Развитие конечностей
| Развитие конечностей | |
|---|---|
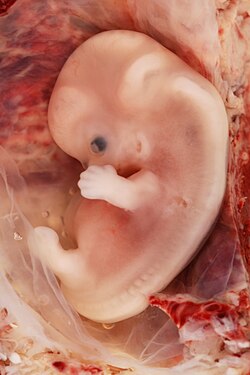
 Иллюстрация человеческого эмбриона на сроке беременности шесть недель. | |
 9-недельный плод человека от внематочной беременности | |
| Анатомическая терминология |
Развитие конечностей у позвоночных является областью активных исследований как в биологии развития , так и в эволюционной биологии , причем большая часть последних работ сосредоточена на переходе от плавника к конечностям . [1]
Формирование конечностей начинается в морфогенетическом поле конечностей , когда мезенхимальные клетки из мезодермы латеральной пластинки пролиферируют до такой степени, что они вызывают эктодермы выпячивание выше, образуя зачаток конечностей . Фактор роста фибробластов (FGF) индуцирует образование организатора на конце зачатка конечности, называемого апикальным эктодермальным гребнем (AER), который направляет дальнейшее развитие и контролирует гибель клеток . Запрограммированная смерть клеток необходима для устранения перепонки между пальцами .
Поле конечностей — это область, определяемая экспрессией определенных Hox-генов , подмножества гомеозисных генов и T-box факторов транскрипции — Tbx5 для развития передних конечностей или крыльев и Tbx4 для развития ног или задних конечностей. Установление поля передних конечностей (но не поля задних конечностей) требует передачи сигналов ретиноевой кислоты в развивающемся туловище эмбриона, из которого появляются зачатки конечностей. [2] [3] Кроме того, хотя избыток ретиноевой кислоты может изменить формирование паттерна конечностей путем эктопической активации экспрессии Shh или Meis1/Meis2, генетические исследования на мышах, которые устраняют синтез ретиноевой кислоты, показали, что RA не требуется для формирования паттерна конечностей. [4]
остается Зачаток конечностей активным на протяжении большей части развития конечностей, поскольку он стимулирует создание и сохранение положительной обратной связи двух сигнальных областей: AER и его последующее создание зоны поляризующей активности (ZPA) с мезенхимальными клетками . [5] В дополнение к дорсально-вентральной оси, создаваемой эктодермальной экспрессией конкурентных Wnt7a и сигналов BMP соответственно, эти сигнальные центры AER и ZPA имеют решающее значение для правильного формирования конечности, которая правильно ориентирована с соответствующей осевой полярностью в развивающемся организме. [6] [7] Поскольку эти сигнальные системы взаимно поддерживают активность друг друга, развитие конечностей по существу автономно после того, как эти сигнальные области были установлены. [5]
Формирование конечностей
[ редактировать ]Зачаток конечности
[ редактировать ]Формирование конечностей начинается в морфогенетическом поле конечностей . Формирование конечностей происходит в результате серии реципрокных тканевых взаимодействий между мезенхимой латеральной пластинки мезодермы и вышележащими эпителиальными клетками эктодермального происхождения. Клетки из латеральной пластинки мезодермы и миотома мигрируют в область конечности и пролиферируют до такой степени, что вызывают эктодермы выпячивание расположенной выше , образуя зачаток конечности . Клетки латеральной пластинки производят хрящевые и скелетные части конечности, а клетки миотома производят мышечные компоненты.
Мезодермальные клетки латеральной пластинки секретируют факторы роста фибробластов ( FGF7 и FGF10 ), чтобы побудить вышележащую эктодерму сформировать организатор на конце зачатка конечности, называемый апикальным эктодермальным гребнем (AER), который направляет дальнейшее развитие и контролирует гибель клеток . [8] AER секретирует дополнительные факторы роста FGF8 и FGF4 , которые поддерживают сигнал FGF10 и индуцируют пролиферацию в мезодерме. [ нужна ссылка ] Положение экспрессии FGF10 регулируется двумя сигнальными путями Wnt : Wnt8c в задних конечностях и Wnt2b в передних конечностях. Передняя и задняя конечности определяются их положением вдоль передней/задней оси и, возможно, двумя факторами транскрипции : Tbx5 и Tbx4 соответственно. [9] [10]
Предхрящевые уплотнения
[ редактировать ]Скелетные элементы конечности представляют собой плотные агрегаты, известные как клеточные конденсации предхрящевых мезенхимальных клеток. [11] Мезенхимальная конденсация опосредуется внеклеточным матриксом и молекулами клеточной адгезии . [12] В процессе хондрогенеза хондроциты хрящ дифференцируются из конденсаций и образуют , дающий начало скелетным зачаткам . При развитии большинства конечностей позвоночных (но не у некоторых земноводных) на более поздних стадиях развития хрящевой скелет заменяется костью .
