Гомология (биология)

В биологии — это сходство , гомология обусловленное общим происхождением пары структур или генов в разных таксонах . Типичным примером гомологичных структур являются передние конечности позвоночных животных , где крылья летучих мышей и птиц , руки приматов , передние ласты китов и передние ноги четвероногих позвоночных, таких как собаки и крокодилы, произошли от одного и того же предка. строение тетрапода . Эволюционная биология объясняет гомологичные структуры, приспособленные для разных целей, результатом происхождения с модификациями от общего предка . Этот термин был впервые применен к биологии в неэволюционном контексте анатомом Ричардом Оуэном в 1843 году. Позднее гомология была объяснена теорией эволюции Чарльза Дарвина в 1859 году, но наблюдалась и раньше, начиная с Аристотеля , и это было подробно проанализирован Пьером Белоном в 1555 году.
В биологии развития органы, которые развились у эмбриона одинаковым образом и из схожего происхождения, например, из совпадающих зачатков в последовательных сегментах одного и того же животного, являются серийно гомологичными . Примеры включают ноги многоножки , верхнечелюстные щупики и губные щупики насекомого, а также остистые отростки последовательных позвонков в позвоночном столбе . Мужские и женские репродуктивные органы гомологичны, если они развиваются из одной и той же эмбриональной ткани, как и яичники и яички млекопитающих, включая человека. [ нужна ссылка ]
Гомология последовательностей между последовательностями белков или ДНК определяется аналогичным образом с точки зрения общего происхождения. Два сегмента ДНК могут иметь общее происхождение либо в результате события видообразования ( ортологи ), либо в результате события дупликации ( паралоги ). Гомология белков или ДНК определяется сходством их последовательностей. Значительное сходство является убедительным доказательством того, что две последовательности связаны дивергентной эволюцией от общего предка. Выравнивание нескольких последовательностей используется для обнаружения гомологичных областей.
остается спорной Гомология в поведении животных , но есть убедительные доказательства того, что, например, иерархии доминирования гомологичны у приматов .
История
[ редактировать ]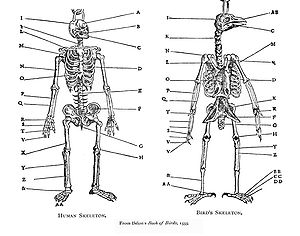
Гомологию заметил Аристотель (ок. 350 г. до н.э.), [ 1 ] и был подробно проанализирован Пьером Белоном в его «Книге птиц» 1555 года , где он систематически сравнивал скелеты птиц и людей. Модель сходства интерпретировалась как часть статичной великой цепи бытия в период Средневековья и раннего Нового времени: тогда она не рассматривалась как подразумевающая эволюционные изменения. В немецкой традиции натурфилософии гомология представляла особый интерес как демонстрация единства в природе. [ 2 ] [ 3 ] В 1790 году Гете изложил свою лиственную теорию в своем эссе «Метаморфозы растений», показав, что части цветка происходят из листьев. [ 4 ] Серийная гомология конечностей была описана в конце XVIII века. Французский зоолог Этьен Жоффруа Сен-Илер показал , что структуры являются общими для рыб, рептилий, птиц и млекопитающих. в 1818 году в своей теории аналогов («теории гомологов») [ 5 ] Когда Жоффруа пошел дальше и стал искать сходство между Жоржа Кювье , ответвлениями такими как позвоночные и моллюски, его утверждения вызвали дебаты 1830 года Кювье-Жоффруа . Жоффруа сформулировал принцип связей, а именно то, что важно взаимное расположение различных структур и их связи друг с другом. [ 3 ] Эмбриолог Карл Эрнст фон Бэер в 1828 году сформулировал то, что сейчас называется законами фон Бэра , отметив, что родственные животные начинают свое развитие как сходные эмбрионы, а затем расходятся: таким образом, животные в одном семействе более тесно связаны и расходятся позже, чем животные, находящиеся только в того же порядка и имеют меньше гомологий. Теория фон Бэера признает, что каждый таксон (например, семейство) имеет отличительные общие черты и что эмбриональное развитие соответствует таксономической иерархии: это не то же самое, что