Полет насекомого

Насекомые - единственная группа беспозвоночных , которые развили крылья и полеты . Насекомые впервые летали в каменноугольном , около 300-350 миллионов лет назад, что делает их первыми животными, которые развивались. Крылья, возможно, развивались из придаток на боковых сторонах существующих конечностей, у которых уже были нервы, суставы и мышцы, используемые для других целей. Первоначально они могут быть использованы для плавания по воде или для замедления скорости происхождения при скольжении.
Две группы насекомых, стрекозы и майки , имеют мышцы полетов, прикрепленные непосредственно к крыльям. У других крылатых насекомых летные мышцы прикрепляются к грудной клетке, что делает его колебаниями, чтобы побудить крылья бить. Из этих насекомых некоторые ( мухи и некоторые жуки ) достигают очень высоких частот крыла посредством эволюции «асинхронной» нервной системы, в которой грудная клетка колеблется быстрее, чем скорость нервных импульсов.
Не все насекомые способны к полету. Ряд жареных насекомых вторично потеряли свои крылья из -за эволюции , в то время как другие более базальные насекомые, такие как серебряные рыбы, никогда не эволюционировали крылья. У некоторых евросоциальных насекомых, таких как муравьи и термиты , только репродуктивные касты Алат развивают крылья в течение сезона спаривания , а затем проливают свои крылья после спаривания, в то время как члены других каст без крыла их жизни.
Некоторые очень маленькие насекомые используют не стационарную аэродинамику , а из механизма Weis-Fogh Clap и Thring , генерируя большие силы подъема за счет износа на крыльях. Многие насекомые могут парить, поддерживать высоту и контролировать их положение. Некоторые насекомые, такие как мотыльки, имеют передние крылья в сочетании с задними крыльями, поэтому они могут работать в унисон.
Механизмы
[ редактировать ]Прямой полет
[ редактировать ]В отличие от других насекомых, мышцы крыльев эфемероптер (майффи) и одонаты (дракоза и пломли) вставляют непосредственно на основания крыла, которые зависают так, чтобы небольшое нисходящее движение основания крыла поднимает само крыло, очень похожее на греблю через воздух. Драконы и самого себя имеют передние и задние крылья, похожие по форме и размеру. Каждый работает независимо, что дает степень тонкого контроля и мобильности с точки зрения резкости, с которой они могут изменить направление и скорость, не видимые у других летающих насекомых. Одонаты - все это воздушные хищники, и они всегда охотились на других воздушных насекомых. [ 1 ]
-
 Прямой полет: мышцы, прикрепленные к крыльям. Только большие насекомые
Прямой полет: мышцы, прикрепленные к крыльям. Только большие насекомые -
 Одоната (стрекозы и самодовольные ) имеют прямую мускулатуру прямого полета, как и Mayflies .
Одоната (стрекозы и самодовольные ) имеют прямую мускулатуру прямого полета, как и Mayflies . -
Медленное движение Dragonfly


Косвенный полет
[ редактировать ]Помимо двух заказов с прямыми летными мышцами, все остальные живые крылатые насекомые летают, используя другой механизм, включающий косвенные мышцы полета. Этот механизм развивался один раз и является определяющей особенностью ( синапоморфий ) для инфракласных неоптер ; Это соответствует, вероятно, не случайно, с появлением механизма сшивания крыльев, который позволяет неоптерановым насекомым складывать крылья обратно через живот, когда они находятся в состоянии покоя (хотя эта способность была потеряна вторичным в некоторых группах, таких как бабочки ) Полем [ 1 ]
Тем не менее, все неоптери, так это то, как мышцы в грудной клетке работают: эти мышцы, а не прикрепляются к крыльям, прикрепляются к грудной клетке и деформируются; Поскольку крылья являются расширениями грудного экзоскелета , деформации грудной клетки также вызывают движения крыльев. Набор продольных мышц вдоль спины сжимает грудную клетку спереди к спине, вызывая дорсальную поверхность грудной клетки ( notum ), чтобы поклониться вверх, заставляя крылья сбиваться вниз. Еще один набор мышц от тергума до грудины снова тянет ноту вниз, в результате чего крылья переворачиваются вверх. [ 1 ] [ 2 ]
-
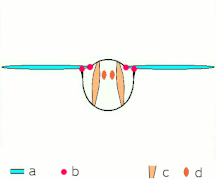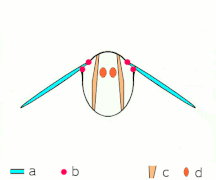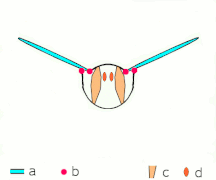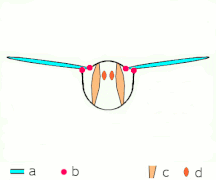 Косвенное полете: мышцы делают грудь колебаться у большинства насекомых
Косвенное полете: мышцы делают грудь колебаться у большинства насекомых -
 Неоптера и большинство , включая бабочек других насекомых, имеют косвенную мускулатуру полета
Неоптера и большинство , включая бабочек других насекомых, имеют косвенную мускулатуру полета


