Тип
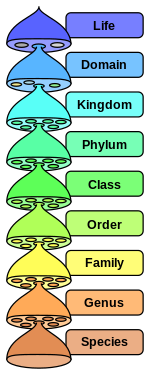
В биологии тип ниже ( / ˈ f aɪ l əm / ; мн.: тип ) это уровень классификации или таксономический ранг — царства и выше класса . Традиционно в ботанике термин «деление вместо термина «тип» использовался », хотя Международный кодекс номенклатуры водорослей, грибов и растений принимает эти термины как эквивалентные. [1] [2] [3] В зависимости от определений царство животных Animalia включает около 31 типа, царство растений Plantae — около 14 типов, а царство грибов Fungi — около 8 типов. Текущие исследования в области филогенетики раскрывают взаимоотношения между типами внутри более крупных клад, таких как Ecdysozoa и Embryophyta .
Общее описание
[ редактировать ]Термин «филум» был придуман в 1866 году Эрнстом Геккелем от греческого «филон» ( φῦλον , «раса, род»), связанного с типом ( φυλή , «племя, клан»). [4] [5] Геккель отмечал, что виды постоянно эволюционировали в новые виды, которые, по-видимому, сохраняли мало последовательных черт между собой и, следовательно, мало черт, отличающих их как группу («самостоятельное единство»): «возможно, такое реальное и вполне самодостаточное единство есть совокупность всех видов, которые постепенно развились из одной и той же общей исходной формы, как, например, все позвоночные животные. Мы называем эту совокупность [а] Штаммом [т. е. запасом] ( Филон )». [а] В таксономии растений Август В. Эйхлер (1883) разделил растения на пять групп , названных подразделениями - термин, который используется и сегодня для групп растений, водорослей и грибов. [1] [6] Определения зоологических типов изменились с момента их возникновения в шести классах Линнея и четырех ответвлениях Жоржа Кювье . [7]
Неофициально типы можно рассматривать как группы организмов, основанные на общей специализации строения тела . [8] По своей сути тип можно определить двумя способами: как группу организмов с определенной степенью морфологического сходства или сходства в развитии ( фенетическое определение) или как группу организмов с определенной степенью эволюционного родства ( филогенетическое определение). ). [9] Попытка определить уровень линнеевской иерархии без ссылки на (эволюционное) родство неудовлетворительна, но фенетическое определение полезно при решении вопросов морфологического характера, например, насколько успешными были различные конструкции тела. [ нужна ссылка ]
Определение, основанное на генетическом родстве
[ редактировать ]Наиболее важной объективной мерой в приведенных выше определениях является «определенная степень», которая определяет, насколько разные организмы должны быть членами разных типов. Минимальное требование состоит в том, что все организмы одного типа должны быть явно более тесно связаны друг с другом, чем с какой-либо другой группой. [9] Даже это проблематично, потому что требование зависит от знания взаимоотношений организмов: по мере того, как становится доступно больше данных, особенно в результате молекулярных исследований, мы можем лучше определять отношения между группами. Таким образом, типы могут быть объединены или разделены, если станет очевидно, связаны они друг с другом или нет. Например, бородатые черви были описаны как новый тип (погонофоры) в середине 20-го века, но молекулярные исследования почти полвека спустя обнаружили, что это группа кольчатых червей , поэтому типы были объединены (бородатые черви теперь являются семейством кольчатых червей ). [10] С другой стороны, высокопаразитический тип Mesozoa был разделен на два типа ( Orthonectida и Rhombozoa ), когда было обнаружено, что Orthonectida, вероятно, являются вторичноротыми , а Rhombozoa — протостомами . [11]
Эта изменчивость типов побудила некоторых биологов призвать отказаться от концепции типа в пользу распределения таксонов по кладам без какого-либо формального ранжирования размера группы. [9]
Определение на основе плана кузова
[ редактировать ]Определение типа, основанное на строении тела, было предложено палеонтологами Грэмом Баддом и Сёреном Йенсеном (как это сделал Геккель столетием ранее). Это определение было дано потому, что вымершие организмы труднее всего классифицировать: они могут быть ответвлениями, которые отошли от линии типа до того, как были приобретены все признаки, определяющие современный тип. По определению Бадда и Дженсена, тип определяется набором признаков, общих для всех его живых представителей.
Этот подход приносит некоторые небольшие проблемы — например, некоторые из членов типа могли утратить наследственные признаки, общие для большинства членов типа. Кроме того, это определение основано на произвольном моменте времени: настоящем. Однако, поскольку он основан на характере, его легко применить к летописи окаменелостей. Более серьезная проблема заключается в том, что это зависит от субъективного решения о том, какие группы организмов следует считать типами.
Этот подход полезен, поскольку позволяет легко классифицировать вымершие организмы как « стволовые группы » по типам, с которыми они имеют наибольшее сходство, основываясь только на таксономически важных сходствах. [9] Однако доказать, что ископаемое принадлежит кроновой группе типа, сложно, поскольку оно должно иметь характер, уникальный для подмножества кроновой группы. [9] Более того, организмы в стволовой группе типа могут обладать «планом тела» типа без всех характеристик, необходимых для его соответствия. Это ослабляет идею о том, что каждый тип представляет собой отдельный план тела. [12]
На классификацию, использующую это определение, может сильно повлиять случайное выживание редких групп, что может сделать тип гораздо более разнообразным, чем он был бы в противном случае. [13]
Известные типы
[ редактировать ]Животные
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( февраль 2013 г. ) |
Общие цифры являются приблизительными; цифры разных авторов сильно различаются, не в последнюю очередь потому, что некоторые из них основаны на описанных видах, [14] некоторые об экстраполяции на количество неописанных видов. Например, описано около 25 000–27 000 видов нематод, тогда как опубликованные оценки общего числа видов нематод включают 10 000–20 000; 500 000; 10 миллионов; и 100 миллионов. [15]
| Протостом | билатерия | Нефрозоа | |
| вторичноротый | |||
| Базальный/спорный | Не-билатерия | ||
| Вендобионта | |||
| Паразоа | |||
| Другие | |||
| Тип | Значение | Общее имя | Отличительная характеристика | Таксоны описаны |
|---|---|---|---|---|
| Аннелиды | Маленькое кольцо [16] : 306 | Сегментированные черви | Несколько круглых сегментов | 22 000 + сохранившиеся |
| бодрствующий | Фрагментированный | Агматы | Известковые конические раковины | 5 видов, вымершие |
