Крыло насекомого




Крылья насекомых — это взрослые выросты насекомых экзоскелета , которые позволяют насекомым летать . Они встречаются на втором и третьем грудных сегментах ( среднегруди и заднегруди ), и эти две пары часто называют передними и задними крыльями соответственно, хотя у некоторых насекомых задние крылья отсутствуют, даже зачатки. Крылья укреплены рядом продольных жилок, часто имеющих поперечные соединения, образующие закрытые «ячейки» в перепонке (крайние примеры включают стрекоз и златоглазок ). Паттерны, возникающие в результате слияния и перекрестного соединения жилок крыльев, часто являются диагностическими для различных эволюционных линий и могут использоваться для идентификации на уровне семейства или даже рода у многих отрядов насекомых.
Физически одни насекомые двигают летательными мышцами напрямую, другие — опосредованно. У насекомых с прямым полетом мускулы крыла непосредственно прикрепляются к основанию крыла, так что небольшое движение основания крыла вниз поднимает само крыло вверх. У насекомых с непрямым полетом есть мышцы, которые прикрепляются к грудной клетке и деформируют ее, заставляя крылья также двигаться.
Крылья присутствуют только у одного пола (часто мужского) у некоторых групп, таких как бархатные муравьи и Strepsiptera , или избирательно утрачены у «работников» общественных насекомых, таких как муравьи и термиты . Редко самка имеет крылья, а самец нет, как у инжирных ос . В некоторых случаях крылья образуются только в определенные моменты жизненного цикла, например, на этапе расселения тли . Структура и окраска крыльев часто различаются в зависимости от морф , например, у тлей , миграционных фаз саранчи и полиморфных бабочек . В состоянии покоя крылья можно держать ровно или складывать несколько раз по определенному рисунку; чаще всего складываются задние крылья, но в некоторых группах, таких как веспидовые осы , это передние крылья.
Эволюционное происхождение крыльев насекомых дискутируется. В XIX веке вопрос об эволюции крыльев насекомых первоначально стоял на двух основных позициях. Одна постулировала, что крылья насекомых произошли от ранее существовавших структур, тогда как вторая предполагала, что крылья насекомых представляли собой совершенно новые образования. [ 1 ] [ 2 ] «Новая» гипотеза предполагала, что крылья насекомых образовались не из ранее существовавших предковых придатков, а скорее как выросты из стенки тела насекомого. [ 3 ]
Уже давно исследования происхождения крыльев насекомых основываются на позиции «ранее существовавших структур», которая была первоначально предложена в 19 веке. [ 2 ] В недавней литературе указано, что несколько наследственных структур сыграли важную роль в происхождении крыльев насекомых. К ним относятся: жабры, дыхательные придатки ног, а также боковые (паранотальные) и задне-боковые выступы грудной клетки, и это лишь некоторые из них. [ 4 ]
Согласно более современной литературе, возможные кандидаты включают жаберные структуры, паранотальную долю и тергальную пластинку ракообразных. Последнее основано на недавних генетических исследованиях насекомых, которые показывают, что насекомые являются членистоногими панракообразных с прямым предком из ракообразных и общими генетическими механизмами развития конечностей. [ 3 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ]
Другими теориями происхождения крыльев насекомых являются теория паранотальных долей, теория жабр и двойная теория эволюции крыльев насекомых. Эти теории постулируют, что крылья либо развились из паранотальных долей, продолжений грудной терги ; [ 5 ] что они представляют собой модификации подвижных брюшных жабр обнаруженных у водных наяд подёнок , ; [ 5 ] или что крылья насекомых возникли в результате слияния ранее существовавших структур эндита и выхода, каждая из которых имела ранее существовавшие сочленения и трахеи. [ 9 ] [ 10 ]
Морфология
[ редактировать ]
Внутренний
[ редактировать ]Каждое из крыльев состоит из тонкой перепонки, поддерживаемой системой жилок. Мембрана образована двумя плотно прилегающими слоями покровов, тогда как вены образуются там, где эти два слоя остаются отдельными; иногда нижняя кутикула толще и сильнее склеротизирована под веной. Внутри каждой из крупных вен имеется нерв и трахея , и, поскольку полости вен соединены с гемоцелем , гемолимфа может оттекать в крылья. [ 11 ]
По мере развития крыла дорсальный и вентральный покровные слои на большей части своей площади плотно прилегают друг к другу, образуя перепонку крыла. Остальные участки образуют каналы, будущие вены, в которых могут располагаться нервы и трахеи. Кутикула, окружающая жилки, утолщается и сильно склеротизируется, что придает крылу прочность и жесткость. На крыльях могут встречаться два типа волос: микротрихии, которые мелкие и неравномерно разбросаны, и макротрихии, которые более крупные, с гнездами и могут ограничиваться жилками. Чешуя Lepidoptera и Trichoptera представляет собой сильно модифицированные макротрихии. [ 12 ]
Жилкование
[ редактировать ]
У некоторых очень мелких насекомых жилкование может быть сильно редуцировано. Например, у хальцидоидных ос имеются только подкоста и часть лучевой кости. И наоборот, увеличение жилкования может происходить за счет разветвления существующих жилок с образованием добавочных жилок или за счет развития дополнительных вставочных жилок между исходными, как в крыльях прямокрылых (кузнечиков и сверчков). У некоторых насекомых присутствует большое количество поперечных жилок, и они могут образовывать сетку, как на крыльях Odonata (стрекозы и стрекозы), так и у основания передних крыльев Tettigonioidea и Acridoidea (катидиды и кузнечики соответственно). [ 11 ]
Архедиктион . — название гипотетической схемы жилкования крыльев, предложенной для самого первого крылатого насекомого Он основан на сочетании предположений и ископаемых данных. Поскольку считается, что все крылатые насекомые произошли от общего предка, архедиктион представляет собой «шаблон», который был модифицирован (и оптимизирован) естественным отбором в течение 200 миллионов лет. Согласно современным догмам, архедиктион содержал 6–8 продольных жилок. Эти вены (и их ответвления) названы в соответствии с системой, разработанной Джоном Комстоком и Джорджем Нидхэмом, — системой Комстока-Нидхэма : [ 13 ]
- Коста (C) – передняя кромка крыла
- Subcosta (Sc) - вторая продольная жилка (за ребром), обычно неразветвленная.
- Радиус (R) – третья продольная жилка, до края крыла доходят от одной до пяти ветвей.
- Медиа (М) - четвертая продольная жилка, до края крыла доходят от одной до четырех ветвей.
- Cubitus (Cu) - пятая продольная жилка, до края крыла доходит от одной до трех ветвей.
- Анальные вены (А1, А2, А3) – неразветвленные вены позади локтевой кости.
Коста (С) — ведущая краевая жилка большинства насекомых. Иногда над ребром имеется небольшая жилка, называемая прекостой, хотя почти у всех современных насекомых [ 14 ] : 41–42 прекоста слита с костой. Ребро редко разветвляется, поскольку оно находится на переднем крае, который у его основания связан с плечевой пластинкой. Трахея реберной вены, возможно, является ветвью подреберной трахеи. После ребра расположена третья вена, подкоста, которая разветвляется на две отдельные вены: переднюю и заднюю. Основание подкосты связано с дистальным концом шейки первой подмышечной кости (см. раздел ниже). Четвертая жилка — лучевая (R), которая разветвляется на пять отдельных жилок. Радиус обычно является самой сильной жилкой крыла. Ближе к середине крыла он раздваивается на первую неделимую ветвь (R1) и вторую ветвь, называемую радиальным сектором (Ra), которая дихотомически подразделяется на четыре дистальные ветви (R2, R3, R4, R5). В основании лучевая кость гибко соединена с передним концом второй подмышечной кости (2Ах). [ 15 ]
Пятая жилка крыла — средства массовой информации. В архетипическом паттерне (А) медиа разветвляется на две основные ветви: переднюю медиану (МА), которая делится на две дистальные ветви (МА1, МА2), и срединный сектор, или медиа заднюю (МП), который имеет четыре концевые ответвления (М1, М2, М3, М4). У большинства современных насекомых передняя медиания утрачена, и привычной «медией» является четырехветвистая медиа-задняя с общим базальным стеблем. У Ephemerida, согласно современным интерпретациям жилкования крыльев, сохраняются обе ветви сред, тогда как у Odonata сохранившимися средами являются примитивная передняя ветвь. Ствол медиальной кости часто соединяется с лучевой кости, но когда она встречается в виде отдельной вены, ее основание связано с дистальной срединной пластинкой (m') или непрерывно склеротизировано с последней. Кубитус, шестая жилка крыла, преимущественно двуветвистый. Первичное раздвоение происходит у основания крыла, образуя две основные ветви (Cu1, Cu2). Передняя ветвь может распадаться на ряд второстепенных ветвей, но обычно она разветвляется на две дистальные ветви. Вторую ветвь кубитуса (Cu2) у перепончатокрылых, трихоптер и чешуекрылых Комсток и Нидэм приняли за первый анальный. Проксимально основной стебель кубитуса связан с дистальной срединной пластинкой (m') основания крыла. [ 15 ]
Посткубит (Pcu) — первый анальный элемент системы Комстока-Нидхэма. Посткубитус, однако, имеет статус самостоятельной жилки крыла и должен быть признан таковым. [ нужна ссылка ] В нимфальных крыльях ее трахея возникает между локтевой трахеей и группой ванальных трахей. В зрелых крыльях более генерализованных насекомых Postcubitus всегда связан проксимально с cubitus и никогда не связан тесно со склеритом-сгибателем (3Ax) основания крыла. У Neuroptera, Mecoptera и Trichoptera посткубитус может быть более тесно связан с ванальными жилками, но его основание всегда свободно от последних. Посткубит обычно неразветвленный; это примитивно двухразветвленный. Ваннальные вены (от lV до nV) — это анальные вены, которые непосредственно связаны с третьей подмышечной веной и на которые напрямую влияет движение этого склерита, вызывающее сгибание крыльев. Число ваннанных жилок различается. от 1 до 12 в зависимости от расширения венальной площади крыла. Ваннальные трахеи обычно возникают из общего ствола трахеи у нимфальных насекомых, а вены рассматриваются как ветви одной анальной вены. Дистально ванальные жилки простые или разветвленные. Скуловые вены (J) скуловой доли крыла часто заняты сетью неправильных жилок или могут быть целиком перепончатыми; но иногда он содержит одну или две отдельные мелкие вены: первую скуловую вену, или дуговую вену, и вторую скуловую вену, или vena cardinalis (2J). [ 15 ]
- Перекрестные вены C-Sc - проходят между ребром и подкостой.
- R поперечные жилки – проходят между соседними ветвями лучевой кости.
- Поперечные вены RM – проходят между лучевой и медиальной поверхностью.
- Поперечные жилки M-Cu - проходят между медией и локтевой пластинкой.