Насекомые, которые побеждают свои крылья менее чем в сто раз в секунду, используют синхронную мышцу. Синхронная мышца - это тип мышц, который сокращается один раз для каждого нервного импульса. Обычно это вызывает меньшую мощность и является менее эффективным, чем асинхронная мышца, которая учитывает независимую эволюцию асинхронных мышц полета в нескольких отдельных кладах насекомых. [ 3 ]
Насекомые, которые бьют их крылья быстрее, такие как шмель , используют асинхронную мышцу; Это тип мышцы, который сокращается более одного раза на нервный импульс. Это достигается тем, что мышцы стимулируются, чтобы снова сокращаться путем высвобождения в напряжении в мышцах, что может произойти быстрее, чем за счет простой нервной стимуляции. [ 4 ] Это позволяет частоте битов крыла превышать скорость, с которой нервная система может отправлять импульсы. Асинхронная мышца является одним из последних уточнений, которые появились в некоторых высших неоптерах ( Coleoptera , Diptera и Hymenoptera ). Общий эффект заключается в том, что многие более высокие неоптери могут победить свои крылья намного быстрее, чем насекомые с прямыми мышцами лета. [ 1 ]
Асинхронная мышца, по определению, под относительно грубым контролем нервной системы. Чтобы сбалансировать этот эволюционный компромисс, насекомые, развивающиеся косвенные полеты, также разработали отдельную нервно-мышечную систему для мелкозернистого контроля над крыльями. [ 5 ] Известные как «прямые мышцы», эти мышцы прикрепляются непосредственно к склеритам, которые составляют шарнир крыла и сокращаются с импульсами 1: 1 от моторных нейронов. [ 6 ] Недавняя работа начала решать сложную нелинейную мышечную динамику в шарнире крыла и ее влияние на путь к крыла. [ 7 ]
Аэродинамика
[ редактировать ]Существуют две основные аэродинамические модели полета насекомых: создание вихря переднего края и использование CLAP и броска. [ 8 ] [ 9 ]
Ведущий вихрек
[ редактировать ]Большинство насекомых используют метод, который создает спирального переднего края вихрь . Эти хлопающие крылья проходят через два основных полусущих. Вниз запускается и обратно и погружается вниз и вперед. Затем крыло быстро переворачивается ( супинация ), так что передний край направлен назад. Затем вверх ударил крыло вверх и назад. Затем крыло снова переворачивается ( пронация ), и может произойти еще один удар. Частотный герц диапазон у насекомых с синхронными летными мышцами обычно составляет от 5 до 200 ( Гц). У тех, у кого асинхронные летные мышцы частота взрыва крыла может превышать 1000 Гц. Когда насекомое колеблется, два удара занимают одинаковое количество времени. Однако более медленный нагрузочный удар обеспечивает тягу . [ 8 ] [ 9 ]
Выявление основных сил имеет решающее значение для понимания полета насекомых. Первые попытки понять хлопающие крылья предположили квази-устроенное состояние. Это означает, что воздушный поток через крыло в любое время предполагалось, что поток будет превышать неподвижное, стабильное крыло под тем же углом атаки. Разделяя взмахивающее крыло на большое количество неподвижных позиций, а затем анализируя каждую позицию, можно было бы создать график мгновенных сил на крыле в каждый момент. Было обнаружено, что рассчитанный подъемник был слишком мал в три, поэтому исследователи поняли, что должны быть неустойчивые явления, обеспечивающие аэродинамические силы. Было несколько разработанных аналитических моделей, пытающихся приблизиться к потоку, близко к хлопчивому крылу. Некоторые исследователи предсказывали силу пики в супинации. С динамически масштабированной моделью фруктовой мухи эти предсказанные силы позже были подтверждены. Другие утверждали, что пики силы во время супинации и пронации вызваны неизвестным вращательным эффектом, который в основном отличается от трансляционных явлений. Есть некоторые разногласия с этим аргументом. Через Вычислительная динамика жидкости , некоторые исследователи утверждают, что нет никакого вращательного эффекта. Они утверждают, что высокие силы вызваны взаимодействием с пробуждением предыдущего инсульта. [ 8 ] [ 9 ]
Аналогично эффекту вращения, упомянутому выше, явления, связанные с хлопающими крыльями, не совсем понятны и не согласованы. Поскольку каждая модель является приближением, разные модели не позволяют эффектам, которые считаются незначительными. Например, эффект Вагнера , предложенный Гербертом А. Вагнером в 1925 году, [ 10 ] говорит, что циркуляция медленно поднимается до его устойчивого состояния из-за вязкости, когда наклонное крыло ускоряется от отдыха. Это явление объяснит подъемную ценность, которая меньше, чем предсказано. Как правило, дело было в том, чтобы найти источники для дополнительного подъема. Утверждалось, что этот эффект незначителен для потока с числом Рейнольдса , который типичен для полета насекомых. Число Рейнольдса является мерой турбулентности ; Поток является ламинарным (гладким), когда число Рейнольдса низкое и турбулентное, когда оно высокое. [ 11 ] Эффект Вагнера был проигнорирован сознательно, по крайней мере, в одной модели. [ 9 ] Одним из наиболее важных явлений, которое возникает во время полета насекомых, является всасывание переднего края. Эта сила важна для расчета эффективности. Концепция всасывания переднего края сначала была выдвинута DG Ellis и JL Stollery в 1988 году, чтобы описать вихревой лифт на Delta Therta с острыми краями . [ 12 ] Под высокими углами атаки поток отделяется по передней части края, но переживает, прежде чем достичь заднего края. В пределах этого пузыря разделенного потока находится вихрь. Поскольку угол атаки настолько высок, большая часть импульса переносится вниз в поток. Эти две функции создают большое количество подъемной силы, а также некоторое дополнительное сопротивление. Важной особенностью, однако, является лифт. Поскольку поток разделен, но он все еще обеспечивает большое количество подъема, это явление называется задержкой стойла , впервые замеченную на авиационных пропеллерах Х. Химмельскампом в 1945 году. [ 13 ] Этот эффект наблюдался при перелетевшем полете насекомых, и, как было доказано, способен обеспечить достаточный подъем, чтобы учесть недостаток в квазистаичественных моделях. Этот эффект используется каноистами в ходе удара . [ 8 ] [ 9 ]
Все эффекты на взмахивающее крыло могут быть уменьшены до трех основных источников аэродинамических явлений: вихревой вихревой края, устойчивые аэродинамические силы на крыле и контакт крыла с его следами предыдущих ударов. Размер летающих насекомых варьируется от 20 микрограммов до примерно 3 граммов. Когда масса тела насекомых увеличивается, площадь крыла увеличивается и частота удара крыла уменьшается. Для больших насекомых число Рейнольдса (Re) может достигать 10000, где поток начинает становиться турбулентным. Для меньших насекомых это может быть всего 10. Это означает, что вязкие эффекты гораздо важнее для меньших насекомых. [ 9 ] [ 14 ] [ 15 ]


Еще одна интересная особенность полета насекомых - это наклон тела. По мере увеличения скорости полета тело насекомых имеет тенденцию наклоняться в нос вниз и становится более горизонтальным. Это уменьшает фронтальную область и, следовательно, сопротивление тела. Поскольку сопротивление также увеличивается по мере увеличения скорости вперед, насекомое делает свой полет более эффективным, поскольку эта эффективность становится более необходимой. Кроме того, изменяя геометрический угол атаки на нисходящем ударе, насекомое может сохранить свой полет с оптимальной эффективностью через как можно больше маневров. Развитие общей тяги относительно невелико по сравнению с силами подъемника. Силы подъема могут быть более чем в три раза больше веса насекомых, в то время как тяга даже на самых высоких скоростях может составлять всего 20% от веса. Эта сила разрабатывается в первую очередь благодаря менее мощному вверх по удару движения. [ 9 ] [ 16 ]
Хлопать и бросить
[ редактировать ]

Clap and Fling, или механизм Weis-Fogh, обнаруженный датским зоологом Торкелем Вайс-Фог , представляет собой метод генерации лифта, используемый во время полета небольших насекомых. [ 17 ] По мере того, как размеры насекомых становятся менее 1 мм, вязкие силы становятся доминирующими, а эффективность обработки подъема из аэродинамического профиля резко уменьшается. Начиная с положения хлопа, два крыла разбиваются и вращаются вокруг заднего края. Затем крылья отделяются и подметают горизонтально до конца удара вниз. Затем крылья проводятся и используют передний край во время гребного движения наверх. Когда начинается движение по хлопоту, ведущие края встречаются и вращаются вместе, пока разрыв не исчезнет. Первоначально считалось, что крылья касались, но несколько инцидентов указывают на разрыв между крыльями и предполагают, что это обеспечивает аэродинамическое преимущество. [ 18 ] [ 19 ]
Генерация подъема из механизма хлопчала и броска происходит во время нескольких процессов на протяжении всего движения. Во-первых, механизм опирается на взаимодействие крыла, так как одно крыло не дает достаточного подъема. [ 20 ] [ 21 ] [ 22 ] Когда крылья вращаются вокруг заднего края в движении броска, воздух бросается в созданный зазор и генерирует сильный вихревой вихревой края, а второй разрабатывает на крылах. Третий, более слабый вихрь развивается на заднем крае. [ 19 ] Сила развивающихся вихрей зависит от начального разрыва между крыльями в начале движения броска. С уменьшенным промежутком меж крылья, указывающим на большую обработку лифта, за счет более крупных сил сопротивления. Реализация движения поднимающегося во время фриста, [ 23 ] гибкие крылья, [ 21 ] и было обнаружено, что задержка с задержкой киоска усиливает стабильность и прикрепление вихря. [ 24 ] Наконец, чтобы компенсировать общее более низкое производство подъема во время полета с низким числом Рейнольдса (с ламинарным потоком ), крошечные насекомые часто имеют более высокую частоту хода для генерации скоростей крыла, которые сравнимы с большими насекомыми. [ 24 ]
В целом крупнейшие ожидаемые силы сопротивления происходят во время движения дорсального фантазии, поскольку крылья должны отделяться и вращаться. [ 22 ] Затухание больших сил сопротивления происходит через несколько механизмов. Было обнаружено, что гибкие крылья уменьшают сопротивление в броске движения на 50% и еще больше уменьшают общее сопротивление через весь ход крыла по сравнению с жесткими крыльями. [ 21 ] Компания на краях крыла, как видно из Formosa Encarsia , вызывают пористость в потоке, которая увеличивает и уменьшает силы сопротивления, за счет более низкого образования лифта. [ 25 ] Кроме того, межступенчатое разделение перед Фриптом играет важную роль в общем эффекте сопротивления. По мере увеличения расстояния между крыльями общее сопротивление уменьшается. [ 18 ]
Механизм хлопок и броска также используется морской морской лимачиной Хелисиной , морской бабочкой. [ 26 ] [ 27 ] Некоторые насекомые, такие как майнер овощных листьев Liriomyza sativae (муха), используют частичный хлопок и бросок, используя механизм только на внешней части крыла, чтобы увеличить подъем примерно на 7% при паря. [ 24 ]
- Механизм полета хлопает и бросает в SANE 2003
-
 Хлоп 1: крылья закрываются назад
Хлоп 1: крылья закрываются назад -
 Хлоп 2: Связанные с ребрами, крыло вращается вокруг ведущего края , вихри
Хлоп 2: Связанные с ребрами, крыло вращается вокруг ведущего края , вихри -
 Хлоп 3: Закрытые края закрываются, вихри проливают, крылья закрывают, давая тягу
Хлоп 3: Закрытые края закрываются, вихри проливают, крылья закрывают, давая тягу