теория рекапитуляции . [ 3 ] Термин «гомология» впервые был использован в биологии анатомом Ричардом Оуэном в 1843 году при изучении сходства плавников и конечностей позвоночных, определив его как «один и тот же орган у разных животных при любом разнообразии форм и функций». [ 6 ] и противопоставил его соответствующему термину «аналогия», который он использовал для описания различных структур с той же функцией. Оуэн систематизировал три основных критерия определения гомологичности признаков: положение, развитие и состав. В 1859 году Чарльз Дарвин объяснил гомологичные структуры как означающие, что соответствующие организмы разделяют строение тела от общего предка и что таксоны являются ветвями единого древа жизни . [ 2 ] [ 7 ] [ 3 ]
Определение
[ редактировать ]


Слово гомология, придуманное примерно в 1656 году, происходит от греческого ὁμόλογος homologos от ὁμός homos «тот же» и λόγος logos «отношение». [ 8 ] [ 9 ] [ а ]
Сходные биологические структуры или последовательности в разных таксонах гомологичны, если они произошли от общего предка . Таким образом, гомология подразумевает дивергентную эволюцию . Например, многие насекомые (например, стрекозы ) обладают двумя парами летающих крыльев . У жуков первая пара крыльев превратилась в пару твёрдых надкрыльев . [ 12 ] в то время как у двукрылых вторая пара крыльев превратилась в небольшие жужжальца, используемые для равновесия. [ б ] [ 13 ]
Точно так же передние конечности предков позвоночных эволюционировали в передние ласты китов , крылья птиц бегущие передние ноги собак , оленей и лошадей , короткие передние ноги лягушек и ящериц и хватательные руки приматов , , включая человека. Одни и те же основные кости предплечья ( плечевая , лучевая и локтевая кости). [ с ] ) встречаются в окаменелостях лопастноперых рыб, таких как Eusthenopteron . [ 14 ]
Гомология против аналогии
[ редактировать ]
Противоположностью гомологичных органов являются аналогичные органы, выполняющие сходную работу у двух таксонов, которые не присутствовали у их самого недавнего общего предка , а скорее развились отдельно . Например, крылья насекомых и птиц развивались независимо в широко разделенных группах и функционально сходились, чтобы обеспечить полет с помощью двигателя , поэтому они аналогичны. Точно так же крылья семя клена платана и крылья птицы аналогичны, но не гомологичны, поскольку развиваются из совершенно разных структур. [ 15 ] [ 16 ] Структура может быть гомологичной на одном уровне, но аналогичной только на другом. Крылья птерозавра , птицы и летучей мыши аналогичны крыльям, но гомологичны передним конечностям, поскольку этот орган служил предплечьем (а не крылом) у последнего общего предка четвероногих и развивался по-разному в трех группах. Таким образом, у птерозавров «крыло» включает в себя как переднюю, так и заднюю конечности. [ 17 ] аналогия называется гомоплазией В кладистике , а в эволюционной биологии — конвергентной или параллельной эволюцией . [ 18 ] [ 19 ]
В кладистике
[ редактировать ]В таксономических исследованиях используются специализированные термины. Первичная гомология - это первоначальная гипотеза исследователя, основанная на схожей структуре или анатомических связях, предполагающая, что состояние признака у двух или более таксонов является общим из-за общего происхождения. Первичную гомологию можно концептуально разбить дальше: мы можем рассматривать все состояния одного и того же характера как «гомологичные» части одной, неопределённой серии трансформаций. Это было названо топографической корреспонденцией. Например, в выровненной матрице последовательности ДНК все A, G, C, T или подразумеваемые пробелы в данном нуклеотидном сайте гомологичны таким образом. Идентичность состояния признака — это гипотеза о том, что конкретное состояние в двух или более таксонах «одинаково» с точки зрения нашей схемы кодирования признаков. Таким образом, предполагается, что два аденина в одном и том же выровненном нуклеотидном сайте гомологичны, если эта гипотеза впоследствии не будет опровергнута другими доказательствами. Вторичная гомология подразумевается экономным анализом. , где гомологичным считается состояние символа, возникающее на дереве только один раз. [ 20 ] [ 21 ] Как подразумевается в этом определении, многие кладисты считают вторичную гомологию синонимом синапоморфии , общего производного признака или состояния черты , которое отличает кладу от других организмов. [ 22 ] [ 23 ] [ 24 ]