Все жилки крыла подвержены вторичному разветвлению и слиянию поперечными жилками. У некоторых отрядов насекомых поперечные жилки столь многочисленны, что весь жилковый рисунок представляет собой плотную сеть ветвящихся жилок и поперечных жилок. Однако обычно имеется определенное количество поперечных жилок, имеющих определенное расположение. Более постоянными поперечными венами являются плечевая поперечная вена (h) между ребрами и подкостами, радиальная поперечная вена (r) между R и первой вилкой Rs, секториальная поперечная вена (s) между двумя развилками Rs. R8, срединная поперечная вена (m-m) между M2 и M3 и медиально-кубитальная поперечная вена (m-cu) между средой и кубитом. [ 15 ]
Жилки крыльев насекомых характеризуются выпукло-вогнутым расположением, как у поденок (т. е. вогнутость — «вниз», выпуклость — «вверх»), которые регулярно чередуются, а также триадным типом ветвления; всякий раз, когда вена разветвляется, между двумя ветвями всегда имеется интерполированная жилка противоположного положения. Вогнутая жилка раздваивается на две вогнутые жилки (при этом интерполированная жилка становится выпуклой), при этом закономерное чередование жил сохраняется. [ 16 ] Жилки крыла кажутся волнообразными в зависимости от того, имеют ли они тенденцию складываться вверх или вниз, когда крыло расслаблено. Базальные стволы вен выпуклые, но каждая вена дистально разветвляется на переднюю выпуклую ветвь и заднюю вогнутую ветвь. Таким образом, ребра и подкосты рассматриваются как выпуклые и вогнутые ветви первичной первой вены, Rs — вогнутая ветвь лучевой кости, задняя медиальная ветвь — вогнутая ветвь средней жилки, Cu1 и Cu2 — соответственно выпуклая и вогнутая, тогда как примитивные Postcubitus и у первой ваннали есть передняя выпуклая ветвь и задняя вогнутая ветвь. Выпуклая или вогнутая природа жилок использовалась в качестве доказательства при определении идентичности сохранившихся дистальных ветвей жилок современных насекомых, но не было продемонстрировано, что она одинакова для всех крыльев. [ 11 ] [ 15 ]
Поля
[ редактировать ]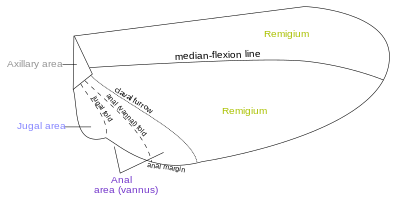
Области крыла ограничены и разделены линиями сгиба, по которым крыло может складываться, и линиями сгиба , по которым крыло может сгибаться во время полета. Фундаментальное различие между линиями сгиба и линиями сгиба часто размыто, поскольку линии сгиба могут обеспечивать некоторую гибкость или наоборот. Две константы, которые встречаются почти у всех крыльев насекомых, - это ключичные (линия сгибания) и скуловые складки (или линия сгиба); формирование изменчивых и неудовлетворительных границ. Складывание крыльев может быть очень сложным: поперечная складчатость наблюдается на задних крыльях Dermaptera и Coleoptera , а у некоторых насекомых анальная область может быть сложена веером. [ 14 ] На крыльях насекомых имеется около четырех различных полей:
- Гребля
- Анальная область (ваннус)
- Югальная область
- Подмышечная область
- Они непростые
Большинство вен и поперечных вен происходит в передней области ремигиума , который отвечает за большую часть полета, приводимого в движение грудными мышцами. Заднюю часть ремигия иногда называют клавусом ; два других задних поля — анальная и скуловая области . [ 14 ] Когда ваннанная складка имеет обычное положение кпереди от группы анальных вен, в ремигиуме присутствуют реберные, подреберные, радиальные, медиальные, кубитальные и посткубитальные вены. В согнутом крыле ремигиум поворачивается кзади на гибком базальном соединении лучевой кости со второй подмышечной, а основание медио-кубитального поля сложено медиально на подмышечной области по plica basalis (bf) между срединными пластинками (m, m'). ) основания крыла. [ 15 ]
Ваннус ограничен ванальной складкой, которая обычно возникает между посткубитусом и первой ванальной жилкой. У прямокрылых он обычно занимает такое положение. Однако у переднего крыла Blattidae единственная складка в этой части крыла лежит непосредственно перед посткубитусом. У Plecoptera ваннанная складка расположена позади посткубитуса, но проксимальнее пересекает основание первой ванальной жилки. У цикады ванна складка лежит сразу за первой ванной жилкой (lV). Эти небольшие изменения в фактическом положении ванальной складки, однако, не влияют на единство действия ваннанных жилок, контролируемых сгибателем склерита (3Ах), при сгибании крыла. На задних крыльях большинства прямокрылых вторичная делимая вена образует ребро в ванальной складке. Ваннус обычно имеет треугольную форму, а его вены обычно расходятся от третьей подмышечной вены, как веерные ребра. Некоторые из венальных вен могут быть разветвленными, а вторичные жилки могут чередоваться с первичными. Ваннальная область обычно лучше всего развита в заднем крыле, где она может быть увеличена, образуя опорную поверхность, как у Plecoptera и Orthoptera. Большие веерные расширения задних крыльев Acrididae явно представляют собой ваннальные области, поскольку все их жилки опираются на третьи подмышечные склериты оснований крыльев, хотя Мартынов (1925) относит большую часть веерных областей у Acrididae к скуловым областям крылья. Истинная югума крыла саранчовых представлена лишь небольшой перепончатой (Ju) месадой последней ваннанной жилки. Югум более развит у некоторых других Polyneoptera, например у Mantidae. У большинства высших насекомых с узкими крыльями ванна редуцируется и ванна складка теряется, но даже в таких случаях согнутое крыло может изгибаться по линии между послекубитусом и первой ванальной жилкой. [ 15 ]
Скуловая область, или Неала, представляет собой область крыла, которая обычно представляет собой небольшую перепончатую область проксимальнее основания ваннуса, укрепленную несколькими небольшими неравномерными жилеобразными утолщениями; но когда он хорошо развит, он представляет собой отдельную часть крыла и может содержать одну или две скуловые жилки. Когда скуловая область переднего крыла развивается как свободная лопасть, она выступает под плечевой угол заднего крыла и, таким образом, служит для соединения двух крыльев вместе. В группе Jugatae чешуекрылых он имеет длинную пальцеобразную лопасть. Скуловую область назвали неалой («новое крыло»), потому что она, очевидно, является вторичной и недавно развившейся частью крыла. [ 15 ]
Подмышечная область представляет собой область, содержащую подмышечные склериты, и имеет в целом форму лестничного треугольника. Основание треугольника (ab) — шарнир крыла с корпусом; вершина (в) — дистальный конец третьего подмышечного склерита; более длинная сторона находится впереди вершины. Точка d на передней стороне треугольника отмечает место сочленения лучевой вены со вторым подмышечным склеритом. Линия между d и c представляет собой базальную складку (bf) или складку крыла у основания медиокубитального поля. [ 15 ] Термен — это внешний край крыла, между вершиной и задним или анальным углом. [ 17 ] [ 18 ]
В заднем углу основания крыла у некоторых двукрылых имеется пара перепончатых лопастей (чешуек, или чашелистиков), известных как алула. У комнатной мухи хорошо развит алула. Наружная чешуя (в) начинается от основания крыла позади третьего подмышечного склерита (3Ах) и, очевидно, представляет собой скуловую лопасть других насекомых (А, D); более крупная внутренняя чешуя (d) начинается от заднего щиткового края тергума крылатого сегмента и образует защитный капюшонообразный навес над жужжальца. В согнутом крыле наружная чешуя alula перевернута вверх дном над внутренней чешуей, причем последняя не затрагивается движением крыла. У многих двукрылых глубокий разрез анальной области перепонки крыла позади единственной ванальной вены отделяет проксимальную крыльную лопасть, дистальнее наружной чешуи алулы. [ 15 ]
Суставы
[ редактировать ]
Различные движения крыльев, особенно у насекомых, которые в состоянии покоя сгибают крылья горизонтально над спиной, требуют более сложной суставной структуры у основания крыла, чем простое соединение крыла с телом. Каждое крыло прикреплено к телу перепончатой базальной областью, но суставная мембрана содержит ряд небольших суставных склеритов, известных под общим названием птералии. Птералии включают переднюю плечевую пластинку у основания реберной вены, группу подмышечных желез (Ax), связанных с подреберной, радиальной и ванной венами, и две менее выраженные срединные пластинки (m, m') у основания реберной вены. медиокубитальная зона. Подмышечные ветви специально развиты только у насекомых, сгибающих крылья, где они составляют сгибательный механизм крыла, приводимый в действие мышцей-сгибателем, возникающей на плевронах. Для основания крыла характерна также небольшая лопасть на переднем крае суставной области проксимальнее плечевой пластинки, которая у передних крыльев некоторых насекомых развита в крупный плоский чешуевидный лоскут — тегулу, перекрывающий друг друга. основание крыла. Сзади суставная перепонка часто образует обширную лопасть между крылом и телом, а ее край обычно утолщен и гофрирован, создавая вид связки, так называемого подмышечного тяжа, продолжающегося мезиально с задней краевой щитковой складкой тергальной кости. пластина, несущая крыло. [ 15 ]
Суставные склериты, или птералии основания крыльев сгибающих крылья насекомых и их отношение к телу и жилкам крыла, показанные схематически, следующие:
- Плечевые пластины
- Первая подмышечная
- Вторая подмышечная
- Третья подмышечная
- Четвертая подмышечная
- Срединные плиты ( м , м' )
Плечевая пластинка обычно представляет собой небольшой склерит на переднем крае основания крыла, подвижный и сочлененный с основанием реберной вены. У Odonata плечевая пластинка сильно увеличена. [ 15 ] с двумя мышцами, отходящими от эпистерна, вставленными в плечевые пластинки, и двумя от края эпимерона, вставленными в подмышечную пластинку. [ 11 ]
Первый подмышечный склерит (lAx) — передняя шарнирная пластинка основания крыла. Его передняя часть опирается на передний нотальный крыльевой отросток тергума (ANP); его задняя часть сочленяется с тергальным краем. Передний конец склерита обычно представляет собой тонкое плечо, вершина которого (д) всегда связана с основанием подреберной вены (Sc), но не соединена с последней. Тело склерита латерально сочленяется со второй подмышечной впадиной. Второй аксиллярный склерит (2Ах) более изменчив по форме, чем первый, но его механические связи не менее определенны. Она наклонно шарнирно прикреплена к наружному краю тела первой подмышечной вены, а к ее переднему концу (г) всегда гибко прикрепляется лучевая вена (R). Вторая подмышечная в основании крыла имеет как дорсальную, так и вентральную склеротизацию; его вентральная поверхность опирается на опорный крыльевой отросток плевры. Таким образом, вторая подмышечная кость представляет собой основной склерит основания крыла и конкретно управляет радиальной жилкой. [ 15 ]
Третий подмышечный склерит (3Ах) лежит в задней части суставной области крыла. Форма его весьма разнообразна и часто неправильная, но третья подмышечная кость представляет собой склерит, к которому прикрепляется мышца-сгибатель крыла (D). Мезально он сочленяется спереди (е) с задним концом второй подмышечной кости, а сзади (б) с задним крыльевым отростком тергума (ПНП) или с небольшой четвертой подмышечной веткой, если последняя имеется. Дистально третья подмышечная артерия удлинена в виде отростка, который всегда связан с основаниями группы жилок анальной области крыла, называемой здесь ванальными жилками (V). Таким образом, третья подмышечная мышца обычно представляет собой заднюю шарнирную пластинку основания крыла и является активным склеритом сгибательного механизма, который непосредственно управляет ванальными венами. Сокращение мышцы-сгибателя (D) вращает третью подмышечную мышцу в ее мезальных сочленениях (b, f) и тем самым поднимает ее дистальную часть; это движение вызывает сгибание крыла. Четвертый подмышечный склерит не является постоянным элементом основания крыла. Если он присутствует, то обычно представляет собой небольшую пластинку, находящуюся между третьей подмышечной впадиной и задним отростком нотального крыла и, вероятно, представляет собой отдельную часть последнего. [ 15 ]
Срединные пластинки (m, m') также представляют собой склериты, не столь четко дифференцированные как специфические пластинки, как три основные подмышечные, но тем не менее они являются важными элементами сгибательного аппарата. Они лежат в срединной области основания крыла дистальнее второй и третьей подмышечных костей и отделены друг от друга косой линией (bf), образующей при сгибании крыла выраженную выпуклую складку. Проксимальная пластинка (m) обычно прикрепляется к дистальному плечу третьей подмышечной впадины и, возможно, ее следует рассматривать как часть последней. Дистальная пластинка (m') менее постоянно представлена в виде отчетливого склерита и может быть представлена общей склеротизацией основания медио-кубитального поля крыла. Когда вены этой области различимы у основания, они связаны с наружной срединной пластинкой. [ 15 ]
Мышцы
[ редактировать ]
Мышцы, управляющие полетом насекомых, могут занимать от 10% до 30% общей массы тела. Мышцы, управляющие полетом, различаются в зависимости от двух типов полета насекомых: непрямого и прямого. У насекомых, которые используют первый, непрямой способ передвижения, мышцы прикрепляются к тергуму, а не к крыльям, как следует из названия. Когда мышцы сокращаются, грудной отдел деформируется, передавая энергию крылу. Есть два «пучка» мышц: те, которые простираются параллельно тергуму, дорсо-продольные, и те, которые прикрепляются к тегуму и доходят до грудины, дорсовентральные. [ 19 ] В прямой мышце соединение идет непосредственно от плевры (грудной стенки) к отдельным склеритам, расположенным у основания крыла. Субалярные и базилярные мышцы имеют связки, прикрепляющиеся к субалярным и базилярным склеритам. Здесь резилин — высокоэластичный материал — образует связки, соединяющие летательные мышцы с крыльевым аппаратом.
