Зона поляризующей активности
| Зона поляризующей активности | |
|---|---|
 Апикальный эктодермальный гребень представляет собой утолщенный эпителий на самом дистальном конце зачатка конечности. Зона поляризующей активности находится в задней части зачатка конечности. | |
| Подробности | |
| Идентификаторы | |
| латинский | зона активности полярисантиса |
| Акроним(ы) | ЗПА |
| ТО | поляризующей активности_by_E5.0.3.0.0.1.5 E5.0.3.0.0.1.5 |
| Анатомическая терминология | |
Зона поляризующей активности (ZPA) представляет собой область мезенхимы , содержащую сигналы, которые дают команду развивающемуся зачатку конечности формироваться вдоль передней/задней оси. Зачаток конечности представляет собой недифференцированную мезенхиму, окруженную покровом эктодермы . Со временем из зачатка конечности развиваются кости, сухожилия, мышцы и суставы. Развитие зачатков конечностей зависит не только от ZPA, но и от множества различных генов, сигналов и уникальной области эктодермы, называемой апикальным эктодермальным гребнем (AER). Исследования Сондерса и Гасселинга в 1948 году выявили AER и его последующее участие в проксимально-дистальном отростке. [1] Двадцать лет спустя та же группа провела исследования по трансплантации зачатков конечностей кур и идентифицировала ZPA. Лишь в 1993 году Тодт и Фэллон показали, что AER и ZPA зависят друг от друга. [2]
Узоры
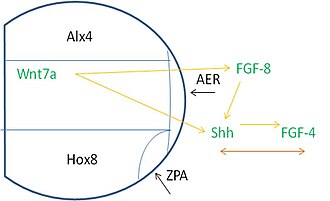
[ редактировать ]Формирование паттерна вдоль зачатка конечности требует сигналов из многих источников. В частности, белки, называемые факторами транскрипции (TF), помогают контролировать скорость транскрипции гена. Зачаток конечности экспрессирует ТФ, называемый ALX4, в передней части мезодермы , а ТФ HOXB8 экспрессируется в задней части. Область Alx4, медиальная область и область экспрессии Hox8 встречаются в проксимальной области, где развивается AER. ZPA формируется там, где регион Hox8 присоединяется к AER.

Эти регионы зависят от передачи сигналов для возникновения соответствующих индукционных событий. AER экспрессирует FGF8 , который индуцирует экспрессию Shh в задней мезодерме. Затем Shh стимулирует FGF4 экспрессию в задней части AER. После этих событий возникает взаимная зависимость между FGF-4 и Shh для их последующей экспрессии и поддержания. Кроме того, Wnt7a экспрессируется в дорсальной эктодерме и необходим для поддержания передачи сигналов FGF и Shh. [3]
Апикальный эктодермальный гребень
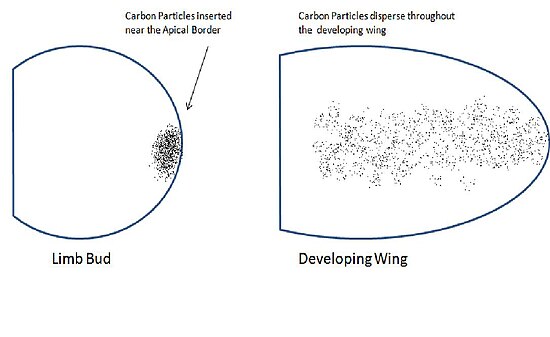
[ редактировать ]Сондерс и Гасселинг опубликовали данные в Журнале экспериментальной биологии в 1948 году, показав, что контрольные метки, вставленные возле края апикальной границы зачатка крыла, разбросаны по всему предплечью крыла. [1] Это заставило их поверить, что апикальная эктодерма может играть роль в формировании частей крыла. Чтобы проверить это, они удалили апикальную эктодерму из зачатков крыльев, в результате чего крылья были деформированы. Когда они удалили дорсальную эктодерму, образовались нормальные крылья. Эти результаты показали, что клетки апикальной эктодермы имеют четкую судьбу, формирующую определенные области крыла.