Периодичности рисунка конечностей
[ редактировать ]
Конечность разделена на три отдела: шилопод , зевгопод и аутопод (в порядке от проксимального к дистальному ). Зеугопод и автопод содержат ряд периодических и квазипериодических мотивов узора . Зеугопод состоит из двух параллельных элементов вдоль переднезадней оси, а аутопод содержит от трех до пяти (в большинстве случаев) элементов вдоль одной оси. Пальцы также имеют квазипериодическое расположение вдоль проксимодистальной оси, состоящее из тандемных цепочек скелетных элементов. Генерация базового плана конечностей во время развития является результатом формирования паттерна мезенхимы в результате взаимодействия факторов, которые способствуют конденсации предхряща, и факторов, которые ее ингибируют. [13]
Разработка основного плана конечности сопровождается возникновением локальных различий между элементами. Например, лучевая и локтевая кости передней конечности, а также большеберцовая и малоберцовая кости задней конечности зевгопода отличаются друг от друга, как и разные пальцы рук и ног автопода. Эти различия можно рассматривать схематически, рассматривая, как они отражаются в каждой из трех основных осей конечностей.
По общему мнению, формирование паттерна скелета конечностей включает один или несколько типа Тьюринга механизмов реакции-диффузии . [1]
Эволюция и развитие
[ редактировать ]Эволюция конечностей из парных плавников была областью большого внимания. [1] Было проведено множество исследований, направленных на выяснение конкретных генов и факторов транскрипции, ответственных за развитие конечностей (см. Таблицу 1 ниже). В частности, исследования показали, что семейства генов SHH , DACH1 и BMP , HOX , T-box , FGF и WNT играют решающую роль в успешном развитии и формировании конечностей. [14] [15] для изучения генов, участвующих в развитии конечностей (и, следовательно, в эволюции), является уменьшение и потеря конечностей у змей . Дополнительным подходом [16] Консервативные последовательности, участвующие в развитии конечностей, сохраняются в геномах змей. Определенные последовательности энхансеров конечностей также консервативны между различными типами придатков, такими как конечности и фаллос . [16] [17] Напр., передача сигналов развития конечностей играет роль как в развитии конечностей, так и генитального бугорка у мышей. [16] [17] Изучение уменьшения и потери конечностей помогает раскрыть генетические пути, контролирующие развитие конечностей. [16] Система Тьюринга позволила получить ряд возможных результатов на этапах эволюции сетей формирования паттернов. [1]
| Таблица 1: Различные гены, которые, как известно, отвечают за развитие конечностей (отдельные по семействам генов) |
|---|
| Бмп2, Бмп4, Бмп7 |
| Дача1 |
| En1 |
| Fgf4, Fgf8, Fgf9, Fgf10, Fgf17, Fgfr1 |
| Гли3 |
| Гремлин1 |
| Рука2 |
| Хокса13, Хокса13 |
| Мсх1, Мсх2 |
| Питкс1 |
| Тссс |
| ТВх4, ТВх5 |
| Внт3, Внт5а |
Многие из генов, перечисленных в Таблице 1, играют важную роль в эмбриональном развитии, особенно во время формирования скелетного паттерна и зачатков конечностей. [18] Ген Shh и гены, принадлежащие к семействам BMP, Hox, T-box, FGF и Wnt, играют ключевую роль в передаче сигналов и дифференцировке клеток , регулируя и способствуя успешному формированию конечностей. Различные другие гены, перечисленные выше, одним из примеров является Dach1, представляют собой ДНК-связывающие белки, которые регулируют экспрессию генов. Сложная комбинация экспрессии, регуляции, активации и деактивации генов позволяет этим генам производить конечности во время эмбрионального развития. Интересно, что многие из этих генов сохраняются даже у животных, не имеющих конечностей, например у змей.