| Археоциата | Древние чашки | Археоциатиды | Вымерший таксон губчатых организмов, строивших рифы, обитавший в теплых тропических и субтропических водах в раннем кембрии. | 3 известных класса (Вымершие) |
| Членистоногие | Суставная стопа | Членистоногие | Сегментированные тела и суставчатые конечности с хитиновым экзоскелетом . | 1 250 000 + сохранившиеся; [14] 20 000+ вымерших |
| Брахиоподы | Рука нога [16] : 336 | Лампы [16] : 336 | Лофофор и ножка | 300–500 сохранились; 12000+ вымерших |
| Мшанки ( Ectoprocta ) | Моховые животные | Моховые животные, морские коврики, эктопрокты. [16] : 332 | Лофофор, ножка отсутствует, с ресничками щупальца , анус снаружи кольца ресничек. | 6000 сохранившихся [14] |
| Четогната | Длинношерстная челюсть | Стрелочные черви [16] : 342 | Хитиновые шипы по обе стороны головы, плавники. | ок. 100 сохранившихся |
| Хордовые | Со шнуром | Хордовые | Полый спинной нервный тяж , хорда , глоточные щели , эндостиль , постанальный хвост . | ок. 55 000 + [14] |
| Книдарийцы | Крапива жгучая | Книдарийцы | Нематоцисты (стрекательные клетки) | ок. 16 000 [14] |
| Гребневик | Гребенчатый носитель | Гребенчатое желе [16] : 256 | Восемь «гребенчатых рядов» сросшихся ресничек. | ок. 100–150 сохранились |
| Циклиофора | Перевозка колес | Симбион | Круглый рот, окруженный маленькими ресничками, мешкообразными телами. | 3 + |
| Иглокожие | Колючая кожа | Иглокожие [16] : 348 | Пятикратная радиальная симметрия у живых форм, мезодермальные кальцинированные шипики. | ок. 7500 сохранились; [14] ок. 13 000 вымерших |
| Энтопрокта | Внутри ануса [16] : 292 | Бокалообразные черви | Анус внутри кольца ресничек | ок. 150 |
| Гастротрича | Волосатый живот [16] : 288 | Желудочные черви | Две концевые клейкие трубки | ок. 690 |
| Гнатостомулида | Челюстное отверстие | Челюстные черви [16] : 260 | Крошечные черви, относящиеся к коловраткам, не имеющие полости тела. | ок. 100 |
| полухордовые | Половина шнура [16] : 344 | Желудевые черви, полухордовые | Стомохорд в воротнике, глоточные щели | ок. 130 сохранившихся |
| Кинорхинча | Движение морды | Грязевые драконы | Одиннадцать сегментов, каждый со спинной пластинкой. | ок. 150 |
| лорицифера | Броненосец | Насадки щетки | Зонтикообразные чешуйки на каждом конце. | ок. 122 |
| Микрогнатозоа | Животные с крошечными челюстями | Никто | в форме гармошки Растяжимая грудная клетка | 1 |
| Моллюска | Мягкий [16] : 320 | Моллюски / моллюски | Мускулистая нога и мантия, круглая раковина. | 85 000 + сохранившиеся; [14] 80 000+ вымерших [17] |
| Монобластозоа ( Имя для поиска ) | Никто | отчетливые передняя и задняя части и густо покрыты ресничками, особенно вокруг «рта» и «ануса». | 1 | |
| Нематоды | Нравится тема | Круглые черви, нитчатые черви [16] : 274 | Круглое сечение, кератиновая . кутикула | 25,000 [14] |
| Нематоморфа | Форма резьбы [16] : 276 | Черви конского волоса, гордиевые черви [16] : 276 | Длинные и тонкие паразитические черви, тесно связанные с нематодами. | ок. 320 |
| Немертеа | Морская нимфа [16] : 270 | Ленточные черви, ринхоцела [16] : 270 | Несегментированные черви с хоботком, расположенным в полости, происходящей от целома, называемой ринхоцелем. | ок. 1200 |
| Онихофора | Носитель когтей | Бархатные черви [16] : 328 | Червеобразное животное с хитиновыми когтями на ногах. | ок. 200 сохранившихся |
| Ортонектида | Ортонектид | Паразитические, микроскопические, простые, червеобразные организмы. | 20 | |
| Петалонамяэ | В форме листьев | Никто | Вымерший тип эдиакарского региона. Они обитают на дне и неподвижны, имеют форму листьев (фрондоморфы), перьев или веретена. | 3 класса, вымершие |
| Форониды | любовница Зевса | Подковообразные черви | U-образная кишка | 11 |
| Плакозоа | Тарелочные животные | Трихоплаксы [16] : 242 | Дифференцированные верхняя и нижняя поверхности, два слоя реснитчатых клеток, клетки амебоидных волокон между ними. | 4 + |
| Платихельминты | Плоский червь [16] : 262 | Плоские черви [16] : 262 | Уплощенные черви без полости тела. Многие из них являются паразитами. | ок. 29 500 [14] |
| Порифера | носитель пор | Губки [16] : 246 | Перфорированная внутренняя стенка, самое простое из всех известных животных. | 10 800 сохранившихся [14] |
| Приапулида | Маленький Приап | Черви пениса | Черви в форме пениса | ок. 20 |
| Выраженный | Прежде чем артикулировать | Проартикулирует | Вымершая группа матрасоподобных организмов, обладающих «симметрией скольжения». Найден во времена Эдиакария. | 3 класса, вымершие |
| Ромбозоа ( Дицемида ) | Леденцы животное | Ромбозои [16] : 264 | Одиночные переднезадние осевые клеточные эндопаразиты, окруженные реснитчатыми клетками. | 100 + |
| коловратка | Колесный подшипник | Коловушки [16] : 282 | Передняя коронка ресничек | ок. 3500 [14] |
| Саккоритида | Саккус: «карман» и «морщина». | Никто | Saccorhytus имеет размер всего около 1 мм (1,3 мм) и характеризуется сферическим или полусферическим телом с выступающим ртом. Его тело покрыто толстой, но гибкой кутикулой. Над ртом имеется узелок. Вокруг его тела расположены 8 отверстий в виде усеченного конуса с радиальными складками. Считается вторичноротым. [18] или ранний экдизозой . [19] | 2 вида, вымершие |
| тихоходка | Медленный шаг | Водяные медведи, моховые поросята | Микроскопические родственники членистоногих с четырехсегментным телом и головой. | 1,000 |
| Трилобоза | Трехлопастное животное | Трилобозой | Таксон преимущественно дискоидных организмов, обладающих трицентрической симметрией. Все эдиакарского возраста. | 18 родов, вымершие |
| Старая школа | Древний житель | Старик | Возможно, это подтип хордовых. Их тело состоит из двух частей: большой передней части, покрытой большим «ртом» и сотней круглых предметов с каждой стороны, которые интерпретируются как жабры или отверстия возле глотки. Их задняя глотка состоит из 7 сегментов. | 15 видов, вымершие |
| Ксенацеломорфа | Странная полая форма | Подтип Acoelomorpha и ксенотурбеллиды | Маленькие, простые животные. Билатерий , но без типичных билатеральных структур, таких как полости кишечника, анусы и системы кровообращения. [20] | 400 + |
| Всего: 40 | 1,525,000 [14] |
Растения
[ редактировать ]Царство Plantae определяется разными биологами по-разному (см. Современные определения Plantae ). Все определения включают живые эмбриофиты (наземные растения), к которым можно добавить два отдела зеленых водорослей, Chlorophyta и Charophyta , образуя кладу Viridiplantae . Таблица ниже соответствует влиятельной (хотя и спорной) системе Кавальера-Смита, приравнивающей «Plantae» к Archaeplastida . [21] группа, содержащая Viridiplantae и подразделения водорослей Rhodophyta и Glaucophyta .