Соник ёжик
[ редактировать ]
В 1968 году Сондерс и Гасселинг провели исследования по трансплантации тканей из зачатков куриных конечностей. [4] Удалив клетки из задней области конечности, они пересадили их в переднюю область и заметили, что в передней области образовались дополнительные пальцы, и эти пальцы были зеркальным отражением нормальных. Этой задней мезенхимой была ZPA, которая, как теперь известно, экспрессирует белок sonic hedgehog (Shh). Одна из гипотез состоит в том, что при высоких концентрациях этот неизвестный морфоген вызывает образование мезенхимы на задней стороне, тогда как низкие концентрации индуцируют образование мезенхимы на переднем конце. [5] Идентификация этого морфогена стала следующим решающим шагом.Первая гипотеза исходила от Tickle et al. которые показали, что при помещении ретиноевой кислоты на передний край зачатка конечности возникают зеркальные удвоения. [6] Однако концентрации ретиноевой кислоты, которые вызывают зеркальные дупликации, индуцируют высокие уровни нижестоящего гена, рецептора ретиноевой кислоты бета, который не наблюдается в задней области. [7] Теперь известно, что эндогенная ретиноевая кислота действует разрешительно до зарождения зачатков конечностей, позволяя начать процесс зачатков. [8] и что специфический морфоген, предположительно Shh, [9] обычно экспрессируется независимо от ретиноевой кислоты в задней области зачатка конечности. Если посмотреть на сигнальные гомологи других организмов, то ген сегментации дрозофилы , ежа, оказался жизнеспособным кандидатом. [10]
Идея о том, что Shh необходим для правильной передачи сигналов ZPA и формирования передних/задних конечностей, нуждалась в проверке. Риддл и др. перенесли открытия Сондерса и Гасселингса на следующий шаг и доказали, что Shh является морфогеном внутри ZPA, который необходим для формирования передне-заднего паттерна. [9] Путем выделения гена Shh и имплантации его в зачаток передней конечности образовались зеркальные дубликаты пальцев.
Выделение проводили путем создания праймеров для ПЦР , соответствующих последовательностям Shh, консервативным у дрозофилы и мышей и участвующим в формировании зачатков конечностей. Затем клон использовали в качестве матрицы для скрининга библиотеки кДНК из РНК зачатка конечности на стадии 22. Группа эктопически экспрессировала ген, воспользовавшись ретровирусным вектором для вставки кДНК в куриные клетки. Существуют уникальные типы этого ретровирусного вектора, которые заражают только определенные штаммы видов птиц. Поэтому эта группа использовала ретровирусный вектор, названный RCAS-E, в котором отсутствует оболочечный белок типа E и который способен инфицировать определенные фибробласты куриных эмбрионов Shh.
![]()
Результаты показали дублирование цифр, наиболее частыми из которых были 4-3-3-4, при этом цифра 2 отсутствовала. Хотя существовала вариабельность, она явно соответствовала паттерну переднего и заднего положения. Вариации были обусловлены количеством пересаженной ткани и расположением трансплантата. Эти данные показывают, что Shh может заменить функцию ZPA. Таким образом, Shh достаточно для действия ZPA.
Медиаторы
[ редактировать ]Shh может быть критическим сигналом, регулирующим функцию ZPA, но гены, участвующие в передаче сигналов Shh, находятся под контролем нескольких других факторов, которые необходимы для поддержания и функционирования ZPA, включая Hand2 и Hoxb-8. Ретиноевая кислота , важная сигнальная молекула, необходимая на протяжении всего эмбриогенеза, действует через Hox-гены. Первоначально предполагалось, что ретиноевая кислота индуцирует ген Hoxb-8. [11] но эта гипотеза не была подтверждена генетическими исследованиями на эмбрионах мышей, лишенных синтеза ретиноевой кислоты, которые все еще экспрессируют Hoxb-8 в конечностях. [8] Передача сигналов Hoxb-8 активна у ранних эмбрионов, начиная с заднего конца мезодермы латеральной пластинки и распространяясь на переднюю область. По мере того как Hoxb-8 распространяется на большее количество передних областей, Shh индуцируется в области, которая станет ZPA. Shh индуцируется только в передней области из-за сигналов от AER. Эксперименты, проведенные Heikinheimo et al. показали, что при удалении AER шариков, экспрессирующих FGF, достаточно, чтобы индуцировать передачу сигналов Shh в ZPA. [12] Таким образом, вероятным сигнальным фактором AER является FGF.

Кроме того, когда AER удаляется, Shh больше не экспрессируется, и ZPA больше не может поддерживаться. Действуя по механизму положительной обратной связи, FGF-4 экспрессируется вблизи ZPA. [13] FGF-4 поддерживает экспрессию Shh, тогда как Shh поддерживает экспрессию FGF-4. В то же время Wnt-7a экспрессируется в дорсальной эктодерме и обеспечивает дополнительную положительную обратную связь с FGF-4 и Shh. [14] Без этой системы конечности и пальцы либо значительно уменьшаются, либо отсутствуют.