Эволюция змей и потеря конечностей
[ редактировать ]
Интересным аспектом в понимании развития конечностей является рассмотрение вопроса о том, как змеи потеряли ноги. Змеи являются особенно хорошим примером для изучения потери конечностей, поскольку на протяжении своей эволюции они неоднократно теряли и восстанавливали конечности, прежде чем окончательно потеряли ноги. Большая часть экспрессии генов во время эмбрионального развития регулируется посредством пространственно-временной и хемотаксической передачи сигналов. [20] как показано на изображении справа. Недавние данные свидетельствуют о том, что высококонсервативные гены, ответственные за развитие конечностей (таблица 1), все еще присутствуют у позвоночных без конечностей. [21] указывая на то, что во время эмбрионального развития образование конечностей или их отсутствие лучше всего можно объяснить регуляцией генов.
Возможная роль энхансера Shh в потере конечностей змей
[ редактировать ]Одна теория предполагает, что деградация последовательностей энхансеров могла способствовать прогрессирующей потере конечностей в змей эволюции . В частности, многие исследования были сосредоточены на регуляторной последовательности ZPA (ZRS) — энхансере гена Sonic Hedgehog ( Shh ). Этот энхансер дальнего действия необходим для правильного формирования конечностей у нескольких позвоночных видов , при этом известно, что мутации в этой последовательности вызывают деформации конечностей. [17] По существу, эта последовательность высоко консервативна у множества видов позвоночных.

Сравнение основных ZRS у нескольких видов змей с последовательностями мышей и ящериц показывает наличие специфичных для змей изменений. [23] Ядро ZRS, как оказалось, в основном сохранилось у базальных змей, таких как удав и питон , которые все еще содержат тазового пояса. кости [22] Напротив, у продвинутых змей, таких как гадюка и кобра , у которых не осталось структур скелетных конечностей, наблюдается гораздо более высокий уровень нуклеотидных изменений по сравнению с ZRS мышей и ящериц. [22] Считается, что эти кумулятивные изменения в ZRS змей указывают на прогрессирующую потерю функции этого энхансера на протяжении эволюции змей. [22]

Дальнейшее исследование этих изменений показало повышенную скорость замен в сайтах связывания для факторов транскрипции, таких как ETS1 , чье связывание с ZRS, как было показано, активирует транскрипцию Shh. [24] Эта деградация ZRS предполагает, что этот энхансер может быть важен для дальнейшего изучения молекулярных механизмов, которые стимулировали морфологическую эволюцию змей.
Текущие выводы и ограничения
[ редактировать ]Змеи не являются обычным модельным организмом, т.е. их нелегко генетически подчинить. Кроме того, их данные о последовательностях генома неполны и страдают от плохих аннотаций и качества. Эти факторы затрудняют понимание механизма потери конечностей змей с использованием генетического подхода, определения и наблюдения за присутствием и активностью этих генов и их регуляторных усилителей. Многие из генов, необходимых для формирования конечностей, все еще сохраняются у змей, поэтому потерю конечностей, вероятно, нельзя объяснить потерей генов. [ нужна ссылка ]
Осевое образование рисунка и связанные с ним проблемы
[ редактировать ]Развивающаяся конечность должна выровняться относительно трех осей симметрии. [25] Это краниокаудальная (от головы к хвосту), дорсовентральная (сзади вперед) и проксимодистальная (от ближней к дальней) оси. [25]
На многие исследования развития структуры скелета конечностей повлияла концепция позиционной информации, предложенная Льюисом Вулпертом в 1971 году. [26] В соответствии с этой идеей были предприняты усилия по идентификации диффузных сигнальных молекул ( морфогенов ), которые пересекают ортогональные оси развивающихся конечностей и определяют расположение и идентичность элементов скелета в зависимости от концентрации.
Проксимодистальный паттерн
[ редактировать ]Hox-гены способствуют спецификации шилоподов , зевгоподов и автоподов . Мутации в Hox-генах приводят к проксимодистальным потерям или аномалиям. [27] Для объяснения структуры этих регионов были предложены три разные модели.
Модель зоны прогресса
[ редактировать ]Апикальный эктодермальный гребень (AER) создает и поддерживает зону пролиферации клеток, известную как зона прогресса . [28] Считается, что здесь клетки получают позиционную информацию, необходимую им для перемещения в предназначенное положение. [28] Было высказано предположение, что их позиционное значение определялось продолжительностью времени, в течение которого клетки находились в зоне прогресса, но это еще предстоит доказать (по состоянию на 2001 г.). [28] Проксимальные структуры предполагалось образовывать первыми покинувшими зону клетками, а дистальные — клетками, вышедшими позже. [28]
Модель зоны прогресса была предложена 30 лет назад, но недавние данные противоречили этой модели. [29]
Экспериментальные доказательства:
- Удаление AER на более позднем периоде развития приводит к меньшему разрушению дистальных структур, чем если бы AER был удален на ранних стадиях развития.
