Ca 2+ проницаемость и входящий поток Ca 2+ . Каналы ASIC1a дополнительно облегчают активацию потенциалзависимых Ca2+-каналов и каналов рецепторов NMDA при начальной деполяризации, способствуя значительному увеличению внутриклеточного кальция, что приводит к гибели клеток. [10] Возможный механизм гибели клеток, опосредованной каналом ASIC1a, обусловлен активацией других каналов, что приводит к повышению уровня кальция. 2+ который создает сигнальные пути апоптоза и некроза в клетке. [5] Исследования нокаута генов, а также блокады ASIC показали, что они уменьшают объем инфаркта головного мозга на целых 60%, что позволяет предположить, что каналы ASIC играют важную роль в развитии патологических состояний, возникающих в результате ацидоза и повреждения нейронов, вызванного ишемией . [10] Эффекты блокады ASIC и NMDA были изучены, чтобы определить роль обоих каналов в Ca. 2+ токсичности и оценить их соответствующий вклад. Использование блокады обоих каналов обеспечивает большую нейропротекцию, чем использование блокады только одного канала, а блокада ASIC создает пролонгированную эффективность блокады NMDA. [10]
Фармакология
[ редактировать ]Из-за роли кислоточувствительных ионных каналов в восприятии боли и некоторых патофизиологических процессах они имеют фармакологическое значение в качестве мишени для ингибирования. Ионные каналы, чувствительные к кислоте, обнаружены как в центральных, так и в периферических нейронах. Модуляция активности ASIC может дополнительно контролировать неблагоприятные поведенческие и эмоциональные симптомы хронической боли, такие как тревога и депрессия.
Обнаружено, что ионные каналы, чувствительные к кислоте (ASIC), активируются при pH ниже ~ 6 с вариациями, зависящими от типа канала и его местоположения. Снижение pH может быть вызвано множеством причин, включая воспаление тканей, ишемический инсульт, накопление молочной кислоты из-за усиления клеточного метаболизма. Активация канала вызывает повышенную проницаемость ионов натрия, что деполяризует клетку и вызывает возникновение потенциала действия . Полученные потенциалы действия можно модулировать с помощью низкомолекулярных ингибиторов.
Амилорид является примером ингибитора ASIC, хотя он и не считается высокоэффективным из-за значения IC50 в микромолярном диапазоне, но позволил провести исследования влияния ингибирования ASIC на мигрень. Во время мигрени наблюдается распространяющаяся кортикальная депрессия, которая вызывает ионный дисбаланс и высвобождение заряженных молекул, которые могут активировать ASIC. Испытание амилорида на грызунах показало уменьшение кортикальной распространяющейся депрессии во время мигрени. Исследования показали, что амилорид действует как конкурентный ингибитор глав ASIC. Использование амилорида также показало побочные эффекты у грызунов из-за ингибирования обменников натрия/кальция. Ингибирование этих обменников нарушает клеточный гомеостаз кальция и вызывает высокий уровень кальция в клетке, что объясняет снижение нейропротекторной эффективности при использовании амилорида. Результаты, полученные благодаря ингибированию ASIC амилоридом, являются многообещающими и подтверждают терапевтический потенциал. Однако из-за недостаточной специфичности и эффективности амилорида перед его выпуском необходимо будет провести дальнейшую разработку его структуры. [11] [10]
Низкомолекулярный ингибитор А-317567 демонстрирует больший терапевтический потенциал, чем амилорид, с более высокой специфичностью к каналам ASIC и повышенной эффективностью. Хотя A-317567 проявляет небольшую селективность в отношении различных типов ASIC-каналов, результаты in vivo показали, что побочных эффектов, наблюдаемых при использовании амилорида, можно избежать благодаря специфичности A-317567 к ASIC. Кроме того, А-317567 обладает способностью поддерживать ингибирование постоянных токов, что может быть перспективным, особенно при хронических состояниях, опосредованных ацидозом. [10]
Наиболее эффективным и известным ингибитором ASIC является PcTX1. PcTX1 специфически ингибирует ASICa и имеет значение IC50 в наномолярном диапазоне - меньшее значение IC50, чем у всех других известных ингибиторов ASIC, находящихся в микромолярном диапазоне. Кроме того, PcTX1 не ингибирует другие потенциал-управляемые ионные каналы или лиганд-управляемые каналы . Структура этого ингибитора представляет собой 40 аминокислот, связанных дисульфидными связями . Он был идентифицирован как пептидный токсин южноамериканского тарантула Psalmopoeus Cambridge . [10] Когда PcTX1 вводили в базолатеральную миндалину крыс, симптомы, связанные с эмоциями и тревогой, связанные с болью, значительно уменьшались. [12] Мамбалгины, выделенные из яда черной мамбы, также были идентифицированы как мощные ингибиторы ASIC. [13]
широко используемые нестероидные противовоспалительные препараты ( НПВП Было обнаружено, что ) играют роль в ингибировании ASIC, что способствует модуляции боли. Хорошо известным механизмом действия НПВП является ингибирование ими синтеза простагландинов, основного воспалительного соединения. Однако результаты показывают, что НПВП ибупрофен и аспирин ингибируют ASIC со значениями IC50 350 мкм и 260 мкм соответственно. НПВП, вероятно, ингибируют ток ASIC во время острой боли, особенно вызванной воспалением тканей, и, таким образом, подавляют сигнал к болевым нейронам. [10]
Продолжая исследования фармакологического потенциала ингибирования ASIC, пациенты, страдающие хронической болью и различными патологиями, связанными с ацидозом, могут иметь более широкие возможности лечения в будущем. Кроме того, исследования по поиску новых лекарств на основе ASIC дают более глубокие знания о функции самих каналов и их физиологическом значении.