Общие состояния наследственных признаков, симплезиоморфии, представляют собой либо синапоморфии более инклюзивной группы, либо дополнительные состояния (часто отсутствие), которые не объединяют ни одну естественную группу организмов. Например, наличие крыльев является синапоморфией для крыловидных насекомых и симплезиоморфией для голометаболических насекомых. Отсутствие крыльев у некрылокрылых насекомых и других организмов является дополнительной симплезиоморфией, не объединяющей ни одну группу (например, отсутствие крыльев не свидетельствует об общем происхождении чешуйниц, пауков и кольчатых червей). С другой стороны, отсутствие (или вторичная потеря) крыльев является синапоморфией для блох. Подобные закономерности заставляют многих кладистов считать концепции гомологии и концепции синапоморфии эквивалентными. [ 25 ] [ 24 ] Некоторые кладисты следуют докладистическому определению гомологии Хааса и Симпсона, [ 26 ] и рассматривать как синапоморфии, так и симплезиоморфии как гомологичные состояния характера. [ 27 ]
В разных таксонах
[ редактировать ]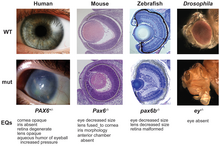
Гомологии обеспечивают фундаментальную основу для всей биологической классификации, хотя некоторые из них могут быть весьма противоречивыми. Например, глубокие гомологии , такие как гены pax6 , которые контролируют развитие глаз позвоночных и членистоногих, были неожиданными, поскольку органы анатомически различны и, по-видимому, развивались совершенно независимо. [ 28 ] [ 29 ]
У членистоногих
[ редактировать ]Эмбриональные сегменты тела ( сомиты ) разных таксонов членистоногих разошлись от простого строения тела со многими сходными придатками, которые серийно гомологичны, к множеству строений тела с меньшим количеством сегментов, снабженных специализированными придатками. [ 30 ] Гомологии между ними были обнаружены путем сравнения генов в эволюционной биологии развития . [ 28 ]

| Сомит (тело сегмент) |
Трилобит ( Трилобитоморфа ) 
|
Паук ( Хелицерата ) 
|
Многоножки ( Многоножек ) |
Насекомое ( Шестиногий ) 
|
Креветка ( Ракообразные ) 
|
|---|---|---|---|---|---|
| 1 | усики | хелицеры (челюсти и клыки) | усики | усики | 1-е усики |
| 2 | первые ноги | педипальпы | - | - | 2-е усики |
| 3 | 2-е ноги | первые ноги | челюсти | челюсти | нижние челюсти (челюсти) |
| 4 | третьи ноги | 2-е ноги | 1- я щека | 1-я щека | 1-я щека |
| 5 | 4-е ноги | третьи ноги | 2-я щека | 2-я щека | 2-я щека |
| 6 | 5-й этап | 4-е ноги | шея (без ног) | первые ноги | первые ноги |
| 7 | 6-й этап | - | первые ноги | 2-е ноги | 2-е ноги |
| 8 | 7-й этап | - | 2-е ноги | третьи ноги | третьи ноги |
| 9 | 8-й этап | - | третьи ноги | - | 4-е ноги |
| 10 | 9-й этап | - | 4-е ноги | - | 5-й этап |
Среди насекомых жало самки медоносной пчелы представляет собой модифицированный яйцеклад , гомологичный яйцекладу других насекомых, таких как прямокрылые , полужесткокрылые и перепончатокрылые без жала. [ 31 ]
У млекопитающих
[ редактировать ]Три маленькие косточки среднего уха млекопитающих, включая человека, молоточек , наковальня и стремечко , сегодня используются для передачи звука от барабанной перепонки во внутреннее ухо . Молоточек и наковальня развиваются у эмбриона из структур, формирующих челюстные кости (квадратную и суставную), у ящериц и окаменелостей ящерицеобразных предков млекопитающих. Обе линии доказательств показывают, что эти кости гомологичны и имеют общего предка. [ 32 ]
Среди многих гомологий репродуктивных систем млекопитающих гомологичны и яички . яичники [ 33 ]
