N- связанное гликозилирование

N -связанное гликозилирование — это присоединение олигосахарида , углевода, состоящего из нескольких молекул сахара, иногда также называемого гликаном , к атому азота ( амидному азоту остатка аспарагина (Asn) белка ) в процессе, называемом N -гликозилирование , изучаемое в биохимии . [ 1 ] Полученный белок называется N-связанным гликаном или просто N-гликаном .
Этот тип связи важен как для структуры, так и для [ 2 ] и функция [ 3 ] многих эукариотических белков. Процесс N -связанного гликозилирования встречается у эукариот и широко у архей , но очень редко у бактерий . Природа N -связанных гликанов, присоединенных к гликопротеину, определяется белком и клеткой, в которой он экспрессируется. [ 4 ] Это также варьируется в зависимости от вида . Разные виды синтезируют разные типы N -связанных гликанов.
Энергетика образования связей
[ редактировать ]В гликопротеине присутствуют связи двух типов: связи между остатками сахаридов в гликане и связь между гликановой цепью и молекулой белка.
Сахарные фрагменты связаны друг с другом в гликановой цепи посредством гликозидных связей . Эти связи обычно образуются между атомами углерода 1 и 4 молекул сахара. Образование гликозидной связи энергетически невыгодно, поэтому реакция сопряжена с гидролизом двух молекул АТФ . [ 4 ]
С другой стороны, присоединение остатка гликана к белку требует распознавания консенсусной последовательности . N -связанные гликаны почти всегда присоединены к атому азота боковой цепи аспарагина (Asn), которая присутствует как часть консенсусной последовательности Asn–X– Ser / Thr , где X – любая аминокислота, кроме пролина (Pro). [ 4 ]
В клетках животных гликан, присоединенный к аспарагину, почти всегда представляет собой N- ацетилглюкозамин (GlcNAc) в β-конфигурации. [ 4 ] Эта β-связь аналогична гликозидной связи между фрагментами сахара в гликановой структуре, как описано выше. Вместо того, чтобы быть присоединенным к гидроксильной группе сахара, аномерный атом углерода присоединен к амидному азоту. Энергия, необходимая для этой связи, поступает в результате гидролиза молекулы пирофосфата . [ 4 ]
Биосинтез
[ редактировать ]
Биосинтез N -связанных гликанов происходит в три основных этапа: [ 4 ]
- Синтез долихолсвязанного олигосахарида-предшественника
- Блоковый перенос олигосахарида-предшественника в белок
- Обработка олигосахарида
Синтез, перенос единым блоком и начальная обрезка олигосахарида- предшественника происходят в эндоплазматическом ретикулуме (ЭР). Последующая обработка и модификация олигосахаридной цепи осуществляются в аппарате Гольджи .
Таким образом, синтез гликопротеинов пространственно разделен в разных клеточных компартментах. Следовательно, тип синтезируемого N -гликана зависит от его доступности для различных ферментов, присутствующих в этих клеточных компартментах.
Однако, несмотря на разнообразие, все N -гликаны синтезируются общим путем с общей гликановой структурой ядра. [ 4 ] Основная структура гликана по существу состоит из двух N -ацетилглюкозамина и трех остатков маннозы . Этот основной гликан затем дорабатывается и модифицируется, что приводит к появлению разнообразного спектра N -гликановых структур. [ 4 ]
Синтез предшественника олигосахарида
[ редактировать ]Процесс N -связанного гликозилирования начинается с образования долихол -связанного сахара GlcNAc. Долихол представляет собой липидную молекулу, состоящую из повторяющихся изопрена единиц . Эта молекула прикреплена к мембране ЭР. Молекулы сахара присоединяются к долихолу посредством пирофосфатной связи. [ 4 ] (один фосфат изначально был связан с долихолом, а второй фосфат происходил из нуклеотидного сахара). Затем олигосахаридная цепь удлиняется за счет поэтапного добавления различных молекул сахара с образованием олигосахарида-предшественника.
Сборка этого олигосахарида-предшественника происходит в две фазы: фаза I и II. [ 4 ] Фаза I происходит на цитоплазматической стороне ЭР, а фаза II - на люминальной стороне ЭР.