У более производных отрядов насекомых, таких как двукрылые (мухи) и перепончатокрылые (оса), непрямые мышцы занимают наибольший объем птероторакса и служат основным источником энергии для взмахов крыльев. Сокращение дорсо-продольных мышц вызывает сильное выгибание нотума, которое прижимает крыло, в то время как сокращение дорсовентральных мышц вызывает противоположное движение нотума. Самые примитивные из ныне живущих летающих насекомых, Ephemeroptera (подёнки) и Odonata (стрекозы), используют прямые мышцы, отвечающие за развитие необходимой силы для взмахов вверх и вниз. [ 19 ] [ 20 ]
Мышцы крыльев насекомых представляют собой строго аэробную ткань. На единицу белка он потребляет топливо и кислород со скоростью, наблюдаемой в очень концентрированной и высокоорганизованной ткани, так что стационарные скорости на единицу объема представляют собой абсолютный рекорд в биологии. Кровь, богатая топливом и кислородом, переносится к мышцам посредством диффузии, происходящей в больших количествах, чтобы поддерживать высокий уровень энергии, используемой во время полета. Многие мышцы крыльев большие и могут достигать 10 мм в длину и 2 мм в ширину. Более того, у некоторых двукрылых волокна имеют гигантские размеры. Например, у очень активной Rutilia поперечное сечение составляет 1800 мкм в длину и более 500 мкм в ширину. Поэтому транспорт топлива и кислорода из окружающей среды к местам потребления и обратный транспорт углекислого газа представляют собой задачу для биолога как в отношении транспорта в жидкой фазе, так и в сложной системе воздушных трубок, т. е. в трахеи. система. [ 21 ]
Датчики
[ редактировать ]несколько типов сенсорных нейронов На крыльях насекомых обнаружено : вкусовые щетинки , [ 22 ] механосенсорные щетинки, [ 23 ] колокольчатые сенсиллы , [ 24 ] и хордотональные органы . [ 25 ] Эти сенсоры обеспечивают нервную систему как внешней, так и внутренней проприоцептивной обратной связью, необходимой для эффективного полета. [ 26 ] и уход. [ 27 ]
Сцепление, складывание и другие функции
[ редактировать ]У многих видов насекомых переднее и заднее крыло могут быть соединены вместе, что повышает аэродинамическую эффективность полета за счет объединения переднего и заднего крыльев в одно большее крыло. Наиболее распространенный механизм сцепления (например, у Hymenoptera и Trichoptera ) представляет собой ряд маленьких крючков на переднем крае заднего крыла, или « хамули », которые фиксируются на переднем крыле, удерживая их вместе (крючковидное соединение). У некоторых других видов насекомых (например, Mecoptera , Lepidoptera и некоторых Trichoptera ) скуловая лопасть переднего крыла покрывает часть заднего крыла (скуловое соединение), или края переднего и заднего крыла широко перекрываются (амплексиформное соединение), или щетинки заднего крыла, или уздечка, зацепляются под удерживающую структуру или удерживатель на переднем крыле. [ 14 ] : 43
В состоянии покоя у большинства насекомых крылья держат за спиной, что может включать продольное складывание перепонки крыла, а иногда и поперечное складывание. Иногда по линиям сгибания может возникать складчатость. Хотя линии сгиба могут быть поперечными, как на задних крыльях жуков и уховерток, они обычно радиальны к основанию крыла, что позволяет сгибать соседние части крыла друг над другом или под друг другом. Наиболее распространенной линией сгиба является скуловая складка, расположенная сразу за третьей анальной веной. [ 12 ] хотя у большинства Neoptera есть скуловая складка сразу за жилкой 3А на передних крыльях. Иногда он присутствует и на задних крыльях. Там, где анальная область заднего крыла большая, как у прямокрылых и Blattodea, вся эта часть может загибаться под переднюю часть крыла по ванальной складке немного позади ключичной борозды. Кроме того, у прямокрылых и Blattodea анальное поле веерообразно сложено вдоль жилок, при этом анальные жилки выпуклые, на гребнях складок, а добавочные жилки вогнутые. Если ключичная борозда и скуловая складка, вероятно, гомологичны у разных видов, то положение ваннальной складки у разных таксонов различается. Складывание производится мышцей, возникающей на плевре и прикрепляющейся к третьему подмышечному склериту таким образом, что при сокращении склерит поворачивается вокруг точек сочленения с задним нотальным отростком и вторым подмышечным склеритом. [ 11 ]
В результате дистальное плечо третьего подмышечного склерита вращается вверх и внутрь, так что, наконец, его положение полностью меняется на противоположное. Анальные жилки сочленены со склеритом таким образом, что при его движении они увлекаются за ним и изгибаются над спиной насекомого. Активность той же мышцы в полете влияет на выходную мощность крыла, поэтому она также важна для управления полетом. У ортоптероидных насекомых эластичность кутикулы приводит к сгибанию венальной области крыла вдоль жилок. Следовательно, энергия расходуется на раскрытие этой области, когда крылья перемещаются в полетное положение. В целом разгибание крыльев, вероятно, является результатом сокращения мышц, прикрепленных к базилярному склериту или, у некоторых насекомых, к субалярному склериту. [ 11 ]
Полет
[ редактировать ]
Механизмы полета
[ редактировать ]У двух групп относительно крупных насекомых, Ephemeroptera ( подёнки ) и Odonata ( стрекозы и стрекозы ), летательные мышцы прикреплены непосредственно к крыльям; крылья могут биться не быстрее, чем скорость, с которой нервы посылают импульсы, приказывающие мышцам биться. [ 28 ] Все другие живые крылатые насекомые летают, используя другой механизм, задействуя мышцы непрямого полета, которые вызывают вибрацию грудной клетки; крылья могут биться быстрее, чем скорость, с которой мышцы получают нервные импульсы. Этот механизм возник однажды и является определяющей особенностью ( синапоморфией ) инфракласса Neoptera . [ 28 ]
Аэродинамика
[ редактировать ]Существуют две основные аэродинамические модели полета насекомых. Большинство насекомых используют метод, который создает спиральный на передней кромке вихрь . [ 29 ] [ 30 ] Некоторые очень маленькие насекомые используют механизм «броска и хлопка» или механизм Вейса-Фога, при котором крылья хлопают вместе над телом насекомого, а затем разлетаются в стороны. Когда они раскрываются, воздух засасывается и создает вихрь над каждым крылом. Этот связанный вихрь затем движется поперек крыла и при хлопке действует как стартовый вихрь для другого крыла. Циркуляция и подъемная сила увеличиваются за счет износа крыльев. [ 29 ] [ 30 ]
Многие насекомые могут парить, быстро взмахивая крыльями, что требует боковой стабилизации и подъемной силы. [ 31 ]
Некоторые насекомые используют планирующий полет без использования тяги.
Эволюция
[ редактировать ]Где-то в каменноугольном периоде , около 350 миллионов лет назад, когда существовало всего два крупных массива суши, насекомые начали летать. Однако как и почему развились крылья насекомых, до сих пор не совсем понятно, во многом из-за нехватки соответствующих окаменелостей периода их развития в нижнем карбоне. Три основные теории происхождения полета насекомых заключаются в том, что крылья развились из паранотальных долей, продолжений грудной терги ; что они представляют собой модификации подвижных брюшных жабр обнаруженных у водных наяд подёнок , ; или что они развились из грудных выступов, используемых в качестве излучателей . [ 32 ]
Окаменелости
[ редактировать ]
Все окаменелости девона ( 400 миллионов лет назад) были бескрылыми, но к каменноугольному периоду (320 миллионов лет назад) более 10 различных родов насекомых имели полностью функциональные крылья. Переходных форм между двумя периодами сохранилось мало. Самые ранние крылатые насекомые относятся к этому периоду времени ( Pterygota ), в том числе Blattoptera , Caloneurodea стеблевой группы , примитивные Ephemeropterans , Orthoptera и Palaeodictyopteroidea . Очень ранние Blattopterans (в каменноугольном периоде) имели очень большую дискоидную переднеспинку и кожистые передние крылья с отчетливой жилкой CuP (неразветвленная жилка крыла, лежащая возле ключичной складки и достигающая заднего края крыла). [ 33 ] : 399 Несмотря на то, что самым старым возможным ископаемым насекомым является девонский Rhyniognatha hirsti , возраст которого оценивается в 396–407 миллионов лет, он обладал двумыщелковыми нижними челюстями, особенностью, свойственной крылатым насекомым. [ 34 ] хотя позже его считают возможным многоножкой . [ 35 ]
В пермский период стрекозы ) были доминирующими воздушными хищниками и, вероятно , ( Odonata также доминировали среди наземных насекомых. Настоящая одоната появилась в Перми. [ 36 ] [ 37 ] и все земноводные . Их прототипами являются древнейшие крылатые окаменелости, [ 38 ] восходят к девонскому периоду и во всех отношениях отличаются от других крыльев. [ 39 ] Их прототипы, возможно, имели зачатки многих современных признаков даже к позднему каменноугольному периоду , и вполне возможно, что они захватывали даже мелких позвоночных, поскольку некоторые виды имели размах крыльев 71 см. [ 37 ] Самые ранние жукообразные виды в перми имели заостренные, кожистые передние крылья с ячейками и ямками. Hemiptera , или настоящие клопы, появились в виде Arctiniscytina и Paraknightia, имеющих передние крылья с необычным жилкованием, возможно, расходящихся от Blattoptera . [ 33 ] : 186
Единственное крупное крыло одного из видов двукрылых триаса ( 10 мм вместо обычных 2–6 мм) обнаружено в Австралии (гора Кросби). Это семейство Tilliardipteridae, несмотря на многочисленные «типулоидные» признаки, следует включить в состав Psychodomorpha. sensu Hennig из-за потери выпуклой дистальной части 1А, достигающей края крыла, и образования анальной петли. [ 40 ]
Гипотезы
[ редактировать ]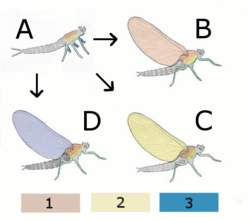
Гипотетический бескрылый предок
Б. Паранотальная теория:
Гипотетическое насекомое с крыльями сзади (Notum)
C Гипотетическое насекомое с крыльями плевры.
