Анкирин повтор
Эта статья , возможно, содержит оригинальные исследования . ( Июль 2015 г. ) |
| Повторяющийся домен Анкирина | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | Анк | ||
| Пфам | PF00023 | ||
| ИнтерПро | ИПР002110 | ||
| УМНЫЙ | SM00248 | ||
| PROSITE | PDOC50088 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1awc / SCOPe / СУПФАМ | ||
| |||
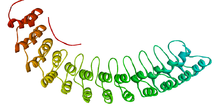
Анкириновый повтор из 33 остатков представляет собой мотив в белках, состоящий из двух альфа-спиралей, разделенных петлями , впервые обнаруженный в сигнальных белках у дрожжей Cdc10 и Drosophila Notch . Домены, состоящие из анкириновых тандемных повторов, опосредуют белок-белковые взаимодействия и являются одними из наиболее распространенных структурных мотивов в известных белках. Они появляются в бактериальных , архейных и эукариотических белках, но гораздо чаще встречаются у эукариот. Белки с анкириновыми повторами, хотя и отсутствуют в большинстве вирусов, широко распространены среди поксвирусов . Большинство белков, содержащих этот мотив, имеют от четырех до шести повторов, хотя его тезка анкирин содержит 24, а наибольшее известное количество повторов составляет 34, что предсказано в белке, экспрессируемом Giardia lamblia . [ 2 ]
Анкириновые повторы обычно складываются вместе, образуя единую линейную соленоидную структуру, называемую доменами повторов анкирина . Эти домены являются одной из наиболее распространенных платформ межбелкового взаимодействия в природе. Они встречаются в большом количестве функционально разнообразных белков, преимущественно эукариот . Несколько известных примеров прокариот и вирусов могут быть результатом горизонтального переноса генов. [ 3 ] Повтор был обнаружен в белках с разнообразными функциями, таких как инициаторы транскрипции, регуляторы клеточного цикла , цитоскелеты , переносчики ионов и преобразователи сигналов . Анкириновая складка, по-видимому, определяется ее структурой, а не функцией, поскольку не существует конкретной последовательности или структуры, которая была бы для нее общепризнанной.
Учитывая атомную структуру отдельных анкириновых повторов, петля часто представляет собой петлю бета-выпуклости типа 1 , тогда как обе альфа-спирали обычно имеют петлю Шеллмана на своем N-конце .
Роль в сворачивании белка
[ редактировать ]Мотив последовательности анкириновых повторов был изучен с использованием множественного выравнивания последовательностей для определения консервативных аминокислотных остатков, имеющих решающее значение для сворачивания и стабильности. Остатки на широкой латеральной поверхности повторяющихся структур анкирина вариабельны, часто гидрофобны и участвуют в основном в обеспечении белок-белковых взаимодействий. дизайн искусственного белка, основанный на консенсусной последовательности, Был синтезирован полученной в результате выравнивания последовательностей, и было обнаружено, что он стабильно сворачивается , представляя собой первый разработанный белок с множественными повторами. [ 4 ] В более обширных стратегиях проектирования использовались комбинаторные последовательности для «развития» анкириновых повторов, которые распознают определенные белковые мишени, метод, который был представлен в качестве альтернативы дизайну антител для приложений, требующих высокоаффинного связывания. [ 5 ] Структурное исследование с участием ряда анкириновых белков известных структур показывает, что анкириновые белки, основанные на консенсусе, очень стабильны, поскольку они максимизируют энергетический разрыв между структурами сворачивания и разворачивания, кодируя плотно связанную сеть благоприятных взаимодействий между мотивами консервативных последовательностей. , как мотив TPLX. [ 6 ] То же исследование показывает, что вставки в каноническую структуру анкириновых повторов обогащены конфликтными взаимодействиями, связанными с функцией. То же самое относится и к взаимодействиям вокруг горячих точек удаления. Это может быть связано со сложными переходами складывания/раскладывания, которые важны для распознавания и взаимодействия партнера.
Белки с анкириновыми повторами представляют собой необычную проблему при изучении сворачивания белков , которое в основном сосредоточено на глобулярных белках , которые образуют четко определенную третичную структуру, стабилизированную дальними, нелокальными контактами остаток-остаток . Анкириновые повторы, напротив, содержат очень мало таких контактов (т. е. имеют низкий порядок контактов ). Большинство исследований показали, что анкириновые повторы сворачиваются по механизму сворачивания в двух состояниях , что предполагает высокую степень кооперативности сворачивания, несмотря на локальные контакты между остатками и очевидную необходимость успешного сворачивания с различным количеством повторов. Некоторые данные, основанные на синтезе усеченных версий белков с природными повторами, [ 7 ] и при проверке значений Фи , [ 8 ] предполагает, что С-конец образует центр складчатой нуклеации.