Молекула-предшественник, готовая к переносу в белок, состоит из двух молекул GlcNAc, девяти молекул маннозы и трех молекул глюкозы .

Фаза I
| |
|---|---|
Шаги |
Расположение
|
|
Цитоплазматическая сторона ЭР
|
|
В этот момент связанный с липидами гликан транслоцируется через мембрану, делая его доступным для ферментов в просвете эндоплазматического ретикулума. Этот процесс транслокации до сих пор плохо изучен, но предполагается, что он осуществляется ферментом, известным как флиппаза .
| |
| Фаза II | |
|
Люминальная сторона ER
|
Перенос гликана в белок
[ редактировать ]Как только образуется олигосахарид-предшественник, завершенный гликан затем переносится в образующийся полипептид в просвете мембраны ЭР. Эта реакция осуществляется за счет энергии, выделяющейся при разрыве пирофосфатной связи между молекулой долихол-гликана. Прежде чем гликан перенесется в образующийся полипептид, необходимо выполнить три условия: [ 4 ]
- Аспарагин должен располагаться в определенной консенсусной последовательности первичной структуры (Asn-X-Ser или Asn-X-Thr или, в редких случаях, Asn-X-Cys). [ 5 ]
- Аспарагин должен располагаться соответствующим образом в трехмерной структуре белка (сахара являются полярными молекулами и, следовательно, должны быть присоединены к аспарагину, расположенному на поверхности белка, а не скрытому внутри белка).
- связанное гликозилирование , аспарагин должен находиться на люминальной стороне эндоплазматического ретикулума N- Чтобы инициировать . Целевые остатки обнаруживаются либо в секреторных белках, либо в участках трансмембранного белка, обращенных к просвету.
Олигосахарилтрансфераза представляет собой фермент, ответственный за распознавание консенсусной последовательности и перенос гликана-предшественника на акцептор полипептида, который транслируется в просвете эндоплазматического ретикулума. Таким образом, N- связанное гликозилирование является событием котрансляции.
Переработка гликана
[ редактировать ]
Процессинг N -гликанов осуществляется в эндоплазматическом ретикулуме и телецe Гольджи. Начальная обрезка молекулы-предшественника происходит в ЭР, а последующий процессинг происходит в Гольджи.
При переносе завершенного гликана на образующийся полипептид из структуры удаляются два остатка глюкозы. Ферменты, известные как гликозидазы, удаляют некоторые остатки сахара. Эти ферменты могут разрывать гликозидные связи, используя молекулу воды. Эти ферменты являются экзогликозидазами, поскольку они воздействуют только на остатки моносахаридов , расположенные на невосстанавливающем конце гликана. [ 4 ] Считается, что этот начальный этап обрезки действует как этап контроля качества в ER для мониторинга сворачивания белка .
Как только белок сложен правильно, два остатка глюкозы удаляются глюкозидазой I и II. Удаление последнего третьего остатка глюкозы сигнализирует о том, что гликопротеин готов к транзиту из ЭР в цис -Гольджи. [ 4 ] ЭР-маннозидаза катализирует удаление этой конечной глюкозы. Однако, если белок не свернут должным образом, остатки глюкозы не удаляются и, таким образом, гликопротеин не может покинуть эндоплазматический ретикулум. Белок -шаперон ( кальнексин / кальретикулин ) связывается с развернутым или частично свернутым белком, помогая сворачиванию белка.
Следующий шаг включает дальнейшее добавление и удаление остатков сахара в цис-Гольджи. Эти модификации катализируются гликозилтрансферазами и гликозидазами соответственно. В цис -Гольджи ряд маннозидаз удаляют некоторые или все четыре остатка маннозы в α-1,2-связях. [ 4 ] Тогда как в медиальной части Гольджи гликозилтрансферазы добавляют остатки сахаров к основной гликановой структуре, образуя три основных типа гликанов: высокоманозные, гибридные и сложные гликаны.