D Эпикоксальная теория
Гипотетическое насекомое с крыльями из придатков ног.
1 Нотум (сзади)
2 Плевра
3 Выход (внешние крепления ножек)
- Паранотальная гипотеза : эта гипотеза предполагает, что крылья насекомого развились из паранотальных долей - преадаптации , обнаруженной в окаменелостях насекомых , которая, как полагают, способствовала стабилизации при прыжках или падении. В пользу этой гипотезы говорит склонность большинства насекомых, испуганных при лазании по ветвям, спасаться бегством, падая на землю. Такие доли служили бы парашютами и позволяли насекомому мягче приземляться. Теория предполагает, что эти доли постепенно увеличивались и на более поздней стадии образовали соединение с грудной клеткой. Еще позже появились мышцы, способные двигать этими грубыми крыльями. Эта модель подразумевает постепенное повышение эффективности крыльев, начиная с парашютного полета , затем планирования и, наконец, активного полета . Тем не менее, отсутствие существенных ископаемых свидетельств развития суставов и мышц крыльев представляет собой серьезную трудность для теории, равно как и кажущееся спонтанным развитие сочленений и жилкования. [ 32 ]
- Эпикоксальная гипотеза : эта теория, впервые предложенная в 1870 году Карлом Гегенбауром, предполагала, что возможным источником крыльев насекомых могли быть подвижные брюшные жабры, обнаруженные у многих водных насекомых, например, наяд подёнок у . [ 41 ] Согласно этой теории, жабры трахеи , которые начинали как выходы дыхательной системы и со временем превратились в двигательные функции, в конечном итоге превратились в крылья. Жабры трахеи оснащены маленькими крылышками, которые постоянно вибрируют, и имеют собственные крошечные прямые мышцы. [ 32 ]
- Гипотеза эндита-выхода . Эта гипотеза основана на адаптации эндитов и выходов, придатков на соответствующих внутренних и внешних сторонах конечностей примитивных членистоногих. Его разработал Труман [ 42 ] на основе исследования Гольдшмидта в 1945 году на Drosophila melanogaster , в котором вариация стручка продемонстрировала мутацию, трансформирующую нормальные крылья в то, что было интерпретировано как трехчленное расположение ног с некоторыми дополнительными придатками, но без предплюсны, где обычно находится реберная поверхность крыла. быть. Эта мутация была переинтерпретирована как убедительное доказательство слияния дорсального выхода и эндита, а не ноги, при этом придатки гораздо лучше соответствуют этой гипотезе. Иннервация, артикуляция и мускулатура, необходимые для развития крыльев, уже присутствуют в подомерах. [ 32 ]
- Гипотеза рекрутирования генов «паранота плюс ноги» (также известная как гипотеза двойного происхождения) : ископаемые личинки Coxoplectoptera предоставили важные новые ключи к разгадке спорного вопроса об эволюционном происхождении крыльев насекомых. До открытия окаменелостей личинок паранотальная гипотеза и гипотеза выхода ног считались несовместимыми альтернативными объяснениями, оба из которых были подтверждены рядом доказательств из летописи окаменелостей , сравнительной морфологии , биологии развития и генетики . Экспрессия генов ног в онтогенезе крыльев насекомых повсеместно рассматривается как убедительное доказательство в пользу гипотезы выхода ног, которая предполагает, что крылья насекомых произошли от подвижных придатков ног (выходов). Однако личинки Coxoplectoptera показывают, что брюшные жабры поденок и их предков, которые обычно считаются структурами, соответствующими крыльям насекомых, сочленяются внутри дорсальных тергитных пластинок. У современных личинок поденок этого не наблюдается, поскольку тергиты и стерниты брюшка у них срослись в кольца, не оставляя никаких следов даже в эмбриональном развитии. Если личиночные жабры и крылья представляют собой соответствующие («серийные гомологичные») структуры и, таким образом, имеют одно и то же эволюционное происхождение, новые результаты по Coxoplectoptera демонстрируют, что крылья также имеют тергальное происхождение, как предполагает классическая паранотальная гипотеза. Станичек, Бехли и Годунко (2011) [ 32 ] [ 43 ] поэтому предложил новую гипотезу, которая могла бы согласовать явно противоречивые данные палеонтологии и генетики развития : крылья сначала возникли как жесткие выросты тергальных пластинок ( паранота ), и только позже в эволюции стали подвижными, сочлененными придатками посредством вторичного рекрутирования генов ног. [ 32 ] Более поздний анализ окаменелостей палеозойских подушечек крыльев нимф обеспечивает дополнительное подтверждение слияния элементов параноты и генов ног членистоногих. [ 44 ]
Были высказаны предположения, что крылья, возможно, изначально развились для плавания по поверхности воды, как это наблюдается у некоторых веснянок . [ 45 ] Альтернативная идея состоит в том, что оно происходит от направленного воздушного планирующего спуска — предполетного явления, обнаруженного у некоторых аптеригот , бескрылых сестринских таксонов крылатых насекомых. [ 46 ] Самые ранние летающие существа были похожи на стрекоз с двумя наборами крыльев, мускулами прямого полета и отсутствием способности складывать крылья на брюшке . Большинство современных насекомых, которые произошли от тех первых летателей, упростились до одной пары крыльев или двух пар, функционирующих как одна пара и использующих систему мышц непрямого полета. [ 32 ]
Естественный отбор сыграл огромную роль в совершенствовании крыльев, систем управления и сенсорики , а также всего остального, что влияет на аэродинамику и кинематику . Одна примечательная черта — поворот крыльев. Крылья большинства насекомых закручены, как и лопасти вертолета, с большим углом атаки у основания. Угол поворота обычно составляет от 10 до 20 градусов. Помимо этого поворота, поверхности крыльев не обязательно плоские или безликие; У большинства более крупных насекомых перепонки крыльев искажены и расположены под углом между жилками таким образом, что поперечное сечение крыльев напоминает аэродинамический профиль . Таким образом, базовая форма крыла уже способна создавать небольшую подъемную силу при нулевом угле атаки . Большинство насекомых управляют своими крыльями, регулируя наклон, жесткость и частоту взмахов крыльев с помощью крошечных мышц грудной клетки (внизу). У некоторых насекомых появились другие особенности крыльев, которые не выгодны для полета, но играют роль в чем-то другом, например, в спаривании или защите . [ 32 ]
| Эволюция способов создания крыльев в покое тела | ||
| крылья не складываются назад (недавние археоптеры) |
распространение вбок (большие пузыри) | |
| через спину друг против друга (стрекозы, подёнки) | ||
| Складной (Неоптера) | ||
| крылья не складные (например, веснянки) | ||
| Складной | веерообразно сложенные (например, передние крылья осы) | |
| Поперечная складка (например, заднее крыло жука) | ||
| Складывающиеся предметы (например, заднее крыло уховертки) | ||
Некоторым насекомым, занимающим те биологические ниши, которые они занимают, необходимо быть невероятно маневренными. Они должны находить пищу в ограниченном пространстве и быть в состоянии спастись от более крупных хищников – или они сами могут быть хищниками и им необходимо ловить добычу. Их маневренность с аэродинамической точки зрения обеспечивается большой подъемной и тяговой силой. Типичные летающие насекомые могут развивать подъемную силу, в три раза превышающую их вес, и силу горизонтальной тяги, в пять раз превышающую их вес. Существует два существенно разных механизма полета насекомых, и каждый из них имеет свои преимущества и недостатки - то, что у стрекозы более примитивный механизм полета, не означает, что они менее способны летать; в определенном смысле они более гибкие, чем все, что появилось впоследствии. [ 32 ]
Морфогенез
[ редактировать ]В то время как развитие крыльев у насекомых четко выражено у представителей Endopterygota , претерпевающих полный метаморфоз ; у этих видов крыло развивается на стадии куколки жизненного цикла насекомых. Однако у насекомых, претерпевающих неполный метаморфоз , стадия куколки отсутствует, поэтому у них должен быть другой морфогенез крыльев . У насекомых, обладающих гемиметаболизмом, есть крылья, которые начинаются с почек, которые находятся под экзоскелетом и не обнажаются до последней возрастной стадии нимфы . [ 47 ]
Первым признаком появления зачатков крыльев является утолщение гиподермы, которое можно наблюдать у насекомых как на ранних стадиях эмбрионального развития, так и на самых ранних стадиях жизненного цикла. Во время развития морфологических особенностей в эмбрионе, или эмбриогенезе растет скопление клеток , под эктодермой , которые на более позднем этапе развития, после того как латеральная эктодерма разрастается дорсально, образуют ветряной имагинальный диск. Пример развития зачатков крыльев у личинок можно увидеть у белых бабочек ( Pieris ). На втором этапе гистобласты становятся более заметными и теперь образуют карманообразную структуру. Начиная с третьего и четвертого возрастов гистобласты становятся более удлиненными. Эта сильно расширенная и выпуклая часть и становится крылом. К концу последней, или пятой, стадии крыло выталкивается из крылового кармана, хотя и на стадии предкуколки продолжает лежать под старой личиночной кутикулой. Только когда бабочка достигает стадии куколки, зачаток крыла обнажается, и вскоре после этого При эклозии крыло начинает расширяться и принимать окончательную форму. [ 47 ]
Развитие трахеации крыльев начинается еще до образования гистобластов крыльев, так как важно отметить, что они развиваются вблизи большой трахеи . В течение четвертого возраста клетки эпителия этой трахеи сильно увеличиваются и проникают в полость зачатка крыла, при этом каждая клетка образует плотно скрученную трахеолу . Каждая трахколе имеет одноклеточное происхождение и сначала находится внутриклеточно; тогда как трахеи имеют многоклеточное происхождение и просвет каждой из них межклеточный. Развитие трахеол, каждая из которых свернута внутри одной клетки эпителия трахеи, и последующее открытие сообщения между трахеолами и просветом трахеи, а также разворачивание и вытягивание трахеол, так что они достигают всех частей крыла. [ 47 ]
На более ранних стадиях своего развития крыльевая зачатка не снабжена специальными органами дыхания, например трахеей, так как в этом отношении она напоминает другие участки гиподермы, частью которой она еще является. Гистобласт развивается вблизи крупной трахеи, поперечный разрез которой показан на рисунке, представляющем участки этих частей первого, второго, третьего и четвертого возрастов соответственно. При этом трахеолы разворачиваются и вытягиваются пучками в образующихся жилках-полостях зачатка крыла. Во время линьки, знаменующей начало стадии куколки, они становятся функциональными. При этом личиночные трахеолы перерождаются; их функция заменена крыльевыми трахеями. [ 47 ]
Номенклатура
[ редактировать ]Большая часть номенклатуры отрядов насекомых основана на древнегреческом слове, обозначающем крыло, πτερόν ( птерон ), как суффиксе -ptera .
