Диатомовая водоросль
| Диатомовая водоросль Временной диапазон: | |
|---|---|
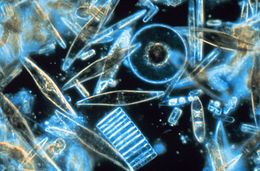 | |
| Световая микроскопия образца морских диатомей, обнаруженных между кристаллами ежегодного морского льда в Антарктиде, демонстрирующая разнообразие размеров и форм. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Клэйд : | Страменопилес |
| Тип: | Гириста |
| Подтип: | Охрофитина |
| Инфрафилум: | Диатомиста |
| Сорт: | Бацилляриофицеи Дэнджерд , 1933 год. [1] |
| Подклассы [2] | |
| Синонимы | |
| |
водоросль Диатомовая ( неолатинская диатома ) [а] Это любой член большой группы, состоящей из нескольких родов водорослей микроводорослей , в частности , обитающих в океанах, водных путях и почвах мира. Земли Живые диатомовые водоросли составляют значительную часть биомассы : они генерируют от 20 до 50 процентов кислорода, производимого на планете каждый год. [11] [12] поглощают более 6,7 миллиардов тонн кремния из вод, в которых они обитают, ежегодно [13] и составляют почти половину органического материала, обнаруженного в океанах. Раковины Сахары мертвых диатомей могут достигать глубины до полумили (800 м) на дне океана, а весь бассейн Амазонки ежегодно удобряется 27 миллионами тонн пыли панцирей диатомей, переносимой трансатлантическими ветрами из африканской , что намного его часть из впадины Боделе , которая когда-то состояла из системы пресноводных озер. [14] [15]
Диатомеи — одноклеточные организмы : они встречаются либо в виде одиночных клеток, либо в колониях , которые могут иметь форму лент, вееров, зигзагов или звезд. Размер отдельных клеток варьируется от 2 до 2000 микрометров. [16] При наличии достаточного количества питательных веществ и солнечного света совокупность живых диатомей удваивается примерно каждые 24 часа за счет бесполого множественного деления ; максимальная продолжительность жизни отдельных клеток составляет около шести дней. [17] Диатомеи имеют две различные формы: некоторые ( центрические диатомеи ) радиально симметричны, тогда как большинство ( пеннатные диатомеи ) имеют широкую двустороннюю симметрию.
Уникальной особенностью диатомовых водорослей является то, что они окружены клеточной стенкой из кремнезема (гидратированного диоксида кремния ), называемой панцирем . [18] Эти панцири дают структурную окраску , поэтому их называют «морскими драгоценностями» и «живыми опалами».
Движение диатомей в основном происходит пассивно в результате как океанских течений , так и ветровой турбулентности воды ; однако мужские гаметы центрических диатомей имеют жгутики , позволяющие активно двигаться в поисках женских гамет. Подобно растениям , диатомеи преобразуют световую энергию в химическую энергию посредством фотосинтеза , но их хлоропласты приобретаются разными способами. [19]
Что необычно для автотрофных организмов, диатомовые водоросли обладают циклом мочевины , особенностью, которую они разделяют с животными , хотя этот цикл используется для различных метаболических целей у диатомовых водорослей. Семейство Rhopalodiaceae также обладает цианобактериальным эндосимбионтом, называемым сфероидным телом. Этот эндосимбионт утратил свои фотосинтетические свойства, но сохранил способность осуществлять азотфиксацию , что позволяет диатомеям фиксировать атмосферный азот . [20] Другие диатомеи в симбиозе с азотфиксирующими цианобактериями относятся к родам Hemiaulus , Rhizosolenia и Chaetoceros . [21]
Динотомы — это диатомовые водоросли, ставшие эндосимбионтами внутри динофлагеллят. Исследования динофлагеллят Durinskia baltica и Glenodinium foliaceum показали, что событие эндосимбионта произошло настолько недавно, с точки зрения эволюции, что их органеллы и геном все еще не повреждены с минимальной потерей генов или вообще без нее. Основное различие между этими и свободноживущими диатомовыми водорослями заключается в том, что они утратили клеточную стенку из кремнезема, что делает их единственными известными диатомовыми водорослями без панциря. [22]
Изучение диатомовых водорослей — раздел психологии . Диатомовые водоросли классифицируются как эукариоты , организмы с ядерной оболочкой , связанным с клеточным ядром , которое отделяет их от прокариотов, архей и бактерий . Диатомовые водоросли — это тип планктона , называемый фитопланктоном , наиболее распространенный из типов планктона. Диатомеи также растут прикрепленными к бентосным субстратам, плавающим обломкам и макрофитам . Они составляют неотъемлемый компонент сообщества перифитона . [23] Другая классификация делит планктон на восемь типов в зависимости от размера: в этой схеме диатомеи относят к микроводорослям. Существует несколько систем классификации отдельных видов диатомей .
Ископаемые данные свидетельствуют о том, что диатомовые водоросли возникли во время или до раннего юрского периода, то есть примерно 150–200 миллионов лет назад. Самым старым ископаемым свидетельством существования диатомовых водорослей является образец современного рода Hemiaulus из янтаря позднеюрского периода из Таиланда. [24]
Диатомовые водоросли используются для мониторинга прошлых и настоящих условий окружающей среды и обычно используются в исследованиях качества воды. Кизельгур (диатомит) представляет собой совокупность панцирей диатомей, обнаруженных в земной коре. Это мягкие осадочные породы, содержащие кремнезем, которые легко измельчаются в мелкий порошок и обычно имеют размер частиц от 10 до 200 мкм. Кизельгур используется для различных целей, в том числе для фильтрации воды, в качестве мягкого абразива, в кошачьих туалетах и в качестве стабилизатора динамита.
Отображает наложения четырех флуоресцентных каналов.
(b) Голубой: [флуоресценция PLL-A546] - общий контрастный краситель для визуализации поверхностей эукариотических клеток.
(в) Синий: [флуоресценция Хехста] - окрашивает ДНК, идентифицирует ядра.
(d) Красный: [автофлуоресценция хлорофилла] - расщепляет хлоропласты. [27]
Обзор
[ редактировать ]Диатомеи — это протисты, которые образуют массовое ежегодное весеннее и осеннее цветение в водной среде и, по оценкам, отвечают за около половины фотосинтеза в мировых океанах. [28] Эта предсказуемая динамика ежегодного цветения способствует повышению трофических уровней и инициирует доставку углерода в глубоководный биом океана. Диатомовые водоросли имеют сложную стратегию жизненного цикла, которая, как предполагается, способствовала их быстрому генетическому разнообразию до примерно 200 000 видов. [29] которые распределены между двумя основными группами диатомей: центриками и пеннатами. [30] [31]
Морфология
[ редактировать ]Размер диатомей обычно составляет от 20 до 200 микрометров. [32] с несколькими более крупными видами. Их желтовато-коричневые хлоропласты , место фотосинтеза, типичны для гетероконтов , имеют четыре клеточные мембраны и содержат такие пигменты , как каротиноид фукоксантин . Особи обычно лишены жгутиков , но они присутствуют в мужских гаметах центрических диатомей и имеют обычное гетероконтное строение, включая волоски ( мастигонемы ), характерные для других групп.
Диатомовые водоросли часто называют «морскими драгоценностями» или «живыми опалами» из-за их оптических свойств. [33] Биологическая функция этой структурной окраски не ясна, но предполагается, что она может быть связана с общением, камуфляжем, теплообменом и/или защитой от ультрафиолета. [34]
Диатомовые водоросли строят сложные твердые, но пористые клеточные стенки, называемые панцирями, состоящие в основном из кремнезема . [35] : 25–30 Эта кремниевая стена [36] может иметь разнообразный узор с различными порами, ребрами, мелкими шипами, краевыми гребнями и возвышениями; все это можно использовать для разграничения родов и видов.
Сама клетка состоит из двух половин, каждая из которых содержит по существу плоскую пластинку, или створку, и краевую соединительную, или поясковую полоску. Одна половина, гипотека , немного меньше другой половины, эпитека . Морфология диатомей варьируется. Хотя форма ячейки обычно круглая, некоторые ячейки могут быть треугольными, квадратными или эллиптическими. Их отличительной особенностью является твердая минеральная оболочка или панцирь, состоящий из опала (гидратированной полимеризованной кремниевой кислоты).

- Ядро ; содержит генетический материал
- Ядрышко ; расположение хромосом
- аппарат Гольджи ; модифицирует белки и выводит их из клетки
- Клеточная стенка ; внешняя мембрана клетки
- Пиреноид ; центр фиксации углерода
- хроматофор ; мембранная структура, несущая пигмент
- Вакуоли ; везикула клетки, содержащая жидкость, связанную мембраной
- Цитоплазматические тяжи; держать ядро
- Митохондрии ; создавать АТФ (энергию) для клетки
- Клапаны/Стрии; позволяют питательным веществам проникать в клетку и выводить их из клетки
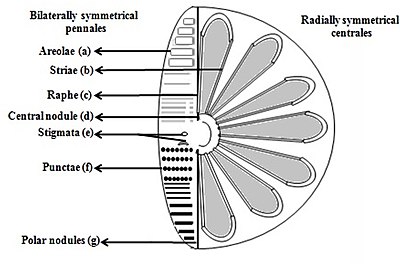
- Ареолы (шестиугольные или многоугольные коробчатые перфорации с ситом на поверхности диатомей)
- Стрии (поры, точки, пятна или точки, расположенные в линию на поверхности)
- Раф (разрез в клапанах)
- Центральный узел (утолщение стенки в середине шва)
- Стигматы (отверстия на поверхности створки, которые снаружи выглядят закругленными, но с щелью, как внутри)
- Точки (пятна или небольшие перфорации на поверхности)
- Полярные узелки (утолщение стенки на дистальных концах шва) [37] [38]

Диатомеи делятся на две группы, отличающиеся формой панциря: центрические диатомеи и пеннатные диатомеи .
Пеннатные диатомеи двусторонне-симметричны. Каждый из их клапанов имеет отверстия в виде прорезей вдоль швов , а их оболочки обычно вытянуты параллельно этим швам. Они вызывают движение клеток через цитоплазму, которая течет по швам, всегда перемещаясь по твердым поверхностям.
Центрические диатомеи радиально симметричны. Они состоят из верхних и нижних створок – эпитеки и гипотеки – каждая из которых состоит из створки и пояска, которые могут легко скользить друг под другом и расширяться, увеличивая содержание клеток по мере продвижения диатомовых водорослей. Цитоплазма центрической диатомеи расположена вдоль внутренней поверхности раковины и образует полую выстилку вокруг крупной вакуоли, расположенной в центре клетки. Эта большая центральная вакуоль заполнена жидкостью, известной как «клеточный сок», которая похожа на морскую воду, но варьируется в зависимости от конкретного содержания ионов. Цитоплазматический слой является домом для нескольких органелл, таких как хлоропласты и митохондрии. Прежде чем центрическая диатомея начнет расширяться, ее ядро находится в центре одной из створок и начинает двигаться к центру цитоплазматического слоя еще до завершения деления. Центрические диатомеи имеют разнообразную форму и размеры в зависимости от того, от какой оси отходит раковина и имеются ли шипы.

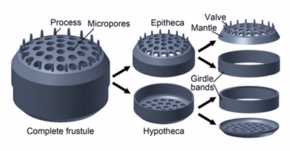
окремнение
[ редактировать ]Клетки диатомей заключены в уникальную клеточную стенку из кремнезема , известную как панцирь, состоящую из двух створок, называемых теками , которые обычно перекрывают друг друга. [40] Биогенный кремнезем, составляющий клеточную стенку, внутриклеточно путем полимеризации мономеров кремниевой кислоты синтезируется . Затем этот материал экструдируется на внешнюю поверхность ячейки и добавляется к стене. У большинства видов, когда диатомея делится с образованием двух дочерних клеток, каждая клетка сохраняет одну из двух половин и вырастает внутри нее меньшую половину. В результате после каждого цикла деления средний размер клеток диатомей в популяции уменьшается. Как только такие клетки достигают определенного минимального размера, они не просто делятся, а обращают этот процесс вспять, образуя ауксоспору , обычно посредством мейоза и полового размножения, но существуют исключения. Ауксоспора увеличивается в размерах, образуя гораздо более крупную клетку, которая затем возвращается к делениям, уменьшающимся в размерах. [41]



Точный механизм переноса кремнезема, поглощенного диатомовыми водорослями, на клеточную стенку неизвестен. Большая часть секвенирования генов диатомовых водорослей связана с поиском механизма поглощения и отложения кремнезема в наноразмерных структурах панциря . Наибольшего успеха в этой области добились два вида: Thalassiosira pseudonana , который стал модельным видом, поскольку был секвенирован весь геном и установлены методы генетического контроля, и Cylindrotheca fusiformis , у которого впервые были обнаружены важные белки отложения кремнезема - силафины. обнаруженный. [43] Силафины, наборы поликатионных пептидов , были обнаружены в клеточных стенках C. fusiformis и могут создавать сложные структуры кремнезема. В этих структурах наблюдались поры размеров, характерных для структуры диатомей. Когда T. pseudonana подвергся анализу генома, было обнаружено, что он кодирует цикл мочевины , включая большее количество полиаминов , чем большинство геномов, а также три различных гена транспорта кремнезема. [44] В филогенетическом исследовании генов транспорта кремнезема из 8 различных групп диатомовых водорослей было обнаружено, что транспорт кремнезема обычно группируется в зависимости от вида. [43] Это исследование также обнаружило структурные различия между переносчиками кремнезема пеннатных (двусторонняя симметрия) и центрических (радиальная симметрия) диатомей. Последовательности, сравниваемые в этом исследовании, были использованы для создания разнообразного фона с целью идентификации остатков, которые дифференцируют функции в процессе осаждения кремнезема. Кроме того, то же исследование показало, что ряд областей консервативен внутри видов, что, вероятно, является базовой структурой транспорта кремнезема.
Эти транспортные белки кремнезема уникальны для диатомовых водорослей, их гомологи не обнаружены у других видов, таких как губки или рис. Дивергенция этих генов транспорта кремнезема также указывает на структуру белка, развивающуюся из двух повторяющихся единиц, состоящих из пяти мембраносвязанных сегментов, что указывает либо на дупликацию гена, либо на его димеризацию . [43] Было высказано предположение, что отложение кремнезема, происходящее из мембраносвязанных везикул диатомовых водорослей, является результатом активности силафинов и длинноцепочечных полиаминов. Эта везикула отложения кремнезема (SDV) характеризуется как кислый отсек, слитый с везикулами, полученными из аппарата Гольджи. [45] Было показано, что эти две белковые структуры in vivo создают листы узорчатого кремнезема с порами неправильной формы, напоминающими панцири диатомовых водорослей . Одна из гипотез относительно того, как эти белки создают сложную структуру, заключается в том, что остатки консервативны внутри SDV, что, к сожалению, трудно идентифицировать или наблюдать из-за ограниченного числа доступных разнообразных последовательностей. Хотя точный механизм высокоравномерного отложения кремнезема пока неизвестен, гены Thalassiosira pseudonana , связанные с силафинами, рассматриваются как мишени для генетического контроля наномасштабного отложения кремнезема.
Способность диатомей создавать на основе кремнезема клеточные стенки была предметом восхищения на протяжении веков. Все началось с микроскопического наблюдения анонимного английского сельского дворянина в 1703 году, который наблюдал объект, похожий на цепочку правильных параллелограммов, и спорил, были ли это просто кристаллы соли или растение. [46] Зритель решил, что это растение, потому что параллелограммы не разделялись при встряхивании и не меняли внешний вид при сушке или воздействии теплой водой (в попытке растворить «соль»). Сам того не зная, замешательство зрителя уловило суть диатомовых водорослей — растений, использующих минералы. Неясно, когда было установлено, что клеточные стенки диатомовых водорослей состоят из кремнезема, но в 1939 году в оригинальной статье этот материал охарактеризовался как кремниевая кислота в «субколлоидном» состоянии. [47] Идентификация основного химического компонента клеточной стенки стимулировала исследования того, как она устроена. Эти исследования включали и стимулировались различными подходами, включая микроскопию, химию, биохимию, характеристику материалов , молекулярную биологию , омику и трансгенные подходы. Результаты этой работы позволили лучше понять процессы формирования клеточной стенки, установив фундаментальные знания, которые можно использовать для создания моделей, которые контекстуализируют текущие результаты и поясняют, как работает этот процесс. [48]
Процесс построения клеточной стенки на основе минералов внутри клетки, а затем ее экспорта наружу — это масштабное событие, в котором должно участвовать большое количество генов и их белковых продуктов. Процесс построения и экзоцитоза этого крупного структурного объекта за короткий период времени, синхронизированный с развитием клеточного цикла способностей клетки , требует существенных физических движений внутри клетки, а также использования значительной части биосинтетических . [48]
Первые характеристики биохимических процессов и компонентов, участвующих в окварцевании диатомей, были сделаны в конце 1990-х годов. [49] [50] [51] За этим последовало понимание того, как может происходить сборка кремнеземных структур более высокого порядка. [52] [53] [54] В более поздних отчетах описывается идентификация новых компонентов, участвующих в процессах более высокого порядка, динамика, документированная с помощью визуализации в реальном времени, и генетические манипуляции со структурой кремнезема. [55] [56] Подходы, установленные в этих недавних работах, предоставляют практические возможности не только идентифицировать компоненты, участвующие в формировании клеточной стенки кремнезема, но и выяснить их взаимодействия и пространственно-временную динамику. Этот тип целостного понимания будет необходим для достижения более полного понимания синтеза клеточной стенки. [48]
Поведение
[ редактировать ]
Отлично, 1897 г.
Большинство центрических и арафидных пеннатных диатомей неподвижны , а их относительно плотные клеточные стенки заставляют их легко тонуть. Планктонные формы в открытой воде обычно полагаются на турбулентное перемешивание верхних слоев океанических вод ветром, чтобы удерживать их во взвешенном состоянии в освещенных солнцем поверхностных водах. Многие планктонные диатомеи также развили особенности, замедляющие скорость их погружения, такие как шипы или способность расти колониальными цепочками. [57] Эти приспособления увеличивают соотношение площади поверхности к объему и сопротивление , что позволяет им дольше оставаться в толще воды. Отдельные клетки могут регулировать плавучесть с помощью ионного насоса. [58]
Некоторые пеннатные диатомеи способны к типу передвижения, называемому «скольжением», который позволяет им перемещаться по поверхности с помощью клейкой слизи, выделяемой через шовную структуру, называемую швом. [59] [60] Чтобы клетка диатомеи могла скользить, она должна иметь твердую подложку, к которой прилипает слизь.
Клетки одиночные или объединены в колонии разного типа, которые могут быть связаны кремниевыми структурами; слизистые подушечки, стебли или трубочки; аморфные массы слизи; или нитями хитина (полисахарида), которые выделяются через расположенные в клетке отростки.