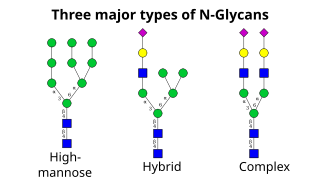
- По сути, высокоманноза представляет собой всего лишь два N -ацетилглюкозамина со многими остатками маннозы, часто почти такими же, как в олигосахаридах-предшественниках до того, как они прикрепятся к белку.
- Сложные олигосахариды названы так потому, что они могут содержать практически любое количество других типов сахаридов, включая больше, чем исходные два N -ацетилглюкозамина.
- Гибридные олигосахариды содержат остатки маннозы на одной стороне ветви, а на другой стороне N -ацетилглюкозамин инициирует сложную ветвь.
Порядок присоединения сахаров к растущим гликановым цепям определяется субстратной специфичностью ферментов и их доступом к субстрату при продвижении по секреторному пути . Таким образом, организация этого механизма внутри клетки играет важную роль в определении того, какие гликаны образуются.
Ферменты в Гольджи
[ редактировать ]Ферменты Гольджи играют ключевую роль в определении синтеза различных типов гликанов. Порядок действия ферментов отражается в их положении в стопке Гольджи:
| Ферменты | Расположение в Гольджи |
|---|---|
| Маннозидаза I | цис -Гольджи |
| GlcNAc трансферазы | медиальный Гольджи |
| Галактозилтрансфераза и сиалилтрансфераза | транс -Гольджи |
У архей и прокариот
[ редактировать ]Сходный путь биосинтеза N -гликанов был обнаружен у прокариот и архей. [ 6 ] Однако по сравнению с эукариотами окончательная структура гликана у эубактерий и архей, по-видимому, не сильно отличается от исходного предшественника, созданного в эндоплазматическом ретикулуме. У эукариот исходный олигосахарид-предшественник сильно модифицируется на пути к поверхности клетки. [ 4 ]
Функция
[ редактировать ]N -связанные гликаны имеют внутренние и внешние функции. [ 4 ] [ 7 ]
В иммунной системе N -связанные гликаны на поверхности иммунных клеток помогают определить характер миграции клетки, например, иммунные клетки, которые мигрируют в кожу, имеют специфические гликозилирования, способствующие возвращению в этот участок. [ 8 ] Паттерны гликозилирования различных иммуноглобулинов, включая IgE, IgM, IgD, IgA и IgG, наделяют их уникальными эффекторными функциями за счет изменения их сродства к Fc и другим иммунным рецепторам. [ 8 ] Гликаны также могут участвовать в различении «своих» и «чужих», что может иметь отношение к патофизиологии различных аутоиммунных заболеваний. [ 8 ]
| Внутренний | |
| Внешний |
|
В некоторых случаях взаимодействие между N-гликаном и белком стабилизирует белок посредством сложных электронных эффектов. [ 11 ]
Клиническое значение
[ редактировать ]Изменения N -связанного гликозилирования связаны с различными заболеваниями, включая ревматоидный артрит , [ 12 ] диабет 1 типа , [ 13 ] болезнь Крона , [ 14 ] и раковые заболевания. [ 15 ] [ 16 ]
Мутации в восемнадцати генах, участвующих в N -связанном гликозилировании, приводят к множеству заболеваний, большинство из которых затрагивают нервную систему . [ 3 ] [ 16 ]
Значение терапевтических белков
[ редактировать ]Многие терапевтические белки, представленные на рынке, представляют собой антитела , которые представляют собой N -связанные гликопротеины. Например, этанерцепт , инфликсимаб и ритуксимаб представляют собой N- гликозилированные терапевтические белки.

Важность N -связанного гликозилирования становится все более очевидной в области фармацевтики . [ 17 ] Хотя бактериальные или дрожжевые системы производства белка обладают значительными потенциальными преимуществами, такими как высокий выход и низкая стоимость, проблемы возникают, когда интересующий белок представляет собой гликопротеин. Большинство прокариотических систем экспрессии, таких как E. coli, не могут осуществлять посттрансляционные модификации . С другой стороны, эукариотические хозяева экспрессии, такие как дрожжи и клетки животных, имеют разные паттерны гликозилирования. Белки, продуцируемые этими экспрессирующими хозяевами, часто не идентичны человеческим белкам и, таким образом, вызывают иммуногенные реакции у пациентов. Например, S.cerevisiae (дрожжи) часто продуцируют иммуногенные гликаны с высоким содержанием маннозы.