Рудиментарные органы, такие как копчик человека , в настоящее время значительно уменьшенные в своем функциональном состоянии, легко воспринимаются как признаки эволюции ; объяснение состоит в том, что они были сокращены естественным отбором из функционирующих органов, когда их функции больше не были нужны, но не имеют никакого смысла. вообще, если виды считаются фиксированными. Копчик гомологичен хвостам других приматов. [ 34 ]
В растениях
[ редактировать ]Листья, стебли и корни
[ редактировать ]У многих растений защитные или накопительные структуры образуются в результате изменений в развитии первичных листьев , стеблей и корней . Листья по-разному модифицируются из фотосинтетических структур, образуя кувшины для ловли насекомых у кувшинчатых растений , челюсти для ловли насекомых у венериной мухоловки и колючки кактусов , причем все они гомологичны. [ 35 ]
| Первичные органы | Оборонительные сооружения | Структуры хранения |
|---|---|---|
| Листья | Шипы | Опухшие листья (например, у суккулентов ) |
| Стебли | Шипы | Клубни (например, картофеля ), корневища (например, имбиря ), мясистые стебли (например, кактусов ) |
| Корни | - | Корневые клубни (например, сладкий картофель ), стержневой корень (например, морковь ) |
Некоторые сложные листья цветковых растений частично гомологичны как листьям, так и побегам, поскольку их развитие произошло на основе генетической мозаики развития листьев и побегов. [ 36 ] [ 37 ]
-
 Один перистый лист ясеня европейского.
Один перистый лист ясеня европейского. -
 Деталь пальмового листа
Деталь пальмового листа -

-
 Очень большие листья банана Musa acuminata.
Очень большие листья банана Musa acuminata. -
 Сочный запасающий воду. лист алоэ,
Сочный запасающий воду. лист алоэ, -

-

-









Части цветка
[ редактировать ]
Четыре типа частей цветка, а именно плодолистики , тычинки , лепестки и чашелистики , гомологичны листьям и происходят из них, как правильно заметил Гете в 1790 году. Развитие этих частей происходит посредством паттерна экспрессии генов в зонах роста ( меристемах ). описывается ABC-моделью развития цветка . Каждый из четырех типов частей цветка последовательно повторяется в концентрических мутовках, контролируемых небольшим количеством генов, действующих в различных комбинациях. Таким образом, гены А, работающие отдельно, приводят к образованию чашелистиков; A и B вместе образуют лепестки; B и C вместе образуют тычинки; Только C производит плодолистики. Когда ни один из генов не активен, образуются листья. Еще две группы генов: D для формирования семяпочек и E для цветочных мутовок, завершают модель. Гены, очевидно, древние, столь же древние, как и сами цветковые растения . [ 4 ]
Биология развития
[ редактировать ]
Биология развития может идентифицировать гомологичные структуры, возникшие из одной и той же ткани в эмбриогенезе . Например, у взрослых змей нет ног, но у их ранних эмбрионов есть зачатки задних ног, которые вскоре теряются по мере развития эмбрионов. Предположение о том, что у предков змей были задние ноги, подтверждается ископаемыми свидетельствами: у меловой змеи Pachyrhachispromaticus задние ноги были в комплекте с бедренными костями ( подвздошная , лобковая , седалищная кость ), бедренной костью ( бедренная кость ), костями ног ( большая берцовая кость , малоберцовая кость ) и кости стопы ( пяточная кость , таранная кость ), как у современных четвероногих, с ногами. [ 38 ]
Гомология последовательностей
[ редактировать ]
Как и в случае с анатомическими структурами, гомология последовательностей между последовательностями белков или ДНК определяется с точки зрения общего происхождения. Два сегмента ДНК могут иметь общее происхождение либо в результате события видообразования ( ортологи ), либо в результате события дупликации ( паралоги ). Гомологию белков или ДНК обычно определяют по сходству их последовательностей. Значительное сходство является убедительным доказательством того, что две последовательности связаны дивергентной эволюцией общего предка. Выравнивание нескольких последовательностей используется для указания того, какие области каждой последовательности являются гомологичными. [ 40 ]