Определение и классификация растений на уровне подразделения также варьируются от источника к источнику и постепенно менялись в последние годы. Так, некоторые источники относят хвощи к отделу Arthrophyta, а папоротники - к отделу Monilophyta. [22] в то время как другие помещают их обоих в Monilophyta, как показано ниже. Отдел Pinophyta можно использовать для всех голосеменных растений (т.е. включая саговники, гинкго и гнетофиты). [23] или только для хвойных пород, как показано ниже.
С момента первой публикации системы APG в 1998 году, в которой предлагалась классификация покрытосеменных растений до уровня порядков , многие источники предпочитали рассматривать ранги выше порядков как неформальные клады. Там, где были предусмотрены формальные ранги, традиционные подразделения, перечисленные ниже, были сокращены до гораздо более низкого уровня, например, подклассов . [24]
| Наземные растения | Зеленые растения | |
| Зеленые водоросли | ||
| Другие водоросли ( Biliphyta ) [21] | ||
| Разделение | Значение | Общее имя | Отличительные характеристики | Описанные виды |
|---|---|---|---|---|
| Антоцеротофиты [25] | Антоцероподобные растения | Роголистники | роговидные Спорофиты , сосудистой системы нет. | 100 -300+ |
| Мохообразные [25] | Брюмоподобные растения, моховые растения | мхи | Стойкие неразветвленные спорофиты , нет сосудистой системы. | ок. 12 000 |
| Харофита | Чараподобные растения | Харофиты | ок. 1000 | |
| Хлорофита | (Желто-)зеленые растения [16] : 200 | Хлорофиты | ок. 7000 | |
| Цикадофиты [26] | Цикасоподобные растения, пальмоподобные растения | Саговники | Семена, крона сложных листьев | ок. 100 -200 |
| Гинкгофита [27] | Гинкгоподобные растения | Гинкго, девичье дерево | Семена, не защищенные плодами (единичные живые виды) | только 1 сохранившийся; 50+ вымерших |
| Глаукофита | Сине-зеленые растения | Глаукофиты | 15 | |
| Гнетофиты [28] | Гнетумоподобные растения | Гнетофиты | Семена и деревянистая сосудистая система с сосудами. | ок. 70 |
| Ликоподиофиты , [23] | Lycopodium -подобные растения Волчьи растения | Клубухи и колючие мхи | Микрофилл листьев , сосудистая система | 1290 сохранившихся |
| Магнолиофита | Магнолиеподобные растения | Цветковые растения, покрытосеменные | Цветы и плоды, сосудистая система с сосудами | 300,000 |
| Марчантиофита , [30] Гепатофиты [25] | Маршанциеподобные растения Печеночные растения | Печеночники | Эфемерные неразветвленные спорофиты , без сосудистой системы. | ок. 9000 |
| Полиподиофиты , | полиподии Растения, похожие на | Папоротники | мегафилла Листья , сосудистая система | ок. 10 560 |
| Пикозоа | Чрезвычайно маленькие животные | Пикозои, пикобилифиты, билифиты | 1 | |
| Пинофита , [23] Хвойные [31] | Сосновые растения Шишконосное растение | Хвойные деревья | Шишки, содержащие семена и древесину, состоящую из трахеид. | 629 сохранившихся |
| Празинодермофита | Празинодермоподобные растения | Пикозои, пикобилифиты, билифиты | 8 | |
| Родофита | Розовые растения | Красные водоросли | Используйте фикобилипротеины в качестве вспомогательных пигментов . | ок. 7000 |
| Всего: 14 |
Грибы
[ редактировать ]| Разделение | Значение | Общее имя | Отличительные характеристики | Описанные виды |
|---|---|---|---|---|
| Аскомикота | Грибок мочевого пузыря [16] : 396 | Аскомицеты, [16] : 396 мешок грибов | Имеют тенденцию иметь плодовые тела (аскокарпий). [32] Нитчатые, образующие гифы, разделенные перегородками. Могут размножаться бесполым путем. [33] | 30,000 |
| Базидиомикота | Маленький базовый гриб [16] : 402 | Базидиомицеты, [16] : 402 клубные грибы | Крошечные грибы, поганки, головня и ржавчина. Половое размножение. [34] | 31,515 |
| Бластокладиомикота | Ответвительный ветвящийся гриб [35] | Бластоклады | Менее 200 | |
| Хитридиомикота | Маленький грибок в кастрюле [36] | Хитриды | Преимущественно водные сапротрофы или паразиты. Имеют задний жгутик . Обычно одноклеточные, но могут быть и многоклеточными. [37] [38] [39] | 1000+ |
| Гломеромикота | Клубок грибка из пряжи [16] : 394 | Гломеромицеты, АМ- грибы [16] : 394 | В основном присутствуют арбускулярные микоризы, наземные, с небольшим присутствием на водно-болотных угодьях. Размножение бесполое, но требует корней растений. [34] | 284 |
| Микроспоридии | Мелкие семена [40] | Микроспораны [16] : 390 | 1400 | |
| Неокаллимастигомикота | Новый красивый плетеный гриб [41] | Неокаллимастигомицеты | Преимущественно локализуется в пищеварительном тракте травоядных животных. Анаэробные, наземные и водные. [42] | ок. 20 [43] |
| Зигомикота | Парный гриб [16] : 392 | Зигомицеты [16] : 392 | Большинство из них являются сапробами и размножаются половым и бесполым путем. [42] | ок. 1060 |
| Всего: 8 |
Тип Microsporidia обычно включается в царство Грибов, хотя его точное родство остается неопределенным. [44] считает его простейшим. и Международное общество протистологов [45] (см. Протиста ниже). Молекулярный анализ Zygomycota показал, что он полифилетичен (его представители не имеют общего предка), [46] что многие биологи считают нежелательным. Соответственно, есть предложение об упразднении типа Zygomycota. Его члены будут разделены на тип Glomeromycota и четыре новых подтипа incertae sedis (неопределенного местоположения): Entomophthoromycotina , Kickxellomycotina , Mucoromycotina и Zoopagomycotina . [44]
Протиста
[ редактировать ]Королевство Protista (или Protoctista) включено в традиционную модель пяти или шести царств, где его можно определить как содержащее всех эукариотов , которые не являются растениями, животными или грибами. [16] : 120 Protista — парафилетический таксон. [47] что менее приемлемо для современных биологов, чем в прошлом. Были сделаны предложения разделить его между несколькими новыми королевствами, такими как Простейшие и Хромиста в системе Кавалера-Смита . [48]