Нисходящие сигналы
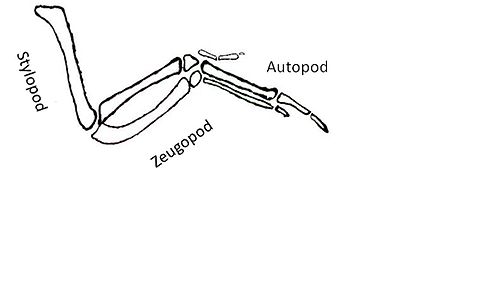
[ редактировать ]Цели, расположенные ниже по течению, которые активируются в ответ на «Тсс», представляют собой еще одну проблему. Гены, являющиеся мишенями передачи сигналов Shh, кодируют факторы, которые приводят к образованию аутоподов , шилоподов и зевгоподов .
Активация факторов транскрипции цинковых пальцев Gli происходит через сигнальный путь Hedgehog . Существует три фактора Gli, которые необходимы для развития конечностей: Gli1 , Gli2 и Gli3 . Без Shh Gli2 и Gli3 преобразуются в репрессорную форму и отправляются в ядро, чтобы подавить ответ Shh. Но когда присутствует Shh, непроцессированные Gli2 и Gli3 способны проходить в ядро и стимулировать экспрессию генов-мишеней Shh, включая Gli1. Исследования на мышах показывают, что нокауты Gli3 имеют полидактилию пальцев. [15] По сути, Shh устраняет репрессию Gli3. Когда Shh диффундирует из ZPA, он преобладает в задней области зачатка конечности, активируя Gli3 в задней области, в то время как репрессор все еще активен в передней области. Это приводит к активации других генов, таких как гены Hox, гены FGF и гены BMP в задней области, устанавливая формирование рисунка пальцев. BMP играет роль в морфологии конечностей, в частности, в расположении пальцев, но конкретная регуляция BMP неясна.

В частности, Hox-гены A и D, вероятно, контролируются Shh внутри ZPA. [16] Три фазы активации Hox-генов приводят к образованию паттерна конечностей, параллельному экспрессии Hox-генов по вложенному паттерну. Активация этих генов приводит к образованию новой оси конечностей, что в конечном итоге приводит к развитию пальцев, возможно, интерпретируя экспрессию генов для определения идентичности пальцев. В целом, молекулярный ZPA требует входной информации для нескольких сигнальных центров, но сам действует как организатор, индуцируя передне-заднее постукивание зачатка конечности цыпленка.
Ссылки
[ редактировать ]- ^ Jump up to: а б Сондерс Дж.В. (декабрь 1998 г.). «Проксимально-дистальная последовательность происхождения частей куриного крыла и роль эктодермы. 1948». Журнал экспериментальной зоологии . 282 (6): 628–68. doi : 10.1002/(SICI)1097-010X(19981215)282:6<628::AID-JEZ2>3.0.CO;2-N . ISSN 0022-104X . ПМИД 9846378 .
- ^ Тодт В.Л., Фэллон Дж.Ф. (1 ноября 1987 г.). «Удаление заднего апикального эктодермального гребня в зачатке крыла цыпленка запускает серию событий, приводящих к формированию дефектного переднего рисунка» . Разработка . 101 (3): 501–15. ISSN 0950-1991 . ПМИД 3502993 .
- ^ Пирс Р.В., Табин С.Дж. (декабрь 1998 г.). «Молекулярный ZPA». Журнал экспериментальной зоологии . 282 (6): 677–90. doi : 10.1002/(SICI)1097-010X(19981215)282:6<677::AID-JEZ4>3.0.CO;2-F . ISSN 0022-104X . ПМИД 9846380 .
- ^ Сондерс Дж.В., Гасселинг М.Т. (1968). «Эктодермально-мезенхимальные взаимодействия в возникновении симметрии конечностей». Эпителиально-мезенхимальные взаимодействия : 78–97.
- ^ Вулперт Л. (октябрь 1969 г.). «Позиционная информация и пространственная картина клеточной дифференциации». Журнал теоретической биологии . 25 (1): 1–47. дои : 10.1016/S0022-5193(69)80016-0 . ISSN 0022-5193 . ПМИД 4390734 .
