Потенциал покоя
Относительно статический мембранный потенциал, который обычно называют основным значением трансмембранного напряжения.
/К +
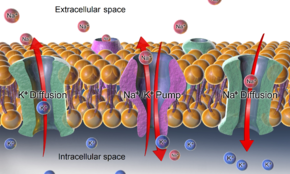
-АТФаза , а также эффекты диффузии участвующих ионов являются основными механизмами поддержания потенциала покоя через мембраны клеток животных.
Относительно статический мембранный потенциал покоящихся (или напряжением покоя), в отличие клеток называется мембранным потенциалом покоя от специфических динамических электрохимических явлений, называемых потенциалом действия и градуированным мембранным потенциалом .
Помимо двух последних, которые встречаются в возбудимых клетках ( нейронах , мышцах и некоторых секреторных клетках желез ), мембранное напряжение в большинстве невозбудимых клеток также может претерпевать изменения в ответ на внешние или внутриклеточные стимулы. Потенциал покоя существует из-за различий в проницаемости мембран для калия , натрия , кальция и хлорида ионов , что, в свою очередь, является результатом функциональной активности различных ионных каналов , переносчиков ионов и обменников. Условно мембранный потенциал покоя можно определить как относительно стабильное основное значение трансмембранного напряжения в клетках животных и растений.
Поскольку проницаемость мембраны для калия намного выше, чем для других ионов, а также из-за сильного химического градиента для калия, ионы калия перетекают из цитозоля во внеклеточное пространство, неся положительный заряд, пока их движение не уравновешивается накоплением. отрицательного заряда на внутренней поверхности мембраны. Опять же, из-за высокой относительной проницаемости для калия результирующий мембранный потенциал почти всегда близок к реверсивному потенциалу калия . Но для того, чтобы этот процесс произошел, необходимо сначала создать градиент концентрации ионов калия. Эта работа выполняется ионными насосами/переносчиками и/или обменниками и обычно приводится в действие АТФ .
В случае мембранного потенциала покоя на плазматической мембране животной клетки градиенты калия (и натрия) устанавливаются Na + /К + -АТФаза (натрий-калиевый насос), переносящая 2 иона калия внутрь и 3 иона натрия наружу за счет 1 молекулы АТФ. В других случаях, например, мембранный потенциал может быть установлен за счет подкисления внутренней части мембранного компартмента (например, протонного насоса, который генерирует мембранный потенциал через мембраны синаптических пузырьков ). [ нужна ссылка ]
Электронейтральность
[ редактировать ]В большинстве количественных обработок мембранного потенциала, таких как вывод уравнения Гольдмана , электронейтральность предполагается ; то есть нет измеримого избытка заряда по обе стороны мембраны. Таким образом, хотя на мембране существует электрический потенциал из-за разделения зарядов, не существует реальной измеримой разницы в общей концентрации положительных и отрицательных ионов на мембране (как это оценивается ниже ), то есть не существует фактической измеримой разницы в общей концентрации положительных и отрицательных ионов на мембране. заряжайте излишки с обеих сторон. Это происходит потому, что влияние заряда на электрохимический потенциал намного сильнее, чем влияние концентрации, поэтому необнаружимое изменение концентрации приводит к значительному изменению электрического потенциала. [ нужна ссылка ]
Генерация потенциала покоя
[ редактировать ]Клеточные мембраны обычно проницаемы только для определенного набора ионов. Обычно к ним относятся ионы калия, ионы хлорида, ионы бикарбоната и другие. Чтобы упростить описание ионной основы мембранного потенциала покоя, полезнее всего сначала рассмотреть только один вид ионов, а остальные рассмотреть позже. Поскольку потенциалы трансплазматической мембраны почти всегда определяются в первую очередь проницаемостью калия, именно с этого и следует начинать.