- Черные (изогнутые) стрелки: поток; Синие стрелки: индуцированная скорость; Оранжевые стрелки: чистая сила на крыле
-
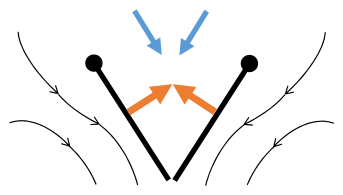 Переплет 1: Крылья вращаются вокруг затяжного края, чтобы разбрасываться
Переплет 1: Крылья вращаются вокруг затяжного края, чтобы разбрасываться -
 Переплетение 2: передний край уходит, воздух бросается, увеличивает лифт
Переплетение 2: передний край уходит, воздух бросается, увеличивает лифт -
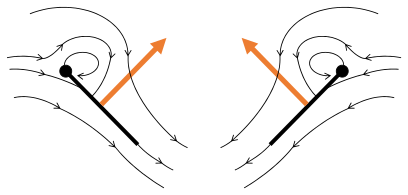 Трип 3: Новые вихревые формы на переднем крае, вихревые вихри с краями отменяют друг друга, возможно, помогая потоку расти быстрее ( Weis-Fogh 1973)
Трип 3: Новые вихревые формы на переднем крае, вихревые вихри с краями отменяют друг друга, возможно, помогая потоку расти быстрее ( Weis-Fogh 1973)



Управляющие уравнения
[ редактировать ]Крыло, движущееся в жидкости, испытывает жидкость , которая следует за конвенциями, обнаруженными в аэродинамике. Компонент силы, нормальный к направлению потока, относительно крыла, называется подъемным ( L ), а компонентом силы в противоположном направлении потока является перетаскивание ( D ). На номерах Рейнольдса, рассмотренных здесь, подходящая силовая единица составляет 1/2 (ρu 2 S), где ρ - плотность жидкости, S область крыла и u с скоростью крыла. Силы без измерений называются коэффициентами подъема ( C L ) и перетаскивания ( C D ), то есть: [ 8 ]

C L и C D являются постоянными, только если поток устойчив. Специальный класс объектов, таких как аэродинамический профиль, может достигать устойчивого состояния, когда он просачивается через жидкость под небольшим углом атаки. В этом случае невидимый поток вокруг аэродинамического профиля может быть аппроксимирован потенциальным потоком, удовлетворяющим граничное условие отсутствия. Теорема Кутта -Жуковского 2D-профиля еще более предполагает, что поток покидает острый задний край плавно, и это определяет полную циркуляцию вокруг аэродинамической промышленности. Соответствующий подъем дается принципом Бернулли ( теорема Блазиуса ): [ 8 ]

Потоки вокруг птиц и насекомых можно считать несжимаемыми : число Маха или скорость относительно скорости звука в воздухе обычно составляет 1/300, а частота крыла составляет около 10–103 Гц. Используя руководящее уравнение в качестве уравнения Навье-Стоукса, подвергающегося ограничению без скольжения, уравнение: [ 8 ]

Где u (x, t) - это поле потока, p давление, ρ плотность жидкости, ν кинематической вязкости, u достигает скорости на границе и u s скорость твердого вещества. Выбирая масштаб длины, L и шкала скорости, U, уравнение может быть выражено в неразмерной форме, содержащей число Рейнольдса, r e = ul/ν. Существует два очевидных различия между крылом насекомых и аэродинамическим профилем: крыло насекомых намного меньше, и оно просачивается. Используя дракоза в качестве примера, ее аккорд (C) составляет около 1 см (0,39 дюйма), длина крыла (L) около 4 см (1,6 дюйма) и его частота крыла (F) около 40 Гц. Скорость наконечника (U) составляет около 1 м/с (3,3 фута/с), а соответствующее число Рейнольдса около 103. На меньшем конце типичная хальцидоидная оса имеет длину крыла около 0,5–0,7 мм (0,020–0,028 в) и бьет его крыло около 400 Гц. Его номер Рейнольдса составляет около 25. Диапазон номера Рейнольдса в полете насекомых составляет от 10 до 10 4 , который находится между двумя пределами, которые удобны для теорий: непревзойденные устойчивые потоки вокруг аэродинамического профиля и потока Стокса, испытываемых плавательной бактерией. По этой причине этот промежуточный диапазон не совсем понятен. С другой стороны, это, пожалуй, самый вездесущий режим среди вещей, которые мы видим. Падающие листья и семена, рыбы и птицы сталкиваются с неустойчивыми потоками, похожими на то, что видели вокруг насекомого. [ 8 ] Номер Рейнольдса chordwise может быть описан:

и


Где Средняя длина аккорда, скорость кончика крыла, Амплитуда хода, это частота ритма, Радиус вращения, это область крыла, и Длина крыла, включая наконечник крыла.







