Экзоцитоз
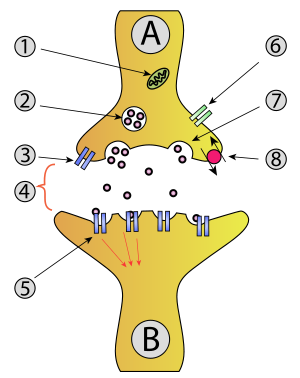
- Митохондрия
- Синаптический везикула с нейротрансмиттерами
- Ауторецептор
- Синапс с высвобождением нейротрансмиттера ( серотонина )
- Постсинаптические рецепторы активируются нейромедиатором (индукция постсинаптического потенциала)
- Кальциевый канал
- Экзоцитоз пузырька
- Возвращенный нейромедиатор
Экзоцитоз ( / ˌ ɛ k s oʊ s aɪ ˈ t oʊ s ɪ s / [ 1 ] [ 2 ] ) — форма активного транспорта и объемного транспорта , при которой клетка транспортирует молекулы (например, нейротрансмиттеры и белки ) из клетки ( экзо- + цитоз ). Будучи активным транспортным механизмом, экзоцитоз требует использования энергии для транспортировки материала. Экзоцитоз и его аналог эндоцитоз используются всеми клетками, поскольку большинство химических веществ важных для них представляют собой большие полярные молекулы, которые не могут пройти через гидрофобную часть клеточной мембраны пассивным путем . Экзоцитоз — это процесс, при котором высвобождается большое количество молекул; таким образом, это форма перевозки массовых грузов. Экзоцитоз происходит через секреторные порталы в плазматической мембране клетки, называемые поросомами . Поросомы представляют собой постоянные липопротеиновые структуры чашеобразной формы на плазматической мембране клетки, где секреторные пузырьки временно стыкуются и сливаются, высвобождая внутривезикулярное содержимое из клетки.
При экзоцитозе мембраносвязанные секреторные везикулы переносятся на клеточную мембрану , где они стыкуются и сливаются с поросомами , а их содержимое (т.е. водорастворимые молекулы) секретируется во внеклеточную среду. Эта секреция возможна, поскольку везикула временно сливается с плазматической мембраной. В контексте нейротрансмиссии нейротрансмиттеры обычно высвобождаются из синаптических пузырьков в синаптическую щель посредством экзоцитоза; однако нейромедиаторы также могут высвобождаться путем обратного транспорта через мембранные транспортные белки .
Экзоцитоз также является механизмом, с помощью которого клетки способны вставлять мембранные белки (такие как ионные каналы и рецепторы клеточной поверхности ), липиды и другие компоненты в клеточную мембрану. Везикулы, содержащие эти мембранные компоненты, полностью сливаются с внешней клеточной мембраной и становятся ее частью.
История
[ редактировать ]Термин был предложен Де Дювом в 1963 году. [ 3 ]
Типы
[ редактировать ]У эукариот различают два типа экзоцитоза: 1) Ка 2+ запускаемый неконститутивный (т.е. регулируемый экзоцитоз) и 2) некальций 2+ триггерный конститутивный (т.е. нерегулируемый).
Что 2+ для запуска неконститутивного экзоцитоза необходим внешний сигнал, специфический сигнал сортировки на везикулах, клатриновой оболочке, а также увеличение внутриклеточного кальция. В многоклеточных организмах этот механизм инициирует многие формы межклеточной коммуникации, такие как синаптическая передача, секреция гормонов нейроэндокринными клетками и секреция иммунных клеток. В нейронах и эндокринных клетках SNARE-белки и SM-белки катализируют слияние, образуя комплекс, который объединяет две слитые мембраны. Например, в синапсах комплекс SNARE образуется синтаксином-1 и SNAP25 на плазматической мембране и VAMP2 на мембране пузырька. [ 4 ] нейронов Экзоцитоз в химических синапсах – это Ca 2+ срабатывает и служит межнейронной передаче сигналов. Сенсоры кальция, запускающие экзоцитоз, могут взаимодействовать либо с комплексом SNARE, либо с фосфолипидами сливающихся мембран. Синаптотагмин признан основным сенсором Ca. 2+ вызывали экзоцитоз у животных. [ 5 ] Однако белки синаптотагмины отсутствуют у растений и одноклеточных эукариот. Другими потенциальными сенсорами кальция для экзоцитоза являются белки EF-hand (например: кальмодулин) и белки, содержащие домен C2 (например: Ferlins, E-синаптотагмин, Doc2b). Неясно, как различные сенсоры кальция могут взаимодействовать друг с другом и определенным образом опосредовать кинетику экзоцитоза, запускаемую кальцием. [ 6 ]