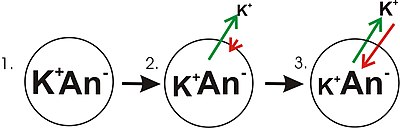
- На панели 1 диаграммы показано схематическое изображение простой ячейки, в которой уже установлен градиент концентрации. Эта панель нарисована так, как будто мембрана не проницаема для каких-либо ионов. Мембранного потенциала нет, поскольку, несмотря на наличие градиента концентрации калия, суммарного дисбаланса зарядов на мембране нет. Если бы мембрана стала проницаемой для типа иона, который более сконцентрирован на одной стороне мембраны, тогда этот ион внес бы вклад в мембранное напряжение, потому что проникающие ионы будут перемещаться через мембрану с итоговым перемещением ионов этого типа вниз по концентрации. градиент. Будет чистое движение со стороны мембраны с более высокой концентрацией ионов в сторону с более низкой концентрацией. Такое движение одного иона через мембрану приведет к суммарному дисбалансу заряда мембраны и мембранного потенциала. Это общий механизм, с помощью которого многие клетки устанавливают мембранный потенциал.
- На панели 2 диаграммы клеточная мембрана сделана проницаемой для ионов калия, но не для анионов (An − ) внутри клетки. Эти анионы в основном представлены белком. В градиенте концентрации ионов калия запасается энергия, которая может быть преобразована в электрический градиент, когда калий (K + ) ионы выходят из клетки. Обратите внимание, что ионы калия могут перемещаться через мембрану в обоих направлениях, но в результате чисто статистического процесса, возникающего из-за более высокой концентрации ионов калия внутри клетки, больше ионов калия будет выходить из клетки. Поскольку внутри клеток имеется более высокая концентрация ионов калия, их случайное молекулярное движение с большей вероятностью столкнется с проницаемой порой ( ионным каналом ), как это происходит с ионами калия, находящимися снаружи и с более низкой концентрацией. Внутренний К + просто «с большей вероятностью» покинет клетку, чем внеклеточный K + это войти в него. Речь идет о диффузии , совершающей работу по рассеиванию градиента концентрации. Покидая клетку, калий оставляет после себя анионы. Поэтому разделение зарядов развивается как K + покидает клетку. Такое разделение зарядов создает трансмембранное напряжение. Это трансмембранное напряжение и есть мембранный потенциал. Поскольку калий продолжает покидать клетку, разделяя все больше зарядов, мембранный потенциал будет продолжать расти. Длина стрелок (зеленые указывают на градиент концентрации, красные указывают на напряжение) представляют собой величину движения ионов калия за счет каждой формы энергии. Направление стрелки указывает направление приложения этой конкретной силы. Таким образом, напряжение на мембране здания представляет собой возрастающую силу, которая действует вопреки тенденции к чистому движению ионов калия вниз по градиенту концентрации калия.
- На панели 3 мембранное напряжение выросло до такой степени, что его «сила» теперь соответствует градиентам концентрации. Поскольку эти силы (приложенные к K + ) теперь имеют одинаковую силу и ориентированы в противоположных направлениях, система теперь находится в равновесии . Другими словами, тенденция калия покидать клетку, снижая градиент своей концентрации, теперь сочетается с тенденцией мембранного напряжения притягивать ионы калия обратно в клетку. К + продолжает двигаться через мембрану, но скорость, с которой он входит в клетку и покидает ее, одинакова, поэтому чистый ток калия отсутствует. Потому что К + находится в равновесии, мембранный потенциал стабилен или «покоится» (EK ) .
Напряжение покоя является результатом работы нескольких ферментов, переносящих ионы ( унипортеров , котранспортеров и насосов ) в плазматической мембране, постоянно работающих параллельно, при этом каждый ион-транслокатор имеет свою характерную электродвижущую силу (= обратный потенциал = «равновесное напряжение»). , в зависимости от конкретных концентраций субстрата внутри и снаружи ( внутренний АТФ в некоторых насосах включен ). ЧАС + экспорт АТФазы делает мембранное напряжение у растений и грибов гораздо более отрицательным, чем в более тщательно изученных животных клетках, где напряжение покоя в основном определяется селективными ионными каналами.
У большинства нейронов потенциал покоя имеет значение примерно −70 мВ. Потенциал покоя в основном определяется концентрацией ионов в жидкостях по обе стороны клеточной мембраны и переносчиками ионов белками- , которые находятся в клеточной мембране. Ниже показано, как концентрации ионов и мембранных транспортных белков влияют на величину потенциала покоя.
Потенциал покоя клетки можно наиболее полно понять, рассматривая его в терминах равновесных потенциалов. На приведенном здесь примере диаграммы в модельную ячейку был добавлен только один проникающий ион (калий). В этом случае потенциал покоя этой клетки будет таким же, как и равновесный потенциал калия.