| Научное название | лингвистический корень | Перевод научного названия | Английское имя |
|---|---|---|---|
| Anisoptera | анизо- ( анизо- ) | Неравные крылья | Стрекоза |
| Аптера | ἀ- ( а- ), не | Бескрылый | Аптериготаны , ныне устаревшие |
| Аптеригота | птеригион маленькое крыло [ нужна ссылка ] ἀ- ( а- ), не |
Бескрылый | Аптериготы |
| Жесткокрылые | Колеос ( колеос , оболочка) | Закаленные крылья | Жуки |
| Дермаптера | Дерма ( дерма , кожа, кожа) | Кожаные крылья | Уховертки |
| Диафаноптеродея | Прозрачный ( диафан , прозрачный или полупрозрачный) | С прозрачными крыльями | диафаноптероидеи |
| Диктиоптеры | Диктён ( диктион , сеть) | Крылья с сетчатым жилкованием. | Тараканы, богомолы и термиты |
| двукрылые | Дё- (дё-, два ) | Два крыла | Мухи |
| Эмбиоптеры | ἐν- ( en , внутри; биос биос , жизнь) | Внутренние живущие крылатые насекомые | Вебспиннеры |
| Эндоптеригота | entós ( entos , внутри; птеригион, маленькое крыло) | Внутренние крылья | Голометаболические насекомые |
| Эфемероптера | ἐφήμερος ( эфемеры продолжительностью около одного дня) | Недолгоживущие крылатые насекомые. | Подёнки |
| Экзотеригота | ἔξο ( экзо , внешний) | Внешние крылья | Насекомые, претерпевающие неполный метаморфоз (и, следовательно, имеющие внешне видимые зачатки крыльев, как у нимф) |
| полужесткокрылые | ἡμι- ( полу- , половина) | Полукрылые насекомые | Hemiptera (настоящие клопы, цикадки, тля и др.) |
| полужесткокрылые | гетеро- ( гетеро- , разные) | Разные крылатые | Настоящие ошибки |
| Равнокрылые | ὅμο- ( гомо- , подобный) | Такой же крылатый | теперь устарело |
| Перепончатокрылые | гимений ( гименион , перепонка) | Насекомые с крыльями из тонких перепонок. | пчелы, осы, муравьи и т. д. |
| Изоптера | ион ( изон , равно) | Такой же крылатый | Термиты |
| Чешуекрылые | Лепис ( лепис , масштаб) | Чешуйчатые крылья | Бабочки и мотыльки |
| Лончоптериды | Ланс ( лонче , копье) | Крылья копья | Лэнс летает |
| Мекоптера | μῆκος ( мекос , длина) | Длинные крылья | Змея летает и т. д. |
| Мегалоптера | Мегало- ( мегало- , большой) | Большие крылья | Добсономухи, рыбные мухи |
| Нейрокрылые | нейрон ( нейрон , вена) | Прожилковое крыло | Златоглазки, совы, муравьиные львы и т. д. |
| Неоптера | neos ( неос , новый, молодой) | Новые крылья | Включает все существующие в настоящее время отряды летающих насекомых, кроме подёнок и стрекоз. |
| Олигонеоптеры | ὀλιγον- ( олигон- , немногие) новый нео или |
Новый, с небольшими прожилками. | Отдел Неоптеры |
| Прямокрылые | ὀρθο ( орто- , прямой) | Прямые крылья | Кузнечики, кузнечики и сверчки |
| Палеодиктиоптера | Старый ( палеос- , старый) сеть диктион, что означает |
Старые крылья с прожилками | Примитивные палеозойские палеокрылые насекомые |
| Палеоптеры | Палеос ( Палеос , старый) | Старые крылья | Подёнки, стрекозы и несколько отрядов ископаемых. |
| Паранеоптеры | Пара- ( Пара- ) новый ( неос , новый) |
Часть Neoptera, преимущественно с пронзительным ротовым аппаратом. | Настоящие клопы, вши, корые власы, трипсы |
| Фтираптера | Φθείρ ( фтих , вши) ἀ, а- , нет |
Вши без крыльев | Животные вши |
| Плекоптера | Плекеин ( плекеин , складка) | Сложенные крылья | веснянки |
| Многонеоптеры | Полисы ( полисы , много неос новый ) |
Много жилистых крыльев | Neoptera с гемиметаболическим развитием |
| Псокоптера | Ψώχω ( псохо , тереть) | Потирание крыльев | Барклице, книжница |
| Птеригота | Птеригион ( птеригион , крыло) | Крылатые насекомые | В классе, в отличие от Apterygota, имеются крылатые и бескрылые вторичные системы. |
| Рафидиоптеры | ῥαφίς ( рафис , игла) | Игольчатые крылья | Змеи |
| Сифонаптера | Сифон ( сифон , трубка) ἀ- или без |
Бескрылый сифон | Блохи |
| Стрепсиптеры | Стрепсис ( стрепсис , развернуться) | Вращающиеся или скрученные крылья | кривокрылые паразиты |
| Тизаноптера | Тисаной ( thysanoi , бахрома) | Бахрома крылатая | Трипсы |
| Трихоптера | Трихома ( трихома , волосы) | Волосатые крылья | ручейники |
| Зораптера | Ζωρός ( zōros означает сильный) | Сильные крылья | Зораптераны |
| Зигоптера | пара ( zeugos означает пара) | Парные крылья | Стрекоза |
Адаптации
[ редактировать ]Вариация
[ редактировать ]Крылья насекомых имеют основополагающее значение для идентификации и классификации видов, поскольку не существует другого набора структур, более значимого для изучения насекомых. Каждый отряд и семейство насекомых имеет отличительные формы и особенности крыльев. Во многих случаях даже виды можно отличить друг от друга по цвету и рисунку. Например, только по положению можно идентифицировать виды, хотя и в гораздо меньшей степени. Хотя большинство насекомых складывают крылья в состоянии покоя, стрекозы и некоторые стрекозы отдыхают, раскинув крылья горизонтально, в то время как такие группы, как ручейники , веснянки , ольхи и златоглазки , держат крылья над спиной, наподобие крыши. Некоторые мотыльки обвивают крыльями свое тело, тогда как многие мухи и большинство бабочек смыкают крылья прямо вверх над спиной. [ 48 ]
Во многих случаях форма крыльев коррелирует с типом полета насекомых. У лучших летающих насекомых обычно длинные и тонкие крылья. У многих видов Sphingidae (мотыльков-сфинксов) передние крылья большие и остро заостренные, образующие с маленькими задними крыльями треугольник, напоминающий крылья быстрых современных самолетов. Другая, возможно, более важная корреляция — это зависимость размера и силы мышц от скорости и силы полета. У мощно летающих насекомых крылья наиболее приспособлены к нагрузкам и аэродинамике полета. Жилки толще, сильнее и ближе друг к другу по направлению к переднему краю (или «переднему краю») и тоньше, но гибкие по направлению к заднему краю (или «заднему краю»). Это делает крыло насекомого превосходно сконструированным аэродинамическим профилем, способным развивать как тягу, так и подъемную силу, при этом минимизируя сопротивление . [ 48 ]
Также могут наблюдаться различия в взмахе крыльев не только у разных видов, но даже у особей в разное время. В общем, частота зависит от соотношения мощности мышц крыльев и сопротивления нагрузки. Бабочки с большими крыльями и легким телом могут иметь частоту взмахов крыльев 4–20 в секунду, тогда как мухи с маленькими крыльями и тяжелым телом и пчелы совершают взмахи крыльями более 100 раз в секунду, а комары могут взмахивать крыльями до 988–1046 раз. секунда. То же самое касается полета; хотя обычно сложно оценить скорость полета насекомых, большинство насекомых, вероятно, в природе могут летать быстрее, чем в контролируемых экспериментах. [ 48 ]
Жесткокрылые
[ редактировать ]У видов жесткокрылых ( жуков ) единственными функциональными крыльями являются задние. Задние крылья длиннее надкрылий , сложенные продольно и поперечно под надкрыльями. Крыло поворачивается вперед на основании в полетное положение. В результате этого действия крыло расправилось и развернулось в продольном и поперечном направлении. В конструкции крыла имеется пружинный механизм, иногда с помощью движения брюшка, удерживающий крыло в сложенном положении. Жилкование крыльев жуков редуцировано и видоизменено за счет складчатого строения, к которому относятся: [ 49 ]
| Крестообразные складки в крыльях жуков | |

|
Заднее крыло расправлено: линиями сгиба оно разделено на пять полей, каждое из которых завершается сзади. |

|
То же крыло, сложенное наполовину: два сустава поперечного сгиба образуют тупой угол. Правая уже в сложенных в три слоя крыльях. При большем разрешении видна третья арка окантовки крыла на первом и втором. Слева от пятой арки появляется четвертая. |

|
То же крыло, полностью сложенное: пять полей выровнены (надкрылья удалены). |
- Costa (C) , Subcosta posterior (ScP) – у края переднего крыла, сросшиеся на большей части длины.
- Передний радиус (RA) – разделен на две ветви за серединой крыла.
- Задняя лучевая кость (RP) – базальное соединение утрачено.
- Media posterior (MP) – разветвления, длинная и сильная жилка.
- передний локоть (CuA)
- Анальные вены (АА, АР) – вены позади локтевой кости, разделенные анальной складкой.