Гомологичные последовательности являются ортологичными, если они произошли от одной и той же предковой последовательности, разделенной событием видообразования : когда вид расходится на два отдельных вида, копии одного гена в двух полученных видах называются ортологичными . Термин «ортолог» был придуман в 1970 году молекулярным эволюционистом Уолтером Фитчем . [ 41 ]
Гомологичные последовательности являются паралогичными, если они были созданы в результате дупликации внутри генома. Что касается событий дупликации гена , если ген в организме дублируется, две копии являются паралогичными. Они могут формировать структуру целых геномов и, таким образом, в значительной степени объяснять эволюцию генома. Примеры включают гены гомеобокса ( Hox ) у животных. Эти гены претерпели не только дупликации генов внутри хромосом , но и дупликации всего генома . В результате Hox-гены у большинства позвоночных распределены по множеству хромосом: кластеры HoxA-D изучены лучше всего. [ 42 ]
Некоторые последовательности гомологичны, но они разошлись настолько, что сходство их последовательностей недостаточно для установления гомологии. Однако многие белки сохранили очень схожие структуры, и структурное выравнивание можно использовать для демонстрации их гомологии. [ 43 ]
В поведении
[ редактировать ]Было высказано предположение, что некоторые виды поведения могут быть гомологичными, основанными либо на общем отношении между родственными таксонами, либо на общем происхождении поведения в развитии человека; однако понятие гомологичного поведения остается спорным, [ 44 ] во многом потому, что поведение более склонно к множественной реализации, чем другие биологические черты. Например, Д. У. Раецки и Рэндалл К. Флэнери, используя данные о людях и приматах , утверждают, что модели поведения в иерархиях доминирования гомологичны у всех приматов. [ 45 ]

Как и в случае с морфологическими особенностями или ДНК, общее сходство в поведении свидетельствует об общем происхождении. [ 46 ] Гипотеза о том, что поведенческий признак не гомологичен, должна основываться на неконгруэнтном распределении этого признака по отношению к другим чертам, которые, как предполагается, отражают истинную структуру отношений. Это приложение Вилли Хеннига. [ 47 ] вспомогательный принцип .
Примечания
[ редактировать ]- ^ Альтернативные термины «гомогенность» и «однородный» также использовались в конце 1800-х - начале 1900-х годов. Однако эти термины в настоящее время являются архаичными в биологии, и термин «гомогенный» теперь обычно встречается как неправильное написание термина « гомогенный », который относится к однородности смеси. [ 10 ] [ 11 ]
- ^ Если две пары крыльев рассматривать как взаимозаменяемые гомологичные структуры, это можно описать как параллельное уменьшение количества крыльев, но в противном случае каждое из этих двух изменений представляет собой расходящиеся изменения в одной паре крыльев.
- ^ На главном изображении они окрашены в цвет: плечевая кость коричневая, лучевая кость бледно-желтая, локтевая кость красная.
Ссылки
[ редактировать ]- ^ Jump up to: а б Панчен, Алабама (1999). «Гомология — история понятия». Симпозиум 222 Фонда Novartis — Гомология . Симпозиумы Фонда Новартис. Том. 222. стр. 5–18, обсуждение 18–23. дои : 10.1002/9780470515655.ch2 . ISBN 9780470515655 . ПМИД 10332750 .
- ^ Jump up to: а б Панчен, Алабама (1999). «Гомология — история понятия». Симпозиум 222 Фонда Novartis — Гомология . Симпозиумы Фонда Новартис. Том. 222. стр. 5–18. дои : 10.1002/9780470515655.ch2 . ISBN 9780470515655 . ПМИД 10332750 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б с д Бригандт, Инго (23 ноября 2011 г.). «Эссе: Гомология» . Энциклопедия проекта «Эмбрион» .
- ^ Jump up to: а б Дорнелас, Марсело Карнье; Дорнелас, Одаир (2005). «От листа к цветку: Возвращаясь к концепциям Гете о «метаморфозе» растений» . Бразильский журнал физиологии растений . 17 (4): 335–344. дои : 10.1590/S1677-04202005000400001 .
- ^ Жоффруа Сен-Илер, Этьен (1818). Анатомическая философия. Полет. 1: Органы дыхания в отношении определения и идентичности их костных частей . Полет. 1. Париж: Ж.Б. Байьер.