Однако реальная клетка устроена сложнее: она обладает проницаемостью для многих ионов, каждый из которых вносит свой вклад в потенциал покоя. Чтобы лучше понять, рассмотрим клетку, в которую проникают только два иона: калия и натрия. Рассмотрим случай, когда эти два иона имеют равные градиенты концентрации, направленные в противоположные стороны, и что проницаемость мембраны для обоих ионов одинакова. К + клетки будет иметь тенденцию смещать мембранный потенциал в сторону EK выход из . На + попадание в клетку будет иметь тенденцию смещать мембранный потенциал в сторону обратного потенциала натрия E Na . Поскольку проницаемости для обоих ионов были равны, мембранный потенциал в конце Na + /К + перетягивание каната, в конечном итоге оказываемся на полпути E Na и EK между . Поскольку E Na и E K были равны, но имели противоположные знаки, посередине между ними находится ноль, что означает, что мембрана будет находиться при напряжении 0 мВ.
Обратите внимание, что хотя мембранный потенциал при 0 мВ стабилен, он не является состоянием равновесия, поскольку ни один из вносящих вклад ионов не находится в равновесии. Ионы диффундируют по своим электрохимическим градиентам через ионные каналы, но мембранный потенциал поддерживается постоянным K + приток и Na + отток через переносчики ионов . Такая ситуация с одинаковой проницаемостью для противодействующих ионов, таких как калий и натрий, в клетках животных может быть чрезвычайно дорогостоящей для клетки, если эта проницаемость относительно велика, поскольку АТФ для перекачки ионов обратно требуется много энергии . Поскольку ни одна реальная клетка не может обеспечить такую равную и большую ионную проницаемость в состоянии покоя, потенциал покоя животных клеток определяется преобладающей высокой проницаемостью для калия и регулируется до необходимого значения путем модуляции и градиентов проницаемости для натрия и хлоридов.
В здоровой животной клетке Na + проницаемость составляет около 5% от K + проницаемость или даже меньше, тогда как соответствующие реверсивные потенциалы составляют +60 мВ для натрия ( E Na ) и -80 мВ для калия ( E K ). Таким образом, мембранный потенциал не будет соответствовать EK EK , а скорее будет деполяризован от на EK величину примерно 5% от разницы в 140 между мВ и E Na . Таким образом, потенциал покоя клетки составит около -73 мВ.
В более формальных обозначениях мембранный потенциал представляет собой средневзвешенное значение равновесного потенциала каждого вносящего вклад иона. Размер каждой гири представляет собой относительную проводимость каждого иона. В обычном случае, когда три иона вносят вклад в мембранный потенциал:
- ,

где
- Em . — мембранный потенциал, измеряемый в вольтах
- E X — равновесный потенциал иона X, также в вольтах.
- g X / g tot — относительная проводимость иона X, которая безразмерна
- g tot — общая проводимость всех проникающих ионов в произвольных единицах (например, в сименсах для электропроводности), в данном случае g K + + г Na + + г Cl −
Мембранные транспортные белки
[ редактировать ]Для определения мембранных потенциалов двумя наиболее важными типами мембранных белков-переносчиков ионов являются ионные каналы и переносчики ионов . Белки ионных каналов создают пути через клеточные мембраны, по которым ионы могут пассивно диффундировать без прямых затрат метаболической энергии. Они обладают селективностью по отношению к определенным ионам, поэтому существуют калий- , хлорид- и натрий-селективные ионные каналы . Разные клетки и даже разные части одной клетки ( дендриты , тела клеток , узлы Ранвье ) будут иметь разное количество различных белков-переносчиков ионов. Обычно количество определенных калиевых каналов наиболее важно для контроля потенциала покоя (см. ниже). Некоторые ионные насосы, такие как Na+/K+-АТФаза, являются электрогенными, то есть они вызывают дисбаланс зарядов клеточной мембраны, а также могут непосредственно способствовать мембранному потенциалу. Большинство насосов для своего функционирования используют метаболическую энергию (АТФ).