У большинства видов жуков передняя пара крыльев модифицирована и склеротизирована (затвердела), образуя надкрылья , и они защищают нежные задние крылья, сложенные внизу. [ 33 ] Надкрылья соединены с крылогруди; его называют так, потому что именно здесь соединяются крылья ( птерон по-гречески означает «крыло»). Надкрылья не используются для полета , но имеют тенденцию закрывать заднюю часть тела и защищать вторую пару крыльев ( крылья ). Надкрылья должны быть подняты, чтобы приводить в движение задние маховые крылья. Летные крылья жука пересечены жилками, после приземления складываются, часто вдоль этих жилок, и располагаются под надкрыльями. У некоторых жуков утрачена способность летать. К ним относятся некоторые жужелицы (семейство Carabidae) и некоторые «настоящие долгоносики» (семейство Curculionidae ), а также некоторые пустынные и пещерные виды других семейств. У многих из этих видов два надкрылья слиты вместе, образуя прочный щит над брюшком. В некоторых семьях утрачены как способность летать, так и надкрылья, наиболее известным примером являются светлячки семейства Phengodidae , у которых самки имеют личиночную форму . на протяжении всей жизни [ 13 ] [ 33 ]
Чешуекрылые
[ редактировать ]Две пары крыльев находятся на среднем и третьем сегментах, или среднегруди и заднегруди соответственно. У более поздних родов крылья второго сегмента гораздо более выражены, однако у некоторых более примитивных форм крылья обоих сегментов одинаковы по размеру. Крылья покрыты чешуйками, расположенными в виде черепицы, образующими необычайное разнообразие цветов. Среднегруди эволюционировала, чтобы иметь более мощные мышцы, позволяющие перемещать мотылька или бабочку по воздуху, при этом крыло указанного сегмента имеет более прочную структуру жилок. [ 33 ] : 560 У крупнейшего надсемейства Noctuidae крылья модифицированы так, что они действуют как тимпанальные органы или органы слуха. [ 50 ] Изменения жилкования крыла включают: [ 49 ]
- Коста (C) – не встречается в «Бабочках».
- Подкоста (Sc) + Radius 1 (Sc+R1) – у переднего края крыла сросшаяся или очень сближенная на большей части длины, в заднем крыле сросшаяся и хорошо развитая в плечевой области, у бабочки подкоста никогда не разветвляется.
- Радиус (R2-R5) - радиус делится на ветви за серединой крыла, у Papilionidae до пяти ветвей. На переднем крыле у всех бабочек, за исключением Hesperiidae, последняя буква R имеет ножку.
- Сектор радиуса (Rs) – в заднем крыле.
- Медиа (М1-М3) – базальный отдел утрачен.
- Передний кубитус (CuA1-CuA2) – участок CuP утерян.
- Анальные вены (А, 1А+2А, 3А) – либо одна вена А, либо две вены 1А+2А, 3А.
- Плечевая вена . На заднем крыле большинства бабочек имеется плечевая вена, за исключением Lycaenidae. Плечевая область заднего крыла увеличена и перекрывается с передним крылом. Плечевая вена укрепила область перекрытия заднего крыла, чтобы два крыла лучше соединялись.
Крылья, головные части груди и брюшка чешуекрылых покрыты мелкой чешуей , от которой отряд чешуекрылых получил свое название, слово «лептерон» на древнегреческом языке означает «чешуйка». Большинство чешуек пластинчатые или пластинчатые и прикреплены к ножке, в то время как другие формы могут быть похожими на волосы или специализироваться как вторичные половые признаки. [ 51 ] Просвет, или поверхность ламели, имеет сложное строение. Он придает цвет либо за счет содержащихся внутри пигментных красок, либо за счет своей трехмерной структуры . [ 52 ] Чешуя выполняет ряд функций, в том числе изоляцию, терморегуляцию, помощь в планирующем полете, среди прочих, наиболее важной из которых является большое разнообразие ярких или нечетких рисунков, которые они обеспечивают, которые помогают организму защитить себя путем маскировки , мимикрии и поиска. приятели. [ 51 ]
Одоната
[ редактировать ]Оба вида Odonata (стрекозы и стрекозы) имеют по две пары крыльев примерно одинакового размера и формы и прозрачного цвета. Их пять, если R+M считать за 1, на крыльях стрекозы и стрекозы отходят основные жилки, жилки крыльев слиты у их оснований и крылья не могут складываться над телом в состоянии покоя, к ним также относятся: [ 49 ]
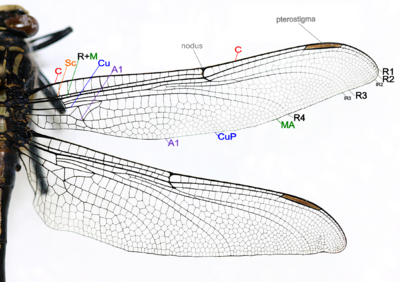

- Коста (С) – у передней кромки крыла, сильная и краевая, доходит до вершины крыла.
- Subcosta (Sc) – вторая продольная жилка, неразветвленная, присоединяется к С в узле.
- Радиус и медиа (R+M) – третья и четвертая продольные жилки, самые сильные жилки на крыле, с ветвями R1-R4 доходят до края крыла, передние медиа (МА) также доходят до края крыла. IR2 и IR3 — вставочные вены позади R2 и R3 соответственно.
- Cubitus (Cu) – пятая продольная жилка, Cubitus posterior (CuP) неразветвленная и доходит до края крыла.
- Анальные вены (А1) – неразветвленные вены позади локтевой кости.
- Узел образуется там, где вторая главная жилка встречается с передней кромкой крыла. Черная птеростигма расположена возле кончика крыла.
Основные жилки и поперечные жилки образуют рисунок жилкования крыла. Схема жилкования у разных видов различна. Поперечных жилок может быть очень много или, скорее, мало. Крылья австралийской плоскокрылой стрекозы представляют собой один из немногих узоров жилок. Характер жилкования полезен для идентификации вида. [ 49 ] Почти все Anisoptera располагаются с крыльями, откинутыми в сторону или немного вниз, однако большинство Zygoptera располагаются со скрепленными крыльями и соприкасающимися дорсальными поверхностями. Грудная клетка Zygoptera настолько наклонена, что крылья аккуратно прилегают к верхней части брюшка. Кажется, что они не держатся прямо вверх, как у бабочек или подёнок. У некоторых семейств двукрылых крылья в состоянии покоя располагаются горизонтально, а у одного рода бескрылых (например, Cordulephya , Corduliidae ) крылья удерживаются в типичном для стрекоз положении покоя. Взрослые виды обладают двумя парами равных или почти равных крыльев. Кажется, существует только пять основных стволов вен. Узелок образуется в месте соприкосновения второй главной жилки (подкосты) с передней кромкой крыла. У большинства семейств возле кончика крыла имеется заметная птеростигма. Идентификация Odonata может быть основана на жилковании. Единственная вероятная путаница связана с некоторыми златоглазками (отряд Neuroptera), у которых на крыльях много поперечных жилок. До начала 20-го века Odonata часто считались родственниками златоглазок и получали порядковое название Paraneuroptera, но любое сходство между этими двумя отрядами является совершенно поверхностным. У Anisoptera заднее крыло шире переднего, и в обоих крыльях поперечная жилка разделяет дискоидальную клетку на треугольник и супертреугольник. [ 53 ]
Прямокрылые
[ редактировать ]
У видов прямокрылых (кузнечиков и сверчков) передние крылья представляют собой жесткие непрозрачные надкрылья, узкие, которые обычно закрывают задние крылья и брюшко в состоянии покоя. Задние крылья пластинчато-перепончатые, сложенные веерообразно, имеют следующее жилкование: [ 49 ]
- Коста (С) – по переднему краю переднего и заднего крыла, неразветвленный.
- Subcosta (Sc) – вторая продольная жилка, неразветвленная.
- Радиус (R) – третья продольная жилка, разветвляющаяся на Rs на переднем и заднем крыле.
- Media anterior (MA) – четвертая продольная жилка, разветвленная в базальной части как Media posterior (MP).
- Cubitus (Cu) - пятая продольная жилка, на переднем и заднем крыле разделяющаяся у основания крыла на разветвленную CuA и неразветвленную CuP.
- Анальные жилки (А) – жилки позади локтевого сустава, неразветвленные, две на переднем крыле, много на заднем крыле.
Фазматода
[ редактировать ]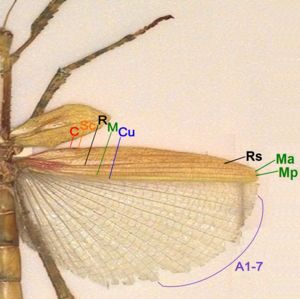
- Коста (С) – на переднем крае заднего крыла, неразветвленная, на переднем крыле отсутствует.
- Subcosta (Sc) – вторая продольная жилка, неразветвленная.
- Радиус (R) – третья продольная жилка, разветвленная до Rs на заднем крыле, неразветвленная на переднем крыле.
- Media anterior (MA) – четвертая продольная жилка, разветвленная в базальной части как Media posterior (MP).
- Локоть (Cu) – пятая продольная жилка, неразветвленная.
- Анальные жилки (А) – жилки позади локтевого сустава, неразветвленные, две на переднем крыле, много на заднем крыле 1А-7А в одной группе, остальные в другой группе.
У палочника передние крылья жесткие, непрозрачные надкрылья, короткие и покрывают только основание задних крыльев в состоянии покоя. Задние крылья от Коста до Кубитуса жесткие и непрозрачные, как и передние. Большая анальная область перепончатая и сложена веерообразно. В жилках крыльев палочников нет или очень мало разветвлений. [ 49 ]
Дермаптера
[ редактировать ]| Разворачивание крыла уховертки | |

|
Передние и задние крылья в состоянии покоя: переднее крыло покрывает большую часть заднего крыла, только суставы выступают вперед в виде четверти круга с белым пятном в центре под передним крылом. С правой стороны переднее крыло открыто вправо (синяя стрелка), что с этой точки зрения кажется уже, чем при полностью сложенном заднем крыле. . |

|
Переднее крыло открыто влево (синяя стрелка), правая часть переднего крыла удалена; заднее крыло полуоткрыто. С большим разрешением показано многократное сгибание, напоминающее веер, параллельный линиям b и c. Стрелка указывает на точку e, где вентилятор снова закрывается, сложившись на 180°. . |
Другие отряды, такие как Dermaptera ( уховертки ), Orthoptera ( кузнечики , сверчки ), Mantodea ( богомолы ) и Blattodea ( тараканы ), имеют жесткие кожистые передние крылья, которые не взмахивают во время полета, иногда называемые надкрыльями (мн. tegmina ), надкрыльями (мн. ) надкрылья , или псевдоэлитрон . [ 13 ]
полужесткокрылые
[ редактировать ]У Hemiptera (настоящие клопы) передние крылья могут быть затвердевшими, хотя и в меньшей степени, чем у жуков. передняя часть передних крыльев Например, у клопов-вонючек затвердевшая, а задняя — перепончатая. Их называют гемелитроном (мн.ч. hemelytra ). Они встречаются только в подотряде Heteroptera ; крылья равнокрылых , таких как цикады , обычно полностью перепончатые. И передние, и задние крылья цикады перепончатые. Большинство видов похожи на стекло, хотя некоторые из них непрозрачны. Цикады не очень хорошие летчики, и большинство из них летают всего несколько секунд. В полете переднее и заднее крыло соединяются друг с другом рифленой муфтой вдоль ребра и края переднего крыла. Большинство видов имеют основное жилкование, как показано на следующем рисунке. [ 49 ]
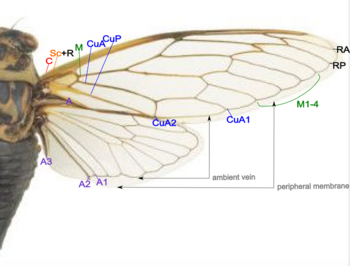
- Коста (C) – на переднем крае крыла, на переднем крыле доходит до узла и лежит близко к Sc+R.