Систематика протистов долгое время была нестабильной. [49] с различными подходами и определениями, что приводит к появлению множества конкурирующих схем классификации. Многие из перечисленных ниже типов используются в « Каталоге жизни» . [50] и соответствуют схеме Protozoa-Chromista, [45] с обновлениями из последней (2022 г.) публикации Cavalier-Smith . [51] Другие типы обычно используются другими авторами и адаптированы из системы, используемой Международным обществом протистологов (ISP). Некоторые описания основаны на версии эукариот, опубликованной ISP в 2019 году. [52]
| Страменопилес | « Хромиста » | |
| Альвеолата | ||
| Ризария | ||
| « Хакробия » | ||
| « Саркомастигота ». | « Простейшие » | |
| " Экскавата " | ||
| Сиротские группы | ||
| Тип | Значение | Общее имя | Отличительные характеристики | Описанные виды | Изображение |
|---|---|---|---|---|---|
| амебозоа | Аморфные животные | Амебозои | Наличие псевдоподий для амебоидного движения, трубчатых крист . [52] | ок. 2400 [53] |  |
| Апикомплекса | Апикальные складки [54] | Апикомплексаны, споровики | Преимущественно паразитический, по крайней мере, одна стадия жизненного цикла с уплощенными субпелликулярными пузырьками и полным апикальным комплексом, нефотосинтезирующим апикопластом . [52] | более 6000 [54] |  |
| Когда он женится ( парафилетический ) | Апусомонас -подобные животные | Скользящие биреснитчатые клетки с двумя или тремя коннекторами между центриолями. | 32 |  | |
| Бигира | Два кольца | Страменопилы с двойной спиралью в цилиарной переходной зоне. |  | ||
| Церкозоа | Жгутиковое животное | Церкозоа | Определяется молекулярной филогенией , не имеет отличительных морфологических или поведенческих признаков. [52] |  | |
| Хромрида | Хромероподобные организмы | Хромподеллиды, хромриды, колподеллиды [55] | Двужгутики, хлоропласты с четырьмя мембранами, неполный апикальный комплекс, кортикальные альвеолы, трубчатые кристы. [52] | 8 [56] |  |
| Чоанозоа ( парафилетический ) | Воронкообразные животные [16] | Опистоконтные протисты | Филозные ложноножки ; некоторые с воротником из микроворсинок, окружающих жгутик | ок. 300 [53] |  |
| Цилиофора | Носители ресничек | Инфузории | Наличие множественных ресничек и цитостома . | ок. 4500 [57] |  |
| Криптиста | Скрытый [16] | Определяется молекулярной филогенией , плоские кристы. [52] | 246 [56] [52] |  | |
| Динофлагелляты | Кружащиеся жгутиконосцы [16] | Динофлагелляты | Двужгутики с поперечным лентообразным жгутиком с множеством волн, отходящих слева от клетки, и продольным жгутиком, бьющимся назад только с одной или несколькими волнами. [52] | 2957 сохранившихся 955 ископаемых [56] |  |
| Эндомикса | Внутри слизи [16] [58] | Определяется молекулярной филогенией , [52] обычно плазмодии эндопаразиты других эукариот. [58] |  | ||
| Эолука ( парафилетический ) | Ранний ритм [59] | Гетеротрофные двужгутики с вентральной питающей бороздкой. [59] | 23 |  | |
| эвгленозоа | Настоящие глазные животные | Двужгутики, одна из двух ресничек , вставленная в апикальный или субапикальный карман, уникальная ресничная конфигурация. [52] | 2037 сохранившихся 20 ископаемых [56] |  | |
| Охрофита , Гетероконтофиты | Растения охры , гетероконтные растения | Гетероконтовые водоросли, страменохромы, охрофиты, гетероконтофиты. | Двужгутиковые с трехраздельными мастигонемами, хлоропластами с четырьмя мембранами и хлорофиллами а и с , трубчатыми кристами. [52] | 21 052 сохранившихся 2262 ископаемых [56] |  |
| Хаптиста | Закрепить [16] | Тонкие придатки на основе микротрубочек для питания (гаптонема у гаптофитов , аксоподии у центрохелид ), сложные минерализованные чешуйки. [52] | 517 сохранившихся 1205 ископаемых [56] |  | |
| Гемимастигофора | Неполные или атипичные жгутиковые. [60] | Гемимастиготы [61] | Эллипсоидные или червеобразные фаготрофы, два слегка спиральных ряда примерно по 12 ресничек в каждом, текальные пластинки под мембраной, поддерживаемые микротрубочками, и ротационно-симметричные трубчатые и мешотчатые кристы. [52] [60] | 10 [62] |  |
| Малавимонада | Malawimonas -подобные организмы | Малавимонады | Мелкие свободноживущие бицилаты с двумя кинетосомами, одной или двумя лопастями в задней ресничке. | 3 [63] |  |
| Метамонада | Средние монады | Метамонады | Анаэробные или микроаэрофильные , некоторые без митохондрий ; четыре кинетосомы на кинетид |  | |
| Опистоспоридии (часто считается грибами ) | опистоконта Споры [64] | Паразиты с хитиновыми спорами и экструзивным аппаратом инвазии в хозяина |  | ||
| перколозоа | Перколомонадоподобные животные | Сложный жизненный цикл , содержащий амебы, жгутиконосцы и цисты . [52] | |||
| Перкинсозоа | Перкинсусоподобные животные | Перкинзозои, перкинсиды | Паразитические двужгутики, неполный апикальный комплекс, образование зооспорангиев или недифференцированных клеток через гифоподобную трубку. [52] | 26 |  |
| Provora | Пожирающие прожорливых протистов [65] | Определенные методом молекулярной филогении свободноживущие эукариофаги-гетеротрофные двужгутики с вентральной бороздкой и экструсомами. [65] | 7 [65] |  | |
| Псевдогрибы | Ложный гриб | Определяется молекулярной филогенией , фаготрофные гетероконты со спирально-ресничной переходной зоной. [66] | более 1200 [67] |  | |
| Ретария | Организмы, несущие ретикулоподии [58] | Питание ретикулоподий (или аксоподий ) обычно осуществляется посредством различных типов скелета, закрытого митоза. [68] | 10 000 сохранившихся 50 000 ископаемых |  | |