На этой проекции стопки конфокальных изображений диатомовых водорослей показаны клеточная стенка (голубой), хлоропласты (красный), ДНК (синий), мембраны и органеллы (зеленый).
Жизненный цикл
[ редактировать ]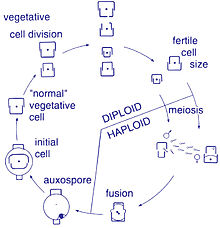
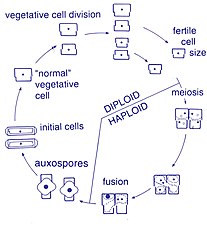
Размножение и размер клеток
[ редактировать ]Размножение этих организмов бесполое путем бинарного деления , во время которого диатомея делится на две части, образуя две «новые» диатомеи с идентичными генами. Каждый новый организм получает одну из двух панцирей — одну большую, другую меньшую — принадлежащую родительскому организму, которая теперь называется эпитекой ; и используется для построения второго, меньшего панциря, гипотеки . Диатомея, получившая панцирь большего размера, становится того же размера, что и ее родитель, но диатомея, получившая панцирь меньшего размера, остается меньше своего родителя. Это приводит к уменьшению среднего размера клеток этой популяции диатомей. [16] Однако было замечено, что некоторые таксоны обладают способностью делиться, не вызывая уменьшения размера клеток. [61] Тем не менее, чтобы восстановить размер клеток популяции диатомей для тех, которые переносят уменьшение размера, ауксоспор . должно произойти половое размножение и образование [16]
Деление клеток
[ редактировать ]Вегетативные клетки диатомей диплоидны (2N), поэтому может происходить мейоз , в результате которого образуются мужские и женские гаметы, которые затем сливаются, образуя зиготу . Зигота сбрасывает кремнеземную теку и вырастает в большую сферу, покрытую органической мембраной — ауксоспорой. Внутри ауксоспоры образуется новая клетка диатомовой водоросли максимального размера, исходная клетка, начиная новое поколение. Покоящиеся споры также могут образовываться в ответ на неблагоприятные условия окружающей среды, а прорастание происходит при улучшении условий. [35]
Определяющей характеристикой всех диатомей является их ограничительная и двудольная клеточная стенка из кремнезема, которая заставляет их постепенно сокращаться во время бесполого деления клеток. При критически малом размере клеток и при определенных условиях ауксоспоруляция восстанавливает размер клеток и предотвращает гибель клонов . [62] [63] [64] [65] [66] Описаны полные жизненные циклы лишь нескольких диатомовых водорослей, и половые события редко фиксируются в окружающей среде. [31]
Половое размножение
[ редактировать ]Большинство эукариот способны к половому размножению с участием мейоза . Половое размножение, по-видимому, является обязательной фазой жизненного цикла диатомовых водорослей, особенно потому, что размер клеток уменьшается при последовательных вегетативных делениях. [67] Половое размножение включает образование гамет и их слияние с образованием зиготы , в которой восстанавливается максимальный размер клеток. [67] Передача сигналов, запускающая сексуальную фазу, благоприятствует, когда клетки накапливаются вместе, так что расстояние между ними сокращается, а контакты и/или восприятие химических сигналов облегчаются. [68]
Исследование геномов пяти диатомовых водорослей и транскриптома одной диатомовой водоросли привело к идентификации 42 генов , потенциально участвующих в мейозе. [69] Таким образом, набор мейотических инструментов, по-видимому, консервативен у этих шести видов диатомей. [69] что указывает на центральную роль мейоза у диатомовых водорослей, как и у других эукариот.
Подвижность сперматозоидов
[ редактировать ]Диатомовые водоросли в основном неподвижны ; однако сперматозоиды, обнаруженные у некоторых видов, могут быть жгутиковыми , хотя подвижность обычно ограничивается скользящим движением. [35] У центрических диатомей мелкие мужские гаметы имеют один жгутик , а женские гаметы крупные и неподвижные ( огамные ). И наоборот, у пеннатных диатомей обе гаметы лишены жгутиков ( изогамны ). [16] Некоторые виды арафид, то есть пеннатные диатомеи без шва (шва), были зарегистрированы как анизогамные и, следовательно, считаются переходной стадией между центрическими и рафидными пеннатными диатомеями, диатомеями со швом. [61]
Разложение микробами
[ редактировать ]Некоторые виды бактерий в океанах и озерах могут ускорить растворение кремнезема в мертвых и живых диатомовых водорослях, используя гидролитические ферменты для расщепления органического материала водорослей. [70] [71]
Экология
[ редактировать ]| Часть серии о |
| Планктон |
|---|
 |


в зависимости от концентрации силиката [72]
Распределение
[ редактировать ]Диатомовые водоросли — широко распространенная группа, их можно найти в океанах , пресной воде , почве и на влажных поверхностях. Они являются одним из доминирующих компонентов фитопланктона в богатых питательными веществами прибрежных водах и во время весеннего цветения океана, поскольку могут делиться быстрее, чем другие группы фитопланктона. [73] Большинство из них живут пелагически в открытой воде, хотя некоторые живут в виде поверхностных пленок на границе раздела вода-отложения ( бентос ) или даже во влажных атмосферных условиях. Они особенно важны в океанах, где исследование 2003 года показало, что на их долю приходится около 45% общего первичного производства органического материала в океане. [74] Однако, по оценкам более недавнего исследования 2016 года, это число приближается к 20%. [75] Пространственное распространение видов морского фитопланктона ограничено как по горизонтали, так и по вертикали. [76] [35]
Рост
[ редактировать ]Планктонные диатомеи в пресноводной и морской среде обычно ведут образ жизни « бум и спад » (или « расцвет и спад»). При благоприятных условиях в верхнем перемешанном слое (питательные вещества и свет) (как весной ) их конкурентоспособность и высокая скорость роста [73] позволяет им доминировать в сообществах фитопланктона («бум» или «цветение»). По существу их часто классифицируют как оппортунистические r-стратеги (т.е. те организмы, экология которых определяется высокой скоростью роста r ).
Влияние
[ редактировать ]Пресноводная диатомовая водоросль Didymosphenia geminata , широко известная как Didymo, вызывает серьезную деградацию окружающей среды в водотоках, где она цветет, производя большое количество коричневого желеобразного материала, называемого «коричневыми солями» или «каменными солями». Эта диатомовая водоросль произрастает в Европе и является инвазивным видом как среди антиподов , так и в некоторых частях Северной Америки . [77] [78] Проблема чаще всего фиксируется в Австралии и Новой Зеландии . [79]
Когда условия становятся неблагоприятными, обычно из-за истощения питательных веществ, клетки диатомовых водорослей обычно увеличивают скорость погружения и покидают верхний смешанный слой («перебор»). Это погружение вызвано либо потерей контроля над плавучестью, синтезом слизи, которая склеивает клетки диатомей, либо образованием тяжелых покоящихся спор . Опускание верхнего смешанного слоя выводит диатомовые водоросли из условий, неблагоприятных для роста, включая популяции травоядных и более высокие температуры (которые в противном случае увеличили бы клеточный метаболизм ). Клетки, достигающие более глубокой воды или мелкого морского дна, могут затем отдохнуть, пока условия снова не станут более благоприятными. В открытом океане многие тонущие клетки теряются на глубине, но вблизи термоклина могут сохраняться убежища .
В конечном итоге клетки диатомей в этих покоящихся популяциях снова попадают в верхний смешанный слой, когда их увлекает вертикальное перемешивание. В большинстве случаев это смешивание также пополняет питательные вещества в верхнем перемешанном слое, создавая условия для следующего цикла цветения диатомей. В открытом океане (вдали от зон непрерывного апвеллинга) [80] ), этот цикл цветения, спада, а затем возвращения к состоянию, предшествующему цветению, обычно происходит в течение годового цикла, при этом диатомеи преобладают только весной и в начале лета. Однако в некоторых местах может произойти осеннее цветение, вызванное нарушением летней стратификации и уносом питательных веществ, в то время как уровень освещенности все еще достаточен для роста. Поскольку вертикальное перемешивание увеличивается, а уровень освещенности падает с приближением зимы, эти цветы меньше и менее продолжительны, чем их весенние эквиваленты.
В открытом океане цветение диатомей (весеннее) обычно заканчивается нехваткой кремния. В отличие от других минералов, потребность в кремнии уникальна для диатомовых водорослей, и он не регенерируется в планктонной экосистеме так эффективно, как, например, азотные или фосфорные питательные вещества. Это можно увидеть на картах приземных концентраций питательных веществ: по мере того, как количество питательных веществ снижается по градиенту, первым истощается кремний (обычно за ним следует азот, а затем фосфор).
Считается, что из-за этого цикла цветения и спада диатомеи играют непропорционально важную роль в экспорте углерода из поверхностных вод океана. [80] [81] (см. также биологический насос ). они также играют ключевую роль в регуляции биогеохимического цикла кремния Примечательно, что в современном океане. [74] [82]
Причина успеха
[ редактировать ]Диатомеи экологически успешны и встречаются практически во всех средах, где есть вода – не только в океанах, морях, озерах и ручьях, но также в почве и водно-болотных угодьях. [ нужна ссылка ] Многие исследователи считают, что использование кремния диатомовыми водорослями является ключом к этому экологическому успеху. Ворон (1983) [83] отметили, что по сравнению с органическими клеточными стенками панцири кремнезема требуют меньше энергии для синтеза (приблизительно 8% от сопоставимой органической стенки), что потенциально обеспечивает значительную экономию общего энергетического баланса клетки. В классическом исследовании Эгге и Акснеса (1992) [72] диатомей обнаружили, что доминирование в сообществах мезокосма напрямую связано с наличием кремниевой кислоты - когда концентрации превышали 2 мкмоль · м. −3 , они обнаружили, что диатомеи обычно составляют более 70% сообщества фитопланктона. Другие исследователи [84] предположили, что биогенный кремнезем в клеточных стенках диатомовых водорослей действует как эффективный pH буферный агент , способствуя превращению бикарбоната в растворенный CO 2 (который легче усваивается). В более общем плане, несмотря на эти возможные преимущества, обеспечиваемые использованием кремния, диатомеи обычно имеют более высокие темпы роста, чем другие водоросли того же соответствующего размера. [73]
Источники для коллекции
[ редактировать ]Диатомовые водоросли можно получить из нескольких источников. [85] Морские диатомовые водоросли можно собирать путем прямого отбора проб воды, а донные формы можно обнаружить путем соскабливания ракушек , устриц и других раковин. Диатомеи часто присутствуют в виде коричневого скользкого налета на погруженных в воду камнях и палках, и их можно увидеть «струящимися» с речным течением. Поверхностный ил пруда, канавы или лагуны почти всегда содержит некоторое количество диатомовых водорослей. Живые диатомеи часто встречаются в больших количествах цепляющимися за нитчатые водоросли или образующими студенистые массы на различных подводных растениях. Cladophora часто покрыта Cocconeis , диатомеей эллиптической формы; Ваухерия часто покрывается мелкими формами. Поскольку диатомовые водоросли составляют важную часть пищи моллюсков , оболочников и рыб , в пищеварительном тракте этих животных часто образуются формы, которые нелегко получить другими способами. Диатомеи можно заставить появиться, наполнив банку водой и грязью, обернув ее черной бумагой и позволив прямым солнечным лучам падать на поверхность воды. В течение дня диатомовые водоросли выйдут на поверхность в виде накипи, и их можно будет изолировать. [85]
Биогеохимия
[ редактировать ]
Кремнеземный цикл
[ редактировать ]На диаграмме показаны основные потоки кремния . в современном океане Большая часть биогенного кремнезема в океане ( кремнезем, образующийся в результате биологической деятельности ) происходит из диатомовых водорослей. Диатомовые водоросли извлекают растворенную кремниевую кислоту из поверхностных вод по мере роста и возвращают ее в толщу воды , когда умирают. Кремний поступает сверху через эоловую пыль , с побережья через реки и снизу через переработку отложений морского дна , выветривание и гидротермальную активность . [82]
Хотя диатомовые водоросли, возможно, существовали со времен триаса , время их господства и «захвата» кремниевого цикла произошло совсем недавно. до фанерозоя Считается, что (до 544 млн лет назад) микробные или неорганические процессы слабо регулировали круговорот кремния в океане. [86] [87] [88] Впоследствии в цикле, по-видимому, доминируют (и более строго регулируются) радиолярии и кремнистые губки , первые как зоопланктон , вторые как оседлые фильтраторы преимущественно на континентальных шельфах . [89] Считается, что за последние 100 млн лет кремниевый цикл оказался под еще более жестким контролем, и это связано с экологическим господством диатомовых водорослей.
Однако точные сроки «перехвата власти» остаются неясными, и разные авторы имеют противоречивые интерпретации летописи окаменелостей. Некоторые свидетельства, такие как смещение кремниевых губок с полок, [90] предполагает, что это поглощение началось в меловом периоде (от 146 до 66 млн лет назад), в то время как данные радиолярий предполагают, что «поглощение» началось только в кайнозое (от 66 млн лет до настоящего времени). [91]
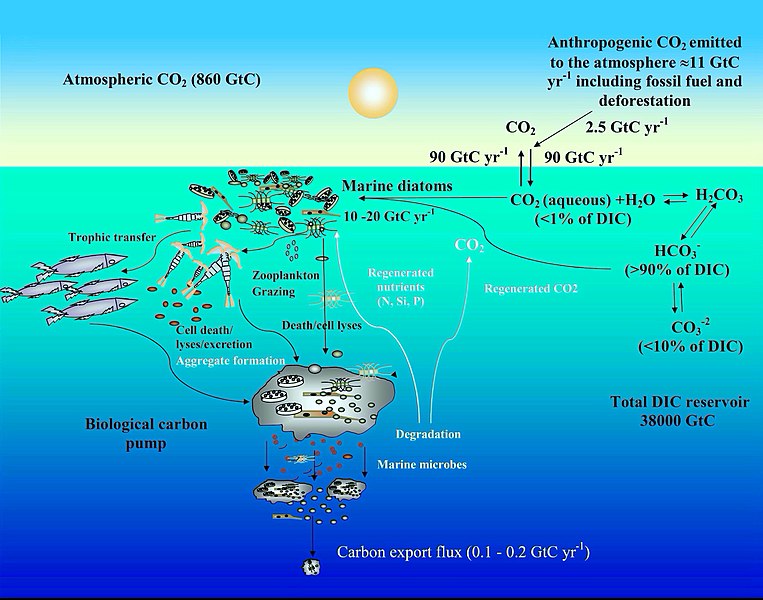 Цикл углерода в океане и механизмы концентрации диатомового углекислого газа [92]
Цикл углерода в океане и механизмы концентрации диатомового углекислого газа [92]