В дополнение к номеру Рейнольдса, существует как минимум два других соответствующих параметров безразмерных. Крыло имеет три шкалы скорости: скорость размахивания относительно тела ( u ), передняя скорость тела ( U 0 ) и скорость качки (ω c ). Соотношения их образуют две безразмерные переменные, u 0 / u и ωc / u , первое часто называют коэффициентом предварительного, и также связано с уменьшенной частотой, Fc / u 0 . [ 8 ]
Если крыло насекомого твердо, например, крыло дрозофилы примерно так, его движение относительно фиксированного тела может быть описано тремя переменными: положение кончика в сферических координатах (θ (t), φ (t) ) и угол качки ψ (t), вокруг оси, соединяющей корень и наконечник. Чтобы оценить аэродинамические силы на основе анализа элементов лезвия, также необходимо определить угол атаки (α). Типичный угол атаки при 70% размах крыльев варьируется от 25 ° до 45 ° у парящих насекомых (15 ° у колибри). Несмотря на богатство данных, доступных для многих насекомых, относительно несколько экспериментов сообщают о изменении времени α во время инсульта. Среди них находятся эксперименты на аэродинамической трубе привязанной саранчи и привязанную муху, а также свободный зависающий полет фруктовой мухи. [ 8 ]
Поскольку их относительно легко измерить, траектории наклона крыла сообщались чаще. Например, выбор только последовательностей полета, которые создали достаточный подъем, чтобы поддержать вес, покажет, что наконечник крыла следует эллиптической форме. Неоткрытые формы также были зарегистрированы для других насекомых. Независимо от их точных форм, движение подключения указывает на то, что насекомые могут использовать аэродинамическое сопротивление в дополнение к подъему для поддержки его веса. [ 8 ]
Пари
[ редактировать ]| Параметры полета [ 28 ] | Скорость (м/с) | Beats/s |
|---|---|---|
| Aeshnid Dragonfly | 7.0 | 38 |
| Шершень | 5.7 | 100 |
| Колибри Хокмот | 5.0 | 85 |
| лошадь | 3.9 | 96 |
| Хуверфляй | 3.5 | 120 |
| Шмель | 2.9 | 130 |
| Пчела | 2.5 | 250 |
| Дом | 2.0 | 190 |
| Сами | 1.5 | 16 |
| Scorpionfly | 0.49 | 28 |
| Большая белая бабочка [ 29 ] | 2.5 | 12 |
| Трипсы (хлопают и бросают) [ 24 ] | 0.3 | 254 |

Многие насекомые могут парить или оставаться в одном месте в воздухе, делая это, быстро избивая свои крылья. Это требует стабилизации боковой стороны, а также производства подъема. Подъемная сила в основном производится вниз по ниду. Когда крылья толкают окружающий воздух, результирующая сила реакции воздуха на крыльях подталкивает насекомого. Крылья большинства насекомых эволюционированы так, что во время восходящего хода сила на крыле маленькая. Поскольку удары пониженного и возврата заставляют насекомое вверх и вниз соответственно, насекомое колеблется и заводится в том же положении. [ 14 ]
Расстояние, которое насекомое падает между битами, зависит от того, насколько быстро бьют его крылья: чем медленнее оно ломается, тем дольше интервал, в котором оно падает, и чем дальше оно падает между каждым крылом. Можно рассчитать частоту крылья, необходимую для того, чтобы насекомое поддержило заданную стабильность в ее амплитуде. Чтобы упростить расчеты, необходимо предположить, что подъемная сила находится на конечном постоянном значении, пока крылья движутся вниз и что оно равна нулю, пока крылья движутся вверх. В течение временного интервала Δ t вверх крыла, насекомое падает на расстояние H под воздействием гравитации. [ 14 ]

Вверх ход затем восстанавливает насекомое в исходное положение. Как правило, может потребоваться, чтобы вертикальное положение насекомых изменялось не более чем на 0,1 мм (то есть H = 0,1 мм). Максимально допустимое время для свободного падения тогда [ 14 ]

Поскольку движения UP и вниз по движениям крыльев примерно равны по продолжительности, период T для полного крыла вверх и вниз-два раза Δ R , то есть, то есть, есть, есть [ 14 ]

Частота ударов, f, что означает количество крыльев в секунду, представлена уравнением: [ 14 ]

В используемых примерах используемая частота составляет 110 ударов/с, что является типичной частотой, обнаруженной у насекомых. Бабочки имеют гораздо более медленную частоту с примерно 10 ударов/с, что означает, что они не могут парить. Другие насекомые могут получить частоту 1000 ударов/с. Чтобы восстановить насекомое в исходном вертикальном положении, средняя сила вверх во время нисходящего удара, F AV , должна быть равна вдвое большему весу насекомого. Обратите внимание, что, поскольку усилия вверх на теле насекомых применяются только в течение половины времени, средняя сила вверх на насекомых - это просто вес. [ 14 ]
Вход питания
[ редактировать ]Теперь можно вычислить мощность, необходимую для поддержания зависания, учитывая снова насекомое с массой M 0,1 г, средняя сила, F AV , применяемая двумя крыльями во время нисходящего удара, в два раза превышает вес. Поскольку давление, приложенное крыльями, равномерно распределено по площади общей крыла, это означает, что можно предположить, что сила, генерируемая каждым крылом, действует через одну точку в середине крыльев. Во время нисходящего хода центр крыльев проходит на вертикальном расстоянии d . [ 14 ] Общая работа, выполняемая насекомым во время каждого нисходящего удара, является продуктом силы и расстояния; то есть,

Если крылья качаются через удар под углом 70 °, то в случае, представленном для насекомых с крыльями длиной 1 см, D составляет 0,57 см. Следовательно, работа, выполняемая во время каждого удара двумя крыльями, составляет: [ 14 ]

Энергия используется для повышения насекомых против гравитации. Энергия E , необходимая для повышения массы насекомых 0,1 мм во время каждого удара, - это: [ 14 ]

Это незначительная доля от общей затраченной энергии, которая, очевидно, ясно, что большая часть энергии тратится в других процессах. Более подробный анализ проблемы показывает, что работа, выполняемая крыльями, преобразуется в основном в кинетическую энергию воздуха, которая ускоряется вниз по нисходящему ходу крыльев. Сила - это объем работы, выполненную за 1 с; У насекомого, используемого в качестве примера, делает 110 нисходящих ударов в секунду. Следовательно, его выходная мощность P - это удары в секунду, а это означает, что его выходная мощность P - это: [ 14 ]

Выходная мощность
[ редактировать ]При расчете мощности, используемой при зависании, в примерах использовались пренебрежение кинетической энергией движущихся крыльев. Крылья насекомых, как они есть, имеют конечную массу; Поэтому, когда они двигаются, они обладают кинетической энергией. Поскольку крылья находятся в роторном движении, максимальная кинетическая энергия во время каждого хода крыла: [ 14 ]

Здесь я - момент инерции крыла, а ω max - максимальная угловая скорость во время хода крыла. Чтобы получить момент инерции для крыла, мы предположим, что крыло может быть аппроксимировано тонким стержнем, поворачированным на одном конце. Момент инерции для крыла тогда: [ 14 ]

Где L - длина крыла (1 см), а М - масса двух крыльев, которые обычно могут быть 10 −3 глин Максимальная угловая скорость, ω max , может быть рассчитана по максимальной линейной скорости , ν max , в центре крыла: [ 14 ]

Во время каждого удара центр крыльев перемещается со средней линейной скоростью ν av, заданной на расстоянии D, проходящем центром крыла, разделенного на продолжительность ΔT хода крыла. Из нашего предыдущего примера D = 0,57 см и ΔT = 4,5 × 10 −3 с Поэтому: [ 14 ]

Скорость крыльев равна нулю как в начале, так и в конце хода крыла, что означает, что максимальная линейная скорость выше средней скорости. Если мы предположим, что скорость колеблется ( синусоидально ) вдоль пути крыла, максимальная скорость в два раза выше средней скорости. Следовательно, максимальная угловая скорость составляет: [ 14 ]

И кинетическая энергия, следовательно, такова: [ 14 ]