Равновесные потенциалы
[ редактировать ]Для большинства животных клеток ионы калия (K + ) являются наиболее важными для потенциала покоя. [1] Благодаря активному транспорту ионов калия концентрация калия внутри клеток выше, чем снаружи. Большинство клеток имеют белки калий-селективных ионных каналов, которые все время остаются открытыми. Через эти калиевые каналы будет происходить чистое движение положительно заряженных ионов калия, что приводит к накоплению избыточного отрицательного заряда внутри клетки. Движение положительно заряженных ионов калия наружу происходит из-за случайного молекулярного движения ( диффузии ) и продолжается до тех пор, пока внутри клетки не накопится достаточно избыточного отрицательного заряда, чтобы сформировать мембранный потенциал, который может уравновесить разницу в концентрации калия внутри и снаружи клетки. «Баланс» означает, что электрическая сила ( потенциал ), возникающая в результате накопления ионного заряда и препятствующая диффузии наружу, увеличивается до тех пор, пока не станет равна по величине, но противоположна по направлению тенденции диффузионного движения калия наружу. Эта точка равновесия представляет собой равновесный потенциал. как чистый трансмембранный поток (или ток ) K + равен нулю. Для хорошего приближения равновесного потенциала данного иона нужны только концентрации по обе стороны мембраны и температура. Его можно рассчитать с помощью уравнения Нернста :
![{\displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}\ln {\frac {[K^{+}]_{o}}{[K^{+}] _{я}}},}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b7eb1afcc5d7a5032ff24be5c403b43b1307b01e)
где
- Е экв,К + - равновесный потенциал калия, измеряемый в вольтах.
- R — универсальная газовая постоянная , равная 8,314 джоуля ·К. −1 ·моль −1
- T — абсолютная температура , измеряемая в кельвинах (= K = градусы Цельсия + 273,15).
- z – число элементарных зарядов рассматриваемого иона, участвующего в реакции
- F — постоянная Фарадея , равная 96 485 кулон ·моль. −1 или СП −1 ·моль −1
- [К + ] o — внеклеточная концентрация калия, измеряемая в моль ·м −3 или ммоль·л −1
- [К + ] i также представляет собой внутриклеточную концентрацию калия
Обычно равновесные потенциалы калия составляют около -80 милливольт (внутри отрицательный). Различия наблюдаются у разных видов, в разных тканях одного и того же животного и в одних и тех же тканях в разных условиях среды. Применяя приведенное выше уравнение Нернста, можно объяснить эти различия изменениями относительного K + концентрации или разницы в температуре.
Для общего использования уравнение Нернста часто дается в упрощенной форме, предполагая типичную температуру человеческого тела (37 ° C), уменьшая константы и переключаясь на логарифмическую систему с основанием 10. (Единицы, используемые для концентрации, не имеют значения, поскольку они будут сокращаться в соотношение). Для калия при нормальной температуре тела можно рассчитать равновесный потенциал в милливольтах как:
![{\displaystyle E_{eq,K^{+}}=61,54 мВ\log {\frac {[K^{+}]_{o}}{[K^{+}]_{i}}},}](https://wikimedia.org/api/rest_v1/media/math/render/svg/9d0b62ffda044574b1a52306113028c6a29c89f2)
Аналогично равновесный потенциал натрия ( Na + ) при нормальной температуре тела человека рассчитывается с использованием той же упрощенной константы. Вы можете рассчитать E, предполагая внешнюю концентрацию [K + ] o , 10 мМ и внутренней концентрацией, [K + ] i , 100 мМ. Для хлорид-ионов (Cl − ) знак константы необходимо поменять на противоположный (-61,54 мВ). Если рассчитать равновесный потенциал кальция (Ca 2+ ) заряд 2+ уменьшает упрощенную константу вдвое до 30,77 мВ. При работе при комнатной температуре, около 21 °C, расчетные константы составляют примерно 58 мВ для K. + и На + , −58 мВ для Cl − и 29 мВ для Ca 2+ . При физиологической температуре около 29,5 °C и физиологических концентрациях (которые различаются для каждого иона) расчетные потенциалы составляют примерно 67 мВ для Na. + , −90 мВ для K + , −86 мВ для Cl − и 123 мВ для Ca 2+ .