Углеродный цикл
[ редактировать ]На диаграмме показаны некоторые механизмы, с помощью которых морские диатомовые водоросли участвуют в биологическом углеродном насосе и влияют на углеродный цикл океана . Антропогенные выбросы CO 2 в атмосферу (в основном вызванные сжиганием ископаемого топлива и вырубкой лесов) составляют почти 11 гигатонн углерода (ГтС) в год, из которых почти 2,5 ГтС поглощается поверхностью океана. В поверхностной морской воде ( рН 8,1–8,4) бикарбонат ( HCO −
3 ) и карбонат- ионы ( CO 2−
3 ) составляют почти 90 и <10% растворенного неорганического углерода (DIC) соответственно, тогда как растворенный CO 2 (водный CO 2 ) составляет <1%. Несмотря на низкий уровень CO 2 в океане и медленную скорость его диффузии в воде, диатомовые водоросли ежегодно фиксируют 10–20 ГтС посредством фотосинтеза благодаря своим механизмам концентрации углекислого газа , что позволяет им поддерживать морские пищевые цепи . Кроме того, 0,1–1% этого органического материала, образующегося в эвфотическом слое , оседает в виде частиц, перенося таким образом поверхностный углерод в глубины океана и изолируя атмосферный CO 2 на тысячи лет или дольше. Оставшееся органическое вещество реминерализируется посредством дыхания. Таким образом, диатомовые водоросли являются одним из основных участников этого биологического углеродного насоса, который, возможно, является наиболее важным биологическим механизмом в системе Земля, позволяющим CO 2 удалять из углеродного цикла на очень длительный период. [93] [92]
 Митохондриальный цикл мочевины в родовой диатомовой клетке и потенциальная судьба промежуточных продуктов цикла мочевины [94]
Митохондриальный цикл мочевины в родовой диатомовой клетке и потенциальная судьба промежуточных продуктов цикла мочевины [94]

Цикл мочевины
[ редактировать ]Особенностью диатомовых водорослей является цикл мочевины , который эволюционно связывает их с животными. В 2011 году Аллен и др. установили, что у диатомей имеется функционирующий цикл мочевины. Этот результат был значительным, поскольку до этого считалось, что цикл мочевины возник у многоклеточных животных , которые появились на несколько сотен миллионов лет раньше диатомовых водорослей. Их исследование показало, что, хотя диатомовые водоросли и животные используют цикл мочевины для разных целей, они считаются эволюционно связанными таким образом, что животные и растения — нет. [95]
Хотя в фотосинтезирующих организмах митохондрии часто упускают из виду, они также играют решающую роль в энергетическом балансе. Имеют значение два пути, связанных с азотом, которые также могут меняться под действием аммония ( NH +
4 ) питание по сравнению с нитратным ( NO −
3 ) питание. Во-первых, у диатомовых водорослей и, вероятно, у некоторых других водорослей существует цикл мочевины. [96] [97] [98] Давно известная функция цикла мочевины у животных заключается в выведении избытка азота, образующегося в результате катаболизма аминокислот ; Как и фотодыхание , цикл мочевины долгое время считался каналом отходов. Однако у диатомовых водорослей цикл мочевины, по-видимому, играет роль в обмене питательных веществ между митохондриями и цитоплазмой и, возможно, пластидой. [99] и может помочь регулировать метаболизм аммония. [96] [97] Из-за этого цикла морские диатомеи, в отличие от хлорофитов , также обзавелись митохондриальным переносчиком мочевины и, по сути, на основе биоинформатики о полном митохондриальном цикле GS-GOGAT . была выдвинута гипотеза [97] [94]
Другой
[ редактировать ]Диатомовые водоросли в основном фотосинтезируют; однако некоторые из них являются облигатными гетеротрофами и могут жить в отсутствие света при наличии соответствующего источника органического углерода. [100] [101]
Фотосинтезирующие диатомеи, оказавшиеся в среде, лишенной кислорода и/или солнечного света, могут переключиться на анаэробное дыхание, известное как нитратное дыхание (ДНРА), и оставаться в состоянии покоя до месяцев и десятилетий. [102] [103]
Основными пигментами диатомовых водорослей являются хлорофиллы А и С, бета-каротин , фукоксантин , диатоксантин и диадиноксантин. [16]
Таксономия
[ редактировать ]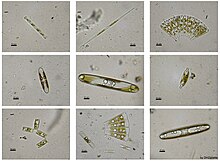



Стефанодиск Ханцский

Нервная истмия Нервная истмия
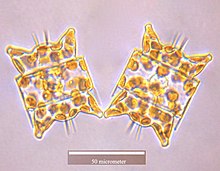
Одонтелла аурита
Диатомеи относятся к большой группе протистов, многие из которых содержат пластиды, богатые хлорофиллами а и с. Группу по-разному называют гетероконтами , хризофитами , хромистами или страменопилями . Многие из них являются автотрофами , например, золотые водоросли и водоросли ; и гетеротрофы, такие как водные плесени , опалиниды и актинофриидные солнечники. Классификация этого ареала протистов до сих пор не определена. С точки зрения ранга к ним относились как к подразделению , типу , королевству или чему-то промежуточному между ними. Следовательно, диатомеи ранжируются от класса, обычно называемого Diatomophyceae или Bacillariophyceae , до подразделения (= типа), обычно называемого Bacillariophyta , с соответствующими изменениями в рангах их подгрупп.
Роды и виды
[ редактировать ]Считается, что существует около 20 000 существующих диатомей видов , из которых около 12 000 получили названия на сегодняшний день, согласно Guiry, 2012. [104] (другие источники дают более широкий диапазон оценок [16] [105] [106] [107] ). Описано около 1000–1300 родов диатомей, как современных, так и ископаемых. [108] [109] из которых около 250–300 существуют только в виде окаменелостей. [110]
Занятия и заказы
[ редактировать ]В течение многих лет диатомовые водоросли, рассматриваемые либо как класс (Bacillariophyceae), либо как тип (Bacillariophyta), были разделены всего на два порядка, соответствующие центрическим и пеннатным диатомовым водорослям ( Centrales и Pennales ). Эта классификация была тщательно пересмотрена Раундом, Кроуфордом и Манном в 1990 году, которые присвоили диатомовым водорослям более высокий ранг (подразделение, соответствующее типу в зоологической классификации) и переместили основные классификационные единицы в классы, сохранив центрические диатомовые водоросли как единый класс Coscinodiscophyceae. , но разделив бывшие пеннатные диатомовые водоросли на два отдельных класса: Fragilariophyceae и Bacillariophyceae (последнее старое название сохранено, но с исправленным определением), между ними имеется 45 отрядов, большинство из которых новые.
Сегодня (написано в середине 2020 г.) признано, что система Round et al. 1990 г. нуждается в пересмотре с появлением новых молекулярных исследований, однако неясно, какая система лучше всего ее заменит, а текущие системы широко используются, такие как AlgaeBase , Всемирный регистр морских видов и входящая в него база данных DiatomBase , а также система для « вся жизнь», представленные в работе Ruggiero et al., 2015, все сохраняют Round et al. в качестве их основы, хотя диатомеи в целом рассматриваются как класс, а не как подраздел/тип, а классы Раунда и др. сокращены до подклассов для лучшего согласования с трактовкой филогенетически соседних групп и содержащих их таксонов. (Ссылки см. в отдельных разделах ниже).
Одно из предложений Линды Медлин и ее коллег, выдвинутое в 2004 году, заключается в том, чтобы некоторые центрические отряды диатомовых водорослей, считающиеся более тесно связанными с пеннатными, выделить в новый класс Mediophyceae, который сам более тесно связан с пеннатными диатомовыми водорослями, чем с пеннатными. остальные центрики. Эта гипотеза, позже названная гипотезой Coscinodiscophyceae-Mediophyceae-Bacillariophyceae или Coscinodiscophyceae+(Mediophyceae+Bacillariophyceae) (CMB), была принята, в частности, Д.Г. Манном, который использует ее в качестве основы для классификации диатомей, представленной в Adl. и др. (2005, 2012, 2019), а также в главе Bacillariophyta Справочника протистов 2017 г. под редакцией Арчибальда и др., с некоторыми изменениями, отражающими очевидную немонофилию Medlin et al. оригинальные «Coscinodiscophyceae». Между тем, группа под руководством EC Theriot выступает за другую гипотезу филогении, которая получила название гипотезы структурной градации (SGH) и не признает Mediophyceae как монофилетическую группу, в то время как другой анализ, проведенный Parks et al., 2018, обнаруживает, что радиальноцентрические диатомеи (Coscinodiscophyceae Медлина и др.) не являются монофилетическими, но поддерживают монофилию Mediophyceae минус Аттея — аномальный род. Обсуждение относительных достоинств этих противоречивых схем продолжается различными участвующими сторонами. [111] [112] [113] [114]
Адл и др., лечение, 2019 г.
[ редактировать ]В 2019 году Адл и др. [115] представил следующую классификацию диатомей, отметив при этом: «Этот пересмотр отражает многочисленные достижения в филогении диатомей за последнее десятилетие. Из-за нашей плохой выборки таксонов за пределами Mediophyceae и пеннатных диатомей, а также известного и ожидаемого разнообразия всех диатомей , многие клады появляются на высоком уровне классификации (а классификация более высокого уровня довольно плоская)». Эта классификация рассматривает диатомеи как тип (Diatomeae/Bacillariophyta), принимает класс Mediophyceae Медлина и его коллег, вводит новые подтипы и классы для ряда изолированных в противном случае родов и перераспределяет ряд ранее установленных таксонов в подклассы. но не перечисляет отряды или семейства. Предполагаемые ранги были добавлены для ясности (Адл и др. не используют ранги, но предполагаемые ранги в этой части классификации очевидны из выбора используемых окончаний в используемой системе ботанической номенклатуры).
- Clade Diatomista Derelle et al. 2016, исправлено. Кавальер-Смит 2017 (диатомовые водоросли плюс подмножество других групп охрофитов)
- Тип Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (диатомовые водоросли)
- Подтип Leptocylindrophytina Д.Г. Манн в Adl et al. 2019 год
- Класс Leptocylindrophyceae DG Mann в Adl et al. 2019 ( Лептоцилиндрус , Тенуицилиндрус )
- Класс Corethrophyceae DG Mann в Adl et al. 2019 ( Коретрон )
- Подтип Ellerbeckiophytina DG Mann в Adl et al. ( Эллербекия 2019
- Подтип Probosciophytina DG Mann в Adl et al. 2019 ( Хоботка )
- Подтип Melosirophytina DG Mann в Adl et al. 2019 ( Аулакосейра , Мелозира , Гиалодиск , Стефанопиксис , Паралия , Эндиктия )
- Подтип Coscinodiscophytina Medlin & Kaczmarska 2004, исправить. ( Actinoptychus , Coscinodiscus , Actinocyclus , Asteromphalus , Aulacodiscus , Stellarima )
- Подтип Rhizosoleniophytina DG Mann в Adl et al. 2019 ( Гинардия , Ризосоления , Псевдосоления )
- Подтип Arachnoidiscophytina DG Mann in Adl et al. 2019 ( Паукообразный )
- Подтип Bacillariophytina Medlin & Kaczmarska 2004, исправить.
- Класс Mediophyceae Jouse и Прошкина-Лавренко в Медлине и Качмарской, 2004 г.
- Подкласс Chaetocerotophycidae Round и RM Crawford в Round et al. 1990, исправлено.
- Подкласс Lithodesmiophycidae Round и RM Crawford в Round et al. 1990, исправлено.
- Подкласс Thalassiosirophycidae Round и RM Crawford в Round et al. 1990 год
- Подкласс Cymatosirophycidae Round и RM Crawford в Round et al. 1990 год
- Подкласс Odontellophycidae DG Mann в Adl et al. 2019 год
- Подкласс Chrysanthemodiscophycidae DG Mann в Adl et al. 2019 год
- Класс Biddulphiophyceae DG Mann в Adl et al. 2019 год
- Подкласс Biddulphiophycidae Round и RM Crawford в Round et al. 1990, исправлено.
- Biddulphiophyceae неопределенный ареал обитания ( Attheya )
- Класс Bacillariophyceae Haeckel 1878, исправить.
- Bacillariophyceae неопределенная среда обитания (Striatellaceae)
- Подкласс Urneidophycidae Medlin 2016
- Подкласс Fragilariophycidae Round in Round, Crawford & Mann 1990, исправить.
- Подкласс Bacillariophycidae DG Mann в Round, Crawford & Mann 1990, исправить.
- Класс Mediophyceae Jouse и Прошкина-Лавренко в Медлине и Качмарской, 2004 г.
- Подтип Leptocylindrophytina Д.Г. Манн в Adl et al. 2019 год
- Тип Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (диатомовые водоросли)
см. в таксономии диатомовых водорослей Более подробную информацию .
Галерея
[ редактировать ]- Сканирование изображений электронного микроскопа
 Диатомовая водоросль Surirella спиральная.
Диатомовая водоросль Surirella спиральная. Диатомовые водоросли Thalassiosira sp. на мембранном фильтре, размер пор 0,4 мкм.
Диатомовые водоросли Thalassiosira sp. на мембранном фильтре, размер пор 0,4 мкм. Диатомовая паралия бороздчатая .
Диатомовая паралия бороздчатая . Диатомовая водоросль Achanthes trinodis
Диатомовая водоросль Achanthes trinodis