Поскольку в каждом цикле движения крыла имеется два удара крыла (наверху и вниз), кинетическая энергия составляет 2 × 43 = 86 ERG . Это примерно столько же энергии, сколько потребляется в самом зависе. [ 14 ]
Эластичность
[ редактировать ]
Насекомые получают кинетическую энергию, обеспечиваемую мышцами, когда крылья ускоряются . Когда крылья начинают замедляться к концу удара, эта энергия должна рассеиваться. Во время нагрузки кинетическая энергия рассеивается самими мышцами и превращается в тепло (это тепло иногда используется для поддержания температуры тела ядра). Некоторые насекомые способны использовать кинетическую энергию в восходящем движении крыльев, чтобы помочь в их полете. Крыловые суставы этих насекомых содержат прокладку из упругого резиноподобного белка, называемого Resilin . Во время подъема крыла релилин растягивается. Кинетическая энергия крыла преобразуется в потенциальную энергию в растянутой релилин, которая хранит энергию, как пружину. Когда крыло движется вниз, эта энергия выпускается и помогает вниз вниз. [ 14 ]
Используя несколько упрощающих допущений, мы можем рассчитать количество энергии, хранящейся в растянутом релилин. Хотя резилин сгибается в сложной форме, приведенный пример показывает расчет как прямой стержень области A и длины. Кроме того, мы будем предполагать, что на протяжении всего растяжения резилин подчиняется закону Гука . Это не строго верно, так как Resilin растягивается значительным количеством, и, следовательно, как область, так и модуль Янга в процессе растяжения. Потенциальная энергия, которую вы сохраняют в растянутой релиции: [ 14 ]

Здесь E - модуль Янга для Resilin, который был измерен как 1,8 × 10 7 / см 2 Полем Обычно у насекомого размером пчела объем резилина может быть эквивалентен цилиндру 2 × 10 −2 CM длиной и 4 × 10 −4 см 2 в области. В приведенном примере длина резилинового стержня увеличивается на 50% при растяжении. То есть Δℓ - 10 −2 см. Следовательно, в этом случае потенциальная энергия, хранящаяся в резилине каждого крыла: [ 14 ]