- Прививка кончика зачатка ранней конечности на позднее крыло приводит к дублированию структур, а прививка кончика зачатка позднего крыла на раннюю конечность приводит к удалению структур.
Модель раннего распределения и расширения предшественников (или модель предварительной спецификации)
[ редактировать ]Клетки определены для каждого сегмента раннего зачатка конечности, и эта популяция клеток увеличивается по мере роста зачатка конечности. Эта модель согласуется со следующими наблюдениями. Деление клеток наблюдается по всему зачатку конечности. Гибель клеток происходит в зоне размером 200 мкм, прилегающей к AER, когда он удаляется; гибель клеток исключает возможность формирования некоторых паттернов. Гранулы, высвобождающие FGF, способны спасти развитие конечностей при удалении AER, предотвращая гибель клеток.
Экспериментальные доказательства:
- Меченые клетки в разных положениях раннего зачатка конечности были ограничены отдельными сегментами конечности. [30]
- В конечностях, в которых отсутствует экспрессия необходимых FGF4 и FGF8, наблюдаются все структуры конечности, а не только проксимальные части. [31]
Однако совсем недавно исследователи, ответственные за модели зоны прогресса и предварительной спецификации, признали, что ни одна из этих моделей не объясняет адекватно имеющиеся экспериментальные данные. [29]
Реакционно-диффузионная модель типа Тьюринга
[ редактировать ]
Эта модель реакции-диффузии, впервые предложенная в 1979 году, [33] основан на самоорганизации свойствах возбудимых сред, описанных Аланом Тьюрингом в 1952 году. [34] Возбудимой средой является мезенхима зачатка конечности, в которой клетки взаимодействуют посредством положительно ауторегуляторных морфогенов, таких как трансформирующий фактор роста бета (TGF-β) и ингибирующие сигнальные пути, включающие фактор роста фибробластов (FGF) и Notch . Проксимодистальная и краниокаудальная оси не считаются независимо определенными, а вместо этого возникают в результате изменения количества параллельных элементов, когда недифференцированная апикальная зона растущего зачатка конечности подвергается изменению формы. [35] Эта модель определяет только шаблон «голого скелета». Вместо этого предполагается, что другие факторы, такие как белки sonic hedgehog (Shh) и Hox, первичные информационные молекулы в других моделях, играют роль точной настройки.
Экспериментальные доказательства:
- Мезенхимальные клетки конечностей, когда их диссоциируют и выращивают в культуре или повторно вводят в эктодермальные «корпуса», могут повторять существенные аспекты формирования паттернов , морфогенеза и дифференцировки . [36] [37]
- Особенности структуры скелета конечностей у мышей-мутантов Doublefoot являются предсказанными результатами механизма типа Тьюринга. [38]
- Прогрессирующее уменьшение количества дистальных Hox-генов на фоне Gli3 -null приводит к постепенно более тяжелой полидактилии, с появлением более тонких и плотно упакованных пальцев, что позволяет предположить (с помощью компьютерного моделирования), что доза дистальных Hox-генов модулирует период или длину волны указанных пальцев. механизмом типа Тьюринга. [39]
Краниокаудальный паттерн
[ редактировать ]
В 1957 году открытие зоны поляризующей активности (ZPA) в зачатке конечности предоставило модель для понимания активности формирования паттерна под действием морфогенного градиента звукового ежа (Shh). [40] Shh признан энхансером, специфичным для конечностей. [41] Shh одновременно достаточно и необходимо для создания ZPA и определения краниокаудального паттерна в дистальной конечности (Shh не является необходимым для полярности шилопода). Shh включается сзади посредством ранней экспрессии генов Hoxd, экспрессии Hoxb8 и экспрессии dHAND. Shh поддерживается сзади через петлю обратной связи между ZPA и AER. Shh индуцирует AER выработку FGF4 и FGF8 , которые поддерживают экспрессию Shh.