 Автономная клетка Bacillaria paxillifer
Автономная клетка Bacillaria paxillifer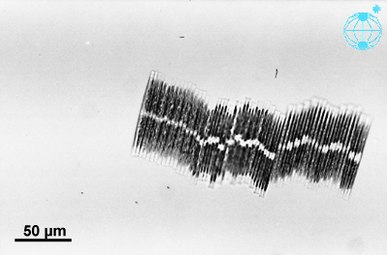 Колониальная группа Bacillaria paxillifer
Колониальная группа Bacillaria paxillifer


| Внешние видео | |
|---|---|
были отправлены три вида диатомей На Международную космическую станцию , в том числе огромные (длиной 6 мм) диатомовые водоросли Антарктиды и эксклюзивная колониальная диатомовая водоросль Bacillaria paradoxa . Клетки Bacillaria двигались рядом друг с другом частично, но в противоположной синхронности с помощью метода микрофлюидики . [116]
Эволюция и летопись окаменелостей
[ редактировать ]Источник
[ редактировать ]Гетероконтные хлоропласты, по-видимому, происходят из красных водорослей , а не непосредственно из прокариот , как это происходит у растений . Это говорит о том, что они имели более недавнее происхождение, чем многие другие водоросли. Однако ископаемые свидетельства скудны, и только с эволюцией самих диатомовых водорослей гетероконты производят серьезное впечатление в летописи окаменелостей.
Самые ранние окаменелости
[ редактировать ]Самые ранние известные ископаемые диатомеи датируются ранней юрой (около 185 млн лет назад ). [117] хотя молекулярные часы [117] и осадочные [118] данные свидетельствуют о более раннем происхождении. Высказано предположение, что их происхождение может быть связано с массовым вымиранием в конце перми многие морские ниши . (~250 млн лет назад), после которого открылись [119] Разрыв между этим событием и временем первого появления ископаемых диатомей может указывать на период, когда диатомеи были неокремненными и их эволюция была загадочной . [120] С момента появления окварцевания диатомовые водоросли оказали значительное влияние на летопись окаменелостей: основные залежи окаменелостей были обнаружены еще в раннем меловом периоде , а некоторые породы, такие как диатомовая земля , почти полностью состоят из них.
Отношение к лугам
[ редактировать ]Считается, что расширение лугов биомов и эволюционная радиация трав . во время миоцена увеличили приток растворимого кремния в океаны, и утверждается, что это способствовало развитию диатомовых водорослей в кайнозойскую эру [121] [122] Недавние исследования показывают, что успех диатомовых водорослей не связан с эволюцией трав, хотя разнообразие как диатомовых водорослей, так и пастбищ сильно увеличилось со среднего миоцена. [123]
Отношение к климату
[ редактировать ]Разнообразие диатомей в кайнозое было очень чувствительным к глобальной температуре, особенно к температурному градиенту экватора и полюса. В прошлом было показано, что более теплые океаны, особенно более теплые полярные регионы, имели значительно меньшее разнообразие диатомей. Будущие теплые океаны с усилением полярного потепления, как прогнозируется в сценариях глобального потепления, [124] Таким образом, теоретически это может привести к значительной потере разнообразия диатомей, хотя на основе современных знаний невозможно сказать, произойдет ли это быстро или только в течение многих десятков тысяч лет. [123]
Метод исследования
[ редактировать ]Летопись окаменелостей диатомовых водорослей в основном была установлена благодаря обнаружению их кремнистых панцирей в морских и неморских отложениях. Хотя диатомовые водоросли имеют как морскую, так и неморскую стратиграфическую летопись, биостратиграфия диатомей , основанная на ограниченном во времени эволюционном возникновении и исчезновении уникальных таксонов, хорошо развита и широко применима только в морских системах. Продолжительность ареала видов диатомовых водорослей была задокументирована посредством изучения океанских кернов и последовательностей горных пород, обнаженных на суше. [125] Там, где диатомовые биозоны хорошо установлены и откалиброваны по временной шкале геомагнитной полярности (например, Южный океан , северная часть Тихого океана , восточная экваториальная часть Тихого океана ), оценки возраста на основе диатомовых водорослей могут быть разрешены с точностью до <100 000 лет, хотя типичное разрешение возраста для кайнозойских комплексов диатомовых водорослей составляет несколько сотен тысяч лет.
широко используются для палеоэкологических реконструкций четвертичного Диатомеи, сохранившиеся в озерных отложениях , климата, особенно для озер закрытого бассейна, испытывающих колебания глубины и солености воды.
Изотопные записи
[ редактировать ]
Когда диатомовые водоросли умирают, их панцири ( панцири ) могут оседать на морском дне и превращаться в микроокаменелости . Со временем эти микрофоссилии погребены в виде отложений опалов в морских отложениях . Палеоклиматология – это изучение климата прошлого. Прокси-данные используются для того, чтобы связать элементы, собранные в современных образцах осадочных пород, с климатическими и океаническими условиями в прошлом. Заместители палеоклимата относятся к сохранившимся или окаменевшим физическим маркерам, которые заменяют прямые метеорологические или океанические измерения. [126] Примером прокси является использование изотопных записей диатомей δ13C , δ18O , δ30Si δ13C ( диатом , диатом δ18O и диатом δ30Si ). В 2015 году Суонн и Снеллинг использовали эти изотопные записи, чтобы задокументировать исторические изменения в условиях фототической зоны северо-западной части Тихого океана , включая снабжение питательными веществами и эффективность биологического насоса мягких тканей , от наших дней до стадии морских изотопов. 5е , что совпадает с последним межледниковьем . Пики продуктивности опалов на морской изотопной стадии связаны с нарушением региональной стратификации галоклина и увеличением поступления питательных веществ в фотическую зону . [127]

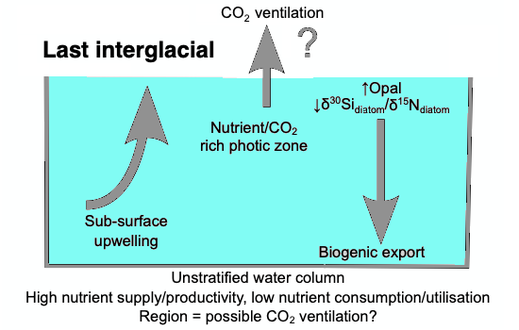
Первоначальное развитие галоклина и стратифицированной толщи воды было связано с началом крупного оледенения в Северном полушарии в 2,73 млн лет назад, которое увеличило приток пресной воды в регион за счет увеличения муссонных осадков и / или талой ледниковой воды, а также температуры поверхности моря . [128] [129] [130] [131] Связанное с этим уменьшение глубинного апвеллинга воды могло способствовать установлению более прохладных условий на глобальном уровне и расширению ледников по Северному полушарию с 2,73 млн лет назад. [129] Хотя галоклин, по-видимому, преобладал на протяжении позднеплиоценового и раннечетвертичного ледниково-межледниковых циклов , [132] другие исследования показали, что граница стратификации могла разрушиться в позднечетвертичное время при окончании ледников и в начале межледниковья. [133] [134] [135] [136] [137] [127]
Диверсификация
[ редактировать ]Меловые находки диатомей ограничены, но недавние исследования показывают постепенное разнообразие типов диатомей. Мел -палеогеновое вымирание , которое в океанах сильно повлияло на организмы с известковым скелетом, по-видимому, оказало относительно небольшое влияние на эволюцию диатомей. [138]
Оборот
[ редактировать ]не наблюдалось массового вымирания морских диатомей Хотя в кайнозое , времена относительно быстрого эволюционного оборота видовых комплексов морских диатомей происходили вблизи границы палеоцена и эоцена . [139] и на границе эоцена и олигоцена . [140] Дальнейший оборот комплексов происходил в разное время между средним миоценом и поздним плиоценом . [141] в ответ на прогрессирующее похолодание полярных регионов и развитие более эндемичных сообществ диатомей.
Глобальная тенденция к более тонким панцирям диатомей была отмечена от олигоцена до четвертичного периода . [125] Это совпадает со все более энергичной циркуляцией поверхности и глубинных вод океана, вызванной увеличением широтных температурных градиентов в начале значительного ледникового покрова расширения Антарктиды и прогрессирующим охлаждением в неогеновом и четвертичном периоде в направлении биполярного ледникового мира. Это заставило диатомовые водоросли поглощать меньше кремнезема для формирования панцирей. Повышенное перемешивание океанов обновляет кремнезем и другие питательные вещества, необходимые для роста диатомей в поверхностных водах, особенно в регионах прибрежного и океанического апвеллинга .
Генетика
[ редактировать ]
Выраженная маркировка последовательностей
[ редактировать ]В 2002 году первые сведения о свойствах репертуара генов Phaeodactylum tricornutum были описаны с использованием 1000 меток экспрессируемых последовательностей (EST). [142] Впоследствии количество EST было увеличено до 12 000, и была создана база данных EST диатомовых водорослей для функционального анализа. [143] Эти последовательности были использованы для сравнительного анализа P. tricornutum и предполагаемых полных протеомов зеленой водоросли Chlamydomonas Reinhardtii , красной водоросли Cyanidioschyzon merolae и диатомовой водоросли Thalassiosira pseudonana . [144] База данных EST диатомовых водорослей в настоящее время состоит из более чем 200 000 EST из клеток P. tricornutum (16 библиотек) и T. pseudonana (7 библиотек), выращенных в различных условиях, многие из которых соответствуют различным абиотическим стрессам. [145]
Секвенирование генома
[ редактировать ]
В 2004 году был секвенирован весь геном центрической диатомеи Thalassiosira pseudonana (32,4 Мб). [146] за ним в 2008 году последовало секвенирование пеннатной диатомеи Phaeodactylum tricornutum (27,4 МБ). [147] Сравнение этих двух геномов показывает, что геном P. tricornutum включает меньше генов (10 402 против 11 776), чем геном T. pseudonana ; между двумя геномами не удалось обнаружить никакой основной синтении (порядка генов). Гены T. pseudonana имеют в среднем ~1,52 интрона на ген по сравнению с 0,79 у P. tricornutum , что позволяет предположить недавнее широкое распространение интронов у центрических диатомей. [147] [148] Несмотря на относительно недавнее эволюционное расхождение (90 миллионов лет), степень молекулярного расхождения между центриками и пеннатами указывает на быстрые темпы эволюции внутри Bacillariophyceae по сравнению с другими группами эукариот . [147] Сравнительная геномика также установила, что определенный класс мобильных элементов , ретротранспозоны, подобные Diatom Copia (или CoDis), значительно амплифицирован в геноме P. tricornutum по отношению к T. pseudonana , составляя 5,8 и 1% соответствующих геномов. [149]
Эндосимбиотический перенос генов
[ редактировать ]Геномика диатомей дала много информации о масштабах и динамике процесса эндосимбиотического переноса генов (EGT). Сравнение белков T. pseudonana с гомологами других организмов показало, что сотни имеют ближайшие гомологи в линии Plantae. EGT по отношению к геномам диатомовых водорослей можно проиллюстрировать тем фактом, что геном T. pseudonana кодирует шесть белков, которые наиболее тесно связаны с генами, кодируемыми Guillardia theta ( криптомонада ) геномом нуклеоморфы . Четыре из этих генов также обнаружены в геномах пластид красных водорослей, что демонстрирует последовательную EGT от пластиды красных водорослей к ядру красных водорослей (нуклеоморфу) и к ядру гетероконта-хозяина. [146] Более поздние филогеномные анализы протеомов диатомовых водорослей предоставили доказательства существования празинофитоподобного эндосимбионта у общего предка хромальвеолатов , что подтверждается тем фактом, что 70% генов диатомовых водорослей происхождения Plantae имеют происхождение из зеленой линии и что такие гены также обнаружены в геноме. других страменопилей . Поэтому было высказано предположение, что хромальвеолаты являются продуктом серийного вторичного эндосимбиоза сначала с зелеными водорослями , а затем второго с красными водорослями , сохранившими геномные следы предыдущего, но вытеснившими зеленые пластиды. [150] Однако филогеномный анализ протеомов диатомей и истории эволюции хромальвеолатов, вероятно, будет использовать дополнительные геномные данные из недостаточно секвенированных линий, таких как красные водоросли.
Горизонтальный перенос генов
[ редактировать ]Помимо EGT, горизонтальный перенос генов (HGT) может происходить независимо от эндосимбиотического события. В публикации генома P. tricornutum сообщается, что по крайней мере 587 генов P. tricornutum, по-видимому, наиболее тесно связаны с бактериальными генами, составляя более 5% протеома P. tricornutum . Около половины из них также обнаружены в геноме T. pseudonana , что свидетельствует об их древнем включении в линию диатомовых водорослей. [147]
Генная инженерия
[ редактировать ]Чтобы понять биологические механизмы, лежащие в основе огромной важности диатомей в геохимических циклах, ученые использовали виды Phaeodactylum tricornutum и Thalassiosira spp. виды как модельные организмы с 90-х годов. [151] В настоящее время доступно несколько инструментов молекулярной биологии для создания мутантов или трансгенных линий: плазмиды, содержащие трансгены, встраиваются в клетки с использованием биолистического метода. [152] или транскоролевская бактериальная конъюгация [153] (с 10 −6 и 10 −4 выход соответственно [152] [153] ), и другие классические методы трансфекции, такие как электропорация или использование ПЭГ , как сообщается, дают результаты с более низкой эффективностью. [153]
Трансфицированные плазмиды могут быть либо случайным образом интегрированы в хромосомы диатомей, либо сохраняться в виде стабильных кольцевых эписом (благодаря центромерной последовательности дрожжей CEN6-ARSH4-HIS3). [153] ). Ген устойчивости к флеомицину/ зеоцину Sh Ble обычно используется в качестве маркера селекции. [151] [154] и различные трансгены были успешно внедрены и экспрессированы в диатомовых водорослях со стабильной передачей из поколения в поколение, [153] [154] или с возможностью его удаления. [154]
Кроме того, эти системы теперь позволяют использовать инструмент редактирования генома CRISPR-Cas , что приводит к быстрому производству функциональных нокаутных мутантов. [154] [155] и более точное понимание клеточных процессов диатомей.
Человеческое использование
[ редактировать ] Кизельгур, состоящий из центрических (радиально-симметричных) и пеннатных (двусторонне-симметричных) диатомей, взвешенных в воде.
Кизельгур, состоящий из центрических (радиально-симметричных) и пеннатных (двусторонне-симметричных) диатомей, взвешенных в воде.
(нажмите 3 раза, чтобы полностью увеличить)

Палеонтология
[ редактировать ]Разложение и распад диатомей приводит к образованию органических и неорганических (в виде силикатов ) осадков, неорганическая составляющая которых может привести к методу анализа прошлых морских сред по кернам океанского дна или илам заливов , поскольку неорганические вещества заключены в отложение глин и ила и образует постоянную геологическую летопись таких морских пластов (см. кремнистый ил ).
Промышленный
[ редактировать ]Диатомовые водоросли и их панцири (панцири), такие как диатомит или диатомит , являются важными промышленными ресурсами, используемыми для тонкой полировки и фильтрации жидкостей. Сложная структура их микроскопических оболочек была предложена в качестве материала для нанотехнологий. [156]
Диатомит считается природным наноматериалом и имеет множество применений и применений, таких как: производство различных керамических изделий, строительной керамики, огнеупорной керамики, специальной оксидной керамики, для производства материалов контроля влажности, используемых в качестве фильтрующего материала, материала в цементе. обрабатывающая промышленность, исходный материал для производства носителей лекарственных средств пролонгированного действия, абсорбирующий материал в промышленных масштабах, производство пористой керамики, стекольная промышленность, используется в качестве носителя катализатора, в качестве наполнителя в пластмассах и красках, очистка промышленных вод, держатель пестицидов, а также для улучшения физических и химических характеристик некоторых почв и других целей. [157] [158] [159]
Диатомовые водоросли также используются для определения происхождения содержащих их материалов, включая морскую воду.
Нанотехнологии
[ редактировать ]Отложение кремнезема диатомовыми водорослями также может оказаться полезным для нанотехнологий . [160] Клетки диатомовых водорослей многократно и надежно производят клапаны различных форм и размеров, что потенциально позволяет диатомовым водорослям создавать микро- или наноструктуры, которые могут быть использованы в ряде устройств, включая: оптические системы; полупроводниковая нанолитография ; и даже машины для доставки наркотиков . При соответствующей процедуре искусственного отбора диатомовые водоросли, производящие створки определенных форм и размеров, могут быть развиты для выращивания в хемостатных культурах для массового производства наноразмерных компонентов. [161] Также было предложено использовать диатомовые водоросли в качестве компонента солнечных элементов, заменив диоксид кремния светочувствительным диоксидом титана , который диатомовые водоросли обычно используют для создания клеточных стенок. [162] Также были предложены варианты производства солнечных панелей из диатомового биотоплива. [163]
- Поддержка и регулирование услуг, предоставляемых морскими диатомовыми водорослями, и некоторые их негативные последствия
 CNN = ядра конденсации облаков , DMS = диметилсульфид , DMSP = диметилсульфониопропионат , VOCs = летучие органические соединения.
CNN = ядра конденсации облаков , DMS = диметилсульфид , DMSP = диметилсульфониопропионат , VOCs = летучие органические соединения.
пунктирная стрелка: отрицательный эффект, сплошная стрелка: положительные эффекты.