- ^ Тикл С., Альбертс Б., Уолперт Л., Ли Дж. (апрель 1982 г.). «Локальное применение ретиноевой кислоты на связь конечностей имитирует действие поляризующей области». Природа . 296 (5857): 564–6. Бибкод : 1982Natur.296..564T . дои : 10.1038/296564a0 . ISSN 0028-0836 . ПМИД 7070499 . S2CID 4242623 .
- ^ Ноно Т., Ноджи С., Кояма Э. и др. (март 1991 г.). «Участие генов гомеобокса курицы Chox-4 в определении переднезадней осевой полярности во время развития конечностей». Клетка . 64 (6): 1197–205. дои : 10.1016/0092-8674(91)90274-3 . ISSN 0092-8674 . ПМИД 1672266 . S2CID 42393794 .
- ^ Jump up to: а б Чжао X, Сирбу И.О., Мик Ф.А. и др. (июнь 2009 г.). «Ретиноевая кислота способствует индукции конечностей за счет воздействия на растяжение осей тела, но не является необходимой для формирования рисунка конечностей» . Курс. Биол . 19 (12): 1050–7. дои : 10.1016/j.cub.2009.04.059 . ПМК 2701469 . ПМИД 19464179 .
- ^ Jump up to: а б Риддл Р.Д., Джонсон Р.Л., Лауфер Э., Табин С. (декабрь 1993 г.). «Звуковой ёж опосредует поляризующую активность ZPA». Клетка . 75 (7): 1401–16. дои : 10.1016/0092-8674(93)90626-2 . ISSN 0092-8674 . ПМИД 8269518 . S2CID 4973500 .
- ^ Ингхэм П.В., Фитц М.Дж. (апрель 1995 г.). «Количественное влияние активности ежей и декапентаплегии на формирование рисунка крыла дрозофилы» . Современная биология . 5 (4): 432–40. дои : 10.1016/S0960-9822(95)00084-4 . ISSN 0960-9822 . ПМИД 7627558 . S2CID 14426793 .
- ^ Лу ХК, Ревелли Дж.П., Геринг Л., Таллер С., Эйхеле Г. (1 мая 1997 г.). «Передача сигналов ретиноидов необходима для установления ZPA и для экспрессии Hoxb-8, медиатора образования ZPA» . Разработка . 124 (9): 1643–51. ISSN 0950-1991 . ПМИД 9165113 .
- ^ Хейкинхеймо М., Лоуше А., Шеклфорд Г.М., Уилсон Д.Б., Макартур, Калифорния (ноябрь 1994 г.). «Экспрессия Fgf-8 у мышей после гаструляции предполагает его роль в развитии лица, конечностей и центральной нервной системы». Механизмы развития . 48 (2): 129–38. дои : 10.1016/0925-4773(94)90022-1 . ISSN 0925-4773 . ПМИД 7873403 . S2CID 8587334 .
- ^ Нисвандер Л., Джеффри С., Мартин Г.Р., Тикл С. (октябрь 1994 г.). «Петля положительной обратной связи координирует рост и формирование конечностей позвоночных». Природа . 371 (6498): 609–12. Бибкод : 1994Natur.371..609N . дои : 10.1038/371609a0 . ISSN 0028-0836 . ПМИД 7935794 . S2CID 4305639 .
- ^ Ян Ю, Нисвандер Л (март 1995 г.). «Взаимодействие между сигнальными молекулами WNT7a и SHH во время развития конечностей позвоночных: дорсальные сигналы регулируют формирование переднезаднего паттерна» . Клетка . 80 (6): 939–47. дои : 10.1016/0092-8674(95)90297-X . ISSN 0092-8674 . ПМИД 7697724 . S2CID 7869066 .
- ^ Литингтунг Ю., Дан Р.Д., Ли Ю., Фэллон Дж.Ф., Чан С. (август 2002 г.). «Shh и Gli3 необязательны для формирования скелета конечностей, но регулируют количество и идентичность пальцев». Природа . 418 (6901): 979–83. Бибкод : 2002Natur.418..979L . дои : 10.1038/nature01033 . ISSN 0028-0836 . ПМИД 12198547 . S2CID 4431757 .
- ^ Нельсон CE, Морган Б.А., Берк AC и др. (1 мая 1996 г.). «Анализ экспрессии гена Hox в зачатке конечностей цыпленка» . Разработка . 122 (5): 1449–66. ISSN 0950-1991 . ПМИД 8625833 .