Цифры 3,4 и 5 указаны временным градиентом Шх. Цифра 2 соответствует диффузионной форме слова «Тсс» на большие расстояния, а цифра 1 не требует «Тсс». Shh расщепляет комплекс транскрипционного репрессора Ci/Gli3, превращая фактор транскрипции Gli3 в активатор, который активирует транскрипцию генов HoxD вдоль краниокаудального отдела. Потеря репрессора Gli3 приводит к образованию родовых (неиндивидуализированных) пальцев в дополнительных количествах. [42]
Дорсовентральный рисунок
[ редактировать ]Формирование дорсовентрального паттерна опосредовано сигналами Wnt7a в вышележащей эктодерме, а не в мезодерме. Wnt7a необходим и достаточен для дорсализации конечности. Wnt7a также влияет на краниокаудальную часть , и потеря Wnt7a приводит к тому, что дорсальная сторона конечностей становится вентральной стороной и вызывает отсутствие задних пальцев. Замена сигналов Wnt7a устраняет этот дефект. Wnt7a также необходим для поддержания экспрессии Shh.
Lmx1b, гена LIM Homeobox (и, следовательно, фактора транскрипции Wnt7a также вызывает экспрессию ). Lmx1b участвует в дорсализации конечности, что было показано при нокауте гена Lmx1b у мышей. [43] У мышей, лишенных Lmx1b, на обеих сторонах лап образовалась вентральная кожа. Считается, что существуют и другие факторы, контролирующие формирование паттерна DV; Engrailed-1 подавляет дорсализующий эффект Wnt7a на вентральной стороне конечностей. [44]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Стюарт, штат Калифорния; Бхат, Р; Ньюман, SA (2017). «Эволюционное происхождение рисунка цифр» . ЭвоДево . 8:21 . дои : 10.1186/s13227-017-0084-8 . ПМЦ 5697439 . ПМИД 29201343 .
- ^ Стратфорд Т., Хортон С., Маден М. (1996). «Ретиноевая кислота необходима для начала роста зачатка конечности цыпленка» . Курр Биол . 6 (9): 1124–33. Бибкод : 1996CBio....6.1124S . дои : 10.1016/S0960-9822(02)70679-9 . ПМИД 8805369 . S2CID 14662908 .
- ^ Чжао X, Сирбу И.О., Мик Ф.А. и др. (июнь 2009 г.). «Ретиноевая кислота способствует индукции конечностей за счет воздействия на растяжение осей тела, но не является необходимой для формирования рисунка конечностей» . Курс. Биол . 19 (12): 1050–7. Бибкод : 2009CBio...19.1050Z . дои : 10.1016/j.cub.2009.04.059 . ПМК 2701469 . ПМИД 19464179 .
- ^ Каннингем, Ти Джей; Дустер, Г. (2015). «Механизмы передачи сигналов ретиноевой кислоты и ее роль в развитии органов и конечностей» . Нат. Преподобный мол. Клеточная Биол . 16 (2): 110–123. дои : 10.1038/nrm3932 . ПМК 4636111 . ПМИД 25560970 .
- ^ Перейти обратно: а б Тикл, К. (октябрь 2015 г.). «Как эмбрион формирует конечность: детерминация, полярность и идентичность» . Журнал анатомии . 227 (4): 418–30. дои : 10.1111/joa.12361 . ПМК 4580101 . ПМИД 26249743 .
- ^ Парр, бакалавр; МакМахон, AP (23 марта 1995 г.). «Дорсализующий сигнал Wnt-7a необходим для нормальной полярности осей DV и AP конечности мыши». Природа . 374 (6520): 350–3. Бибкод : 1995Natur.374..350P . дои : 10.1038/374350a0 . ПМИД 7885472 . S2CID 4254409 .
- ^ Пизетт, С; Абате-Шен, К; Нисвандер, Л. (ноябрь 2001 г.). «BMP контролирует проксимодистальный рост посредством индукции апикального эктодермального гребня и формирования дорсовентрального паттерна в конечностях позвоночных». Разработка . 128 (22): 4463–74. дои : 10.1242/dev.128.22.4463 . ПМИД 11714672 .
- ^ Йоней-Тамура С., Эндо Т., Ядзима Х., Охучи Х., Иде Х., Тамура К. (1999). «FGF7 и FGF10 непосредственно индуцируют апикальный эктодермальный гребень у куриных эмбрионов» . Дев. Биол . 211 (1): 133–43. дои : 10.1006/dbio.1999.9290 . ПМИД 10373311 .