- Subcosta + Radius (Sc+R) – на переднем крыле Sc и R срослись в узел. Радиальный сектор (Rs) возникает вблизи узла и разветвляется.
- Передний радиус (RA)
- Задний радиус (RP)
- Медиа (М) – разветвляется от М1 до М4.
- Cubitus anterior (CuA) – разветвляется на CuA1 и CuA2.
- Задний локоть (CuP) – неразветвляется.
- Анальные жилки (А) – жилки позади локтевой кости, 1А и 2А слиты в переднем крыле, CuP и 2А складчаты.
Также обратите внимание на окружающие вены и периферические мембраны по краям обоих крыльев.
двукрылые
[ редактировать ]У двукрылых ( настоящих мух ) имеется только одна пара функциональных крыльев, при этом задняя пара крыльев редуцирована до жужжальца , которые помогают мухе чувствовать свою ориентацию и движение, а также улучшать баланс, действуя подобно гироскопам. . У Calyptratae самая задняя часть крыльев видоизменена в несколько утолщенные лоскуты, называемые калиптерами , которые покрывают жужжальца. [ 49 ]

- Коста (C) – у двукрылых не встречается.
- Субкоста (Sc) – стала ведущей жилкой крыла, неразветвленной.
- Радиус (R) – разветвлен на R1-R5.
- Медиа (М) – разветвлена на М1-М4.
- Cubitus anterior (CuA) – неразветвленный, у Diptera CuP редуцирован. Некоторые виды CuA и 1A разделяются, некоторые виды встречаются при достижении края крыла, некоторые виды сливаются.
- Анальные жилки (А) - присутствуют только две анальные жилки 1А и 2А, у некоторых видов 2А не выражена.
- Дискальная клетка (dc) – хорошо выражена у большинства видов.
Блаттодея
[ редактировать ]У видов Blattodea ( тараканов ) переднее крыло, также известное как тегмен, более или менее склеротизировано. Он используется в полете, а также в качестве защиты перепончатых задних крыльев. Жилки заднего крыла примерно такие же, как переднего, но с большой анальной лопастью, сложенной в состоянии покоя между CuP и 1А. Анальная лопасть обычно сложена веерообразно. [ 49 ]

- Коста (С) – на передней кромке крыла.
- Subcosta (Sc) – вторая продольная жилка, относительно короткая.
- Радиус (R) – третья продольная жилка со множеством гребенчатых ответвлений.
- Медиа (М) – четвертая продольная жилка, доходит до края крыла.
- Cubitus anterior (CuA) – пятая продольная жилка, с дихотомическими ветвями, занимающими большую часть надкрылья.
- Cubitus posterior (CuP) – неразветвленный, изогнутый, достигает края крыла.
- Анальные вены (А) – вены позади локтевой кости.
Перепончатокрылые
[ редактировать ]| Пример продольной складчатости у ос (Vespidae) | |

|
Основная линия сгиба переднего крыла видна на полпути вверх как яркая горизонтальная линия. Часть крыла, находящаяся за этой линией, повернута обратно вниз. Узкая полоска у передней кромки крыла находится перед первой прочной проволокой, загнутой вперед и вниз. |

|
Таким образом, в состоянии покоя внешняя оболочка образует прочный внешний край крыла, который защищает боковые части брюшка как амортизатор. Заднее крыло в значительной степени прикрыто передним крылом. |
У взрослых перепончатокрылых , включая пилильщиков, ос, пчел и нерабочих муравьев, есть две пары перепончатых крыльев. [ 49 ]
- Коста (C) – у перепончатокрылых не встречается.
- Субкоста (Sc) – неразветвленная.
- Радиус (R) – разветвлен на R1-R5.
- Медиа (М) – М неразветвленная, в переднем крыле М на части длины слита с Rs.
- Cubitus (CuA) – неразветвленный, CuP отсутствует у перепончатокрылых.
- Анальные жилки (А) - присутствуют только две анальные жилки 1А и 2А, у некоторых видов 2А не выражена.
- Соединение крыльев . Ряд крючков на передней кромке заднего крыла зацепляется за задний край переднего крыла, прочно соединяя крылья в полете.
- Линия складывания крыла . У некоторых видов, в том числе у Vespidae, переднее крыло в состоянии покоя сложено продольно вдоль «линии складывания крыла».
- Птеростигма – присутствует у некоторых видов.
На переднем крае заднего крыла имеется несколько крючковатых щетинок, или « хамули », которые прикрепляются к переднему крылу, удерживая их вместе. У более мелких видов может быть только два или три хамули с каждой стороны, но у самых крупных ос их может быть значительное количество, поэтому крылья сжимаются особенно плотно. Крылья перепончатокрылых имеют относительно мало жилок по сравнению со многими другими насекомыми, особенно у более мелких видов. [ 13 ]
Другие семьи
[ редактировать ]Термиты относительно плохо летают, и их легко уносит по ветру при скорости ветра менее 2 км/ч, они сбрасывают крылья вскоре после приземления в приемлемом месте, где они спариваются и пытаются сформировать гнездо во влажной древесине или земле. [ 54 ] Крылья большинства термитов имеют три тяжелые жилки вдоль базальной части переднего края переднего крыла, а поперечные жилки возле кончика крыла расположены под углом, образуя трапециевидные ячейки. Хотя крылья подземных термитов имеют только две основные жилки вдоль переднего края переднего крыла, а поперечные жилки, ведущие к кончику крыла, перпендикулярны этим жилкам, образуя квадратные и прямоугольные клетки. [ 55 ]
Виды Thysanoptera ( трипсы ), Ptiliidae и другие летающие микронасекомые имеют тонкие передние и задние крылья с длинной бахромой из волос, называемые крыльями с бахромой, также называемые птилоптериями. [ 56 ] А виды Trichoptera ( ручейники ) имеют волосистые крылья, передние и задние крылья покрыты щетинками. [ 13 ]
У самцов Strepsiptera также есть жужжальца , которые развились из передних крыльев, а не из задних. Это означает, что только их задние крылья способны летать, в отличие от двукрылых , у которых есть функциональные передние крылья и жужжальца вместо задних крыльев. Также задние крылья у самцов Coccidae редуцированы до жужжальца (или отсутствуют). [ 57 ]
См. также
[ редактировать ]- Придаток
- Система Комстока-Нидхэма
- Крыло
- Роботы, вдохновленные насекомыми: RoboBee , DelFly
Примечания
[ редактировать ]- ^ Крэмптон, Г. (1916). «Филогенетическое происхождение и природа крыльев насекомых согласно паранотальной теории». Журнал Нью-Йоркского энтомологического общества . 24 (1): 1–39. JSTOR 25003692 .
- ^ Перейти обратно: а б Росс, Эндрю (2017). «Эволюция насекомых: происхождение крыльев» . Современная биология . 27 (3): Р113–Р115. дои : 10.1016/j.cub.2016.12.014 . PMID 28171756 – через Web of Science.
- ^ Перейти обратно: а б Авероф, Михалис и С.М. Коэн. (1997). «Эволюционное происхождение крыльев насекомых из предковых жабр». Природа . 385 (6617): 627–630. Бибкод : 1997Natur.385..627A . дои : 10.1038/385627a0 . ПМИД 9024659 . S2CID 4257270 – через Web of Science.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гродницкий, Дмитрий, Л. (1999). Форма и функции крыльев насекомых: эволюция биологических структур . стр. 82–83.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с Александр, Дэвид, Э. (2015). На крыле: насекомые, птерозавры, птицы, летучие мыши и эволюция полета животных . Издательство Оксфордского университета. стр. 74–101.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хауг, Иоахим, К. Хауг и Р. Дж. Гарвуд. (2016). «Эволюция и развитие крыльев насекомых – новые подробности палеозойских нимф» . Биологические обзоры . 91 (1): 53–69. дои : 10.1111/brv.12159 . ПМИД 25400084 . S2CID 21031689 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Альмуди, Изабель; Визуэта, Джоэл; Вятт, Кристофер Д.Р.; де Мендоса, Алекс; Марлетаз, Фердинанд; Фирбас, Панос Н.; Феуда, Роберто; Масьеро, Джулио; Медина, Патрисия; Алькаина-Каро, Ана; Круз, Фернандо (2020). «Геномная адаптация к водной и воздушной жизни у подёнок и происхождение крыльев насекомых» . Природные коммуникации . 11 (1): 2631. Бибкод : 2020NatCo..11.2631A . дои : 10.1038/s41467-020-16284-8 . ISSN 2041-1723 . ПМК 7250882 . ПМИД 32457347 .
- ^ Брюс, Хизер, Н. Х. Патель. (2020). «Нокаут генов, формирующих рисунок ног ракообразных, позволяет предположить, что крылья и стенки тела насекомых произошли от древних сегментов ног». Экология и эволюция природы . 4 (12): 1703–1712. Бибкод : 2020NatEE...4.1703B . дои : 10.1038/s41559-020-01349-0 . ПМИД 33262517 . S2CID 227253368 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кларк-Хэтчел, Кортни (2013). «Понимание происхождения крыльев насекомых получено с помощью функционального анализа рудиментов красного мучного жука Tribolium castaneum» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (42): 16951–16956. Бибкод : 2013PNAS..11016951C . дои : 10.1073/pnas.1304332110 . ПМК 3801059 . ПМИД 24085843 .
- ^ Прокоп, Якуб, Пехарова М., Нел А., Хёрншемейер Т., Кшеминьска Э., Кжеминьский В. и Энгель М. (2017). «Палеозойские подушечки крыльев нимф подтверждают двойную модель происхождения крыльев насекомых» . Современная биология . 27 (2): 263–269. дои : 10.1016/j.cub.2016.11.021 . ПМИД 28089512 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с д и ж Чепмен, РФ (1998). Насекомые: структура и функции (4-е изд.). Кембридж, Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-57048-4 .
- ^ Перейти обратно: а б Гиллиотт, Седрик (август 1995 г.). Энтомология (2-е изд.). Спрингер-Паблишерс Нью-Йорк, ООО. ISBN 0-306-44967-6 .
- ^ Перейти обратно: а б с д и Мейер, Джон Р. (5 января 2007 г.). «Внешняя анатомия: КРЫЛЬЯ» . Кафедра энтомологии Университета штата Северная Каролина. Архивировано из оригинала 16 июля 2011 года . Проверено 21 марта 2011 г.
- ^ Перейти обратно: а б с д Гуллан, Пи Джей; Крэнстон, PS (2004). Насекомые: Очерк энтомологии . Великобритания: Издательство Блэквелл. п. 42 . ISBN 1-4051-1113-5 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Снодграсс, Р.Э. (декабрь 1993 г.). Основы морфологии насекомых . Корнельский университет Пресс. ISBN 0-8014-8125-2 .
- ^ Спит, ХТ (1932). «Новый метод изучения жилок крыльев поденок и некоторые результаты (Эфемериды)» (PDF) . Энтомологические новости . Архивировано из оригинала (PDF) 30 сентября 2011 г.
- ^ «РАЗЪЯСНЕНИЕ ТЕРМИНОВ, ИСПОЛЬЗУЕМЫХ В ЭНТОМОЛОГИИ» . www.gutenberg.org . Проверено 08 января 2024 г.