- ^ Оуэн, Ричард (1843). Лекции по сравнительной анатомии и физиологии беспозвоночных животных, прочитанные в Королевском колледже хирургов в 1843 году . Лонгман, Браун, Грин и Лонгманс. стр. 374, 379.
- ^ Соммер, Р.Дж. (июль 2008 г.). «Гомология и иерархия биологических систем». Биоэссе . 30 (7): 653–658. doi : 10.1002/bies.20776 . ПМИД 18536034 .
- ^ Бауэр, Фредерик Орпен (1906). «Морфология растений» . Конгресс искусств и науки: Универсальная выставка, Сент-Луис, 1904 год . Хоутон, Миффлин. п. 64.
- ^ Уильямс, Дэвид Малкольм; Фори, Питер Л. (2004). Вехи в систематике . ЦРК Пресс. п. 198 . ISBN 978-0-415-28032-7 .
- ^ «Однородный, прил.». ОЭД онлайн. Март 2016 г. Издательство Оксфордского университета. http://www.oed.com/view/Entry/88045 ? (по состоянию на 9 апреля 2016 г.).
- ^ «Однородный, прил.». ОЭД онлайн. Март 2016 г. Издательство Оксфордского университета. http://www.oed.com/view/Entry/88055 ? (по состоянию на 9 апреля 2016 г.).
- ^ Вагнер, Гюнтер П. (2014). Гомология, гены и эволюционные инновации . Издательство Принстонского университета. стр. 53–54. ISBN 978-1-4008-5146-1 .
надкрылья очень мало похожи на типичные крылья, но явно гомологичны передним крыльям. Следовательно, бабочки, мухи и жуки имеют две пары спинных придатков, гомологичных у разных видов.
- ^ Липшиц, Ховард Д. (2012). Гены, развитие и рак: жизнь и работа Эдварда Б. Льюиса . Спрингер. п. 240. ИСБН 978-1-4419-8981-9 .
Например, крыло и жужжальца являются гомологичными, но широко расходящимися органами, которые обычно возникают как дорсальные придатки второго грудного (Т2) и третьего грудного (Т3) сегментов соответственно.
- ^ «Гомология: ноги и конечности» . Калифорнийский университет в Беркли . Проверено 15 декабря 2016 г.
- ^ «Найден секрет полета «вертолетных семян» » . ЖиваяНаука. 11 июня 2009 года . Проверено 2 марта 2017 г.
- ^ Лентинк, Д.; Диксон, ВБ; ван Леувен, JL; Дикинсон, Миннесота (12 июня 2009 г.). «Передовые вихри увеличивают подъем семян автовращающихся растений» (PDF) . Наука . 324 (5933): 1438–1440. Бибкод : 2009Sci...324.1438L . дои : 10.1126/science.1174196 . ПМИД 19520959 . S2CID 12216605 .
- ^ Шотландия, RW (2010). «Глубокая гомология: взгляд из систематики». Биоэссе . 32 (5): 438–449. doi : 10.1002/bies.200900175 . ПМИД 20394064 . S2CID 205469918 .
- ^ См. Батлер, AB: Гомология и гомопластика. В: Сквайр, Ларри Р. (ред.): Энциклопедия неврологии , Academic Press, 2009, стр. 1195–1199.
- ^ «Гомологичная структура против аналогичной структуры: в чем разница?» . Проверено 27 сентября 2016 г.
- ^ де Пинна, MCC (1991). «Концепции и тесты гомологии в кладистической парадигме». Кладистика . 7 (4): 367–394. CiteSeerX 10.1.1.487.2259 . дои : 10.1111/j.1096-0031.1991.tb00045.x . S2CID 3551391 .
- ^ Брауэр, Эндрю В.З.; Шавароч, В. (1996). «Три шага оценки гомологии». Кладистика . 12 (3): 265–272. дои : 10.1111/j.1096-0031.1996.tb00014.x . ПМИД 34920625 . S2CID 85385271 .
- ^ Пейдж, Родерик DM; Холмс, Эдвард К. (2009). Молекулярная эволюция: филогенетический подход . Джон Уайли и сыновья . ISBN 978-1-4443-1336-9 .
- ^ Брауэр, Эндрю В.З.; де Пинна, Марио CC (24 мая 2012 г.). «Гомологии и ошибки» . Кладистика . 28 (5): 529–538. дои : 10.1111/j.1096-0031.2012.00398.x . PMID 34844384 . S2CID 86806203 .