- ^ Охучи Х, Такеучи Дж, Ёсиока Х, Ишимару Ю, Огура К, Такахаши Н, Огура Т, Ноджи С (1998). «Корреляция идентичности крыльев-ног в эктопических химерных конечностях, индуцированных FGF, с дифференциальной экспрессией Tbx5 и Tbx4 у кур». Разработка . 125 (1): 51–60. дои : 10.1242/dev.125.1.51 . ПМИД 9389663 .
- ^ Родригес-Эстебан С, Цукуи Т, Йоней С, Магаллон Дж, Тамура К, Исписуа Бельмонте ХК (1999). «Гены T-box Tbx4 и Tbx5 регулируют рост и идентичность конечностей». Природа . 398 (6730): 814–8. Бибкод : 1999Natur.398..814R . дои : 10.1038/19769 . ПМИД 10235264 . S2CID 4330287 .
- ^ ДеЛиз, AM; Фишер, Л; Туан, Р.С. (сентябрь 2000 г.). «Клеточные взаимодействия и передача сигналов в развитии хряща» . Остеоартрит и хрящ . 8 (5): 309–34. дои : 10.1053/joca.1999.0306 . ПМИД 10966838 .
- ^ Холл Б.К., Мияке Т. (2000). «Все за одного и один за всех: сгущения и начало развития скелета». Биоэссе . 22 (2): 138–47. doi : 10.1002/(SICI)1521-1878(200002)22:2<138::AID-BIES5>3.0.CO;2-4 . ПМИД 10655033 .
- ^ Ньюман С.А., Бхат Р. (2007). «Динамика активатора-ингибитора формирования паттерна конечностей позвоночных». Исследование врожденных дефектов, часть C: Эмбрион сегодня: обзоры . 81 (4): 305–19. CiteSeerX 10.1.1.128.3260 . дои : 10.1002/bdrc.20112 . ПМИД 18228262 .
- ^ Леал, Франциска; Кон, Мартин Дж. (07 ноября 2016 г.). «Потеря и повторное появление ног у змей в результате модульной эволюции Sonic hedgehog и усилителей HOXD» . Современная биология . 26 (21): 2966–2973. Бибкод : 2016CBio...26.2966L . дои : 10.1016/j.cub.2016.09.020 . ISSN 0960-9822 . ПМИД 27773569 . S2CID 4495482 .
- ^ Кон, Мартин Дж. (07 октября 2008 г.), «Механизмы развития эволюции конечностей позвоночных», в Кардью, Гейл; Гуд, Джейми А. (ред.), Молекулярная основа скелетогенеза , Симпозиумы Фонда Novartis, том. 232, Чичестер, Великобритания: John Wiley & Sons, Ltd, стр. 47–62, doi : 10.1002/0470846658.ch5 , ISBN 978-0-471-49433-1 , PMID 11277086
- ^ Перейти обратно: а б с д Инфанте, ЧР; Расис, А.М.; Менке, Д.Б. (январь 2018 г.). «Придатки и сети регуляции генов: уроки без конечностей» . Бытие . 56 (1): e23078. дои : 10.1002/dvg.23078 . ПМЦ 5783778 . ПМИД 29076617 .
- ^ Перейти обратно: а б с Лил Ф., Кон М.Дж. (2016). «Потеря и повторное появление ног у змей в результате модульной эволюции Sonic hedgehog и усилителей HOXD» . Курр Биол . 26 (21): 2966–2973. Бибкод : 2016CBio...26.2966L . дои : 10.1016/j.cub.2016.09.020 . ПМИД 27773569 . S2CID 4495482 .
- ^ Пощекочи, Шерил; Тауэрс, Мэтью (2017). «Сигнализация Sonic Hedgehog в развитии конечностей» . Границы клеточной биологии и биологии развития . 5 : 14. дои : 10.3389/fcell.2017.00014 . ISSN 2296-634X . ПМЦ 5328949 . ПМИД 28293554 .
- ^ Харф, Брайан Д.; Шерц, Пол Дж.; Ниссим, Сахар; Тянь, Хуа; МакМахон, Эндрю П.; Табин, Клиффорд Дж. (20 августа 2004 г.). «Доказательства временного градиента Shh, основанного на расширении, при определении идентичности пальцев позвоночных» . Клетка . 118 (4): 517–528. дои : 10.1016/j.cell.2004.07.024 . ISSN 0092-8674 . ПМИД 15315763 . S2CID 16280983 .