Хранитованная энергия в двух крылах для насекомого размером с пчела составляет 36 ERG, что сопоставимо с кинетической энергией вверх по удару крыльев. Эксперименты показывают, что до 80% кинетической энергии крыла можно хранить в Resilin. [ 14 ]
Крылоф
[ редактировать ]
Некоторые четырехступенчивые заказы на насекомые, такие как чешуекрылые, разработали морфологические механизмы связи крыла у Imago , которые делают эти таксоны функционально двумя крыльями. [ 30 ] Все, кроме самых базальных форм, демонстрируют эту связанную сфере крыла. [ 31 ]
Механизмы из трех разных типов-Югал, Френуло-Ретинакулярный и Амплексельный: [ 32 ]
- Более примитивные группы имеют увеличенную долю, похожую на долю, возле базального заднего края, т.е. у основания переднего крыла, югум , который складывается под задним ходом в полете. [ 31 ] [ 33 ]
- Другие группы имеют френулум на задних крылах, который зацепится под сетчаткой на переднем крыте. [ 33 ]
- Почти во всех бабочках и в бомбикоиде (кроме Sphingidae ) не существует расположения френулы и сетчатки, чтобы соединить крылья. Вместо этого увеличенная плечевая область задних крыльев широко перекрывается передним крылом. Несмотря на отсутствие конкретного механического соединения, крылья перекрываются и работают в фазе. Силовой ход переднего крыла толкает задних крыльев в унисон. Этот тип муфты является вариацией типа FRENATE, но где Frenulum и Retinaculum полностью потеряны. [ 30 ] [ 34 ]
Биохимия
[ редактировать ]Биохимия полета насекомых была в центре внимания. В то время как многие насекомые используют углеводы и липиды в качестве источника энергии для полета, многие жуки и мухи используют аминокислотный пролин в качестве источника энергии. [ 35 ] Некоторые виды также используют комбинацию источников и мотыльков, таких как Manduca Sexta, используют углеводы для разминки перед полетом. [ 36 ]
Сенсорная обратная связь
[ редактировать ]Насекомые используют сенсорную обратную связь для поддержания и управления полетом. Исследования продемонстрировали роль сенсорных структур, таких как антенны , [ 37 ] гантель [ 38 ] и крылья [ 39 ] В управлении полевой осанкой, амплитудой WingBeat и частотой WingBeat.
Эволюция и адаптация
[ редактировать ]
Когда -то в каменноугольный период , от 350 до 300 миллионов лет назад, когда было только две основные земельные массы, насекомые начали летать. [ 40 ] Среди самых старых ископаемых насекомых - делицшала , палеодиктиоптеран из нижнего карбона; [ 41 ] Rhyniognatha старше, от раннего девона , но неясно, было ли у него крылья или действительно было насекомым. [ 42 ] [ 43 ]
Как и почему развивались крылья насекомых, недостаточно понятно, в значительной степени из -за нехватки соответствующих окаменелостей из периода их развития в нижнем углероде. Исторически было три основные теории о происхождении полета насекомых. Первым было то, что они представляют собой модификации подвижных брюшных жабра обнаружено на водных наядах Мэйфлэ , как это . Филогеномный анализ предполагает, что полиоптера , группа крылатых насекомых, которые включают кузнечиков, развивались от наземного предка, что делает эволюцию крыльев из жабр маловероятным. [ 44 ] Дополнительное изучение прыжкового поведения личинок Мэйфля определило, что жабры трахеи не играют никакой роли в направлении происхождения насекомых, предоставляя дополнительные доказательства против этой эволюционной гипотезы. [ 45 ] Это оставляет две основные исторические теории: это крылья развивались от паранотических дол, расширения грудной территории ; или что они возникли из модификаций сегментов ног, которые уже содержали мышцы. [ 46 ]
Эпиксаль (брюшная жабер) гипотеза
[ редактировать ]
Многочисленные [ 47 ] Энтомологи , включая Ландоа в 1871 году, Лаббок в 1873 году, Грабер в 1877 году и Осборн в 1905 году, предположили, что возможным происхождением для насекомых крыльев могло быть подвижные брюшные жабра, найденные у многих водных насекомых, например, на Наядах Мэйфлс. Согласно этой теории, эти трахеальные жабры, которые начались в качестве выхода дыхательной системы и со временем были изменены в локомотивные цели, в конечном итоге превратились в крылья. Трахеальные жабры оснащены маленькими крыльями, которые постоянно вибрируют и имеют свои крошечные прямые мышцы. [ 48 ]
Паранотальная (тергальная) гипотеза
[ редактировать ]Паранотическая доля или гипотеза доля (дорсальная стенка тела), предложенная Фриц Мюллером в 1875 году [ 49 ] и переработано Г. Крэмптоном в 1916 году, [ 47 ] Jarmila Kukalova-Peck в 1978 году [ 50 ] и Александр П. Раснитсин в 1981 году, среди прочих, [ 51 ] предполагает, что крылья насекомых, развивающиеся от паранотических дол, предварительная предварительная, обнаруженная у окаменелостей насекомых , которая помогла бы стабилизации при прыжке или падении. В пользу этой гипотезы является тенденция большинства насекомых, когда они поражаются во время подъема на ветви, сбежать, упав на землю. Такие доли служили парашютами и позволяли насекомому приземлиться более мягко. Теория предполагает, что эти доли постепенно росли больше, а на более поздней стадии разработал сустав с грудной клеткой. Даже позже появится мышцы, чтобы переместить эти сырые крылья. Эта модель подразумевает прогрессивное повышение эффективности крыльев, начиная с парашютирования , затем скользящего и, наконец, активного полета . Тем не менее, отсутствие существенных ископаемых доказательств развития крыловых суставов и мышц создает серьезную трудность теории, как и, казалось бы, спонтанное развитие артикуляции и венеции, и это в значительной степени отвергнуто экспертами в этой области. [ 48 ]
Энти-экстра (плевральная) гипотеза
[ редактировать ]
В 1990 году JWH Trueman предположил, что крыло было адаптировано из эндийтов и выходов , придатками к соответствующим внутренним и внешним аспектам примитивной конечности членистоногих , также называемых гипотезой плеврала. Это было основано на исследовании Goldschmidt в 1945 году на Drosophila melanogaster , в котором вариация, называемая «стручком» (для подомеров, сегментов конечностей), демонстрировала мутация, которая трансформировала нормальные крылья. Результат был истолкован как тройное расположение ног с некоторыми дополнительными придатками, но не хватало лапки, где обычно будет оборотная поверхность крыла. Эта мутация была переосмыслена как убедительные доказательства дорсального выхода и экинтного слияния, а не ноги, причем придатки гораздо лучше подходили к этой гипотезе. Иннервация, артикуляция и мускулатура, необходимые для эволюции крыльев, уже присутствуют в сегментах конечностей. [ 46 ]
Другие гипотезы
[ редактировать ]Другие гипотезы включают предположение Винсента Вигглсворта 1973 года о том, что крылья развивались из грудных выступов, используемых в качестве радиаторов . [ 48 ]
Адриан Томас и Оке Норберг предположили в 2003 году, что крылья могли изначально развиваться для плавания на поверхности воды, как видно в некоторых каменных светах . [ 52 ] [ 53 ]
Стивен П. Яновак и его коллеги предложили в 2009 году, что крыло происходит от направленного воздушного скользящего спуска - феномена предварительного полета, обнаруженного в некоторых Аптериготе , бессмысленном сестринском таксоне до крылатых насекомых. [ 54 ]
Двойное происхождение
[ редактировать ]Биологи, включая Авероф, [ 55 ] Нива, [ 56 ] Элиас-Нето [ 57 ] и их коллеги начали исследовать происхождение крыла насекомых с использованием Evo-Devo в дополнение к палеонтологическим данным. Это говорит о том, что крылья последовательно гомологичны как с тергальными, так и с плевральными структурами, потенциально разрешающие многовековые дебаты. [ 58 ] Jakub Prokop и его коллеги в 2017 году обнаружили палеонтологические данные из палеозойских подушек для нимфы, что крылья действительно имели такое двойное происхождение. [ 59 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Чепмен, AD (2006). Количество живых видов в Австралии и мире . Канберра: Австралийское исследование биологических ресурсов . С. 60pp. ISBN 978-0-642-56850-2 Полем Архивировано с оригинала на 2009-05-19 . Получено 2015-09-15 .
