Пространственно-временная экспрессия генов
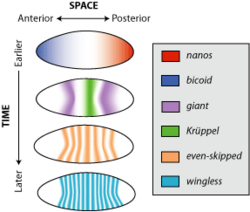
-временная экспрессия генов — это активация генов тканях в определенных Пространственно организма в определенные моменты развития . Паттерны активации генов сильно различаются по сложности. Некоторые из них просты и статичны, например, структура тубулина, которая экспрессируется во всех клетках на всех этапах жизни. Некоторые, с другой стороны, чрезвычайно сложны, их трудно предсказать и смоделировать, при этом экспрессия сильно колеблется от минуты к минуте или от клетки к клетке. Пространственно-временные вариации играют ключевую роль в создании разнообразия типов клеток , обнаруженных в развитых организмах; поскольку идентичность клетки определяется набором генов, активно экспрессируемых внутри этой клетки, если бы экспрессия генов была однородной в пространстве и времени, мог бы существовать не более одного типа клеток.
Рассмотрим ген wingless, член семейства wnt генов . На раннем этапе эмбрионального развития модельного организма Drosophila melanogaster , или плодовой мушки, бескрылость выражена почти по всему эмбриону в виде чередующихся полос, разделенных тремя клетками. Этот образец теряется к тому времени, когда организм превращается в личинку, но бескрылость все еще проявляется в различных тканях, таких как имагинальные диски крыльев , участки ткани, которые разовьются во взрослые крылья. Пространственно -временной паттерн экспрессии генов бескрылых определяется сетью регуляторных взаимодействий, состоящей из эффектов множества различных генов, таких как Even-skiped и Krüppel.
Что вызывает пространственные и временные различия в экспрессии одного гена? Поскольку нынешние паттерны экспрессии строго зависят от предыдущих паттернов экспрессии, существует регрессивная проблема объяснения того, что вызвало первые различия в экспрессии генов. Процесс, посредством которого однородная экспрессия генов становится дифференцированной в пространстве и времени, известен как нарушение симметрии . Например, в случае эмбрионального развития дрозофилы гены nanos и bicoid асимметрично экспрессируются в ооците , поскольку материнские клетки откладывают информационную РНК (мРНК) для этих генов в полюсах яйца до его откладки .

Выявление пространственно-временных закономерностей
[ редактировать ]Один из способов определить характер экспрессии конкретного гена — поместить репортерный ген ниже его промотора. В этой конфигурации ген-промотор будет вызывать экспрессию репортерного гена только там и тогда, когда экспрессируется интересующий ген. Распределение экспрессии репортерного гена можно определить путем его визуализации. Например, зеленый флуоресцентный белок репортерного гена можно визуализировать, стимулируя его синим светом, а затем используя цифровую камеру для регистрации зеленого флуоресцентного излучения.
Если промотор интересующего гена неизвестен, существует несколько способов определить его пространственно-временное распределение. Иммуногистохимия включает получение антитела со специфическим сродством к белку, связанному с интересующим геном. Это распределение этого антитела затем можно визуализировать с помощью такого метода, как флуоресцентное мечение. Преимущество иммуногистохимии состоит в том, что она методологически осуществима и относительно недорога. К его недостаткам относится неспецифичность антитела, приводящая к ложноположительной идентификации экспрессии. Плохая пенетрантность антитела в ткань-мишень может привести к ложноотрицательным результатам. Более того, поскольку иммуногистохимия визуализирует белок, генерируемый геном, если белковый продукт диффундирует между клетками или имеет особенно короткий или длинный период полураспада по сравнению с мРНК , которая используется для трансляции белка, это может привести к искаженной интерпретации, которая клетки экспрессируют мРНК .
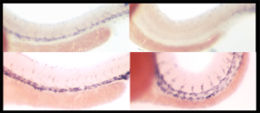
in situ Гибридизация — это альтернативный метод, при котором к ткани добавляется «зонд» — синтетическая нуклеиновая кислота с последовательностью, комплементарной мРНК гена. Затем этот зонд химически помечается, чтобы его можно было позже визуализировать. Этот метод позволяет визуализировать именно клетки, продуцирующие мРНК, без каких-либо артефактов, связанных с иммуногистохимией. Однако это чрезвычайно сложно и требует знания последовательности ДНК , соответствующей интересующему гену.
Метод, называемый скринингом энхансерных ловушек , выявляет разнообразие возможных в организме пространственно-временных паттернов экспрессии генов. В этом методе ДНК, кодирующая репортерный ген, вставляется в геном случайным образом. гена, В зависимости от промоторов расположенных проксимальнее точки вставки, репортерный ген будет экспрессироваться в определенных тканях в определенные моменты развития. Хотя паттерны экспрессии, полученные с помощью энхансерных ловушек, не обязательно отражают реальные паттерны экспрессии конкретных генов, они раскрывают разнообразие пространственно-временных паттернов, доступных для эволюции.
Репортерные гены можно визуализировать в живых организмах, но как иммуногистохимию, так и гибридизацию in situ необходимо проводить в фиксированных тканях. Методы, требующие фиксации тканей, могут генерировать только одну временную точку для каждого отдельного организма. Однако использование живых животных вместо фиксированных тканей может иметь решающее значение для динамического понимания закономерностей экспрессии на протяжении всей жизни человека. В любом случае различия между людьми могут затруднить интерпретацию временных паттернов экспрессии.