Конститутивный экзоцитоз осуществляется всеми клетками и служит высвобождению компонентов внеклеточного матрикса или доставке вновь синтезированных мембранных белков, которые встраиваются в плазматическую мембрану после слияния транспортного пузырька . Нет четкого консенсуса относительно механизмов и молекулярных процессов, которые управляют образованием, почкованием, транслокацией и слиянием пост-Гольджи-везикул с плазматической мембраной. Слияние включает привязку мембран (распознавание) и слияние мембран. До сих пор неясно, различны ли механизмы конститутивной и регулируемой секреции. Механизм, необходимый для конститутивного экзоцитоза, изучен не так хорошо, как механизм регулируемого экзоцитоза. Два привязывающих комплекса связаны с конститутивным экзоцитозом у млекопитающих: ELKS и Exocyst. ELKS представляет собой большой спиральный белок, также участвующий в синаптическом экзоцитозе, отмечая точки слияния «горячих точек» слияния секреторных носителей. Экзоциста представляет собой октамерный белковый комплекс. У млекопитающих компоненты экзоцист локализуются как в плазматической мембране, так и в аппарате Гольджи, а белки экзоцист колокализуются в точке слияния пост-Гольджи-везикул. Мембранное слияние конститутивного экзоцитоза, вероятно, опосредуется SNAP29 и Syntaxin19 на плазматической мембране и YKT6 или VAMP3 на мембране везикул. [ 7 ]
Везикулярный экзоцитоз у прокариот грамотрицательных бактерий является третьим механизмом и последним открытием экзоцитоза. Периплазма отщипывается в виде пузырьков внешней мембраны бактерий (OMV) для передачи микробных биохимических сигналов в эукариотические клетки-хозяева. [ 8 ] или другие микробы, находящиеся поблизости, [ 9 ] осуществление контроля над секретирующим микробом в его среде, включая инвазию в хозяина, эндотоксемию, конкуренцию с другими микробами за питание и т. д. Это обнаружение перемещения мембранных пузырьков, происходящего на границе между хозяином и патогеном, также развеивает миф о том, что экзоцитоз - это чисто эукариотическая клетка. явление. [ 10 ]
Шаги
[ редактировать ]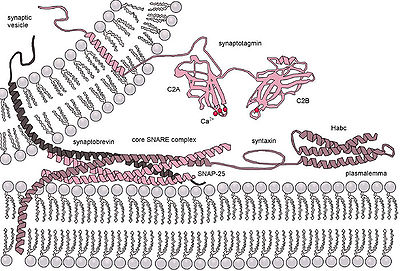
В экзоцитозе участвуют пять этапов:
Торговля везикулами
[ редактировать ]Определенные этапы транспортировки везикул требуют транспортировки везикул на умеренно небольшое расстояние. Например, везикулы, которые транспортируют белки из аппарата Гольджи к поверхности клетки, вероятно, будут использовать моторные белки и цитоскелетные пути, чтобы приблизиться к своей цели. До того, как привязка стала уместной, многие белки, используемые для активного транспорта, вместо этого были настроены на пассивный транспорт, поскольку аппарату Гольджи не требуется АТФ для транспортировки белков. В этих процессах участвуют как актин, так и основа микротрубочек, а также несколько моторных белков . Как только везикулы достигают своих целей, они вступают в контакт с привязывающими факторами, которые могут их сдерживать.
Привязка пузырьков
[ редактировать ]Полезно различать начальную, слабую связь везикул с их целью и более стабильные, упаковывающие взаимодействия. Привязка включает в себя связи на расстояниях более половины диаметра пузырька от данной поверхности мембраны (> 25 нм). Привязывающие взаимодействия, вероятно, участвуют в концентрации синаптических пузырьков в синапсе .
стыковка пузырьков
[ редактировать ]Секреторные везикулы временно стыкуются и сливаются с поросомой плазматической мембраны клетки посредством плотного кольцевого комплекса t-/v-SNARE.
Праймирование везикул
[ редактировать ]При нейрональном экзоцитозе термин «прайминг» использовался для обозначения всех молекулярных перестроек и АТФ-зависимых модификаций белков и липидов, которые происходят после первоначальной стыковки синаптического пузырька, но до экзоцитоза, так что приток ионов кальция — это все, что происходит. необходимо, чтобы вызвать почти мгновенное высвобождение нейротрансмиттера . В других типах клеток, секреция которых является конститутивной (т.е. непрерывной, независимой от ионов кальция, не запускаемой), прайминг отсутствует.
Слияние пузырьков
[ редактировать ]
Временное слияние везикул осуществляется белками SNARE , что приводит к высвобождению содержимого везикул во внеклеточное пространство (или в случае нейронов в синаптической щели).
Слияние донорной и акцепторной мембран решает три задачи:
- Поверхность плазматической мембраны увеличивается (на поверхность сросшегося пузырька). Это важно для регуляции размера клеток, например, во время роста клеток.