- ^ «Определение ТЕРМЕНА» . www.merriam-webster.com . Проверено 08 января 2024 г.
- ^ Перейти обратно: а б Кноспе, Карл Р. (осень 1998 г.). «Механизмы полета насекомых: анатомия и кинематика» (PDF) . Машиностроение и аэрокосмическая инженерия, Университет Вирджинии.
- ^ Вейс-Фог, Т. (июль 1963 г.). «Диффузия в мышцах крыльев насекомых, самой активной из известных тканей» . J Exp Biol . 41 (2): 229–256. дои : 10.1242/jeb.41.2.229 . ПМИД 14187297 .
- ^ Тигс, Огайо (февраль 1954 г.). «Летающие мышцы насекомых - их анатомия и гистология; с некоторыми наблюдениями о строении поперечнополосатых мышц в целом» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 238 (656): 221–348. Бибкод : 1955РСТБ.238..221Т . дои : 10.1098/rstb.1955.0001 . JSTOR 3739600 .
- ^ Он, Же; Ло, Ичен; Шан, Сюэин; Сан, Дженнифер С.; Карлсон, Джон Р. (21 мая 2019 г.). «Хемосенсорные сенсиллы крыла дрозофилы экспрессируют кандидатный рецептор ионотропного феромона» . ПЛОС Биология . 17 (5): e2006619. дои : 10.1371/journal.pbio.2006619 .
- ^ Вальмалетт, Дж. К., Раад, Х., Цю, Н., Охара, С., Каповилла, М. и Робишон, А., 2015. Наноархитектура вкусовых хемосенсорных щетинок и трахеи в крыльях дрозофилы. Научные отчеты, 5(1), стр.1-11.
- ^ Дингес, Г.Ф., Чокли, А.С., Бокемюль, Т., Ито, К., Бланке, А. и Бюшгес, А., 2021. Расположение и расположение колокольчатых сенсилл у Drosophila melanogaster. Журнал сравнительной неврологии, 529 (4), стр. 905-925.
- ^ Филд, Л.Х. и Мэтисон, Т., 1998. Хордотональные органы насекомых. В книге «Достижения в области физиологии насекомых» (том 27, стр. 1–228). Академическая пресса.
- ^ Вольф, Х., 1993. Тегула саранчи: значение для формирования ритма полета, управления движением крыльев и создания аэродинамической силы. Журнал экспериментальной биологии, 182 (1), стр. 229–253.
- ^ Чжан, Н. и Симпсон, Дж. Х., 2022. Пара комиссуральных командных нейронов вызывает уход за крыльями дрозофилы. Исайенс, 25(2), стр.103792.
- ^ Перейти обратно: а б Чепмен, AD (2006). Численность живых видов в Австралии и мире . Канберра: Исследование биологических ресурсов Австралии . стр. 60стр. ISBN 978-0-642-56850-2 . Архивировано из оригинала 19 мая 2009 г. Проверено 18 июня 2012 г.
- ^ Перейти обратно: а б Ван, З. Джейн (2005). «Рассекающий полет насекомых» (PDF) . Ежегодный обзор механики жидкости . 37 (1). Годовые обзоры: 183–210. Бибкод : 2005AnRFM..37..183W . doi : 10.1146/annurev.fluid.36.050802.121940 .
- ^ Перейти обратно: а б Сане, Санджай П. (2003). «Аэродинамика полета насекомых» (PDF) . Журнал экспериментальной биологии . 206 (23): 4191–4208. дои : 10.1242/jeb.00663 . ПМИД 14581590 . S2CID 17453426 .
- ^ Давидовиц, Павел (2008). Физика в биологии и медицине . Академическая пресса. стр. 78–79. ISBN 978-0-12-369411-9 .
- ^ Перейти обратно: а б с д и ж г час я Гримальди, Дэвид; Энгель, Майкл С. (2005). Эволюция насекомых . Нью-Йорк, штат Нью-Йорк: Издательство Кембриджского университета.
- ^ Перейти обратно: а б с д и Пауэлл, Джерри А. (2009). «Жестококрылые» . В Реше, Винсент Х.; Карде, Ринг Т. (ред.). Энциклопедия насекомых (2-е (иллюстрированное) изд.). Академическая пресса. п. 1132. ИСБН 978-0-12-374144-8 .
- ^ Майкл С. Энгель; Дэвид А. Гримальди (2004). «Новый свет, пролитый на старейшее насекомое». Природа . 427 (6975): 627–630. Бибкод : 2004Natur.427..627E . дои : 10.1038/nature02291 . ПМИД 14961119 . S2CID 4431205 .
- ^ Хауг, Кэролин; Хауг, Иоахим Т. (30 мая 2017 г.). «Предположительно старейшее летающее насекомое: скорее многоножка?» . ПерДж . 5 : е3402. дои : 10.7717/peerj.3402 . ISSN 2167-8359 . ПМЦ 5452959 . ПМИД 28584727 .
- ^ Гржимек Х.К. Бернхард (1975) Энциклопедия жизни животных Гржимека, том 22, насекомые. Ван Ностранд Рейнхольд Ко. Нью-Йорк.
- ^ Перейти обратно: а б Риек Э. Ф. Кукалова-Пек Дж (1984). «Новая интерпретация жилкования крыльев стрекозы, основанная на окаменелостях раннего верхнего карбона из Аргентины (Insecta: Odonatoida и основные состояния крыльев Pterygote)». Может. Дж. Зул . 62 (6): 1150–1160. дои : 10.1139/z84-166 .
- ^ Уэйклинг Дж. М. Эллингтон CP (1997). «Требования к подъемной силе и мощности Dragonfly Flight III» . Журнал экспериментальной биологии . 200 (Часть 3): 583–600 (589). дои : 10.1242/jeb.200.3.583 . ПМИД 9318294 .
- ^ Мацуда Р. (1970). «Морфология и эволюция грудной клетки насекомых». Память Энтомол. Соц. Может . 102 (76): 1–431. дои : 10.4039/entm10276fv .
- ^ В.А. Благодеров; Э.Д. Лукашевич; М.Б. Мостовский (2002). «Орден двукрылых Линнея, 1758 год. Настоящие мухи». В исследовании AP; DLJ Quicke (ред.). История насекомых Академическое издательство Kluwer . ISBN 1-4020-0026-Х .
- ^ Гегенбаур, Карл (1870). Основы сравнительной анатомии . В. Энгельманн.
- ^ Труман JWH (1990), Комментарий: эволюция крыльев насекомых: модель выхода конечности и эндита Канадский журнал зоологии
- ^ Станичек, А.Х.; Бехли, Г. и Годунко, Р.Дж. (2011). «Coxoplectoptera, новый ископаемый отряд Palaeoptera (Arthropoda: Insecta), с комментариями к филогении стволовой группы подёнок (Ephemeroptera)» (PDF) . Систематика и эволюция насекомых . 42 (2): 101–138. дои : 10.1163/187631211X578406 . S2CID 4986911 . Архивировано из оригинала (PDF) 3 октября 2011 г.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Прокоп, Якуб; Печарова, Мартина; Нел, Андре; Хёрншемейер, Томас; Кшеминьска, Ева; Кшеминский, Веслав; Энгель, Майкл С. (январь 2017 г.). «Палеозойские подушечки крыльев нимф подтверждают двойную модель происхождения крыльев насекомых» . Современная биология . 27 (2): 263–269. дои : 10.1016/j.cub.2016.11.021 . ПМИД 28089512 .
- ^ Адриан Л.Р. Томас; Р. Оке Норберг (1996). «Скольжение по поверхности — происхождение полета насекомых?». Тенденции в экологии и эволюции . 11 (5): 187–188. дои : 10.1016/0169-5347(96)30022-0 . ПМИД 21237803 .
- ^ Яновяк С.П., Каспари М., Дадли Р. (2009). «Скользящие шестиногие и происхождение воздушного поведения насекомых» . Письма по биологии . 5 (4): 510–2. дои : 10.1098/rsbl.2009.0029 . ПМК 2781901 . ПМИД 19324632 .
- ^ Перейти обратно: а б с д Х. Комсток, Генри (1918). Крылья насекомых . Итака, Нью-Йорк: Издательская компания Comstack. п. 114 .
- ^ Перейти обратно: а б с «Крылья насекомых вообще» . Аэродинамика насекомых . Цислунарная аэрокосмическая отрасль. 1997 год . Проверено 28 марта 2011 г.
- ^ Перейти обратно: а б с д и ж г час я дж Чу, Питер (9 мая 2009 г.). «Крылья насекомых» . Брисбенские насекомые и пауки . Проверено 21 марта 2011 г.
- ^ Скобл, М.Дж. (1992). Чешуекрылые: форма, функции и разнообразие . Оксфордский университет. Нажимать. ISBN 978-1-4020-6242-1 .
- ^ Перейти обратно: а б Скобл (1995). Раздел «Весы» , (стр. 63 – 66).
- ^ Вукусич, П. (2006). «Структурная окраска чешуекрылых» (PDF) . Современная биология . 16 (16): R621–3. дои : 10.1016/j.cub.2006.07.040 . ПМИД 16920604 . S2CID 52828850 . Проверено 11 ноября 2010 г.
- ^ Труман, Джон WH; Ричард Дж. Роу (16 октября 2009 г.). "Одоната. Стрекозы и стрекозы" . Tolweb.org . Проверено 21 марта 2011 г.
- ^ Эйб Т., Бигнелл Д.Э.; Хигаши М (2000). Термиты: эволюция, социальность, симбиозы, экология, эколаборатория . Академические издательства Kluwer. ISBN 0-7923-6361-2 .
- ^ «Термит» . Техасская служба распространения знаний AgriLife. Архивировано из оригинала 13 апреля 2011 г.
- ^ Полилов, Алексей А.; Решетникова Наталья Ивановна; Петров Петр Н.; Фарисенков, Сергей Евгеньевич (январь 2019 г.). «Морфология крыльев жуков-перылов (Coleoptera: Ptiliidae): особенности, связанные с миниатюризацией и анализом функционального масштабирования» . Строение и развитие членистоногих . Специальный выпуск: Миниатюризация панартропод. 48 : 56–70. Бибкод : 2019АртСД..48...56П . дои : 10.1016/j.asd.2019.01.003 .
- ^ Камачо, Эрнесто Робайо; Чонг, Хуанг-Хорнг (2015). «Общая биология и современные подходы к борьбе с мягкочешуйными вредителями (Hemiptera: Coccidae)» (PDF) . Журнал комплексной борьбы с вредителями . 6 (1): 17. дои : 10.1093/jipm/pmv016 . ISSN 2155-7470 . ПМЦ 4725186 . ПМИД 26823990 .
Ссылки
[ редактировать ]- Триплхорн, Чарльз А.; Джонсон Норман Ф. (2005). Введение Боррора и Делонга в изучение насекомых (7-е изд.). Томсон Брукс/Коул. ISBN 0-03-096835-6 .
Внешние ссылки
[ редактировать ]- Курс Брисбенского университета по крыльям насекомых
- Курс Университета штата Северная Каролина по крыльям насекомых
- Рисунки крыльев насекомых
| до |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||