Клиническое значение
[ редактировать ]Белки с анкириновыми повторами связаны с рядом заболеваний человека . Эти белки включают клеточного цикла ингибитор p16 , который связан с раком , и белок Notch (ключевой компонент клеточных сигнальных путей), который может вызывать неврологическое расстройство CADASIL, когда повторяющийся домен разрушается в результате мутаций. [ 2 ]
Специализированное семейство белков анкирина, известное как белки повторов анкирина в мышцах (MARP), участвует в восстановлении и регенерации мышечной ткани после повреждения, вызванного травмой и стрессом. [ 9 ]
Естественная вариация между глутамином и лизином в положении 703 в 11-м анкириновом повторе ANKK1 , известная как аллель TaqI A1, [ 10 ] ему приписывают поощрение аддиктивного поведения, такого как ожирение, алкоголизм, никотиновая зависимость и стиль любви Эроса. [ нужна ссылка ] одновременно препятствуя подростковой преступности и невротизму-тревоге. [ 11 ] [ не удалось пройти проверку ] Это изменение может повлиять на специфичность белковых взаимодействий, осуществляемых протеинкиназой ANKK1 посредством этого повтора. [ нужна ссылка ] .
Белки человека, содержащие этот повтор
[ редактировать ]АБТБ1 ; АБТБ2 ; ACBD6 ; АКТБЛ1 ; АНК1 ; АНК2 ; АНК3 ; ЯКОРЬ ; АНКДД1А ; АНКЕФ1 ; АНКФИ1 ; АНКХД1 ; АНКИБ1 ; АНКК1 ; АНКМИ1 ; АНКМИ2 ; ЯКОР2 ; АНКРД1 ; АНКРД10 ; АНКРД11 ; АНКРД12 ; АНКРД13 ; АНКРД13А ; АНКРД13Б ; АНКРД13С ; АНКРД13Д ; АНКРД15 ; АНКРД16 ; АНКРД17 ; АНКРД18А ; АНКРД18Б ; АНКРД19 ; АНКРД2 ; АНКРД20А1 ; АНКРД20А2 ; АНКРД20А3 ; АНКРД20А4 ; АНКРД21 ; АНКРД22 ; АНКРД23 ; АНКРД24 ; АНКРД25 ; АНКРД26 ; АНКРД27 ; АНКРД28 ; АНКРД30А ; АНКРД30Б ; АНКРД30БЛ ; АНКРД32 ; АНКРД33 ; АНКРД35 ; АНКРД36 ; АНКРД36Б ; АНКРД37 ; АНКРД38 ; АНКРД39 ; АНКРД40 ; АНКРД41 ; АНКРД42 ; АНКРД43 ; АНКРД44 ; АНКРД45 ; АНКРД46 ; АНКРД47 ; ANKRD49 ; АНКРД50 ; АНКРД52 ; АНКРД53 ; АНКРД54 ; АНКРД55 ; АНКРД56 ; АНКРД57 ; АНКРД58 ; АНКРД60 ; АНКРД6 ; АНКРД7 ; АНКРД9 ; АНКС1А ; АНКС3 ; АНКС4Б ; АНКС6 ; АНКЗФ1 ; АСБ1 ; АСБ10 ; АСБ11 ; АСБ12 ; АСБ13 ; АСБ14 ; АСБ15 ; АСБ16 ; АСБ2 ; АСБ3 ; АСБ4 ; АСБ5 ; АСБ6 ; АСБ7 ; АСБ8 ; АСБ9 ; АСЗ1 ; БАРД1 ; БАТ4 ; БАТ8 ; БКЛ3 ; БЦОР ; БКОРЛ1 ; БТБД11 ; КАМТА1 ; КАМТА2 ; БОЧКА1 ; БОЧКА2 ; СКМ1 ; CDKN2A ; CDKN2B ; CDKN2C ; CDKN2D ; ЦЕНТБ1 ; ЦЕНТБ2 ; ЦЕНТБ5 ; ЦЕНТГ1 ; ЦЕНТГ2 ; ЦЕНТГ3 ; КЛИП3 ; КЛИП4 ; КЛПБ ; CTGLF1 ; CTGLF2 ; CTGLF3 ; CTGLF4 ; CTGLF5 ; CTTNBP2 ; ДАПК1 ; ДДЕФ1 ; ДДЕФ2 ; ДДЕФЛ1 ; ДГКИ ; ДГКЗ ; ДП58 ; ДИСФИП1 ; ДЗАНК ; ЭХМТ1 ; ЭХМТ2 ; ЭСПН ; ФАНК1 ; ФЕМ1А ; ФЕМ1Б ; ГАБПБ2 ; ГИТ1 ; ГИТ2 ; ГЛС ; ГЛС2 ; ХАСЕ1 ; ГЕКТД1 ; ИБТК ; ИЛК ; ИНВС ; КИДИНС220 ; КРИТ1 ; ЛРРК1 ; ПОЧТА ; МИБ1 ; МИБ2 ; МФОСФ8 ; МТПН ; МИО16 ; НФКБ1 ; НФКБ2 ; НФКБИА ; НФКБИБ ; НФКБИ ; НФКБИЛ1 ; НФКБИЛ2 ; ВЫРЕЗ1 ; ВЫРЕЗКА2 ; ВЫРЕЗКА3 ; ВЫРЕЗКА4 ; НРАРП ; НУДТ12 ; ОСБПЛ1А ; ОСТФ1 ; ПЛА2Г6 ; ПОТЕ14 ; ПОТЕ15 ; ПОТЕ8 ; ППП1Р12А ; ППП1Р12Б ; ППП1Р12С ; ППП1Р13Б ; ППП1Р13Л ; ППП1Р16А ; ППП1Р16Б ; ПСМД10 ; РАИ14 ; RFXANK ; РИПК4 ; РНАСЕЛ ; ХВОСТОВИК1 ; ХВОСТОВИК2 ; ШАНК3 ; СНКАИП ; ТА-НФКБХ ; ТЭК14 ; ТНКС ; ТНКС2 ; ТННИ3К ; ТП53ВР2 ; ТРП7 ; ТРПА1 ; ТРПК3 ; ТРПК4 ; ТРПК5 ; ТРПК6 ; ТРПК7 ; ТРПВ1 ; ТРПВ2 ; ТРПВ3 ; ТРПВ4 ; ТРПВ5 ; ТРПВ6 ; УАКА ; УШ1Г ; ЗДХХК13 ; ЗДХХК17 ;
См. также
[ редактировать ]- DARPin (созданный белок с повторами анкирина), сконструированный миметик антител, основанный на структуре повторов анкирина.