| Сулькозоа ( парафилетический ) | Животные с бороздками [59] | Аэробные жгутиконосцы (нет, 1, 2 или 4 жгутика) с дорсальной полужесткой пелликулой из одного или двух подмембранных плотных слоев, вентральной питающей бороздкой, ветвящимися вентральными псевдоподиями, обычно нитевидными. [59] | 40 + |  | |
| Телонемия | Телонемоподобные организмы [69] | Телонемиды [70] | Фаготрофные грушевидные двужгутики с уникальным сложным цитоскелетом, трубчатыми кристами, трехраздельными мастигонемами, корковыми альвеолами. [69] [70] | 7 |  |
| Итого: 26 , но см. ниже. | |||||
Число типов протистов сильно варьируется от одной классификации к другой. В Каталог жизни входят Rhodophyta и Glaucophyta царства Plantae. [50] но другие системы считают эти типы частью Protista. [71] Кроме того, менее популярные схемы классификации объединяют Ochrophyta и Pseudofungi в один тип Gyrista , а все альвеоляты, кроме инфузорий , в один тип Myzozoa , позднее пониженный в ранге и включенный в парафилетический тип Miozoa . [51] Даже внутри типа появляются другие ранги на уровне типа, например, в случае Bacillariophyta (диатомовые водоросли) внутри Ochrophyta . Эти различия стали неактуальными после принятия кладистического ISP подхода, при котором таксономические ранги исключаются из классификаций после того, как считаются излишними и нестабильными. Многие авторы предпочитают такое использование, что приводит к устареванию схемы Chromista-Protozoa. [52]
Бактерии
[ редактировать ]В настоящее время существует 40 типов бактерий (не включая « цианобактерии »), которые были официально опубликованы в соответствии с Бактериологическим кодексом. [72]
- Абдитибактериота
- Ацидобактерии , фенотипически разнообразные и в основном некультивируемые.
- Actinomycetota , грамположительные виды с высоким содержанием G+C
- Аквификота , глубоковетвистая.
- Арматимонадота
- Атрибактериота
- Bacillota , грамположительные виды с низким содержанием G+C, такие как спорообразующие бациллы (аэробные) и клостридии (анаэробные).
- Бактероидота
- Бальнеолота
- Бделловибрионота
- Caldisericota , бывший кандидат в дивизион OP5, изгнанник Caldisericum является единственным представителем
- Калдитрихота
- Кампилобактерия
- Хламидиота
- Хлоробиота , зеленые серобактерии.
- Хлорофлексота , зеленые бессерные бактерии.
- Chrysiogenota , всего 3 рода ( Chrysiogenes arsenatis , Desulfurispira natronophila , Desulfurispirillum щелочефилум )
- Копротермобактерия
- Деферрибактериота
- Deinococta , Deinococcus radiodurans и Thermus aquaticus являются «общеизвестными» видами этого типа.
- Диктиогломота
- Elusimicrobiota , бывший кандидат в подразделение Thermite Group 1
- Фибробактерита
- Фузобактериота
- Одаренный драгоценными камнями
- Игнавибактериота
- Киритиматиеллота
- Lentisphaerota , ранее клада VadinBE97
- Mycoplasmatota , известный род: Mycoplasma.
- Миксококкота
- Нитроспинота
- Нитроспирота
- Планктомицетота
- Pseudomonadota , наиболее известный тип, содержащий такие виды, как Escherichia coli или Pseudomonas aeruginosa.
- Родотермота
- Spirochaetota , виды включают Borrelia burgdorferi , вызывающую болезнь Лайма.
- синергист
- Термодесульфобактерии
- Термомикробиота
- Термотогота , глубоковетвящаяся.
- Веррукомикробиота
Архея
[ редактировать ]В настоящее время существует 2 типа, которые были официально опубликованы в соответствии с Бактериологическим кодексом. [72]
- Нитрососферота
- Thermoproteota , второй по распространенности тип архей.
Другие типы, которые были предложены, но не названы правильно, включают:
- « Euryarchaeota », наиболее распространенный тип архей.
- « Корархеота »
- « Наноархеоты », сверхмелкие симбиоты, единичный известный вид.
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ "Но такое реальное и вполне полное единство есть сумма всех видов, постепенно развившихся из одной и той же общей родительской формы, как, например, все позвоночные. Эту сумму мы называем трибой (филоном)".
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Макнил, Дж.; и др., ред. (2012). Международный кодекс номенклатуры водорослей, грибов и растений (Мельбурнский кодекс), принятый Восемнадцатым Международным ботаническим конгрессом в Мельбурне, Австралия, июль 2011 г. (электронное издание). Международная ассоциация систематики растений. Архивировано из оригинала 10 октября 2020 года . Проверено 14 мая 2017 г.
- ^ «Науки о жизни» . Новый словарь культурной грамотности американского наследия (третье изд.). Компания Хоутон Миффлин. 2005 . Проверено 4 октября 2008 г.
Типы в царстве растений часто называют подразделениями.
- ^ Берг, Линда Р. (2 марта 2007 г.). Вводная ботаника: растения, люди и окружающая среда (2-е изд.). Cengage Обучение. п. 15. ISBN 9780534466695 . Проверено 23 июля 2012 г.
- ^ Валентин 2004 , с. 8.
- ^ Геккель, Эрнст (1866). морфология организмов ( Общая на немецком языке). Том 1. Берлин, (Германия): Г. Раймер. стр. 28–29 .
- ^ Наик, В.Н. (1984). Систематика покрытосеменных . Тата МакГроу-Хилл. п. 27. ISBN 9780074517888 .
- ^ Коллинз А.Г., Валентайн Дж.В. (2001). «Определение типа: эволюционные пути к строению тела многоклеточных животных» . Эволюция и развитие . 3 : 432–442. Архивировано из оригинала (PDF) 27 апреля 2020 года . Проверено 5 марта 2013 г.
- ^ Валентин, Джеймс В. (2004). О происхождении Фила . Чикаго: Издательство Чикагского университета. п. 7. ISBN 978-0-226-84548-7 .
Классификации организмов в иерархических системах использовались в семнадцатом и восемнадцатом веках. Обычно организмы группировались в соответствии с их морфологическим сходством, как это воспринималось теми ранними исследователями, а затем эти группы группировались в соответствии с их сходством и так далее, чтобы сформировать иерархию.