- ^ Харф, Брайан Д.; Шерц, Пол Дж.; Ниссим, Сахар; Тянь, Хуа; МакМахон, Эндрю П.; Табин, Клиффорд Дж. (20 августа 2004 г.). «Доказательства временного градиента Shh, основанного на расширении, при определении идентичности пальцев позвоночных» . Клетка . 118 (4): 517–528. дои : 10.1016/j.cell.2004.07.024 . ISSN 0092-8674 . ПМИД 15315763 . S2CID 16280983 .
- ^ Квон Евгений З.; Камнева Ольга К.; Мело, Уира С.; Бароцци, Ирос; Остервальдер, Марко; Мэннион, Брэндон Дж.; Тиссьер, Вирджиния; Пикл, Кэтрин С.; Плайзер-Фрик, Ингрид; Ли, Элизабет А.; Като, Момоэ (20 октября 2016 г.). «Прогрессирующая потеря функции усилителя конечностей во время эволюции змеи» . Клетка . 167 (3): 633–642.e11. дои : 10.1016/j.cell.2016.09.028 . ISSN 0092-8674 . ПМЦ 5484524 . PMID 27768887 .
- ^ Перейти обратно: а б с д и Квон Э.З., Камнева О.К., Мело У.С., Бароцци И., Остервальдер М., Маннион Б.Дж. и др. (2016). «Прогрессирующая потеря функции усилителя конечностей во время эволюции змеи» . Клетка . 167 (3): 633–642.e11. дои : 10.1016/j.cell.2016.09.028 . ПМЦ 5484524 . PMID 27768887 .
- ^ Берджесс DJ (2016). «Развитие: Звуковые змеи и регуляция формирования конечностей» . Нат преподобный Жене . 17 (12): 715. doi : 10.1038/nrg.2016.149 . ПМИД 27843141 . S2CID 28930888 .
- ^ Сюй С, Ян X, Чжоу Х, Ли Ю, Син С, Чжоу Т и др. (2020). «Новый вариант ZRS вызывает преаксиальную полидактилию I типа за счет увеличения экспрессии звукового ежа в развивающейся зачатке конечности» . Генет Мед . 22 (1): 189–198. дои : 10.1038/s41436-019-0626-7 . ПМК 6944640 . ПМИД 31395945 .
- ^ Перейти обратно: а б Ларсен, Уильям (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. п. 335. ИСБН 978-0-443-06583-5 .
- ^ Вулперт Л. (1971). «Глава 6. Информация о положении и формирование шаблона». Позиционная информация и формирование паттернов . Текущие темы биологии развития. Том. 6. стр. 183–224. дои : 10.1016/S0070-2153(08)60641-9 . ISBN 9780121531065 . ПМИД 4950136 .
- ^ Велик Д., Капечки М. (2003). «Гены Hox10 и Hox11 необходимы для глобального формирования структуры скелета млекопитающих». Наука . 301 (5631): 363–7. Бибкод : 2003Sci...301..363W . дои : 10.1126/science.1085672 . ПМИД 12869760 . S2CID 9751891 .
- ^ Перейти обратно: а б с д Ларсен, Уильям (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. п. 337. ИСБН 978-0-443-06583-5 .
- ^ Перейти обратно: а б Табин С., Вольперт Л. (2007). «Переосмысление проксимодистальной оси конечностей позвоночных в молекулярную эпоху» . Генс Дев . 21 (12): 1433–42. дои : 10.1101/gad.1547407 . ПМИД 17575045 .
- ^ Дадли А., Рос М., Табин С. (2002). «Повторное исследование проксимодистального паттерна во время развития конечностей позвоночных». Природа . 418 (6897): 539–44. Бибкод : 2002Natur.418..539D . дои : 10.1038/nature00945 . ПМИД 12152081 . S2CID 1357061 .
- ^ Сан X, Мариани Ф, Мартин Дж (2002). «Функции передачи сигналов FGF от апикального эктодермального гребня в развитии конечностей». Природа . 418 (6897): 501–8. Бибкод : 2002Natur.418..501S . дои : 10.1038/nature00902 . ПМИД 12152071 . S2CID 4409248 .
- ^ Гилберт, Сан-Франциско; Барреси, MJF (май 2017 г.). «Биология развития» . Американский журнал медицинской генетики, часть A. 173 (5) (11-е изд.): 1430. doi : 10.1002/ajmg.a.38166 . ISSN 1552-4825 .