Методы контроля пространственно-временной экспрессии генов
[ редактировать ]В настоящее время разрабатывается несколько методов контроля экспрессии генов в пространстве, времени и в различной степени. Один из методов заключается в использовании системы индуктор/репрессор оперона , которая обеспечивает временной контроль экспрессии генов. Для пространственного контроля экспрессии генов разрабатываются струйные принтеры для печати лигандов на гелевой культуре. [1] Другой популярный метод предполагает использование света для контроля экспрессии генов в пространственно-временном режиме. Поскольку свет также можно легко контролировать в пространстве, времени и степени, существует несколько методов контроля экспрессии генов на уровне ДНК и РНК. [2] разработаны и находятся в стадии изучения. Например, интерференцию РНК можно контролировать с помощью света. [3] [4] а также формирование паттерна экспрессии генов было выполнено в монослое клеток. [5] и на эмбрионах рыбок данио с использованием морфолино в клетках [6] или пептид-нуклеиновая кислота [7] [8] [9] демонстрируя пространственно-временной контроль экспрессии генов. Недавно контроль на основе света был продемонстрирован на уровне ДНК с использованием системы на основе трансгена. [10] или клеточные триплексы, образующие олигонуклеотиды [11]
Ссылки
[ редактировать ]- ^ Коэн, диджей; Морфино, РЦ; Махарбиз, ММ (2009). «Модифицированный потребительский струйный принтер для пространственно-временного контроля экспрессии генов» . ПЛОС ОДИН . 4 (9): е7086. Бибкод : 2009PLoSO...4.7086C . дои : 10.1371/journal.pone.0007086 . ПМК 2739290 . ПМИД 19763256 .
- ^ Андо, Хидеки; Фурута, Тошиаки; Цянь, Роджер Ю.; Окамото, Хитоши (2001). «Фотоопосредованная активация генов с использованием клеточной РНК / ДНК в эмбрионах рыбок данио». Природная генетика . 28 (4): 317–325. дои : 10.1038/ng583 . ПМИД 11479592 . S2CID 6773535 .
- ^ Шах, Самит; Рангараджан, Субхашри; Фридман, Саймон Х. (2005). «Светоактивируемая интерференция РНК». Angewandte Chemie, международное издание . 44 (9): 1328–1332. дои : 10.1002/anie.200461458 . ПМИД 15643658 .
- ^ Микат, Вера; Хекель, Александр (2007). «Светозависимая РНК-интерференция с миРНК, заключенными в нуклеиновые основания» . РНК . 13 (12): 2341–2347. дои : 10.1261/rna.753407 . ПМК 2080613 . ПМИД 17951332 .
- ^ Джайн, Пиюш К.; Шах, Самит; Фридман, Саймон Х. (2011). «Паттерн экспрессии генов с использованием новых фотолабильных групп, примененных к активированным светом РНКи». Журнал Американского химического общества . 133 (3): 440–446. дои : 10.1021/ja107226e . ПМИД 21162570 . S2CID 207058522 .
- ^ Шестопалов Илья А; Чен, Джеймс К. (2011). «Пространственно-временной контроль экспрессии эмбриональных генов с использованием морфолино в клетках». Рыбка данио: генетика, геномика и информатика . Методы клеточной биологии. Том. 104. стр. 151–72. дои : 10.1016/B978-0-12-374814-0.00009-4 . ISBN 9780123748140 . ПМК 4408312 . ПМИД 21924162 .
- ^ Тан, СиньЦзин; Маэгава, Синго; Вайнберг, Эрик С.; Дмоховский, Иван Дж. (2007). «Регулирование экспрессии генов в эмбрионах рыбок данио с использованием светоактивируемых отрицательно заряженных пептид-нуклеиновых кислот». Журнал Американского химического общества . 129 (36): 11000–11001. дои : 10.1021/ja073723s . ПМИД 17711280 .
- ^ Александр Хекель, Гюнтер Майер, «Химическая биология нуклеиновых кислот», глава 13. Светочувствительные нуклеиновые кислоты для пространственно-временного контроля биологических процессов. http://onlinelibrary.wiley.com/doi/10.1002/9780470664001.ch13/summary
- ^ Гован, Дж. М.; Дейтерс (2012) «Активация и деактивация антисмысловой и РНК-интерференционной функции светом А. От последовательностей нуклеиновых кислот к молекулярной медицине (под ред. В. А. Эрдманна и Дж. Барцишевского)», Springer, Гейдельберг, https://doi.org/10.1007% 2F978-3-642-27426-8_11
- ^ Ван, X; Чен, X; Ян, Ю (2012). «Пространственно-временной контроль экспрессии генов с помощью трансгенной системы с переключателем света» . Нат-методы . 9 (3): 266–9. дои : 10.1038/nmeth.1892 . ПМИД 22327833 . S2CID 26529717 .
- ^ Гован, Джин М.; Упрети, Раджендра; Хемфилл, Джеймс; Лайвли, Марк О.; Дейтерс, Александр (2012). «Регуляция транскрипции посредством световой активации и световой деактивации триплекс-образующих олигонуклеотидов в клетках млекопитающих» . АКС хим. Биол . 7 (7): 1247–1256. дои : 10.1021/cb300161r . ПМК 3401312 . ПМИД 22540192 .
Внешние ссылки
[ редактировать ]- Отчет FlyBase о бескрылом проявлении у плодовых мух
- Просмотрите пространственно-временные закономерности экспрессии генов, организованные по числу хромосом человека.
- Пространственно-временная экспрессия генов в Genevestigator
- Поиск генов млекопитающих с определенными паттернами экспрессии
- Паттерны экспрессии во время эмбриогенеза дрозофилы, выявленные с помощью гибридизации in situ. Архивировано 7 августа 2008 г. в Wayback Machine.