- ^ Перейти обратно: а б с д и Бадд, GE; Дженсен, С. (май 2000 г.). «Критическая переоценка летописи окаменелостей двусторонних типов» . Биологические обзоры . 75 (2): 253–295. дои : 10.1111/j.1469-185X.1999.tb00046.x . ПМИД 10881389 . S2CID 39772232 . Архивировано из оригинала 15 сентября 2019 года . Проверено 26 мая 2007 г.
- ^ Роуз, GW (2001). «Кладистический анализ Siboglinidae Caullery, 1914 (Polychaeta, Annelida): ранее относились к типам Pogonophora и Vestimentifera» . Зоологический журнал Линнеевского общества . 132 (1): 55–80. дои : 10.1006/zjls.2000.0263 .
- ^ Павловски Дж., Монтойя-Бургос Дж.И., Фарни Дж.Ф., Вюст Дж., Занинетти Л. (октябрь 1996 г.). «Происхождение Mesozoa установлено на основе последовательностей гена 18S рРНК» . Молекулярная биология и эволюция . 13 (8): 1128–32. doi : 10.1093/oxfordjournals.molbev.a025675 . ПМИД 8865666 .
- ^ Бадд, GE (сентябрь 1998 г.). «Эволюция строения тела членистоногих в кембрии на примере мышц аномалокаридид». Летайя . 31 (3): 197–210. дои : 10.1111/j.1502-3931.1998.tb00508.x .
- ^ Бриггс, Германия ; Форти, РА (2005). «Чудесная борьба: систематика, стволовые группы и филогенетический сигнал кембрийской радиации». Палеобиология . 31 (2 (Приложение)): 94–112. doi : 10.1666/0094-8373(2005)031[0094:WSSSGA]2.0.CO;2 . S2CID 44066226 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Чжан, Чжи-Цян (30 августа 2013 г.). «Биоразнообразие животных: обновление классификации и разнообразия в 2013 году. В: Чжан, З.-К. (ред.) Биоразнообразие животных: план классификации более высокого уровня и исследование таксономического богатства (Дополнения, 2013 г.)» . Зоотакса . 3703 (1): 5. дои : 10.11646/zootaxa.3703.1.3 .
- ^ Фелдер, Дэррил Л.; Кэмп, Дэвид К. (2009). Мексиканский залив Происхождение, воды и биота: биоразнообразие . Издательство Техасского университета A&M. п. 1111. ИСБН 978-1-60344-269-5 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а Маргулис, Линн ; Чепмен, Майкл Дж. (2009). Королевства и владения: Иллюстрированный путеводитель по типам жизни на Земле (4-е исправленное изд.). Лондон: Академическая пресса. ISBN 9780123736215 .
- ^ Фельдкамп, С. (2002) Современная биология . Холт, Райнхарт и Уинстон, США. (стр. 725)
- ^ Хан, Цзянь; Моррис, Саймон Конвей; Оу, Цян; Шу, Деган; Хуан, Хай (2017). «Мейофауна вторичноротых из базального кембрия Шэньси (Китай)» . Природа . 542 (7640): 228–231. Бибкод : 2017Natur.542..228H . дои : 10.1038/nature21072 . ISSN 1476-4687 . ПМИД 28135722 . S2CID 353780 .
- ^ Лю, Юньхуань; Карлайл, Эмили; Чжан, Хуацяо; Ян, Бен; Штайнер, Майкл; Шао, Тецюань; Дуань, Байчуань; Мароне, Федерика; Сяо, Шухай; Донохью, Филип CJ (17 августа 2022 г.). «Saccorhytus — ранний экдисозой, а не самый ранний вторичноротый» . Природа . 609 (7927): 541–546. Бибкод : 2022Natur.609..541L . дои : 10.1038/s41586-022-05107-z . hdl : 1983/454e7bec-4cd4-4121-933e-abeab69e96c1 . ISSN 1476-4687 . ПМИД 35978194 . S2CID 251646316 .
- ^ Кэннон, Джей Ти; Веллутини, Британская Колумбия; Смит, Дж.; Ронквист, Ф.; Йонделиус, У.; Хейнол, А. (4 февраля 2016 г.). «Xenacoelomorpha — сестринская группа Nephrozoa» . Природа . 530 (7588): 89–93. Бибкод : 2016Natur.530...89C . дои : 10.1038/nature16520 . ПМИД 26842059 . S2CID 205247296 .
- ^ Перейти обратно: а б Кавальер-Смит, Томас (22 июня 2004 г.). «Только шесть королевств жизни» . Труды: Биологические науки . 271 (1545): 1251–1262. дои : 10.1098/rspb.2004.2705 . ПМК 1691724 . ПМИД 15306349 .
- ^ Маусет 2012 , стр. 514, 517.
- ^ Перейти обратно: а б с Кронквист, А.; А. Тахтаджан; В. Циммерманн (апрель 1966 г.). «О высших таксонах Embryobionta». Таксон . 15 (4): 129–134. дои : 10.2307/1217531 . JSTOR 1217531 .
- ^ Чейз, Марк В. и Ревил, Джеймс Л. (октябрь 2009 г.), «Филогенетическая классификация наземных растений, сопровождающих APG III», Ботанический журнал Линнеевского общества , 161 (2): 122–127, doi : 10.1111/ j.1095-8339.2009.01002.x
- ^ Перейти обратно: а б с Маусет, Джеймс Д. (2012). Ботаника: Введение в биологию растений (5-е изд.). Садбери, Массачусетс: Джонс и Бартлетт Обучение. ISBN 978-1-4496-6580-7 . п. 489
- ^ Маусет 2012 , стр. 540.
- ^ Маусет 2012 , стр. 542.
- ^ Маусет 2012 , стр. 543.
- ^ Маусет 2012 , стр. 509.
- ^ Крэндалл-Стотлер, Барбара; Стотлер, Раймонд Э. (2000). «Морфология и классификация Marchantiophyta». В А. Джонатане Шоу; Бернар Гоффине (ред.). Биология мохообразных . Кембридж: Издательство Кембриджского университета. п. 21. ISBN 978-0-521-66097-6 .
- ^ Маусет 2012 , стр. 535.
- ^ Вятт, Т.; Вестен, Х.; Дейкстерхейс, Дж. (2013). «Достижения прикладной микробиологии. Глава 2. Споры грибов для распространения в пространстве и времени». Достижения прикладной микробиологии . 85 : 43–91. дои : 10.1016/B978-0-12-407672-3.00002-2 . ПМИД 23942148 .
- ^ «Классификации грибов | Безграничная биология» . Courses.lumenlearning.com . Проверено 5 мая 2019 г.
- ^ Перейти обратно: а б «Архейная генетика | Безграничная микробиология» . Courses.lumenlearning.com .
- ^ Холт, Джек Р.; Иудика, Карлос А. (1 октября 2016 г.). «Бластокладиомикота» . Разнообразие жизни . Университет Саскуэханны . Проверено 29 декабря 2016 г.