Потенциалы покоя
[ редактировать ]Мембранный потенциал покоя не является равновесным потенциалом, поскольку он зависит от постоянного расхода энергии (для ионных насосов , как упоминалось выше) для его поддержания. Именно динамический диффузионный потенциал учитывает этот механизм - в отличие от равновесного потенциала подушки, который верен независимо от природы рассматриваемой системы. В мембранном потенциале покоя доминируют те ионные виды системы, которые обладают наибольшей проводимостью через мембрану. Для большинства клеток это калий. Поскольку калий также является ионом с наиболее отрицательным равновесным потенциалом, обычно потенциал покоя не может быть более отрицательным, чем равновесный потенциал калия. Потенциал покоя можно рассчитать с помощью уравнения напряжения Гольдмана-Ходжкина-Каца, используя концентрации ионов в качестве равновесного потенциала, а также включая относительные проницаемости каждого вида ионов. В нормальных условиях можно с уверенностью предположить, что только калий, sodium (Na + ) и хлорид (Cl − ) ионы играют большую роль для потенциала покоя:
![{\displaystyle E_{m}={\frac {RT}{F}}\ln {\left({\frac {P_{Na^{+}}[Na^{+}]_{o}+P_{ K^{+}}[K^{+}]_{o}+P_{Cl^{-}}[Cl^{-}]_{i}}{P_{Na^{+}}[Na^ {+}]_{i}+P_{K^{+}}[K^{+}]_{i}+P_{Cl^{-}}[Cl^{-}]_{o}}} \верно)}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/aef443116c02ee0a29c6f4d6f5a5fc571e530ec8)
Это уравнение похоже на уравнение Нернста, но содержит член для каждого проникающего иона. Кроме того, z , в результате чего внутриклеточные и внеклеточные концентрации Cl в уравнение был добавлен − быть перевернутым относительно K + и На + , поскольку отрицательный заряд хлорида обрабатывается путем инвертирования дроби внутри логарифмического члена. * Em s — мембранный потенциал, измеренный в вольтах * — относительная проницаемость ионов s * [ R, T и F — как указано выше * P s ] Y — концентрация ионов s в отсеке Y, как указано выше. Другой способ оценить мембранный потенциал, рассматривая вместо этого проводимость ионных каналов, а не проницаемость мембраны, - использовать уравнение Миллмана (также называемое уравнением хордовой проводимости):

или переформулировать

где g tot — совокупная проводимость всех видов ионов, опять же в произвольных единицах. Последнее уравнение изображает мембранный потенциал покоя как средневзвешенное значение реверсивных потенциалов системы, где веса представляют собой относительную проводимость каждого вида ионов ( g X / g tot ). Во время потенциала действия эти веса меняются. Если проводимость Na + и Cl − равны нулю, мембранный потенциал сводится к потенциалу Нернста для K + (как г К + = г общий ). Обычно в состоянии покоя g Na+ и g Cl− не равны нулю, но они намного меньше, чем g K+ , что делает E m близким к E eq,K+ . Медицинские состояния, такие как гиперкалиемия , при которых крови калий сыворотки (который регулирует [K + ] o ) очень опасны, поскольку они смещают E eq,K+ , тем самым влияя Em на . Это может вызвать аритмию и остановку сердца . Использование болюсной инъекции хлорида калия при казнях посредством смертельной инъекции останавливает сердце, сдвигая потенциал покоя на более положительное значение, что деполяризует и окончательно сокращает сердечные клетки, не позволяя сердцу реполяризоваться и , таким образом, войти в диастолу для пополнения запасов. с кровью.
Хотя уравнение напряжения GHK и уравнение Миллмана связаны, они не эквивалентны. Критическое отличие состоит в том, что уравнение Миллмана предполагает, что соотношение ток-напряжение является омическим, тогда как уравнение напряжения GHK учитывает небольшие мгновенные исправления, предсказанные уравнением потока GHK, вызванные градиентом концентрации ионов. Таким образом, более точную оценку мембранного потенциала можно рассчитать с помощью уравнения GHK, чем с помощью уравнения Миллмана. [2]
Измерение потенциалов покоя
[ редактировать ]В некоторых клетках мембранный потенциал постоянно меняется (например, в клетках кардиостимулятора ). Для таких клеток никогда не существует «покоя», а «потенциал покоя» — это теоретическое понятие. Другие клетки с небольшими функциями мембранного транспорта, которые изменяются со временем, имеют мембранный потенциал покоя, который можно измерить, вставив в клетку электрод. [3] Трансмембранные потенциалы также можно измерить оптически с помощью красителей, которые меняют свои оптические свойства в зависимости от мембранного потенциала.