- ^ Jump up to: а б Брауэр, Эндрю В.З.; де Пинна, MCC (2014). «Ни о чём». Кладистика . 30 (3): 330–336. дои : 10.1111/cla.12050 . ПМИД 34788975 . S2CID 221550586 .
- ^ Паттерсон, К. (1982). «Морфологические признаки и гомологии». В К.А. Джойси; AE Friday (ред.). Проблемы филогенетической реконструкции . Лондон и Нью-Йорк: Академическая пресса. стр. 21–74.
- ^ Хаас, О. и Г.Г. Симпсон. 1946. Анализ некоторых филогенетических терминов с попытками их переопределения. Учеб. амер. Фил. Соц. 90 :319-349.
- ^ Никсон, КЦ; Карпентер, Дж. М. (2011). «О гомологии» . Кладистика . 28 (2): 160–169. дои : 10.1111/j.1096-0031.2011.00371.x . ПМИД 34861754 . S2CID 221582887 .
- ^ Jump up to: а б Бруска, Колорадо; Бруска, Дж.Дж. (1990). Беспозвоночные . Синауэр Ассошиэйтс. п. 669 .
- ^ Кэрролл, Шон Б. (2006). Бесконечные формы, самые красивые . Вайденфельд и Николсон. стр. 28, 66–69. ISBN 978-0-297-85094-6 .
- ^ Холл, Брайан (2008). Гомология . Джон Уайли. п. 29. ISBN 978-0-470-51566-2 .
- ^ Шинг, Х.; Эриксон, Э.Х. (1982). «Некоторые ультраструктуры жала пчелы ( Apis mellifera L.)» . Апидология . 13 (3): 203–213. дои : 10.1051/apido:19820301 .
- ^ «Гомология: От челюстей до ушей — необычный пример гомологии» . Калифорнийский университет в Беркли . Проверено 15 декабря 2016 г.
- ^ Хайд, Джанет Шибли ; ДеЛаматер, Джон Д. (июнь 2010 г.). «Глава 5» (PDF) . Понимание человеческой сексуальности (11-е изд.). Нью-Йорк: МакГроу-Хилл . п. 103. ИСБН 978-0-07-338282-1 .
- ^ Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории . Современная библиотека. п. 112. ИСБН 978-0-679-64288-6 .
- ^ «Гомология: оставьте это растениям» . Калифорнийский университет в Беркли . Проверено 7 мая 2017 г.
- ^ Саттлер, Р. (1984). «Гомология — постоянный вызов». Систематическая ботаника . 9 (4): 382–394. дои : 10.2307/2418787 . JSTOR 2418787 .
- ^ Саттлер, Р. (1994). «Гомология, гомеозис и морфология процессов у растений». В зале, Брайан Кейт (ред.). Гомология: иерархическая основа сравнительной биологии . Академическая пресса. стр. 423–75. ISBN 978-0-12-319583-8 .
- ^ «Гомологии: биология развития» . Калифорнийский университет в Беркли . Проверено 15 декабря 2016 г.
- ^ «Кластерный FAQ #Символы» . Кластал . Архивировано из оригинала 24 октября 2016 года . Проверено 8 декабря 2014 г.
- ^ Кунин, Е.В. (2005). «Ортологи, паралоги и эволюционная геномика» . Ежегодный обзор генетики . 39 : 309–38. дои : 10.1146/annurev.genet.39.073003.114725 . ПМИД 16285863 .
- ^ Fitch, WM (июнь 1970 г.). «Отличение гомологичных белков от аналогичных». Систематическая зоология . 19 (2): 99–113. дои : 10.2307/2412448 . JSTOR 2412448 . ПМИД 5449325 .
- ^ Закани, Йожеф; Дюбул, Денис (2007). «Роль Hox-генов в развитии конечностей позвоночных». Текущее мнение в области генетики и развития . 17 (4): 359–366. дои : 10.1016/j.где.2007.05.011 . ISSN 0959-437X . ПМИД 17644373 .