- ^ Смит, Д.С. (1965). «Летные мышцы насекомых». Scientific American . 212 (6): 76–88. Bibcode : 1965sciam.212f..76s . doi : 10.1038/Scientificamerican0665-76 . PMID 14327957 .
- ^ Джозефсон, Роберт К.; Маламуд, Джин Г.; Стоукс, Даррелл Р. (2001). «Эффективность асинхронной летной мышцы из жука». Журнал экспериментальной биологии . 204 (23): 4125–4139. doi : 10.1242/jeb.204.23.4125 . ISSN 0022-0949 . PMID 11809787 .
- ^ «Определение асинхронной мышцы в глоссарии энтомологов» . Кафедра энтомологии, Университет штата Северная Каролина . Получено 2011-03-21 .
- ^ Деора, Танви; Гандия, Намрата; SANE, SANJAY P. (2017-04-15). «Механика грудной клетки в мух» . Журнал экспериментальной биологии . 220 (8): 1382–1395. doi : 10.1242/jeb.128363 . ISSN 1477-9145 . PMID 28424311 . S2CID 207172023 .
- ^ Хайде, Г.Г. (1983). «Нейронные механизмы управления полетом в Diptera». Биона отчет . 2 : 33–52.
- ^ Мелис, Йохан М.; Дикинсон, Майкл Х. (2023-06-30). «Машинное обучение раскрывает механику управления шарниром крыла насекомых» . Biorxiv: сервер препринта для биологии . doi : 10.1101/2023.06.29.547116 . PMC 10327165 . PMID 37425804 . Получено 2023-08-23 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Ван, З. Джейн (2005). «Расширение полета насекомых» (PDF) . Ежегодный обзор механики жидкости . 37 (1): 183–210. BIBCODE : 2005ANRFM..37..183W . doi : 10.1146/annurev.fluid.36.050802.121940 .
- ^ Jump up to: а беременный в дюймовый и фон глин Сэйн, Санджай П. (2003). «Аэродинамика полета насекомых» (PDF) . Журнал экспериментальной биологии . 206 (23): 4191–4208. doi : 10.1242/jeb.00663 . PMID 14581590 . S2CID 17453426 .
- ^ Вагнер, Герберт (1925). «О создании динамической плавучести крыла» [о происхождении динамического подъема аэродинамических профилей]. Журнал прикладной математики и механики (на немецком языке). 5 (1): 17–35. Код BIB : 1925ZAMM .... 5 ... 17W . Doi : 10.1002/Zamm .
- ^ «Переход и турбулентность» . Принстонский университет . Получено 8 июля 2021 года .
- ^ Эллис, DG; Stollery, JL (1988). «Поведение и производительность передовых вихревых клапанов» (PDF) . ICAS 1988 Иерусалим . 1988 . Международный совет авиационных наук: 758–765 . Получено 8 июля 2021 года .
- ^ Himmelskamp, H. (1945) «Расследования профиля на вращающемся воздушном скваре». Докторская диссертация, Университет Геттингена .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v Давидовит, Пол (2008). Физика в биологии и медицине . Академическая пресса . С. 78–79. ISBN 978-0-12-369411-9 .
- ^ Надежда, Даниэль К; Делука, Энтони М.; О'Хара, Райан П. (2018-01-03). «Исследование числа Рейнольдса на биомиметическое крыло хлопья» . Международный журнал микро -воздушных транспортных средств . 10 (1): 106–122. doi : 10.1177/1756829317745319 . ISSN 1756-8293 .
- ^ "Поймать след" . Scientific American . 28 июня 1999 г. Получено 31 марта 2011 года .
- ^ Weis-Fogh, Torkel (1973). «Быстрые оценки полезной физической подготовки у зависающих животных, включая новые механизмы производства подъема». Журнал экспериментальной биологии . 59 : 169–230. doi : 10.1242/jeb.59.1.169 .
- ^ Jump up to: а беременный Santhanakrishnan, A.; Робинсон, А.; Джонс, с.; Низкий, а.; Гади, с.; Хендрик, Т.; Миллер Л. (2014). «Механизм хлопок и броска с взаимодействующим пористым крылом в крошечном полете насекомых» . Журнал экспериментальной биологии . 217 (Pt 21): 3898–4709. doi : 10.1242/jeb.084897 . PMID 25189374 .
- ^ Jump up to: а беременный Kolomenskiy, D; Moffatt, H.; Farge, M.; Schneider, K. (2011). «Двух- и трехмерные численные моделирование хлопкового флеза парящих насекомых» . Журнал жидкостей и структур . 27 (5): 784. Bibcode : 2011JFS .... 27..784K . doi : 10.1016/j.jfluidstructs.2011.05.002 .
- ^ Беннетт Л. (1977). «Clap and Chling Aerodynamics- экспериментальная оценка». Журнал экспериментальной биологии . 69 : 261–272. doi : 10.1242/jeb.69.1.261 .
- ^ Jump up to: а беременный в Miller, L.; Пескин С. (2009). «Гибкий хлопок и брось в крошечный полет насекомых» . Журнал экспериментальной биологии . 212 (19): 3076–3090. doi : 10.1242/jeb.028662 . PMID 19749100 . S2CID 29711043 .
- ^ Jump up to: а беременный Lehmann, F.-O.; SANE, S.; Дикинсон М. (2005). «Аэродинамические эффекты взаимодействия крыла на крылах насекомых» . Журнал экспериментальной биологии . 208 (Pt 16): 3075–3092. doi : 10.1242/jeb.01744 . PMID 16081606 . S2CID 7750411 .
- ^ Lehmann, F.-O.; Pick, S. (2007). «Аэродинамическое преимущество взаимодействия крыла зависит от траектории инсульта при взмахивании крыльев насекомых» . Журнал экспериментальной биологии . 210 (Pt 8): 1362–1377. doi : 10.1242/jeb.02746 . PMID 17401119 . S2CID 23330782 .
- ^ Jump up to: а беременный в дюймовый Ченг, Синь; Sun, Mao (2016-05-11). «Измерение и аэродинамику крыла и аэродинамики у небольшого насекомых при зависании полета» . Научные отчеты . 6 (1): 25706. Bibcode : 2016natsr ... 625706c . doi : 10.1038/srep25706 . PMC 4863373 . PMID 27168523 .
- ^ Kasoju, v.; Сантанакришнан, А. (2021). «Аэродинамическое взаимодействие партированных пар крыльев в флинге». Физика жидкости . 33 (3): 031901. Arxiv : 2011.00939 . Bibcode : 2021phfl ... 33C1901K . doi : 10.1063/5.0036018 . S2CID 226227261 .
- ^ Вайсбергер, Минди (19 февраля 2016 г.). «Плавать, как бабочка? Морская улитка» проходит через воду » . Scientific American . Получено 20 февраля 2016 года .
- ^ Мерфи, Д.; Adhikari, D.; Webster, D.; Йен, Дж. (2016). «Подводной полет у планктонной морской бабочки» . Журнал экспериментальной биологии . 219 (4): 535–543. doi : 10.1242/jeb.129205 . PMID 26889002 .
- ^ «Обсекие крылья в целом» . Аэродинамика насекомых . Cislunar Aerospace. 1997 . Получено 28 марта 2011 года .
- ^ «Бабочки в семье Pieridae (белые)» . Bumblebee.org . Получено 18 марта 2018 года .
- ^ Jump up to: а беременный Дадли, Роберт (2000). Биомеханика полета насекомых: форма, функция, эволюция . Принстон, Нью -Джерси: издательство Принстонского университета . п. 69. JSTOR J.CTV301G2X .
- ^ Jump up to: а беременный Акции, Ян. (2008). Раздел Крыловая связь в Capinera (Ed) (2008) «Encyc. Entom», Vol 4. P. 4266
- ^ Скобл, Малкольм Дж. (1995). Lepidoptera: форма, функция и разнообразие . Музей естественной истории . С. 56–60. ISBN 978-0-19-854952-9 .
- ^ Jump up to: а беременный Пауэлл, Джерри А. Ч. Lepidoptera (стр. 631-664) в Resh, & Cardé (Eds). «Энциклопедия насекомых». 2003.
- ^ Gorb, S. (2001) CH 4.1.5 "Interlocking частей тела". С. 46–50.
- ^ Woiwod, ip; Рейнольдс, доктор; Томас, CD (Eds) 2001. Движение насекомых: механизмы и последствия. Cab International.
- ^ Joos, B. (1987). «Использование углеводов в летных мышцах Manduca Sexta во время прогрева перед полетом». Журнал экспериментальной биологии . 133 : 317–327. doi : 10.1242/jeb.133.1.317 .
- ^ Сейн, Санджай П., Александр Дидидоне, Марк А. Уиллис и Томас Л. Даниэль. «Антеннальные механососенсоры опосредуют управление полетом в мотынах». Science 315, нет. 5813 (2007): 863-866.
- ^ Дикерсон, Брэдли Х., Алиша М. де Соуза, Эйнул Худа и Майкл Х. Дикинсон. «Мухи регулируют движение крыла посредством активного контроля двойного гироскопа». Текущая биология 29, нет. 20 (2019): 3517-3524.
- ^ Волк, Харальд. «Саранча Тегула: значение для генерации ритма полета, управления движением крыла и аэродинамической силы». Журнал экспериментальной биологии 182, нет. 1 (1993): 229-253.
- ^ Misof, B.; Лю, с.; Meusemann, K.; Петерс, рупий; Донат, А.; Mayer, C.; Франдсен, П.; и др. (2014). «Филогеномика разрешает время и паттерн эволюции насекомых». Наука . 346 (6210): 763–767. Bibcode : 2014sci ... 346..763m . doi : 10.1126/science.1257570 . ISSN 0036-8075 . PMID 25378627 . S2CID 36008925 .