Сводная информация о значениях потенциала покоя в различных типах клеток
[ редактировать ]| Типы ячеек | Потенциал покоя |
|---|---|
| Клетки скелетных мышц | -95 мВ [4] |
| Астроглия | от -80 до -90 мВ |
| Нейроны | от -60 до -70 мВ [5] |
| Гладкие мышечные клетки | -60 мВ |
| Аорта Гладкая мышечная ткань | -45мВ [5] |
| Фоторецепторные клетки | -40 мВ |
| Волосковая клетка ( Улитка ) | от -15 до -40 мВ [6] |
| Эритроциты | -8,4 мВ [7] |
| Хондроциты | -8мВ [5] |
История
[ редактировать ]Токи покоя в нервах были измерены и описаны Юлиусом Бернштейном в 1902 году, когда он предложил «мембранную теорию», объясняющую потенциал покоя нервов и мышц как диффузионный потенциал. [8]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Пример эксперимента , электрофизиологического K демонстрирующего важность + для потенциала покоя. Зависимость потенциала покоя от внеклеточной концентрации К + показано на рис. 2.6 книги «Нейронаука» , 2-е издание, авторами Дейлом Первсом, Джорджем Дж. Августином, Дэвидом Фитцпатриком, Лоуренсом К. Кацем, Энтони-Сэмюэлем ЛаМантией, Джеймсом О. Макнамарой, С. Марком Уильямсом. Сандерленд (Массачусетс): Sinauer Associates, Inc.; 2001.
- ^ Хилле, Бертиль (2001) Ионные каналы возбудимых мембран, 3-е изд.
- ^ Иллюстрированный пример измерения мембранных потенциалов с помощью электродов приведен на рисунке 2.1 книги «Нейронаука» Дейла Первеса и др. (см. ссылку № 1 выше).
- ^ «Мышцы» . пользователи.rcn.com . 24 января 2015 г. Архивировано из оригинала 07.11.2015 . Проверено 1 июня 2016 г.
- ^ Перейти обратно: а б с Льюис, Ребекка; Асплин, Кэти Э.; Брюс, Гарет; Дарт, Кэролайн; Мобашери, Али; Барретт-Джолли, Ричард (1 ноября 2011 г.). «Роль мембранного потенциала в регуляции объема хондроцитов» . Журнал клеточной физиологии . 226 (11): 2979–2986. дои : 10.1002/jcp.22646 . ISSN 1097-4652 . ПМЦ 3229839 . ПМИД 21328349 .
- ^ Эшмор, Дж. Ф.; Мич, RW (24 июля 1986 г.). «Ионная основа мембранного потенциала наружных волосковых клеток улитки морской свинки». Природа . 322 (6077): 368–371. Бибкод : 1986Natur.322..368A . дои : 10.1038/322368a0 . ПМИД 2426595 . S2CID 4371640 .
- ^ Ченг, К; Хаспел, ХК; Валлано, ML; Осотимехин, Б; Зоненберг, М. (1980). «Измерение мембранных потенциалов (пси) эритроцитов и белых адипоцитов по накоплению катиона трифенилметилфосфония». Дж. Член. Биол . 56 (3): 191–201. дои : 10.1007/bf01869476 . ПМИД 6779011 . S2CID 19693916 .
- ^ Зайфарт, Эрнст-Август (1 января 2006 г.). «Юлиус Бернштейн (1839-1917): пионер нейробиолог и биофизик». Биологическая кибернетика . 94 (1): 2–8. дои : 10.1007/s00422-005-0031-y . ISSN 0340-1200 . ПМИД 16341542 . S2CID 2842501 .
Внешние ссылки
[ редактировать ]- Нейронаука - онлайн-учебник Первеса и др.
- нейрохимии , Сигел и др. Основные молекулярные, клеточные и медицинские аспекты
- Бертил Хилле Ионные каналы возбудимых мембран , 3-е изд., Sinauer Associates, Сандерленд, Массачусетс (2001). ISBN 0-87893-321-2
- Райт, С.Х. (2004). «Генерация мембранного потенциала покоя». Adv Physiol Educ . 28 (1–4): 139–42. дои : 10.1152/advan.00029.2004 . ПМИД 15545342 . S2CID 5009629 .
- Мембранный потенциал покоя - конспекты онлайн-лекций о мембранном потенциале покоя
- Происхождение потенциала мембран покоя - интерактивное онлайн-руководство