- ^ Холт, Джек Р.; Иудика, Карлос А. (9 января 2014 г.). «Хитридиомикота» . Разнообразие жизни . Университет Саскуэханны . Проверено 29 декабря 2016 г.
- ^ «Chytridiomycota | тип грибов» . Британская энциклопедия . Проверено 5 мая 2019 г.
- ^ МакКоннахи, М (2014). Физико-химические свойства грибов . дои : 10.1016/B978-0-12-801238-3.05231-4 . ISBN 9780128012383 .
- ^ Тейлор, Томас; Крингс, Майкл; Тейлор, Эдит (2015). «Ископаемые грибы, глава 4 — Chytridiomycota». Ископаемые грибы : 41–67. дои : 10.1016/b978-0-12-387731-4.00004-9 .
- ^ Холт, Джек Р.; Иудика, Карлос А. (12 марта 2013 г.). «Микроспоридии» . Разнообразие жизни . Университет Саскуэханны . Проверено 29 декабря 2016 г.
- ^ Холт, Джек Р.; Иудика, Карлос А. (23 апреля 2013 г.). «Неокаллимастигомикота» . Разнообразие жизни . Университет Саскуэханны . Проверено 29 декабря 2016 г.
- ^ Перейти обратно: а б «Виды грибов» . BiologyWise . 22 мая 2009 года . Проверено 5 мая 2019 г.
- ^ Ван, Сюэвэй; Лю, Синчжун; Гроеневальд, Йоханнес З. (2017). «Филогения анаэробных грибов (тип Neocallimastigomycota) с участием яка из Китая» . Антони ван Левенгук . 110 (1): 87–103. дои : 10.1007/s10482-016-0779-1 . ПМК 5222902 . ПМИД 27734254 .
- ^ Перейти обратно: а б Хиббетт Д.С., Биндер М., Бишофф Дж.Ф., Блэквелл М., Кэннон П.Ф., Эрикссон О.Е. и др. (май 2007 г.). «Филогенетическая классификация грибов более высокого уровня» (PDF) . Микологические исследования . 111 (Часть 5): 509–47. CiteSeerX 10.1.1.626.9582 . дои : 10.1016/j.mycres.2007.03.004 . ПМИД 17572334 . S2CID 4686378 . Архивировано из оригинала (PDF) 26 марта 2009 года.
- ^ Перейти обратно: а б Руджеро, Майкл А.; Гордон, Деннис П.; Оррелл, Томас М.; и др. (29 апреля 2015 г.). «Классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (6): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ Уайт, Мерлин М.; Джеймс, Тимоти Ю.; О'Доннелл, Керри; и др. (ноябрь – декабрь 2006 г.). «Филогения Zygomycota на основе данных о ядерных рибосомальных последовательностях». Микология . 98 (6): 872–884. дои : 10.1080/15572536.2006.11832617 . ПМИД 17486964 . S2CID 218589354 .
- ^ Хаген, Джоэл Б. (январь 2012 г.). «Пять королевств, более или менее: Роберт Уиттакер и широкая классификация организмов» . Бионаука . 62 (1): 67–74. дои : 10.1525/bio.2012.62.1.11 .
- ^ Блэквелл, Уилл Х.; Пауэлл, Марта Дж. (июнь 1999 г.). «Примирение королевств с номенклатурными кодексами: необходимо ли это?» . Систематическая биология . 48 (2): 406–412. дои : 10.1080/106351599260382 . ПМИД 12066717 .
- ^ Дэвис, РА (19 марта 2012 г.). «Королевство ПРОТИСТА» . Колледж Маунт-Сент-Джозеф . Проверено 28 декабря 2016 г.
- ^ Перейти обратно: а б «Таксономическое дерево» . Каталог жизни . 23 декабря 2016 г. Архивировано из оригинала 1 августа 2021 г. Проверено 28 декабря 2016 г.
- ^ Перейти обратно: а б Кавалер-Смит Т (2022). «Эволюция ресничной переходной зоны и корень дерева эукариот: значение для происхождения опистоконтов и классификации царств простейших, растений и грибов» . Протоплазма . 259 : 487–593. дои : 10.1007/s00709-021-01665-7 . ПМК 9010356 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л., Хампл В., Хейсс А.А., Хоппенрат М., Джеймс Тай, Карнковска А., Карпов С., Ким Е., Колиско М., Кудрявцев А., Лар DJG, Лара Е., Ле Галл Л., Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл И.А.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Перейти обратно: а б Павловски Дж., Аудик С., Адл С., Басс Д., Белбахри Л., Берни С. и др. (6 ноября 2012 г.). «Рабочая группа CBOL по протистам: штрих-кодирование эукариотического богатства за пределами царств животных, растений и грибов» . ПЛОС Биология . 10 (11): e1001419. дои : 10.1371/journal.pbio.1001419 . ПМК 3491025 . ПМИД 23139639 .
- ^ Перейти обратно: а б Вотыпка Ю, Модри Д, Оборник М, Шлапета Ю, Лукеш Ю (2016). «Апикомплекса». В Арчибальд Дж., Симпсон А.Г.Б., Сламовиц Ч., Маргулис Л., Мелконян М., Чепмен Д.Д., Корлисс Дж.О. (ред.). Справочник протистов . Чам: Спрингер. дои : 10.1007/978-3-319-32669-6_20-1 .
- ^ Ян Янушковец; Денис Тихоненков; Фабьен Бурки; Алексис Т. Хоу; Мартин Колиско; Александр П. Мыльников; Патрик Джон Килинг (25 февраля 2015 г.). «Факторы, опосредующие пластидную зависимость, и причины паразитизма у апикомплексов и их близких родственников» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (33): 10200–10207. Бибкод : 2015PNAS..11210200J . дои : 10.1073/PNAS.1423790112 . ISSN 0027-8424 . ПМЦ 4547307 . ПМИД 25717057 . Викиданные Q30662251 .
- ^ Перейти обратно: а б с д и ж Майкл Д. Гири (21 января 2024 г.). «Сколько существует видов водорослей? Реприза. Четыре царства, 14 типов, 63 класса и все еще растут». Журнал психологии . 00 :1–15. дои : 10.1111/JPY.13431 . ISSN 0022-3646 . ПМИД 38245909 . Викиданные Q124684077 .
- ^ Фойснер, В.; Хоксворт, Дэвид, ред. (2009). Разнообразие и географическое распространение протистов . Темы биоразнообразия и охраны природы. Том. 8. Спрингер Нидерланды. п. 111. дои : 10.1007/978-90-481-2801-3 . ISBN 9789048128006 .