судебно-медицинская экспертиза
[ редактировать ]Основная цель диатомового анализа в судебной медицине - отличить смерть в результате погружения от посмертного погружения тела в воду. Лабораторные исследования могут выявить наличие диатомей в организме. Поскольку скелеты диатомей на основе кремнезема трудно разлагаются, их иногда можно обнаружить даже в сильно разложившихся телах. Поскольку они не встречаются в организме естественным путем, если лабораторные анализы обнаруживают в трупе диатомовые водоросли того же вида, что и в воде, где было обнаружено тело, то это может быть убедительным доказательством утопления как причины смерти . Смесь видов диатомей, обнаруженная в трупе, может быть такой же или отличаться от окружающей воды, что указывает на то, утонул ли жертва в том же месте, где было найдено тело. [164]
История открытия
[ редактировать ]
Первые изображения диатомей встречаются в статье 1703 года в «Трудах Королевского общества», показывающей безошибочные рисунки табелларии . [165] Хотя автором публикации был неназванный английский джентльмен, недавно появились свидетельства того, что это был Чарльз Кинг Стаффордширский. [165] [166] Лишь 80 лет спустя мы находим первую официально идентифицированную диатомовую водоросль, колониальную Bacillaria paxillifera , открытую и описанную в 1783 году датским натуралистом Отто Фридрихом Мюллером . [165] Как и многие другие после него, он ошибочно полагал, что это животное из-за его способности двигаться. Даже Чарльз Дарвин видел останки диатомовых водорослей в пыли, находясь на островах Зеленого Мыса, хотя и не был уверен, что это были за частицы. Лишь позднее они были идентифицированы у него как кремнистые полигастрики. Инфузории, которые Дарвин позже заметил в краске на лице Фуэгинос, коренных жителей Огненной Земли на юге Южной Америки, позже были идентифицированы таким же образом. Еще при его жизни кремнистые многожелудки были выяснены как принадлежащие к диатомовым , и Дарвин изо всех сил пытался понять причины, лежащие в основе их красоты. Он обменялся мнениями по этой теме с известным криптогамистом Г.К. Туэйтсом. В четвертом издании «Происхождения видов» он заявил, что « немногие объекты прекраснее мельчайших кремнистых футляров диатомовых: были ли они созданы для того, чтобы их можно было рассматривать и восхищаться ими под мощным микроскопом »? и пришли к выводу, что их изысканные морфологии должны иметь функциональную основу, а не быть созданы исключительно для того, чтобы люди могли ими восхищаться. [167]
См. также
[ редактировать ]- Сильно разветвленные изопреноиды , длинноцепочечные алкены, вырабатываемые небольшим количеством морских диатомей.
Примечания
[ редактировать ]- ^ От греческого : διατομή , латинизированного : diatomé , «прорезание, разрыв», [7] от греческого : διάτομος , латинизированное : diátomos , «разрезанный пополам, разделенный поровну» [8] от греческого : διατέμνω , латинизированного : diatémno , «разрезать пополам». [9] [10] : 718
Ссылки
[ редактировать ]- ^ Дэнджерд, П. (1933). Traite d'Algologie. Поль Лехвалье и Филс, Париж, [1] Архивировано 4 октября 2015 года в Wayback Machine .
- ^ «Бацилляриофицеи» . ВОРМС . Всемирный реестр морских видов . 2024 . Проверено 9 мая 2024 г.
- ^ Дюмортье, Б.-К. (1822). Ботанические комментарии. Ботанические наблюдения, посвященные Садоводческому обществу Турне (PDF) . Турне: Imprimerie de Ch. Casterman-Dieu, Rue de Pont No. 10. С. [i], [1]-116, [1, табл., ошиб.] Архивировано из оригинала (PDF) 6 октября 2015 г. - через Algaebase.
- ^ Рабенхорст, Л. Европейская флора пресноводных и подводных водорослей (1864–1868). Раздел I. «Диатомовые водоросли» с изображениями всех родов, напечатанными на дереве (1864 г.). стр. 1–359. Лейпциг [Лейпциг]: У Эдуарда Куммерума.
- ^ Геккель, Э. (1878). Das Protistenreich. Архивировано 10 ноября 2014 года в Wayback Machine .
- ^ Энглер, А. и Гилг, Э. (1919). Программа семейств растений: обзор всей растительной системы с особым вниманием к лекарственным и полезным растениям, а также обзор цветочных царств и цветочных регионов мира для использования в лекциях и исследованиях по специальной и медико-фармацевтической ботанике. , 8-е изд., Gebrüder Borntraeger Verlag, Берлин, 395 стр.
- ^ διατομή . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ διάτομος . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ διατέμνω . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Компактный Оксфордский словарь английского языка . Кларендон Пресс. 1971. ISBN 0918414083 .
- ^ «Воздух, которым вы дышите? Это сделала диатомовая водоросль» . Живая наука . 11 июня 2014 года. Архивировано из оригинала 30 апреля 2018 года . Проверено 30 апреля 2018 г.
- ^ «Что такое диатомовые водоросли?» . Диатомовые водоросли Северной Америки. Архивировано из оригинала 25 января 2020 года . Проверено 28 января 2020 г.
- ^ Трегер, П.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Лейнарт, А.; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука . 268 (5209): 375–9. Бибкод : 1995Sci...268..375T . дои : 10.1126/science.268.5209.375 . ПМИД 17746543 . S2CID 5672525 .
- ^ «Королевский колледж Лондона – озеро Мегачад» . www.kcl.ac.uk. Архивировано из оригинала 27 ноября 2018 года . Проверено 5 мая 2018 г.
- ^ Бристоу, CS; Хадсон-Эдвардс, Калифорния ; Чаппелл, А. (2010). «Удобрение Амазонки и экваториальной Атлантики западноафриканской пылью». Геофиз. Рез. Летт . 37 (14): L14807. Бибкод : 2010GeoRL..3714807B . дои : 10.1029/2010GL043486 . S2CID 128466273 .
- ^ Перейти обратно: а б с д и ж Хасл, Грета Р.; Сиверцен, Эрик Э.; Штайдингер, Карен А.; Танген, Карл (25 января 1996 г.). «Морские диатомеи» . В Томасе, Кармело Р. (ред.). Определение морских диатомей и динофлагеллят . Академическая пресса. стр. 5–385. ISBN 978-0-08-053441-1 . Архивировано из оригинала 3 января 2014 года . Проверено 13 ноября 2013 г.
- ^ «Газовые пожиратели» . Архивировано из оригинала 22 мая 2018 года . Проверено 22 мая 2018 г.
- ^ «Подробнее о диатомовых водорослях» . Музей палеонтологии Калифорнийского университета . Архивировано из оригинала 4 октября 2012 года . Проверено 20 мая 2015 г.
- ^ ван ден Хук, К.; Манн, Д.Г.; Янс, Х.М. (1995). Водоросли: введение в психологию . Кембридж: Издательство Кембриджского университета . стр. 165–218. ISBN 978-0-521-31687-3 .
- ^ Накаяма, Т.; Камикава, Р.; Танифудзи, Г.; Касияма, Ю.; Окоучи, Н.; Арчибальд, Дж. М.; Инагаки, Ю. (2014). «Полный геном нефотосинтетической цианобактерии диатомовой водоросли демонстрирует недавнюю адаптацию к внутриклеточному образу жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (31): 11407–11412. Бибкод : 2014PNAS..11111407N . дои : 10.1073/pnas.1405222111 . ПМК 4128115 . ПМИД 25049384 .
- ^ Пьерелла Карлушич, Хуан Хосе; Пеллетье, Эрик; Ломбард, Фабьен; Карсик, Мэдлин; Дворжак, Этьен; Колен, Себастьен; Пишераль, Марк; Корнехо-Кастильо, Франсиско М.; Ацинас, Сильвия Г.; Пепперкок, Райнер; Карсенти, Эрик (6 июля 2021 г.). «Глобальные закономерности распределения морских азотфиксаторов по данным визуализации и молекулярным методам» . Природные коммуникации . 12 (1): 4160. Бибкод : 2021NatCo..12.4160P . дои : 10.1038/s41467-021-24299-y . ISSN 2041-1723 . ПМК 8260585 . ПМИД 34230473 .
- ^ Функциональная связь между хозяином динофлагелляты и его диатомовым эндосимбионтом | Молекулярная биология и эволюция | Оксфорд Академик
- ^ Вер, доктор медицинских наук; Оболочка, Р.Г.; Кочиолек, JP, ред. (2015). Пресноводные водоросли Северной Америки: экология и классификация (2-е изд.). Сан-Диего: Академическая пресса. ISBN 978-0-12-385876-4 .
- ^ Жирар, Винсент; Сен-Мартен, Симона; Баффето, Эрик; Сен-Мартен, Жан-Поль; Неродо, Дидье; Пейро, Даниэль; Роги, Гвидо; Рагацци, Эухенио; Сутитхорн, Варавуд (2020). «Тайский янтарь: понимание ранней истории диатомовых водорослей?» . BSGF — Бюллетень наук о Земле . 191 : 23. дои : 10.1051/bsgf/2020028 . ISSN 1777-5817 .
- ^ Внутреннее пространство субарктического Тихого океана. Архивировано 27 октября 2020 года в на Wayback Machine земных экспедициях НАСА , 4 сентября 2018 года.
 В данную статью включен текст из этого источника, находящегося в свободном доступе .
В данную статью включен текст из этого источника, находящегося в свободном доступе . - ^ Руссо, Сесиль С.; Грегг, Уотсон В. (2015). «Последние десятилетние тенденции в глобальном составе фитопланктона» . Глобальные биогеохимические циклы . 29 (10): 1674–1688. Бибкод : 2015GBioC..29.1674R . дои : 10.1002/2015GB005139 .
- ^ Колин С., Коэльо Л.П., Сунагава С., Боулер К., Карсенти Э., Борк П., Пепперкок Р. и Де Варгас К. (2017) «Количественная 3D-визуализация клеток биология и экология экологических микробных эукариот». eLife , 6 : e26066. два : 10.7554/eLife.26066.002 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine . - ^ Нельсон, Дэвид М.; Трегер, Поль; Бжезинский, Марк А.; Лейнарт, Од; Кегинер, Бернар (1995). «Производство и растворение биогенного кремнезема в океане: пересмотренные глобальные оценки, сравнение с региональными данными и связь с биогенным седиментированием». Глобальные биогеохимические циклы . 9 (3). Американский геофизический союз (AGU): 359–372. Бибкод : 1995GBioC...9..359N . дои : 10.1029/95gb01070 . ISSN 0886-6236 .
- ^ Манн, Дэвид Г. (1999). «Видовая концепция диатомовых водорослей». Психология . 38 (6). Информа Великобритания Лимитед: 437–495. Бибкод : 1999Phyco..38..437M . дои : 10.2216/i0031-8884-38-6-437.1 . ISSN 0031-8884 .
- ^ Симонсен, Р., (1979). «Диатомовая система: идеи о филогении», Bacillaria , 2 : 9–71.
- ^ Перейти обратно: а б Мур, Эрик Р.; Буллингтон, Бриана С.; Вайсберг, Александра Дж.; Цзян, Юань; Чанг, Джефф; Хэлси, Кимберли Х. (7 июля 2017 г.). «Морфологические и транскриптомные доказательства индукции аммонием полового размножения у Thalassiosira pseudonana и других центрических диатомей» . ПЛОС ОДИН . 12 (7). Публичная научная библиотека (PLoS): e0181098. Бибкод : 2017PLoSO..1281098M . дои : 10.1371/journal.pone.0181098 . ISSN 1932-6203 . ПМК 5501676 . ПМИД 28686696 . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Университетский колледж Лондона (2002) диатомей Отдел микропалеонтологии .
- ^ Паркер, Эндрю Р.; Таунли, Хелен Э. (2007). «Биомиметика фотонных наноструктур». Природные нанотехнологии . 2 (6): 347–53. Бибкод : 2007NatNa...2..347P . дои : 10.1038/nnano.2007.152 . ПМИД 18654305 .
- ^ Гордон, Ричард; Лосич, Душан; Тиффани, Мэри Энн; Надь, Стивен С.; Стерренбург, Фритьоф А.С. (2009). «Стеклянный зверинец: диатомовые водоросли для новых применений в нанотехнологиях». Тенденции в биотехнологии . 27 (2): 116–27. дои : 10.1016/j.tibtech.2008.11.003 . ПМИД 19167770 .
- ^ Перейти обратно: а б с д Рита А. Хорнер (2002). Таксономический справочник некоторых распространенных морских фитопланктонов . Биопресс. стр. 25–30. ISBN 978-0-948737-65-7 . Архивировано из оригинала 1 августа 2020 года . Проверено 13 ноября 2013 г.
- ^ «Стекло в природе» . Корнингский музей стекла. Архивировано из оригинала 7 марта 2013 года . Проверено 19 февраля 2013 г.
- ^ Тейлор, Дж. К., Хардинг, В. Р. и Арчибальд, К. (2007). Иллюстрированный путеводитель по некоторым распространенным видам диатомей из Южной Африки . Гезина: Комиссия по водным исследованиям. ISBN 9781770054844 .
- ^ Мишра М., Аруха А.П., Башир Т., Ядав Д. и Прасад ГБКС (2017) «Все новые лица диатомовых водорослей: потенциальный источник наноматериалов и не только». Границы микробиологии , 8 : 1239. два : 10.3389/fmicb.2017.01239 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine .
- ^ Перейти обратно: а б Чжан, Д.; Ван, Ю.; Кай, Дж.; Пан, Дж.; Цзян, X.; Цзян, Ю. (2012). «Технология биопроизводства на основе диатомовой микро- и наноструктуры» . Китайский научный бюллетень . 57 (30): 3836–3849. Бибкод : 2012ЧСБу..57.3836З . дои : 10.1007/s11434-012-5410-x .
- ^ «Диатомовые водоросли» . Архивировано из оригинала 2 февраля 2016 года . Проверено 13 февраля 2016 г.
- ^ Молекулярная жизнь диатомей
- ^ Килиас, Эстель С.; Юнгес, Леандро; Шупраха, Лука; Леонард, Гай; Метфис, Катя; Ричардс, Томас А. (2020). «Распространение хитридиевых грибов и их совместное присутствие с диатомовыми водорослями коррелируют с таянием морского льда в Северном Ледовитом океане» . Коммуникационная биология . 3 (1): 183. дои : 10.1038/s42003-020-0891-7 . ПМЦ 7174370 . ПМИД 32317738 . S2CID 216033140 .
- ^ Перейти обратно: а б с Таматракольн, К.; Алверсон, Эй Джей; Хильдебранд, М. (2006). «Сравнительный анализ последовательностей диатомовых переносчиков кремния: к механистической модели транспорта кремния». Журнал психологии . 42 (4): 822–834. Бибкод : 2006JPcgy..42..822T . дои : 10.1111/j.1529-8817.2006.00233.x . S2CID 86674657 .
- ^ Крегер, Нильс; Дойцманн, Райнер; Манфред, Сампер (ноябрь 1999 г.). «Поликатионные пептиды из диатомового биокремнезема, которые направляют образование кремнеземной наносферы». Наука . 286 (5442): 1129–1132. дои : 10.1126/science.286.5442.1129 . ПМИД 10550045 . S2CID 10925689 . [ постоянная мертвая ссылка ]
- ^ Крогер, Нильс (2007). Справочник по биоминерализации: биологические аспекты и формирование структуры . Вайнхайм, Германия: Wiley-VCH Verlag GmbH. стр. глава 3.
- ^ Аноним (1702 г.). «Два письма от деревенского джентльмена, касающиеся сделки г-на Левенгука, № 283», Philos. Пер. Р. Сок. Лонд. Б , 23 : 1494–1501.
- ^ Рогалл, Э. (1939). « О тонкой структуре кремнеземной мембраны диатомей» , Планта : 279-291.
- ^ Перейти обратно: а б с Хильдебранд, Марк; Лерч, Сара Дж.Л.; Шреста, Рошан П. (11 апреля 2018 г.). «Понимание окварцевания клеточной стенки диатомовых водорослей — движение вперед» . Границы морской науки . 5 . Фронтирс Медиа С.А. дои : 10.3389/fmars.2018.00125 . ISSN 2296-7745 . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хильдебранд, Марк; Вулкани, Бенджамин Э.; Гассманн, Вальтер; Шредер, Джулиан И. (1997). «Семейство генов переносчиков кремния». Природа . 385 (6618). ООО «Спрингер Сайенс энд Бизнес Медиа»: 688–689. Бибкод : 1997Natur.385..688H . дои : 10.1038/385688b0 . ISSN 0028-0836 . ПМИД 9034185 . S2CID 4266966 .
- ^ Крегер, Нильс; Дойцманн, Райнер; Сампер, Манфред (5 ноября 1999 г.). «Поликатионные пептиды из диатомового биокремнезема, которые направляют образование кремнеземной наносферы». Наука . 286 (5442). Американская ассоциация содействия развитию науки (AAAS): 1129–1132. дои : 10.1126/science.286.5442.1129 . ISSN 0036-8075 . ПМИД 10550045 .
- ^ Крегер, Нильс; Дойцманн, Райнер; Бергсдорф, Кристиан; Сампер, Манфред (5 декабря 2000 г.). «Видоспецифические полиамины диатомовых водорослей контролируют морфологию кремнезема» . Труды Национальной академии наук . 97 (26): 14133–14138. Бибкод : 2000PNAS...9714133K . дои : 10.1073/pnas.260496497 . ISSN 0027-8424 . ЧВК 18883 . ПМИД 11106386 .
- ^ Тессон, Бенуа; Хильдебранд, Марк (10 декабря 2010 г.). «Обширная и тесная связь цитоскелета с образованием кремнезема у диатомовых водорослей: контроль над формированием рисунка на мезо- и микромасштабе» . ПЛОС ОДИН . 5 (12). Публичная научная библиотека (PLoS): e14300. Бибкод : 2010PLoSO...514300T . дои : 10.1371/journal.pone.0014300 . ISSN 1932-6203 . ПМК 3000822 . ПМИД 21200414 .
- ^ Тессон, Бенуа; Хильдебранд, Марк (23 апреля 2013 г.). «Характеристика и локализация нерастворимых органических матриц, связанных со стенками клеток диатомей: понимание их роли в формировании клеточной стенки» . ПЛОС ОДИН . 8 (4). Публичная научная библиотека (PLoS): e61675. Бибкод : 2013PLoSO...861675T . дои : 10.1371/journal.pone.0061675 . ISSN 1932-6203 . ПМЦ 3633991 . ПМИД 23626714 .
- ^ Шеффель, Андре; Поульсен, Николь; Шиан, Сэмюэл; Крегер, Нильс (7 февраля 2011 г.). «Белковые микрокольца с наноструктурой из диатомовой водоросли, которые управляют морфогенезом кремнезема» . Труды Национальной академии наук . 108 (8): 3175–3180. дои : 10.1073/pnas.1012842108 . ISSN 0027-8424 . ПМК 3044418 . ПМИД 21300899 .
- ^ Коч, Александр; Грегер, Филип; Павольски, Дамиан; Боманс, Пол Х.Х.; Соммердейк, Нико AJM; Шлирф, Майкл; Крегер, Нильс (24 июля 2017 г.). «Силиканин-1 представляет собой консервативный мембранный белок диатомовой водоросли, участвующий в биоминерализации кремнезема» . БМК Биология . 15 (1). Springer Science and Business Media LLC: 65. doi : 10.1186/s12915-017-0400-8 . ISSN 1741-7007 . ПМЦ 5525289 . ПМИД 28738898 .