Системы экспрессии млекопитающих, отличных от человека, такие как CHO или клетки NS0, обладают механизмом, необходимым для добавления сложных гликанов человеческого типа. Однако гликаны, продуцируемые в этих системах, могут отличаться от гликанов, продуцируемых у человека, поскольку они могут быть кэпированы как N -гликолилнейраминовой кислотой (Neu5Gc), так и N -ацетилнейраминовой кислотой (Neu5Ac), тогда как клетки человека продуцируют только гликопротеины, содержащие N -ацетилнейраминовую кислоту. Кроме того, клетки животных также могут продуцировать гликопротеины, содержащие эпитоп галактоза-альфа-1,3-галактозы , который может вызывать серьезные аллергические реакции, включая анафилактический шок , у людей, страдающих аллергией на альфа-гал .
Эти недостатки были устранены с помощью нескольких подходов, таких как устранение путей, которые производят эти гликановые структуры, посредством генетического нокаута. Кроме того, были генетически сконструированы другие системы экспрессии для производства терапевтических гликопротеинов с человеческими N -связанными гликанами. К ним относятся дрожжи, такие как Pichia Pastoris , [ 18 ] линии клеток насекомых, зеленые растения, [ 19 ] и даже бактерии.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Гликозилирование» . UniProt: Последовательность белка и функциональная информация .
- ^ Империали Б, О'Коннор С.Э. (декабрь 1999 г.). «Влияние N -связанного гликозилирования на структуру гликопептидов и гликопротеинов». Современное мнение в области химической биологии . 3 (6): 643–9. дои : 10.1016/S1367-5931(99)00021-6 . ПМИД 10600722 .
- ^ Jump up to: а б Паттерсон MC (сентябрь 2005 г.). «Метаболические мимики: нарушения N -связанного гликозилирования». Семинары по детской неврологии . 12 (3): 144–51. дои : 10.1016/j.spen.2005.10.002 . ПМИД 16584073 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Дрикамер К., Тейлор М.Э. (2006). Введение в гликобиологию (2-е изд.). Издательство Оксфордского университета, США. ISBN 978-0-19-928278-4 .
- ^ Меллквист Дж.Л., Кастури Л., Спиталник С.Л., Шакин-Эшлеман Ш.Х. (май 1998 г.). «Аминокислота, следующая за секвоном Asn-X-Ser/Thr, является важным фактором, определяющим эффективность N -связанного корового гликозилирования». Биохимия . 37 (19): 6833–7. дои : 10.1021/bi972217k . ПМИД 9578569 .
- ^ Делл А., Галадари А., Састре Ф., Хитчен П. (2010). «Сходства и различия механизмов гликозилирования у прокариот и эукариот» . Международный журнал микробиологии . 2010 : 1–14. дои : 10.1155/2010/148178 . ПМК 3068309 . ПМИД 21490701 .
- ^ ГлиГен. «Словарь структуры гликанов GlyGen» . ГлиГен . Проверено 1 апреля 2021 г.
- ^ Jump up to: а б с Маверакис Э., Ким К., Шимода М., Гершвин М.Е., Патель Ф., Уилкен Р. и др. (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор» . Журнал аутоиммунитета . 57 (6): 1–13. дои : 10.1016/j.jaut.2014.12.002 . ПМЦ 4340844 . ПМИД 25578468 .
- ^ Синклер А.М., Эллиот С. (август 2005 г.). «Гликоинженерия: влияние гликозилирования на свойства терапевтических белков». Журнал фармацевтических наук . 94 (8): 1626–35. дои : 10.1002/jps.20319 . ПМИД 15959882 .
- ^ «N-гликозилирование как защитный механизм эукариот против агрегации белков» . Достижения науки . 10 (5). 2024. doi : 10.1126/sciadv.adk8173 . ПМЦ 10830103 .