- ^ Холм, Лийза; Лайхо, Алекси; Тёрёнен, Петри; Сальгадо, Марко (23 ноября 2022 г.). «DALI проливает свет на отдаленные гомологи: сто открытий» . Белковая наука . 32 (1): e4519. дои : 10.1002/pro.4519 . ISSN 0961-8368 . ПМЦ 9793968 . ПМИД 36419248 .
- ^ Мур, Дэвид С. (2013). «Импорт концепции гомологии из биологии в психологию развития». Развивающая психобиология . 55 (1): 13–21. дои : 10.1002/dev.21015 . ПМИД 22711075 .
- ^ Раецки, Д.В.; Флэнери, Рэндалл К. (2013). Лэмб, Мэн; Браун, Ал. (ред.). Социальный конфликт и доминирование у детей: аргументы в пользу гомологии приматов . Тейлор и Фрэнсис. п. 125. ИСБН 978-1-135-83123-3 .
Наконец, большое количество недавней информации о поведении детей и нечеловекообразных приматов в группах, сочетание достоверных данных о людях и достоверных данных о нечеловекообразных приматах, придает достоверность нашему сравнению. Наш вывод состоит в том, что, основываясь на совпадении нескольких необычных характеристик, паттерны доминирования гомологичны у приматов. Такое совпадение необычных характеристик обнаруживается на нескольких уровнях, включая мелкую моторику, крупную моторику и поведение на групповом уровне.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Венцель, Джон В. 1992. Поведенческая гомология и филогения. Ежегодный обзор экологии и систематики 23: 361-381
- ^ Хенниг, В. 1966. Филогенетическая систематика . Университет Иллинойса Пресс
Дальнейшее чтение
[ редактировать ]- Бригандт, Инго (2011) «Эссе: Гомология». В: Энциклопедия проекта «Эмбрион» . ISSN 1940-5030 . http://embryo.asu.edu/handle/10776/1754
- Кэрролл, Шон Б. (2006). Бесконечные формы, самые красивые . Нью-Йорк: WW Norton & Co. ISBN 978-0-297-85094-6 .
- Кэрролл, Шон Б. (2006). Создание сильнейших: ДНК и окончательная судебно-медицинская экспертиза эволюции . Нью-Йорк: WW Norton & Co. ISBN 978-0-393-06163-5 .
- ДеПинна, MC (1991). «Концепции и тесты гомологии в кладистической парадигме». Кладистика . 7 (4): 367–94. CiteSeerX 10.1.1.487.2259 . дои : 10.1111/j.1096-0031.1991.tb00045.x . S2CID 3551391 .
- Дьюи, Китай; Пахтер, Л. (апрель 2006 г.). «Эволюция на уровне нуклеотидов: проблема множественного выравнивания целого генома» . Молекулярная генетика человека . 15 (спец № 1): R51–R56. дои : 10.1093/hmg/ddl056 . ПМИД 16651369 .
- Fitch, WM (май 2000 г.). «Гомология - личный взгляд на некоторые проблемы». Тенденции в генетике . 16 (5): 227–31. дои : 10.1016/S0168-9525(00)02005-9 . ПМИД 10782117 .
- Гегенбаур, Г. (1898). Сравнительная анатомия позвоночных ... Лейпциг.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - Геккель, Эрнст (1866). Общая морфология организмов . Том 1-2. Берлин.
{{cite book}}: CS1 maint: местоположение ( ссылка ) CS1 maint: отсутствует местоположение издателя ( ссылка ) - Кузняр, А.; ван Хэм, RC; Понгор, С.; Леуниссен, JA (ноябрь 2008 г.). «В поисках ортологов: поиск соответствующего гена в геномах». Тенденции Жене . 24 (11): 539–551. дои : 10.1016/j.tig.2008.08.009 . ПМИД 18819722 .
- Минделл, ДП; Мейер, А. (2001). «Эволюция гомологии» (PDF) . Тенденции экологии и эволюции . 16 (8): 434–40. дои : 10.1016/S0169-5347(01)02206-6 . Архивировано из оригинала (PDF) 27 июня 2010 года.
- Оуэн, Ричард (1847). Об архетипе и гомологиях скелета позвоночных . Лондон: Джон ван Вурст, Патерностер Роу.
Внешние ссылки
[ редактировать ] СМИ, связанные с гомологией, на Викискладе?
СМИ, связанные с гомологией, на Викискладе?