- ^ Тессон, Бенуа; Лерч, Сара Дж.Л.; Хильдебранд, Марк (18 октября 2017 г.). «Характеристика нового семейства белков, связанного с мембраной везикул отложения кремнезема, позволяет генетически манипулировать диатомовым кремнеземом» . Научные отчеты . 7 (1). Springer Science and Business Media LLC: 13457. Бибкод : 2017NatSR...713457T . дои : 10.1038/s41598-017-13613-8 . ISSN 2045-2322 . ПМЦ 5647440 . ПМИД 29044150 .
- ^ Падисак, Юдит; Сорочки-Пинтер, Ева; Резнер, Жужанна (2003), Мартенс, Коэн (редактор), «Свойство погружения некоторых форм фитопланктона и связь устойчивости формы с морфологическим разнообразием планктона - экспериментальное исследование» (PDF) , Водное биоразнообразие: праздничный том в честь Анри Дж. Дюмона , «Развития в гидробиологии», Springer Нидерланды, стр. 243–257, doi : 10.1007/978-94-007-1084-9_18 , ISBN 9789400710849 , заархивировано из оригинала (PDF) 23 июля 2018 г. , получено 4 октября 2019 г.
- ^ Андерсон, Ларс В.Дж.; Суини, Беатрис М. (1 мая 1977 г.). «Дневные изменения характеристик седиментации Ditylum Brightwelli: изменения клеточных липидов и влияние ингибиторов дыхания и модификаторов ионного транспорта1» . Лимнология и океанография . 22 (3): 539–552. Бибкод : 1977LimOc..22..539A . дои : 10.4319/lo.1977.22.3.0539 . ISSN 1939-5590 .
- ^ Поулсен, Николь С.; Спектор, Илан; Сперк, Тимоти П.; Шульц, Томас Ф.; Уэтерби, Ричард (1 сентября 1999 г.). «Скольжение диатомей является результатом системы подвижности актин-миозина». Подвижность клеток и цитоскелет . 44 (1): 23–33. doi : 10.1002/(SICI)1097-0169(199909)44:1<23::AID-CM2>3.0.CO;2-D . ISSN 1097-0169 . ПМИД 10470016 .
- ^ Манн, Дэвид Г. (февраль 2010 г.). «Рафидные диатомеи» . Веб-проект «Древо жизни» . Архивировано из оригинала 27 сентября 2019 года . Проверено 27 сентября 2019 г.
- ^ Перейти обратно: а б Г. Дребес (1 января 1977 г.). «Глава 9: Сексуальность» . У Дитриха Вернера (ред.). Биология диатомей . Ботанические монографии. Том. 13. Издательство Калифорнийского университета. стр. 250–283. ISBN 978-0-520-03400-6 . Архивировано из оригинала 1 августа 2020 года . Проверено 14 ноября 2013 г.
- ^ Льюис, WM-младший (1984). «Половые часы диатомей и их эволюционное значение». Американский натуралист , 123 (1): 73–80.
- ^ Чепурнов В.А., Манн Д.Г., Саббе К., Выверман В. (2004). «Экспериментальные исследования полового размножения диатомовых водорослей». В Чон К.В., изд. Обзор клеточной биологии . Международное обозрение цитологии 237 : 91–154. Лондон.
- ^ Дребес (1977) «Сексуальность». В: Вернер Д., изд. Биология диатомей , Ботанические монографии 13 250–283. Оксфорд: Научные публикации Блэквелла.
- ^ «Уменьшение размера, репродуктивная стратегия и жизненный цикл центрической диатомеи». Философские труды Лондонского королевского общества. Серия Б: Биологические науки . 336 (1277). Королевское общество: 191–213. 29 мая 1992 г. doi : 10.1098/rstb.1992.0056 . ISSN 0962-8436 . S2CID 86332060 .
- ^ Кестер, Джули А.; Броули, Сьюзен Х.; Карп-Босс, Ли; Манн, Дэвид Г. (2007). «Половое размножение морской центрической диатомовой водоросли Ditylum Brightwellii (Bacillariophyta)» . Европейский журнал психологии . 42 (4). Информа UK Limited: 351–366. Бибкод : 2007EJPhy..42..351K . дои : 10.1080/09670260701562100 . ISSN 0967-0262 . S2CID 80737380 .
- ^ Перейти обратно: а б Муже Ж.Л., Гастино Р., Давидович О., Годен П., Давидович Н.А. Свет является ключевым фактором, запускающим половое размножение пеннатной диатомовой водоросли Haslea ostrearia. ФЭМС Микробиол Экол. Август 2009 г.;69(2):194-201. doi: 10.1111/j.1574-6941.2009.00700.x. Epub 2009, 6 мая. PMID 19486155.
- ^ Скалько Э., Стек К., Иудикон Д., Ферранте М.И., Монтрезор М. Динамика половой фазы морской диатомовой водоросли Pseudo-nitzschia multistriata (Bacillariophyceae). Дж. Фикол. Октябрь 2014 г.;50(5):817-28. дои: 10.1111/jpy.12225. Epub, 14 сентября 2014 г. PMID 26988637.
- ^ Перейти обратно: а б Патил С., Мойс С., фон Дассов П., Гюисман М.Дж., Мейплсон Д., Де Вейлдер Л., Санжес Р., Виверман В., Монтрезор М., Ферранте М.И. Идентификация набора мейотических инструментов у диатомовых водорослей и исследование специфичных для мейоза гомологов SPO11 и RAD51 у половых видов Pseudo-nitzschia multistriata и Seminavis Robusta. БМК Геномика. 2015, 14 ноября; 16: 930. дои: 10.1186/s12864-015-1983-5. Опечатка в: BMC Genomics. 5 июля 2019 г.; 20 (1): 544. ПМИД 26572248; PMCID: PMC4647503
- ^ Азам, Фарук; Бидл, Кей Д. (1999). «Ускоренное растворение диатомового кремнезема морскими бактериальными комплексами». Природа . 397 (6719): 508–12. Бибкод : 1999Natur.397..508B . дои : 10.1038/17351 . S2CID 4397909 . ИНИСТ 1755031 .
- ^ Захарова Юлия Р.; Галачанц Юрий П.; Курилкина Мария Ивановна; Лихошвай, Александр Васильевич; Петрова Дарья П.; Шишлянников Сергей М.; Равин, Николай В.; Марданов Андрей Владимирович; Белецкий Алексей Владимирович; Лихошвей, Елена Владимировна (2013). «Структура микробного сообщества и деградация диатомей глубокого придонного слоя озера Байкал» . ПЛОС ОДИН . 8 (4): e59977. Бибкод : 2013PLoSO...859977Z . дои : 10.1371/journal.pone.0059977 . ПМК 3613400 . ПМИД 23560063 .
- ^ Перейти обратно: а б Эгге, Дж. К.; Акснес, Д.Л. (1992). «Силикат как регулирующее питательное вещество в конкуренции фитопланктона» . Мар Экол. Прог. Сер . 83 : 281–289. Бибкод : 1992MEPS...83..281E . дои : 10.3354/meps083281 .
- ^ Перейти обратно: а б с Фурнас, Майлз Дж. (1990). «Темпы роста морского фитопланктона in situ: подходы к измерению, темпы роста сообществ и видов». Журнал исследований планктона . 12 (6): 1117–51. дои : 10.1093/планкт/12.6.1117 . ИНИСТ 5474600 .
- ^ Перейти обратно: а б Юл, Эндрю; Тиррелл, Тоби (2003). «Роль диатомовых водорослей в регулировании круговорота кремния в океане». Глобальные биогеохимические циклы . 17 (4): н/д. Бибкод : 2003GBioC..17.1103Y . CiteSeerX 10.1.1.394.3912 . дои : 10.1029/2002GB002018 . S2CID 16849373 .
- ^ Мальвия, Шрути; Скалько, Элеонора; Аудик, Стефан; Винсент, Флора; Велучами, Алагурадж; Пулен, Жюли; Винкер, Патрик; Иудиконе, Даниэле; де Варгас, Колумбан; Биттнер, Люси; Зингоне, Адриана; Боулер, Крис (29 февраля 2016 г.). «Понимание глобального распределения и разнообразия диатомовых водорослей в мировом океане» . Труды Национальной академии наук . 113 (11): Е1516–Е1525. Бибкод : 2016PNAS..113E1516M . дои : 10.1073/pnas.1509523113 . ISSN 0027-8424 . ПМК 4801293 . ПМИД 26929361 .
- ^ Липпс, Джер Х. (1970). «Эволюция планктона». Эволюция . 24 (1): 1–22. дои : 10.2307/2406711 . JSTOR 2406711 . ПМИД 28563010 .
- ^ «Дидимо, пришельцы среди нас» . Архивировано 7 октября 2015 года на Wayback Machine виртуальной выставке Виртуального музея Канады.
- ^ «DEP сообщает, что Дидимо обнаружен в западном отделении реки Фармингтон» . Архивировано из оригинала 16 февраля 2015 года . Проверено 27 апреля 2015 г.
- ^ «Информация для заинтересованных сторон Didymo – 31 октября 2008 г.» . MAF Biosecurity Новая Зеландия http://www.biosecurity.govt.nz . Архивировано из оригинала 12 февраля 2013 года . Проверено 1 декабря 2013 г.
{{cite web}}: Внешняя ссылка в|publisher= - ^ Перейти обратно: а б Дагдейл, Колорадо; Вилкерсон, ФП (1998). «Силикатное регулирование новой продукции в экваториальном тихоокеанском апвеллинге». Природа . 391 (6664): 270–273. Бибкод : 1998Natur.391..270D . дои : 10.1038/34630 . S2CID 4394149 .
- ^ Сметачек, В.С. (1985). «Роль погружения в жизненных циклах диатомовых водорослей: экологическое, эволюционное и геологическое значение». Мар Биол . 84 (3): 239–251. Бибкод : 1985MarBi..84..239S . дои : 10.1007/BF00392493 . S2CID 85054779 .
- ^ Перейти обратно: а б Трегер, П.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Лейнарт, А.; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука . 268 (5209): 375–9. Бибкод : 1995Sci...268..375T . дои : 10.1126/science.268.5209.375 . ПМИД 17746543 . S2CID 5672525 .
- ^ Рэйвен, Дж. А. (1983). «Транспорт и функция кремния в растениях». Биол. Преподобный . 58 (2): 179–207. дои : 10.1111/j.1469-185X.1983.tb00385.x . S2CID 86067386 .
- ^ Миллиган, Эй Джей; Морель, ФММ (2002). «Роль протонного буфера для кремнезема в диатомовых водорослях». Наука . 297 (5588): 1848–1850. Бибкод : 2002Sci...297.1848M . дои : 10.1126/science.1074958 . ПМИД 12228711 . S2CID 206507070 .
- ^ Перейти обратно: а б Чемберлен, CJ (1901) Методы гистологии растений , Издательство Чикагского университета.
- ^ Р. Сивер; Стивен Генри Шнайдер; Пенелопа Дж. Бостон (январь 1993 г.). «Кремнезем в океанах: биолого-геологическое взаимодействие» . Ученые на Гайе . МТИ Пресс . стр. 287–295 . ISBN 978-0-262-69160-4 . Проверено 14 ноября 2013 г.
- ^ Киддер, Дэвид Л.; Эрвин, Дуглас Х. (2001). «Вековое распределение биогенного кремнезема в фанерозое: сравнение кремнезамещенных окаменелостей и слоистых кремней на уровне серий». Журнал геологии . 109 (4): 509–22. Бибкод : 2001JG....109..509K . дои : 10.1086/320794 . S2CID 128401816 .
- ^ Гренн, Тор; Слэк, Джон Ф. (2003). «Палеозойская и мезозойская морская вода, богатая кремнеземом: данные по отложениям гематитового кремня (яшмы)». Геология . 31 (4): 319–22. Бибкод : 2003Geo....31..319G . doi : 10.1130/0091-7613(2003)031<0319:PAMSRS>2.0.CO;2 . ИНИСТ 14692468 .
- ^ Раки, Г; Корди, Фабрис (2000). «Палеоэкология радиолярий и радиоляриты: является ли настоящее ключом к прошлому?». Обзоры наук о Земле . 52 (1): 83–120. Бибкод : 2000ESRv...52...83R . дои : 10.1016/S0012-8252(00)00024-6 .
- ^ Мальдонадо, Мануэль; Кармона, М. Кармен; Урис, Мария Х.; Крузадо, Антонио (1999). «Уменьшение количества мезозойских губок, строивших рифы, объясняется ограничением кремния». Природа . 401 (6755): 785–8. Бибкод : 1999Natur.401..785M . дои : 10.1038/44560 . S2CID 205034177 . ИНИСТ 1990263 .
- ^ Харпер, Ховард Э.; Нолл, Эндрю Х. (1975). «Кремнезем, диатомовые водоросли и кайнозойская эволюция радиолярий». Геология . 3 (4): 175–177. Бибкод : 1975Geo.....3..175H . doi : 10.1130/0091-7613(1975)3<175:SDACRE>2.0.CO;2 .
- ^ Перейти обратно: а б Пьерелла Карлушич, Хуан Хосе; Боулер, Крис; Бисвас, Хайманти (30 апреля 2021 г.). «Механизмы концентрации углекислого газа в природных популяциях морских диатомовых водорослей: данные из океанов Тары» . Границы в науке о растениях . 12 . Frontiers Media SA: 657821. doi : 10.3389/fpls.2021.657821 . ISSN 1664-462X . ПМЦ 8119650 . ПМИД 33995455 . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Фридлингштейн, Пьер; О'Салливан, Майкл; Джонс, Мэтью В.; Эндрю, Робби М.; и др. (11 декабря 2020 г.). «Глобальный углеродный бюджет 2020» . Данные науки о системе Земли . 12 (4). Коперник ГмбХ: 3269–3340. Бибкод : 2020ESSD...12.3269F . doi : 10.5194/essd-12-3269-2020 . hdl : 10871/126892 . ISSN 1866-3516 .
- ^ Перейти обратно: а б Глиберт, Патрисия М.; Вилкерсон, Фрэнсис П.; Дагдейл, Ричард С.; Рэйвен, Джон А.; Дюпон, Кристофер Л.; Ливитт, Питер Р.; и др. (11 октября 2015 г.). «Плюсы и минусы поглощения и ассимиляции аммония и нитратов фитопланктоном и последствия для продуктивности и состава сообщества с упором на условия, обогащенные азотом». Лимнология и океанография . 61 (1). Уайли: 165–197. дои : 10.1002/lno.10203 . ISSN 0024-3590 . S2CID 16561857 . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Аллен, Эндрю Э.; Дюпон, Кристофер Л.; Оборник, Мирослав; Горак, Алеш; Нуньес-Неси, Адриано; МакКроу, Джон П.; и др. (2011). «Эволюция и метаболическое значение цикла мочевины у фотосинтезирующих диатомей» . Природа . 473 (7346): 203–7. Бибкод : 2011Natur.473..203A . дои : 10.1038/nature10074 . ПМИД 21562560 . S2CID 4350884 . Архивировано из оригинала 19 февраля 2022 года . Проверено 20 мая 2021 г.
- «Животный цикл мочевины в крошечных диатомовых водорослях океана позволяет морскому фитопланктону использовать углерод и азот из окружающей среды» . ScienceDaily (пресс-релиз). 12 мая 2011 г.
- ^ Перейти обратно: а б Армбруст, Э. Вирджиния; Берджес, Джон А.; Боулер, Крис; Грин, Беверли Р.; Мартинес, Диего; Патнэм, Николас Х.; и др. (2004). «Геном диатомовой водоросли Thalassiosira Pseudonana: экология, эволюция и метаболизм» . Наука . 306 (5693). Американская ассоциация развития науки (AAAS): 79–86. Бибкод : 2004Sci...306...79A . дои : 10.1126/science.1101156 . ISSN 0036-8075 . ПМИД 15459382 . S2CID 8593895 .
- ^ Перейти обратно: а б с Аллен, Эндрю Э.; Дюпон, Кристофер Л.; Оборник, Мирослав; Горак, Алеш; Нуньес-Неси, Адриано; МакКроу, Джон П.; и др. (2011). «Эволюция и метаболическое значение цикла мочевины у фотосинтезирующих диатомей» . Природа . 473 (7346). ООО «Спрингер Сайенс энд Бизнес Медиа»: 203–207. Бибкод : 2011Natur.473..203A . дои : 10.1038/nature10074 . ISSN 0028-0836 . ПМИД 21562560 . S2CID 4350884 .
- ^ Вейман, Филип Д.; Бири, Карен; Лефевр, Стефан К.; Ривера, Хосефа; Маккарти, Джеймс К.; Хойбергер, Адам Л.; и др. (10 октября 2014 г.). «Инактивация гена уреазы Phaeodactylum tricornutum с использованием целевого мутагенеза на основе эффекторной нуклеазы, подобной активатору транскрипции» . Журнал биотехнологии растений . 13 (4). Уайли: 460–470. дои : 10.1111/pbi.12254 . ISSN 1467-7644 . ПМИД 25302562 .
- ^ Бендер, Сара Дж.; Паркер, Микаэла С.; Армбруст, Э. Вирджиния (2012). «Совместное влияние света и источника азота на цикл мочевины и метаболизм азота в течение цикла Диеля у морской диатомовой водоросли Thalassiosira pseudonana». Протист . 163 (2). Эльзевир Б.В.: 232–251. дои : 10.1016/j.protis.2011.07.008 . ISSN 1434-4610 . ПМИД 21873112 .
- ^ Армстронг, Э; Роджерсон, А; Левтли, младший (2000). «Использование углерода морских водорослей тремя гетеротрофными протистами, связанными с поверхностью: Stereomyxa ramosa, Nitzschia alba и Labyrinthula sp» . Водная микробная экология . 21 : 49–57. дои : 10.3354/ame021049 . ISSN 0948-3055 .
- ^ Левин, Джойс; Левин, Р.А. (1967). «Культурирование и питание некоторых апохлоротичных диатомей рода Nitzschia» . Микробиология . 46 (3): 361–367. дои : 10.1099/00221287-46-3-361 . ISSN 1350-0872 .
- ^ Камп, А.; Де Бир, Д.; Нитч, Дж.Л.; Лавик, Г.; Стиф, П. (2011). «Диатомовые водоросли дышат нитратами, чтобы выжить в темноте и бескислородных условиях» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (14): 5649–5654. Бибкод : 2011PNAS..108.5649K . дои : 10.1073/pnas.1015744108 . ПМК 3078364 . ПМИД 21402908 .
- ^ Мерц, Элиза; Дик, Грегори Дж.; Пиво, Дирк; Грим, Шэрон; Хюбенер, Томас; Литтманн, Стен; и др. (2021). «Нитратное дыхание и суточная миграция диатомовых водорослей связаны в отложениях под микробным матом» . Экологическая микробиология . 23 (3): 1422–1435. Бибкод : 2021EnvMi..23.1422M . дои : 10.1111/1462-2920.15345 . hdl : 2027.42/167035 . PMID 33264477 . S2CID 227259897 .
- ^ Гири, доктор медицины (2012). «Сколько существует видов водорослей?». Журнал психологии . 48 (5): 1057–1063. Бибкод : 2012JPcgy..48.1057G . дои : 10.1111/j.1529-8817.2012.01222.x . ПМИД 27011267 . S2CID 30911529 .
- ^ Раунд, Фрэнк Эрик; Кроуфорд, РМ; Манн, Д.Г. (1990). Диатомовые водоросли: биология и морфология родов . Издательство Кембриджского университета. ISBN 978-0-521-36318-1 . Архивировано из оригинала 24 мая 2021 года . Проверено 13 ноября 2013 г. [ нужна страница ]
- ^ Кантер-Лунд, Х. и Лунд, JWG (1995). Пресноводные водоросли: объяснение их микроскопического мира , Biopress Limited. ISBN 0-948737-25-5 . [ нужна страница ]
- ^ Манн, Дэвид Г. (2005). «Видовая концепция диатомовых водорослей: свидетельства морфологически различных симпатрических гамодем у четырех эпипелических видов». Систематика и эволюция растений . 164 (1/4): 215–37. дои : 10.1007/BF00940439 . JSTOR 23675282 . S2CID 37684109 .
- ^ Фуртанье, Элизабет; Кочиолек, Дж. Патрик (1999). «Каталог родов диатомовых водорослей». Диатомовые исследования . 14 (1): 1–190. Бибкод : 1999DiaRe..14....1F . дои : 10.1080/0269249X.1999.9705462 .
- ^ По состоянию на июль 2020 года во Всемирном реестре морских видов перечислено 1356 названий родов диатомовых водорослей из всех сред обитания, из которых 1248 «приняты».