- ^ Ардеяни, Мазиар С.; Нудлман, Луи; Пауэрс, Эван Т.; Келли, Джеффри В. (15 марта 2021 г.). «Стереоэлектронные эффекты в стабилизации взаимодействий белок-N-гликан, выявленные экспериментом и машинным обучением» . Природная химия . 13 (5): 480–487. дои : 10.1038/s41557-021-00646-w . ISSN 1755-4349 . ПМК 8102341 . ПМИД 33723379 .
- ^ Накагава Х., Хато М., Такегава Ю., Дегучи К., Ито Х., Такахата М. и др. (июнь 2007 г.). «Обнаружение измененных профилей N -гликанов в цельной сыворотке пациентов с ревматоидным артритом». Журнал хроматографии Б. 853 (1–2): 133–7. дои : 10.1016/j.jchromb.2007.03.003 . hdl : 2115/28276 . ПМИД 17392038 .
- ^ Бермингем М.Л., Коломбо М., МакГурнаган С.Дж., Блэкборн Л.А., Вучкович Ф., Пучич Бакович М. и др. (январь 2018 г.). « Профиль N -гликанов и заболевания почек при диабете 1 типа» . Уход при диабете . 41 (1): 79–87. дои : 10.2337/dc17-1042 . hdl : 20.500.11820/413dce5a-e852-4787-aac9-62c2c6d4389f . ПМИД 29146600 .
- ^ Трбоевич Акмачич И., Вентам Н.Т., Теодорату Э., Вучкович Ф., Кеннеди Н.А., Криштич Дж. и др. (июнь 2015 г.). «Воспалительные заболевания кишечника связаны с провоспалительным потенциалом гликома иммуноглобулина G» . Воспалительные заболевания кишечника . 21 (6): 1237–47. doi : 10.1097/MIB.0000000000000372 . ПМЦ 4450892 . ПМИД 25895110 .
- ^ Кодар К., Стадлманн Й., Клаамас К., Сергеев Б., Куртенков О. (январь 2012 г.). «Профилирование иммуноглобулина G Fc N -гликанов у пациентов с раком желудка с помощью LC-ESI-MS: связь с прогрессированием опухоли и выживаемостью». Гликоконъюгатный журнал . 29 (1): 57–66. дои : 10.1007/s10719-011-9364-z . ПМИД 22179780 . S2CID 254501034 .
- ^ Jump up to: а б Чэнь Г, Ван Ю, Цинь X, Ли Х, Го Ю, Ван Ю и др. (август 2013 г.). «Изменение IgG1 Fc N -связанного гликозилирования при раке легких человека: диагностический потенциал, связанный с возрастом и полом». Электрофорез . 34 (16): 2407–16. дои : 10.1002/elps.201200455 . ПМИД 23766031 . S2CID 11131196 .
- ^ Далзиел М., Криспин М., Сканлан К.Н., Зитцманн Н., Двек Р.А. (январь 2014 г.). «Новые принципы терапевтического использования гликозилирования». Наука . 343 (6166): 1235681. doi : 10.1126/science.1235681 . ПМИД 24385630 . S2CID 206548002 .
- ^ Гамильтон С.Р., Бобровиц П., Бобровиц Б., Дэвидсон Р.К., Ли Х., Митчелл Т. и др. (август 2003 г.). «Производство сложных гликопротеинов человека в дрожжах». Наука . 301 (5637): 1244–6. дои : 10.1126/science.1088166 . ПМИД 12947202 . S2CID 38981893 .
- ^ Штрассер Р., Альтманн Ф., Штайнкельнер Х. (декабрь 2014 г.). «Контролируемое гликозилирование рекомбинантных белков растительного происхождения». Современное мнение в области биотехнологии . 30 :95–100. дои : 10.1016/j.copbio.2014.06.008 . ПМИД 25000187 .
Внешние ссылки
[ редактировать ]- GlycoEP : платформа In silico для прогнозирования N- , O- и C -гликозитов в последовательностях эукариотических белков
- Маверакис Э., Ким К., Шимода М., Гершвин М.Е., Патель Ф., Уилкен Р., Райчаудхури С., Рухаак Л.Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор» . Журнал аутоиммунитета . 57 : 1–13. дои : 10.1016/j.jaut.2014.12.002 . ПМЦ 4340844 . ПМИД 25578468 .