- ^ Ньюман С.А., Фриш Х.Л. (1979). «Динамика формирования скелетного рисунка развивающихся конечностей цыпленка». Наука . 205 (4407): 662–668. Бибкод : 1979Sci...205..662N . дои : 10.1126/science.462174 . ПМИД 462174 . S2CID 44653825 .
- ^ Тьюринг, AM (1952). «Химические основы морфогенеза» . Философские труды Королевского общества Б. 237 (641): 37–72. Бибкод : 1952РСПТБ.237...37Т . дои : 10.1098/rstb.1952.0012 .
- ^ Чжу Дж., Чжан Ю.Т., Альбер М.С., Ньюман С.А. (2010). «Формирование структуры голых костей: основная регуляторная сеть различной геометрии воспроизводит основные особенности развития и эволюции конечностей позвоночных» . ПЛОС ОДИН . 5 (5): е:10892. Бибкод : 2010PLoSO...510892Z . дои : 10.1371/journal.pone.0010892 . ПМЦ 2878345 . ПМИД 20531940 .
- ^ Мофтах М.З., Дауни С.А., Бронштейн Н.Б., Мезенцева Н., Пу Дж., Махер П.А., Ньюман С.А. (2002). «Эктодермальные FGF индуцируют перинодулярное ингибирование хондрогенеза конечностей in vitro и in vivo через рецептор FGF 2» . Дев. Биол . 249 (2): 270–82. дои : 10.1006/dbio.2002.0766 . ПМИД 12221006 .
- ^ Рос М.А., Лайонс Дж.Э., Макем С., Фэллон Дж.Ф. (1994). «Рекомбинантные конечности как модель для изучения регуляции гомеобоксных генов во время развития конечностей». Дев. Биол . 166 (1): 59–72. дои : 10.1006/dbio.1994.1296 . ПМИД 7958460 .
- ^ Миура Т., Шиота К., Моррисс-Кей Дж., Майни ПК (2006). «Смешанный режим в модели реакции-диффузии конечностей мутантной мыши Doublefoot - Тьюринга в растущем домене во время развития конечностей» . J Теория Биол . 240 (4): 562–73. Бибкод : 2006JThBi.240..562M . дои : 10.1016/j.jtbi.2005.10.016 . ПМИД 16364368 .
- ^ Шет Р., Маркон Л., Бастида М.Ф., Юнко М., Кинтана Л., Дан Р., Кмита М., Шарп Дж., Рос М.А. (2012). «Гены Hox регулируют последовательность цифр, контролируя длину волны механизма Тьюринга» . Наука . 338 (6113): 1476–1480. Бибкод : 2012Sci...338.1476S . дои : 10.1126/science.1226804 . ПМЦ 4486416 . ПМИД 23239739 .
- ^ Чжу, Дж; Макем, С. (15 сентября 2017 г.). «ZPA Джона Сондерса, еж Соника и цифровая идентичность — как все это на самом деле работает?» . Биология развития . 429 (2): 391–400. дои : 10.1016/j.ydbio.2017.02.001 . ПМК 5540801 . ПМИД 28161524 .
- ^ Лил Ф., Кон М.Дж. (январь 2018 г.). «Эволюционное, генетическое и геномное понимание эволюционной потери конечностей у змей». Бытие . 56 (1): e23077. дои : 10.1002/dvg.23077 . ПМИД 29095557 . S2CID 4510082 .
- ^ Чан С., Литингтунг И., Харрис М., Симандл Б., Ли Ю., Бичи П., Фэллон Дж. (2001). «Проявление препаттерна конечностей: развитие конечностей при отсутствии функции звукового ежа» . Дев Биол . 236 (2): 421–35. дои : 10.1006/dbio.2001.0346 . ПМИД 11476582 .
- ^ Риддл Р.Д., Энсини М., Нельсон С., Цучида Т., Джесселл Т.М., Табин С. (1995). « Индукция гомеобоксного гена LIM Lmx1b с помощью WNT7a устанавливает дорсовентральный паттерн в конечностях позвоночных» . Клетка . 83 (2): 631–40. дои : 10.1016/0092-8674(95)90103-5 . ПМИД 7585966 . S2CID 16272219 .
- ^ Джонсон Р.Л., Табин С.Дж. (19 сентября 1997 г.). «Молекулярные модели развития конечностей позвоночных» . Клетка . 90 (6): 979–990. дои : 10.1016/S0092-8674(00)80364-5 . ПМИД 9323126 .