- ^ Запросы во Всемирный реестр морских видов, июль 2020 г., возвращают 299 названий родов, «только ископаемых», из которых 285 «приняты».
- ^ Териот, Эдвард С.; Кэннон, Джейми Дж.; Гутелл, Робин Р.; Алверсон, Эндрю Дж. (2009). «Пределы ядерно-кодируемой рДНК SSU для решения филогении диатомей» . Европейский журнал психологии . 4 (3): 277–290. дои : 10.5091/plecevo.2010.418 . ПМЦ 2835975 . ПМИД 20224747 .
- ^ Териот, Эдвард С.; Эшворт, Мэтт; Рак, Элизабет; Наков, Теофил; Янсен, Роберт К. (2010). «Предварительная мультигенная филогения диатомовых водорослей (Bacillariophyta): задачи для будущих исследований» . Экология и эволюция растений . 143 (3): 277–290. дои : 10.1080/09670260902749159 . ПМЦ 2835975 . ПМИД 20224747 .
- ^ Паркс, Мэтью Б.; Уикетт, Норман Дж.; Алверсон, Эндрю Дж. (2018). «Сигнал, неопределенность и конфликт в филогеномных данных для разнообразной линии микробных эукариот (диатомовых, Bacillariophyta)» . Молекулярная биология и эволюция . 35 (1): 80–93. дои : 10.1093/molbev/msx268 . ПМЦ 5850769 . ПМИД 29040712 .
- ^ Медлин, ЛК; Десдевис, Ю. (2020). «Обзор филогенетической реконструкции диатомей с использованием молекулярных инструментов с анализом набора данных из семи генов с использованием нескольких внешних групп и морфологических данных для комплексного подхода к доказательствам» (PDF) . Психология . в прессе. Архивировано (PDF) из оригинала 20 июля 2020 года . Проверено 20 июля 2020 г.
- ^ Адл, Сина М.; Басс, Дэвид; Лейн, Кристофер Э.; Лукеш, Юлиус; Шох, Конрад Л.; Смирнов, Алексей; и др. (26 сентября 2018 г.). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Мишра, Мирамбика; Аруха, Ананта П.; Башир, Туфаил; Ядав, Дхананджай; Прасад, ГБКС (5 июля 2017 г.). «Все новые лица диатомей: потенциальный источник наноматериалов и не только» . Границы микробиологии . 8 . Frontiers Media SA: 1239. doi : 10.3389/fmicb.2017.01239 . ISSN 1664-302X . ПМК 5496942 . ПМИД 28725218 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0. Архивировано 16 октября 2017 г. на Wayback Machine .
- ^ Перейти обратно: а б Кооистра, Вибе ХКФ; Медлин, Линда К. (1996). «Эволюция диатомовых водорослей (Bacillariophyta)». Молекулярная филогенетика и эволюция . 6 (3): 391–407. дои : 10.1006/mpev.1996.0088 . ПМИД 8975694 .
- ^ Шибер, Юрген; Кринсли, Дэйв; Ричипути, Ли (2000). «Диагенетическое происхождение кварцевого ила в аргиллитах и последствия для круговорота кремнезема». Природа . 406 (6799): 981–5. Бибкод : 2000Natur.406..981S . дои : 10.1038/35023143 . ПМИД 10984049 . S2CID 4417951 .
- ^ Медлин, ЛК; Куистра, WHCF; Герсонд, Р.; Симс, Пенсильвания; Уэллброк, У. (1997). «Связано ли происхождение диатомовых водорослей с массовым вымиранием в конце Перми?». Нова Хедвигия . 65 (1–4): 1–11. doi : 10.1127/nova.hedwigia/65/1997/1 . hdl : 10013/epic.12689 .
- ^ Рэйвен, Дж.А.; Уэйт, AM (2004). «Эволюция окварцевания диатомовых водорослей: неизбежное погружение и погружение как спасение?» . Новый фитолог . 162 (1): 45–61. дои : 10.1111/j.1469-8137.2004.01022.x . JSTOR 1514475 .
- ^ Фальковски, П.Г.; Кац, Мириам Э.; Нолл, Эндрю Х.; Куигг, Антониетта; Рэйвен, Джон А.; Шофилд, Оскар; Тейлор, FJR (2004). «Эволюция современного эукариотического фитопланктона». Наука . 305 (5682): 354–60. Бибкод : 2004Sci...305..354F . CiteSeerX 10.1.1.598.7930 . дои : 10.1126/science.1095964 . ПМИД 15256663 . S2CID 451773 .
- ^ Киддер, Д.Л.; Герловски-Кордеш, Э.Х. (2005). «Влияние радиации пастбищ на цикл неморского кремнезема и миоценовый диатомит». ПАЛЕОС . 20 (2): 198–206. Бибкод : 2005Палай..20..198К . дои : 10.2110/palo.2003.p03-108 . JSTOR 27670327 . S2CID 140584104 .
- ^ Перейти обратно: а б Лазарь, Дэвид; Бэррон, Джон; Реноди, Йохан; Дайвер, Патрик; Тюрке, Андреас (2014). «Кайнозойское планктонное морское диатомовое разнообразие и корреляция с изменением климата» . ПЛОС ОДИН . 9 (1): e84857. Бибкод : 2014PLoSO...984857L . дои : 10.1371/journal.pone.0084857 . ПМЦ 3898954 . ПМИД 24465441 .
- ^ Основная группа авторов МГЭИК, 2007. «Изменение климата 2007: сводный отчет». 104.
- ^ Перейти обратно: а б Шерер, Р.П.; Гладенков А. Ю.; Бэррон, Дж. А. (2007). «Методы и приложения кайнозойской биостратиграфии морских диатомей». Документы Палеонтологического общества . 13 : 61–83. дои : 10.1017/S1089332600001467 .
- ^ «Что такое «прокси-данные»? | Национальные центры экологической информации (NCEI), ранее известные как Национальный центр климатических данных (NCDC)» . www.ncdc.noaa.gov . Архивировано из оригинала 8 марта 2020 года . Проверено 20 октября 2020 г.
- ^ Перейти обратно: а б Суонн, GEA; Снеллинг, AM (6 января 2015 г.). «Изменения фототической зоны в северо-западной части Тихого океана от MIS 4–5e» . Климат прошлого . 11 (1). Коперник ГмбХ: 15–25. Бибкод : 2015CliPa..11...15S . дои : 10.5194/cp-11-15-2015 . ISSN 1814-9332 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0. Архивировано 23 февраля 2011 г. на Wayback Machine .
- ^ Сигман, Дэниел М.; Жаккар, Сэмюэл Л.; Хауг, Джеральд Х. (2004). «Стратификация полярного океана в холодном климате» . Природа . 428 (6978). ООО «Спрингер Сайенс энд Бизнес Медиа»: 59–63. Бибкод : 2004Natur.428...59S . дои : 10.1038/nature02357 . ISSN 0028-0836 . ПМИД 14999278 . S2CID 4329978 . Архивировано из оригинала 19 февраля 2022 года . Проверено 15 декабря 2021 г.
- ^ Перейти обратно: а б Хауг, Джеральд Х.; Ганопольский, Андрей; Сигман, Дэниел М.; Розелл-Меле, Антони; Суонн, Джордж Э.А.; Тидеманн, Ральф; и др. (2005). «Сезонность в Северной части Тихого океана и оледенение Северной Америки 2,7 миллиона лет назад» . Природа . 433 (7028). ООО «Спрингер Сайенс энд Бизнес Медиа»: 821–825. Бибкод : 2005Natur.433..821H . дои : 10.1038/nature03332 . ISSN 0028-0836 . ПМИД 15729332 . S2CID 24116155 . Архивировано из оригинала 19 февраля 2022 года . Проверено 15 декабря 2021 г.
- ^ Суонн, Джордж Э.А.; Маслин, Марк А.; Ленг, Мелани Дж.; Слоан, Хилари Дж.; Хауг, Джеральд Х. (24 февраля 2006 г.). «Диатом δ18O свидетельствует о развитии современной системы галоклина в субарктической северо-западной части Тихого океана в начале крупного оледенения Северного полушария» . Палеоокеанография . 21 (1). Американский геофизический союз (AGU): данные отсутствуют. Бибкод : 2006PalOc..21.1009S . дои : 10.1029/2005pa001147 . ISSN 0883-8305 .
- ^ Не, Цзюньшэн; Кинг, Джон; Лю, Чжэнъюй; Клеменс, Стив; Прелл, Уоррен; Фанг, Сяоминь (2008). «Опреснение поверхностных вод: причина начала стратификации северной части Тихого океана, начиная с 2,75 млн лет назад?». Глобальные и планетарные изменения . 64 (1–2). Эльзевир Б.В.: 49–52. Бибкод : 2008GPC....64...49N . дои : 10.1016/j.gloplacha.2008.08.003 . ISSN 0921-8181 .
- ^ Суонн, Джордж Э.А. (2010). «Изменения солености в северо-западной части Тихого океана в позднем плиоцене/начале четвертичного периода от 2,73 млн лет до 2,52 млн лет назад» (PDF) . Письма о Земле и планетологии . 297 (1–2). Эльзевир Б.В.: 332–338. Бибкод : 2010E&PSL.297..332S . дои : 10.1016/j.epsl.2010.06.035 . ISSN 0012-821X . Архивировано (PDF) из оригинала 19 июля 2018 года . Проверено 15 декабря 2021 г.
- ^ Сарнтейн, М.; Гебхардт, Х.; Кифер, Т.; Кучера, М.; Кук, М.; Эрленкейзер, Х. (2004). «Среднеголоценовое происхождение низкой солености морской поверхности в субарктической северной части Тихого океана». Четвертичные научные обзоры . 23 (20–22). Эльзевир Б.В.: 2089–2099. Бибкод : 2004QSRv...23.2089S . doi : 10.1016/j.quascirev.2004.08.008 . ISSN 0277-3791 .
- ^ Жаккар, СЛ; Гэлбрейт, Эд; Сигман, DM; Хауг, GH (2010). «Повсеместная связь между керном антарктического льда и записями субарктических тихоокеанских отложений за последние 800 тысяч лет». Четвертичные научные обзоры . 29 (1–2). Эльзевир Б.В.: 206–212. Бибкод : 2010QSRv...29..206J . doi : 10.1016/j.quascirev.2009.10.007 . ISSN 0277-3791 .
- ^ Гэлбрейт, Эрик Д.; Киенаст, Маркус; Жаккар, Сэмюэл Л.; Педерсен, Томас Ф.; Брюнель, Бриджит Г.; Сигман, Дэниел М.; Кифер, Торстен (23 мая 2008 г.). «Последовательная взаимосвязь между глобальным климатом и поверхностным использованием нитратов в западной субарктической части Тихого океана на протяжении последних 500 тыс. лет назад» (PDF) . Палеоокеанография . 23 (2). Американский геофизический союз (AGU): данные отсутствуют. Бибкод : 2008PalOc..23.2212G . дои : 10.1029/2007pa001518 . ISSN 0883-8305 . S2CID 4082469 . Архивировано (PDF) из оригинала 15 июля 2021 года . Проверено 15 декабря 2021 г.
- ^ Брюнель, Бриджит Г.; Сигман, Дэниел М.; Жаккар, Сэмюэл Л.; Кейгвин, Ллойд Д.; Плессен, Биргит; Шеттлер, Георг; Кук, Меа С.; Хауг, Джеральд Х. (2010). «Ледниковые/межледниковые изменения в снабжении питательными веществами и стратификации в западной субарктической части северной части Тихого океана после предпоследнего ледникового максимума». Четвертичные научные обзоры . 29 (19–20). Эльзевир Б.В.: 2579–2590. Бибкод : 2010QSRv...29.2579B . doi : 10.1016/j.quascirev.2010.03.010 . ISSN 0277-3791 .
- ^ Кофельд, Карен Э.; Чейз, Занна (2011). «Контроль за дегляциальными изменениями биогенных потоков в северной части Тихого океана». Четвертичные научные обзоры . 30 (23–24). Эльзевир Б.В.: 3350–3363. Бибкод : 2011QSRv...30.3350K . doi : 10.1016/j.quascirev.2011.08.007 . ISSN 0277-3791 .
- ^ Харвуд, DM; Николаев, В.А.; Зима, ДМ (2007). «Меловая летопись эволюции, излучения и расширения диатомей». Документы Палеонтологического общества . 13 : 33–59. дои : 10.1017/S1089332600001455 .
- ^ Стрельникова, Н.И. (1990). «Эволюция диатомей в меловой и палеогеновый периоды». В Симоле, Х. (ред.). Материалы Десятого международного симпозиума по диатомовым водорослям . Кенигштейн: Научные книги Кельца. стр. 195–204. ISBN 3-87429-307-6 .
- ^ Балдауф, Дж. Г. (1993). «Обмен цветков диатомовых водорослей от среднего эоцена до раннего миоцена». В Протеро, Д.; Берггрен, штат Вашингтон (ред.). Эоцен-олигоценовая климатическая и биотическая эволюция . Принстон: Издательство Принстонского университета. стр. 310–326. ISBN 0-691-02542-8 .
- ^ Бэррон, Дж. А. (2003). «Появление и вымирание планктонных диатомей за последние 18 млн лет в Тихом и Южном океанах». Диатомовые исследования . 18 : 203–224. дои : 10.1080/0269249x.2003.9705588 . S2CID 84781882 .
- ^ Скала, С.; Карелс, Н.; Фальсиаторе, А; Кьюзано, ML; Боулер, К. (2002). «Свойства генома диатомовой водоросли Phaeodactylum tricornutum» . Физиология растений . 129 (3): 993–1002. дои : 10.1104/стр.010713 . ПМК 166495 . ПМИД 12114555 .
- ^ Махешвари, У.; Монтсант, А; Голл, Дж; Кришнасами, С; Раджьяшри, КР; Пателл, В.М.; Боулер, К. (2004). «База данных диатомовых EST» . Исследования нуклеиновых кислот . 33 (Проблема с базой данных): D344–7. дои : 10.1093/nar/gki121 . ПМК 540075 . ПМИД 15608213 .
- ^ Монтсант, А.; Джаббари, К; Махешвари, У; Боулер, К. (2005). «Сравнительная геномика пеннатной диатомовой водоросли Phaeodactylum tricornutum» . Физиология растений . 137 (2): 500–13. дои : 10.1104/стр.104.052829 . ПМЦ 1065351 . ПМИД 15665249 .
- ^ Махешвари, У.; Мок, Т.; Армбруст, EV; Боулер, К. (2009). «Обновление базы данных Diatom EST: новый инструмент цифровой транскриптомики» . Исследования нуклеиновых кислот . 37 (Проблема с базой данных): D1001–5. дои : 10.1093/нар/gkn905 . ПМК 2686495 . ПМИД 19029140 .
- ^ Перейти обратно: а б Армбруст, EV; Берджес, Джон А.; Боулер, Крис; Грин, Беверли Р.; Мартинес, Диего; Патнэм, Николас Х.; и др. (2004). «Геном диатомовой водоросли Thalassiosira Pseudonana: экология, эволюция и метаболизм». Наука . 306 (5693): 79–86. Бибкод : 2004Sci...306...79A . CiteSeerX 10.1.1.690.4884 . дои : 10.1126/science.1101156 . ПМИД 15459382 . S2CID 8593895 .
- ^ Перейти обратно: а б с д Боулер, Крис; Аллен, Эндрю Э.; Бэджер, Джонатан Х.; Гримвуд, Джейн; Джаббари, Камель; Куо, Алан; и др. (2008). «Геодактилум раскрывает эволюционную историю геномов диатомовых водорослей» . Природа . 456 (7219): 239–244. Бибкод : 2008Natur.456..239B . дои : 10.1038/nature07410 . ПМИД 18923393 . S2CID 4415177 .
- ^ Рой, Юго-Запад; Пенни, Д. (2007). «Очень высокая доля уникальных позиций интронов в богатой интронами диатомовой водоросли Thalassiosira pseudonana указывает на широко распространенное усиление интронов» . Молекулярная биология и эволюция . 24 (7): 1447–57. дои : 10.1093/molbev/msm048 . ПМИД 17350938 .
- ^ Маумус, Флориан; Аллен, Эндрю Э; Мхири, Коринн; Ху, Ханьхуа; Джаббари, Камель; Варди, Асаф; Гранбастьен, Мари-Анжель; Боулер, Крис (2009). «Потенциальное влияние активированных стрессом ретротранспозонов на эволюцию генома морской диатомеи» . БМК Геномика . 10 :624. дои : 10.1186/1471-2164-10-624 . ПМК 2806351 . ПМИД 20028555 .
- ^ Мустафа, А.; Бестери, Б.; Майер, У.Г.; Боулер, К.; Валентин, К.; Бхаттачарья, Д. (2009). «Геномные следы загадочного пластидного эндосимбиоза у диатомовых водорослей» (PDF) . Наука . 324 (5935): 1724–6. Бибкод : 2009Sci...324.1724M . дои : 10.1126/science.1172983 . ПМИД 19556510 . S2CID 11408339 . Архивировано из оригинала (PDF) 21 апреля 2014 года . Проверено 13 января 2019 г.
- ^ Перейти обратно: а б Крот, Питер Г.; Боунс, Атл М.; и др. (октябрь 2018 г.). «Редактирование генома диатомовых водорослей: достижения и цели» . Отчеты о растительных клетках . 37 (10): 1401–1408. дои : 10.1007/s00299-018-2334-1 . HDL : 11250/2590892 . ПМИД 30167805 . S2CID 52133809 . Архивировано из оригинала 19 февраля 2022 года . Проверено 20 мая 2021 г.
- ^ Перейти обратно: а б Фальсиаторе, Анжела; Казотти, Рафаэлла; и др. (май 2015 г.). «Трансформация генов неселектируемых портеров у морских диатомей Re». Морская биотехнология . 1 (3): 239–251. дои : 10.1007/PL00011773 . ПМИД 10383998 . S2CID 22267097 .
- ^ Перейти обратно: а б с д и Карас, Богумил Ж.; Дайнер, Рэйчел Э.; Лефевр, Стефан К.; Маккуэйд, Джефф; Филлипс, Алекс PR; Ноддингс, Чари М.; и др. (21 апреля 2015 г.). «Дизайнерские эписомы диатомей, доставленные путем бактериальной конъюгации» . Природные коммуникации . 6 (1): 6925. Бибкод : 2015NatCo...6.6925K . дои : 10.1038/ncomms7925 . ISSN 2041-1723 . ПМЦ 4411287 . ПМИД 25897682 .
- ^ Перейти обратно: а б с д Слэттери, Сэмюэл С.; Даймонд, Эндрю; Ван, Хелен; Терриен, Жасмин А.; Лант, Джереми Т.; Джейзи, Теа; и др. (16 февраля 2018 г.). «Расширенный набор генетических инструментов на основе плазмид позволяет редактировать геном Cas9 и стабильно поддерживать синтетические пути у Phaeodactylum tricornutum». ACS Синтетическая биология . 7 (2): 328–338. doi : 10.1021/acsynbio.7b00191 . ПМИД 29298053 .
- ^ Наймарк, Марианна; Шарма, Амит Кумар; и др. (июль 2016 г.). «Система CRISPR/Cas9, адаптированная для редактирования генов морских водорослей» . Научные отчеты . 6 (1): 24951. Бибкод : 2016NatSR...624951N . дои : 10.1038/srep24951 . ПМЦ 4842962 . ПМИД 27108533 .
- ^ Мишра, М; Аруха, АП; Башир, Т; Ядав, Д; Гбкс, Прасад (2017). «Все новые лица диатомей: потенциальный источник наноматериалов и не только» . Передний микробиол . 8 : 1239. дои : 10.3389/fmicb.2017.01239 . ПМК 5496942 . ПМИД 28725218 .
- ^ Река, Арианит А.; Павловский, Благой; Макрески, Петре (октябрь 2017 г.). «Новый оптимизированный метод низкотемпературного гидротермального производства пористой керамики с использованием диатомита» . Керамика Интернешнл . 43 (15): 12572–12578. doi : 10.1016/j.ceramint.2017.06.132 . Архивировано из оригинала 19 февраля 2022 года . Проверено 8 апреля 2020 г.
- ^ Река, Арианит; Ановский, Тодор; Богоевский, Слободан; Павловский, Благой; Бошковский, Бошко (29 декабря 2014 г.). «Физико-химические и минералого-петрографические исследования диатомита из месторождения близ села Рожден, Республика Македония» . Геология Македонии . 28 (2): 121–126. ISSN 1857-8586 . Архивировано из оригинала 18 апреля 2020 года . Проверено 8 апреля 2020 г.
- ^ Река, Арианит А.; Павловский, Благой; Адеми, Эгзон; Джашари, Ахмед; Боев, Блазо; Боев, Иван; Макрески, Петре (31 декабря 2019 г.). «Эффект термической обработки трепеля в диапазоне температур 800-1200˚С» . Открытая химия . 17 (1): 1235–1243. doi : 10.1515/chem-2019-0132 .
- ^ Брэдбери, Дж. (2004). «Природные нанотехнологи: раскрывая тайны диатомовых водорослей» . ПЛОС Биология . 2 (10): 1512–1515. дои : 10.1371/journal.pbio.0020306 . ПМК 521728 . ПМИД 15486572 .