Ссылки
[ редактировать ]- ^ ПДБ : 1N11 ; Майкли П., Томчик Д.Р., Мачиус М., Андерсон Р.Г. (декабрь 2002 г.). «Кристаллическая структура стека из 12 повторов ANK человека ANK1» . ЭМБО Дж . 21 (23): 6387–96. дои : 10.1093/emboj/cdf651 . ПМК 136955 . ПМИД 12456646 .
- ^ Jump up to: а б Мосави Л., Каммет Т., Дерозье Д., Пэн З. (2004). «Анкириновый повтор как молекулярная архитектура распознавания белков» . Белковая наука . 13 (6): 1435–48. дои : 10.1110/ps.03554604 . ПМК 2279977 . ПМИД 15152081 . Архивировано из оригинала 7 сентября 2004 г.
- ^ Борк П. (декабрь 1993 г.). «Сотни анкириноподобных повторов в функционально разнообразных белках: мобильные модули, которые пересекают типы горизонтально?». Белки . 17 (4): 363–74. дои : 10.1002/прот.340170405 . ПМИД 8108379 . S2CID 35224626 .
- ^ Мосави Л.К., Минор Д.Л., Пэн З.Ю. (декабрь 2002 г.). «Структурные детерминанты повторного мотива анкирина, полученные на основе консенсуса» . Proc Natl Acad Sci США . 99 (25): 16029–34. Бибкод : 2002PNAS...9916029M . дои : 10.1073/pnas.252537899 . ПМК 138559 . ПМИД 12461176 .
- ^ Бинц Х.К., Амштутц П., Коль А. и др. (май 2004 г.). «Высокоаффинные связыватели, выбранные из разработанных библиотек белков с повторами анкирина». Нат. Биотехнология . 22 (5): 575–82. дои : 10.1038/nbt962 . ПМИД 15097997 . S2CID 1191035 .
- ^ Парра Р.Г., Эспада Р., Верстраете Н., Феррейро Д.У. и др. (декабрь 2015 г.). «Структурная и энергетическая характеристика семейства белков с повторами анкирина» . ПЛОС Компьютер. Биол . 12 (11): 575–82. Бибкод : 2015PLSCB..11E4659P . дои : 10.1371/journal.pcbi.1004659 . ПМК 4687027 . ПМИД 26691182 .
- ^ Чжан Б., Пэн Цзи (июнь 2000 г.). «Минимальная единица сворачивания в белке анкириновых повторов p16 (INK4)». Дж Мол Биол . 299 (4): 1121–32. дои : 10.1006/jmbi.2000.3803 . ПМИД 10843863 .
- ^ Тан К.С., Фершт А.Р., Ицхаки Л.С. (январь 2003 г.). «Последовательное развертывание анкириновых повторов в супрессоре опухоли p16» . Структура . 11 (1): 67–73. дои : 10.1016/S0969-2126(02)00929-2 . ПМИД 12517341 .
- ^ Миллер М.К., Банг М.Л., Витт CC и др. (ноябрь 2003 г.). «Белки с повторами анкирина в мышцах: CARP, ankrd2 / Arpp и DARP как семейство молекул реакции на стресс на основе титиновых нитей». Дж Мол Биол . 333 (5): 951–64. дои : 10.1016/j.jmb.2003.09.012 . ПМИД 14583192 .
- ^ Невилл М.Дж., Джонстон Э.К., Уолтон Р.Т. (июнь 2004 г.). «Идентификация и характеристика ANKK1: новый ген киназы, тесно связанный с DRD2 на участке хромосомы 11q23.1» . Хм. Мутат . 23 (6): 540–5. дои : 10.1002/humu.20039 . ПМИД 15146457 . S2CID 22242611 .
- ^ «Сводка генов NCBI для DRD2» . (временная ссылка)
Внешние ссылки
[ редактировать ]- ресурсов Eukaryotic Linear Motif Класс мотивов LIG_TNKBM_1
- Анкирин + повтор в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)