- ^ Браукманн, Карстен; Schneider, Joerg (1996). «Постоянное насекомые из района Bitterfeld/Delitzsch (Pterygota, Arnsbergium, Германия)» [Нижнее каменноугольное насекомое из области Bitterfeld/Delitzsch (Pterygota, Arnsbergian, Germany)]. Новый ежегодный книга по геологии и палеонтологии - ежемесячные слои (на немецком языке). 1996 (1): 17–30. Doi : 10.1127/njgpm/1996/17 .
- ^ Tillyard, RJ (2009). «Некоторые замечания о девонских ископаемых насекомых с кроватей Rhynie Chert, старого красного песчаника». Сделки Королевского энтомологического общества Лондона . 76 (1): 65–71. doi : 10.1111/j.1365-2311.1928.tb01188.x . ISSN 0035-8894 .
- ^ Carolin Haug & Joachim Haug (2017). "Предполагаемое старое летающее насекомое: скорее всего, бесчисленное множество?" Полем ПЕРЕЙ . 5 : E3402. doi : 10.7717/peerj.3402 . PMC 5452959 . PMID 28584727 .
- ^ Wipfler, Benjamin; и др. (19 февраля 2019 г.). «Эволюционная история полиоптера и ее последствия для нашего понимания ранних крылатых насекомых» . Труды Национальной академии наук . 116 (8): 3024–3029. Bibcode : 2019pnas..116.3024W . doi : 10.1073/pnas.1817794116 . ISSN 0027-8424 . PMC 6386694 . PMID 30642969 .
- ^ Яновак, Стивен П.; Дадли, Роберт (июль 2018 г.). «Прыжок и воздушное поведение личинок водной майфляй (Myobaetis Ellenae, Baetidae)» . Членистоногие структура и развитие . 47 (4): 370–374. Bibcode : 2018artsd..47..370y . doi : 10.1016/j.asd.2017.06.005 . PMID 28684306 . S2CID 205697025 .
- ^ Jump up to: а беременный Trueman, JWH (1990), Комментарий: эволюция крыльев насекомых: модель Expect Execite Plus. Канадский журнал зоологии .
- ^ Jump up to: а беременный Крэмптон, Г. (1916). «Филогенетическое происхождение и природа крыльев насекомых в соответствии с паранотальной теорией». Журнал нью -йоркского энтомологического общества . 24 (1): 1–39. JSTOR 25003692 .
- ^ Jump up to: а беременный в Гримальди, Дэвид; Энгель, Майкл С. (2005). Насекомые выходят на небо . Нью -Йорк: издательство Кембриджского университета . С. 155–159.
{{cite book}}:|work=игнорируется ( помощь ) - ^ Мюллер, Фриц (1875) Йена. Газета Natural Sciences , 9, 241.
- ^ Kukalova-Peck, Jarmila (1978). «Происхождение и эволюция крыльев насекомых и их связь с метаморфозой, как задокументировано ископаемой записи». Журнал морфологии . 156 (1). Wiley: 53–125. doi : 10.1002/jmor.1051560104 . ISSN 0362-2525 . PMID 30231597 . S2CID 52301138 .
- ^ Раснитсин, Александр П. (1981). «Модифицированная паранотическая теория происхождения крыла насекомых». Журнал морфологии . 168 (3): 331–338. doi : 10.1002/jmor.1051680309 . ISSN 0362-2525 . PMID 30110990 . S2CID 52010764 .
- ^ Томас, Адриан Л.Р .; Норберг, Р. Оке (1996). «Смириться с поверхностью - происхождение полета у насекомых?». Тенденции в экологии и эволюции . 11 (5): 187–188. Bibcode : 1996tecoe..11..187t . doi : 10.1016/0169-5347 (96) 30022-0 . PMID 21237803 .
- ^ Марден, Джеймс (2003). «Гипотеза на поверхности по поверхности для эволюции полета насекомых». Acta Zoologica Cracoviensia . 46 : 73–84.
- ^ Яновак, Стивен П.; Kaspari, M.; Дадли Р. (2009). «Скоростные гексапод и происхождение воздушного поведения насекомых» . Биологические письма . 5 (4): 510–2. doi : 10.1098/rsbl.2009.0029 . PMC 2781901 . PMID 19324632 .
- ^ Авероф, Михалис; Коэн, Стивен М. (1997). «Эволюционное происхождение крыльев насекомых из наследственных жабра». Природа . 385 (6617). Springer Science and Business Media LLC: 627–630. Bibcode : 1997natur.385..627a . doi : 10.1038/385627A0 . ISSN 0028-0836 . PMID 9024659 . S2CID 4257270 .
- ^ Нива, Нао; Акимото-Като, ИИ; Ниими, Теруюки; Тоджо, Коджи; Мачида, Рючиро; Хаяси, Шигео (2010-03-17). «Эволюционное происхождение крыла насекомых посредством интеграции двух модулей развития». Эволюция и развитие . 12 (2): 168–176. doi : 10.1111/j.1525-142x.2010.00402.x . ISSN 1520-541X . PMID 20433457 . S2CID 15838166 .
- ^ Элиас-Нето, Мойзс; Belles, Xavier (2016). «Тергальные и плевральные структуры способствуют формированию эктопических проторакальных крыльев в тараканах» . Королевское общество открыто наука . 3 (8). Королевское общество: 160347. Bibcode : 2016rsos .... 360347e . doi : 10.1098/rsos.160347 . ISSN 2054-5703 . PMC 5108966 . PMID 27853616 .
- ^ Томоясу, Йошинори; Оде, Такахиро; Кларк-Хахтель, Кортни (2017-03-14). «Что серийные гомологи могут рассказать нам о происхождении крыльев насекомых» . F1000Research . 6 : 268. doi : 10.12688/f1000research.10285.1 . ISSN 2046-1402 . PMC 5357031 . PMID 28357056 .
- ^ Прокоп, Джакуб; Печарова, Мартина; Нел, Андре; Hörnschemeyer, Thomas; Креминьска, Эва; Креминьски, Вислав; Энгель, Майкл С. (2017). «Палеозойские нимфальные подушки поддерживают двойную модель происхождения крыла насекомых» . Текущая биология . 27 (2): 263–269. Bibcode : 2017cbio ... 27..263p . doi : 10.1016/j.cub.2016.11.021 . ISSN 0960-9822 . PMID 28089512 .
Дальнейшее чтение
[ редактировать ]- Дикинсон, MH; Lehmann, fo; SANE, SP (1999). «Вращение крыла и аэродинамическая основа полета насекомых». Наука . 284 (5422): 1954–60. doi : 10.1126/science.284.5422.1954 . PMID 10373107 .
- Эллингтон, Чарльз П.; Ван Ден Берг, Коэн; Уиллмотт, Александр П.; Томас, Адриан Л.Р. (1996). «Передовые вихри в полете насекомых». Природа . 384 (6610): 626–630. Bibcode : 1996natur.384..626e . doi : 10.1038/384626A0 . S2CID 4358428 .
- Эллингтон, CP (1984). «Аэродинамика парящего полета насекомых. I. квази-устроенный анализ». Философские транзакции Королевского общества B: биологические науки . 305 (1122): 1–15. Bibcode : 1984rsptb.305 .... 1e . doi : 10.1098/rstb.1984.0049 .
- Эллингтон, CP (1999). «Новая аэродинамика полета насекомых: применение на транспортные средства для микроализат» . Журнал экспериментальной биологии . 202 (Pt 23): 3439–3448. doi : 10.1242/jeb.202.23.3439 . PMID 10562527 .
- Левин, GC; Хадж-Харири, Х. (2003). «Моделирование генерации тяги двумерной аэродинамической профиля с воздействием вязкого потока». Журнал жидкой механики . 492 : 339–362. Bibcode : 2003jfm ... 492..339L . doi : 10.1017/s0022112003005743 . S2CID 122077834 .
- Прингл, JWS (1975). Полет насекомого . Оксфордские читатели биологии. Тол. 52
- SANE, SP (2003). «Аэродинамика полета насекомых». Журнал экспериментальной биологии . 206 (Pt 23): 4191–208. doi : 10.1242/jeb.00663 . PMID 14581590 . S2CID 17453426 .
- Savage, SB; Ньюман, BG; Вонг, DTM (1979). «Роль вихрей и неустойчивых последствий во время парящего полета мух драконов» . Журнал экспериментальной биологии . 83 (1): 59–77. doi : 10.1242/jeb.83.1.59 .
- Смит, Т. младший (1985). «Мышечные системы». В Blum, MS Blum (ред.). Основы физиологии насекомых . Джон Уайли и сыновья. С. 227–286.
- Сотавалта, О. (1953). «Записи высокой частоты вибрации с высоким ходом и грудной вибрацией» . Биол. Бык Вудс -дыра . 104 (3): 439–444. doi : 10.2307/1538496 . JSTOR 1538496 .
- Ван ден Берг, C.; Эллингтон, CP (1997). «Пробуждение вихря« парящей »модели Хоукмота» . Философские транзакции Королевского общества B: биологические науки . 352 (1351): 317–328. BIBCODE : 1997RSPTB.352..317V . doi : 10.1098/rstb.1997.0023 . PMC 1691928 .
- Уокер, JA (сентябрь 2002 г.). "Ротационный лифт: что -то отличие или более того же?" Полем Журнал экспериментальной биологии . 205 (Pt 24): 3783–3792. doi : 10.1242/jeb.205.24.3783 . PMID 12432002 .
- Zbikowski, R. (2002). «О аэродинамическом моделировании крыла, похожая на насекомых, в паряке для микрофиналов». Философские транзакции Королевского общества A: математические, физические и инженерные науки . 360 (1791): 273–290. BIBCODE : 2002RSPTA.360..273Z . doi : 10.1098/rsta.2001.0930 . PMID 16210181 . S2CID 2430367 .
Внешние ссылки
[ редактировать ]- Роль насекомого в разработке микро -воздушных транспортных средств
- Микро воздушные автомобили, похожие на насекомые
- Новая аэродинамика полета насекомых: применение на транспортные средства для микроализа
- Аэродинамика полета насекомых
- Летные мышцы архивировали 2004-08-22 на машине Wayback
- Визуализация потока аэродинамических механизмов бабочки
- Полет шмеля
- Полет насекомого