Ссылки
[ редактировать ]- ^ Джасти Дж., Фурукава Х., Гонсалес Э.Б., Гуо Э. (2007). «Структура кислоточувствительного ионного канала 1 с разрешением 1,9 Å и низким pH». Природа . 449 (7160): 316–322. Бибкод : 2007Natur.449..316J . дои : 10.1038/nature06163 . ПМИД 17882215 .
- ^ Jump up to: а б Грюндер С., Пуш М. (июль 2015 г.). «Биофизические свойства кислоточувствительных ионных каналов (ASIC)». Нейрофармакология . 94 : 9–18. doi : 10.1016/j.neuropharm.2014.12.016 . ПМИД 25585135 . S2CID 34111346 .
- ^ Jump up to: а б с д и ж г час я дж к Ханукоглу I (февраль 2017 г.). «Натриевые каналы типа ASIC и ENaC: конформационные состояния и структуры ионно-селективных фильтров» . Журнал ФЭБС . 284 (4): 525–545. дои : 10.1111/февраль 13840 . ПМИД 27580245 . S2CID 24402104 .
- ^ Jump up to: а б с Слука К.А., Зимний ОК, Уэмми Дж.А. (сентябрь 2009 г.). «Кислотно-чувствительные ионные каналы: новая мишень для лечения боли и заболеваний ЦНС» . Текущее мнение об открытии и разработке лекарств . 12 (5): 693–704. ПМЦ 3494879 . ПМИД 19736627 .
- ^ Jump up to: а б с д Ван Ю.З., Сюй Т.Л. (декабрь 2011 г.). «Ацидоз, кислоточувствительные ионные каналы и гибель нейрональных клеток». Молекулярная нейробиология . 44 (3): 350–8. дои : 10.1007/s12035-011-8204-2 . ПМИД 21932071 . S2CID 15169653 .
- ^ Jump up to: а б с Осьмаков Д.И., Андреев Ю.А., Козлов С.А. (2014). «Кислоточувствительные ионные каналы и их модуляторы». Биохимия. Биохимия . 79 (13): 1528–45. дои : 10.1134/S0006297914130069 . ПМИД 25749163 . S2CID 14874830 .
- ^ Шервуд Т.В., Фрей Э.Н., Асквит CC (октябрь 2012 г.). «Структура и активность кислоточувствительных ионных каналов» . Американский журнал физиологии. Клеточная физиология . 303 (7): C699–710. doi : 10.1152/ajpcell.00188.2012 . ПМЦ 3469599 . ПМИД 22843794 .
- ^ Jump up to: а б с д и ж г час Жа ХМ (январь 2013 г.). «Кислоточувствительные ионные каналы: торговля и синаптическая функция» . Молекулярный мозг . 6 :1. дои : 10.1186/1756-6606-6-1 . ПМК 3562204 . ПМИД 23281934 .
- ^ Барон, А.; Вальдманн, Р.; Лаздунски, М. (2002). «ASIC-подобные протонно-активируемые токи в нейронах гиппокампа крысы. Журнал физиологии, 539 (2), 485–494 | 10.1113/jphysicalol.2001.014837» . Журнал физиологии . 539 (Часть 2): 485–494. дои : 10.1113/jphysicalol.2001.014837 . ПМК 2290154 . ПМИД 11882680 .
- ^ Jump up to: а б с д и ж г Сюн З.Г., Пиньятаро Г., Ли М., Чанг С.Ю., Саймон Р.П. (февраль 2008 г.). «Кислотно-чувствительные ионные каналы (ASIC) как фармакологические мишени при нейродегенеративных заболеваниях» . Современное мнение в фармакологии . Нейронауки. 8 (1): 25–32. дои : 10.1016/j.coph.2007.09.001 . ПМК 2267925 . ПМИД 17945532 .
- ^ Барон А, Лингуэлья Э (июль 2015 г.). «Фармакология кислоточувствительных ионных каналов - физиологические и терапевтические перспективы» (PDF) . Нейрофармакология . Кислоточувствительные ионные каналы в нервной системе. 94 : 19–35. doi : 10.1016/j.neuropharm.2015.01.005 . ПМИД 25613302 . S2CID 25550294 .
- ^ Айсуни, Юсеф; Эль Герраб, Абдеррахим; Хамие, Аль-Махди; Ферье, Жереми; Шалюс, Мариз; Лемэр, Дайан; Грегуар, Стефани; Этьен, Моник; Эшалье, Ален (2 марта 2017 г.). «Кислоточувствительный ионный канал 1а в миндалевидном теле участвует в поведении, связанном с болью и тревогой, связанном с артритом» . Научные отчеты . 7 : 43617. Бибкод : 2017NatSR...743617A . дои : 10.1038/srep43617 . ISSN 2045-2322 . ПМК 5340794 . ПМИД 28321113 .
- ^ Диошо С., Барон А., Салинас М., Дуге Д., Скарцелло С., Даберт-Гей А.С. и др. (октябрь 2012 г.). «Пептиды яда черной мамбы воздействуют на кислоточувствительные ионные каналы, чтобы облегчить боль» (PDF) . Природа . 490 (7421): 552–5. Бибкод : 2012Natur.490..552D . дои : 10.1038/nature11494 . ПМИД 23034652 . S2CID 4337253 .