- ^ Драм, Райан В.; Гордон, Ричард (2003). «Репликаторы Звездного пути и диатомовые нанотехнологии». Тенденции в биотехнологии . 21 (8): 325–8. дои : 10.1016/S0167-7799(03)00169-0 . ПМИД 12902165 .
- ^ Джонсон, Р.К. (9 апреля 2009 г.). «Диатомовые водоросли могут утроить эффективность солнечных батарей» . ЭЭ Таймс . Архивировано из оригинала 31 июля 2012 года . Проверено 13 апреля 2009 г.
- ^ Рамачандра, ТВ; Махапатра, Дурга Мадхаб; б, Картик; Гордон, Ричард (2009). «Доение диатомовых водорослей для устойчивой энергетики: биохимическая инженерия против солнечных панелей из диатомовых водорослей, выделяющих бензин». Исследования в области промышленной и инженерной химии . 48 (19): 8769–88. дои : 10.1021/ie900044j .
- ^ Ауэр, Антти (1991). «Качественный диатомовый анализ как инструмент диагностики утопления». Американский журнал судебной медицины и патологии . 12 (3): 213–8. дои : 10.1097/00000433-199109000-00009 . ПМИД 1750392 . S2CID 38370984 .
- ^ Перейти обратно: а б с Пьерелла Карлушич, Хуан Хосе; Ибарбальц, Федерико М; Боулер, Крис (2020). «Исследование морского фитопланктона: от исторического понимания до эпохи омики» . Журнал исследований планктона . 42 : 595–612. дои : 10.1093/plankt/fbaa049 . hdl : 11336/143676 .
- ^ Долан, Джон Р. (1 августа 2019 г.). «Разоблачение «старшего сына отца протозоологии»: Чарльза Кинга» . Протист . 170 (4): 374–384. дои : 10.1016/j.protis.2019.07.002 . ISSN 1434-4610 . ПМИД 31479910 . S2CID 201207778 .
- ^ Дарвин, Ричард (1866). О происхождении видов путем естественного отбора: или сохранение избранных рас в борьбе за жизнь .
Внешние ссылки
[ редактировать ]- База данных Diatom EST , Высшая нормальная школа
- Plankton*Net , таксономическая база данных, включая изображения видов диатомовых водорослей.
- История жизни и экология диатомовых водорослей , Музей палеонтологии Калифорнийского университета
- Диатомовые водоросли: «Мрамор природы» , сайт Эврика, Бергенский университет.
- История жизни и экология диатомовых водорослей , Восстановление и распространение изображений микроископаемых для обучения и образования (ЧУДО), Университетский колледж Лондона
- Страница диатомовых водорослей. Архивировано 8 октября 2009 г. в Wayback Machine , Королевский ботанический сад Эдинбурга.
- Геометрия и закономерности в природе 3: Дыры в тестах радиолярий и диатомей
- Краткая информация о диатоме , Научно-исследовательский институт аквариумов Монтерей-Бей
- База данных изображений водорослей. Архивировано 14 августа 2011 г. в Wayback Machine Академии естественных наук в Филадельфии (ANSP).
- Таксоны диатомовых водорослей. Архивировано 3 октября 2011 г. в Wayback Machine Академии естественных наук в Филадельфии (ANSP).
- Введение в микроскопическое исследование диатомей. Архивировано 5 августа 2022 года в Wayback Machine. Робертом Б. Маклафлином