- ^ Перейти обратно: а б с Т. Кавальер-Смит (март 2002 г.). «Фаготрофное происхождение эукариот и филогенетическая классификация простейших». Международный журнал систематической и эволюционной микробиологии . 52 (2): 297–354. дои : 10.1099/00207713-52-2-297 . ISSN 1466-5026 . ПМИД 11931142 . Викиданные Q28212529 .
- ^ Перейти обратно: а б с д Кавалер-Смит Т (2013). «Ранняя эволюция способов питания эукариот, структурное разнообразие клеток и классификация типов простейших Loukozoa, Sulcozoa и Choanozoa». Европейский журнал протистологии . 49 (2): 115–178. дои : 10.1016/j.ejop.2012.06.001 . ПМИД 23085100 .
- ^ Перейти обратно: а б В. Фойснер; Х. Блаттерер; Я Фойсснер (1 октября 1988 г.). «Гемимастигофора (Hemimastix amphikineta nov. gen., nov. Spec.), новый тип протистанов из гондванских почв». Европейский журнал протистологии . 23 (4): 361–383. дои : 10.1016/S0932-4739(88)80027-0 . ISSN 0932-4739 . ПМИД 23195325 . Викиданные Q85570914 .
- ^ Гордон Лакс; Яна Эглит; Лаура Эме; Эрин М. Бертран; Эндрю Дж. Роджер ; Аластер ГБ Симпсон (14 ноября 2018 г.). «Гемимастигофора - это новая линия эукариот, принадлежащая к сверхцарству». Природа . 564 (7736): 410–414. дои : 10.1038/S41586-018-0708-8 . ISSN 1476-4687 . ПМИД 30429611 . Викиданные Q58834974 .
- ^ Шешкин, Егор (2022). « Spironematella terricola com. n. и Spironematella Goodeyi com. n. (Hemimastigida = Hemimastigea = Hemimastigophora) для Spironema terricola и Spironema Goodeyi с диагнозами рода и семейства Spironematellidae с поправками». Зоотакса . 5128 (2): 295–297. дои : 10.11646/zootaxa.5128.2.8 . ПМИД 36101172 . S2CID 252220401 .
- ^ Хейсс А.А., Уорринг С.Д., Лукач К., Фавате Дж., Ян А., Гьялтшен Ю., Филарди С., Симпсон А.Г.Б., Ким Э. (декабрь 2020 г.). «Описание Imasa heleensis, gen. nov., sp. nov. (Imasidae, fam. nov.), глубоковетвистой морской малавимонады и возможного ключевого таксона в понимании ранней эукариотической эволюции». Журнал эукариотической микробиологии . 68 : е12837. дои : 10.1111/jeu.12837 .
- ^ Карпов Сергей; Мамкаева Мария Александровна; Алеошин Владимир; Нассонова, Елена; Лилье, Осу; Глисон, Фрэнк Х. (1 января 2014 г.). «Морфология, филогения и экология афелид (Aphelidea, Opisthokonta) и предложение о новом супертипе Opisthosporidia» . Границы микробиологии . 5 : 112. дои : 10.3389/fmicb.2014.00112 . ПМЦ 3975115 . ПМИД 24734027 .
- ^ Перейти обратно: а б с Тихоненков Денис Владимирович, Михайлов Кирилл Владимирович, Гаврилюк Райан М.Р., Беляев Артем Олегович, Варша Матур, Карпов Сергей Александрович, Загумённый Дмитрий Григорьевич, Бородина Анастасия Сергеевна, Прокина Кристина Ивановна, Мыльников Александр Петрович, Владимир Владимирович. Алеошин и Патрик Дж. Килинг (7 декабря 2022 г.). «Микробные хищники образуют новую супергруппу эукариотов». Природа . 612 : 714–719. дои : 10.1038/S41586-022-05511-5 . ISSN 1476-4687 . Викиданные Q115933632 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Томас Кавальер-Смит ; Эма Э.Ю. Чао (апрель 2006 г.). «Филогения и мегасистематика фаготрофных гетероконтов (царство Chromista)». Журнал молекулярной эволюции . 62 (4): 388–420. дои : 10.1007/S00239-004-0353-8 . ISSN 0022-2844 . ПМИД 16557340 . Викиданные Q28303534 .
- ^ Тайнс М (2018). «Оомицеты» . Современная биология . 28 (15): Р812–Р813. дои : 10.1016/j.cub.2018.05.062 .
- ^ Т. Кавальер-Смит (1999). «Принципы нацеливания белков и липидов во вторичном симбиогенезе: происхождение эвгленоидных, динофлагеллятных и споровидных пластид и генеалогическое древо эукариот». Журнал эукариотической микробиологии . 46 (4): 347–66. doi : 10.1111/J.1550-7408.1999.TB04614.X . ISSN 1066-5234 . ПМИД 18092388 . Викиданные Q28261633 .
- ^ Перейти обратно: а б Шалчиан-Тебризи, К; Эйкрем, В; Клавенесс, Д; Вало, Д; Минге, Массачусетс; Ле Галль, Ф; Ромари, К; Трондсен, Дж; Ботнен, А; Массана, Р; Томсен, штат Ха; Якобсен, К.С. (28 апреля 2006 г.). «Телонемия, новый тип протистов, родственный хромистическим линиям» . Труды Королевского общества B: Биологические науки . 273 (1595): 1833–1842. дои : 10.1098/rspb.2006.3515 . ПМЦ 1634789 . ПМИД 16790418 .
- ^ Перейти обратно: а б Тихоненков Денис Владимирович; Джейми, Махваш; Бородина Анастасия С.; Беляев Артем О.; Загумённый Дмитрий Георгиевич; Прокина Кристина И.; Мыльников Александр П.; Бурки, Фабьен; Карпов, Сергей А. (2022). «О происхождении ЦАР: морфология, разнообразие и филогения телонемии» . Открытая биология . 12 (3). Королевское общество. дои : 10.1098/rsob.210325 . ISSN 2046-2441 . ПМЦ 8924772 . ПМИД 35291881 .
- ^ Корлисс, Джон О. (1984). «Королевство Протиста и его 45 типов». БиоСистемы . 17 (2): 87–176. дои : 10.1016/0303-2647(84)90003-0 . ПМИД 6395918 .
- ^ Перейти обратно: а б Эузеби Ж.П., Парте AC. «Названия типов» . Список названий прокариот, имеющих номенклатуру (LPSN) . Проверено 3 апреля 2022 г.
Внешние ссылки
[ редактировать ]- Являются ли типы «реальными»? Действительно ли существует четко определенное «количество типов животных», сохранившихся в летописи окаменелостей?
- Основные типы животных. Архивировано 16 июля 2006 г. в Wayback Machine.
| Базы данных органов управления : Национальные |
|---|