- Вещества внутри пузырька высвобождаются наружу. Это могут быть продукты жизнедеятельности или токсины , или сигнальные молекулы, такие как гормоны или нейротрансмиттеры во время синаптической передачи .
- Белки , встроенные в мембрану пузырька, теперь являются частью плазматической мембраны. Сторона белка, которая была обращена внутрь везикулы, теперь обращена к внешней стороне клетки. Этот механизм важен для регуляции трансмембранных и транспортеров.
Извлечение везикул
[ редактировать ]Извлечение синаптических везикул происходит путем эндоцитоза . Большинство синаптических везикул перерабатываются без полного слияния с мембраной ( слияние «поцелуй и беги» ) через поросому . Неконститутивный экзоцитоз и последующий эндоцитоз представляют собой процессы с высокими энергозатратами и, следовательно, зависят от митохондрий . [ 12 ]
Исследование клеток после секреции с помощью электронной микроскопии демонстрирует повышенное присутствие частично пустых везикул после секреции. Это позволило предположить, что в ходе секреторного процесса только часть содержимого везикул способна выйти из клетки. Это могло бы быть возможно только в том случае, если бы везикула временно установила непрерывность с клеточной плазматической мембраной в поросомах , вытеснила часть своего содержимого, затем отделилась, снова запечаталась и вышла в цитозоль (эндоцитоза). Таким образом, секреторный пузырек можно было повторно использовать для последующих раундов экзоэндоцитоза до тех пор, пока он полностью не опорожнится. [ 13 ]
См. также
[ редактировать ]- Эндоцитоз
- Пиноцитоз
- Фагоцитоз
- Мембранная нанотрубка
- Вирусное выделение
- Пресинаптическая активная зона
- Остаточное тело
- Дегрануляция
Ссылки
[ редактировать ]- ^ «Экзоцитоз» . Lexico Британский словарь английского языка . Издательство Оксфордского университета . Архивировано из оригинала 22 марта 2020 г.
- ^ «Экзоцитоз» . Словарь Merriam-Webster.com . Мерриам-Вебстер . Проверено 21 января 2016 г.
- ^ Ригер, Ригомар; Михаэлис, Арнд; Грин, Мелвин М. (06 декабря 2012 г.). Глоссарий генетики: классическая и молекулярная . Springer Science & Business Media. ISBN 978-3-642-75333-6 .
- ^ Шин, Огайо (17 января 2011 г.). Терджунг, Рональд (ред.). Комплексная физиология . Том. 4 (1-е изд.). Уайли. стр. 149–175. дои : 10.1002/cphy.c130021 . ISBN 978-0-470-65071-4 . ПМИД 24692137 .
- ^ Вулфс, Энн С; Дин, Камин (август 2020 г.). «Разнообразие изоформ синаптотагмина» . Современное мнение в нейробиологии . 63 : 198–209. дои : 10.1016/j.conb.2020.04.006 . ПМИД 32663762 . S2CID 220480746 .
- ^ Панг, Чжипин П; Зюдхоф, Томас С. (август 2010 г.). «Клеточная биология экзоцитоза, запускаемого Ca2+» . Современное мнение в области клеточной биологии . 22 (4): 496–505. дои : 10.1016/j.ceb.2010.05.001 . ПМЦ 2963628 . ПМИД 20561775 .
- ^ Сталдер, Даниэль; Гершлик, Дэвид К. (ноябрь 2020 г.). «Прямые пути переноса от аппарата Гольджи к плазматической мембране» . Семинары по клеточной биологии и биологии развития . 107 : 112–125. дои : 10.1016/j.semcdb.2020.04.001 . ПМК 7152905 . ПМИД 32317144 .
- ^ YashRoy RC (1993) Электронно-микроскопические исследования поверхностных волосков и везикул Salmonella 3,10:r:- организмов. Индийский журнал наук о животных , том. 63, стр. 99-102. https://www.researchgate.net/publication/230817087_Electron_microscope_studies_of_surface_pilli_and_vesicles_of_Salmonella_310r-_organisms?ev=prf_pub
- ^ Кадуругамува, JL; Беверидж, Ти Джей (1996). «Бактериолитическое действие мембранных везикул Pseudomonas aeruginosa на другие бактериальные, в том числе возбудители: концептуально новые антибиотики» . Журнал бактериологии . 178 (10): 2767–2774. дои : 10.1128/jb.178.10.2767-2774.1996 . ПМК 178010 . ПМИД 8631663 .
- ^ ЯшРой, RC (1998). «Открытие везикулярного экзоцитоза у прокариот и его роль в инвазии сальмонелл» (PDF) . Современная наука . 75 (10): 1062–1066.
- ^ Георгиев, Данко Д.; Джеймс Ф. Глейзбрук (2007). «Субнейронная обработка информации уединенными волнами и случайными процессами». В Лышевском, Сергей Эдвард (ред.). Справочник по нано- и молекулярной электронике . Серия «Нано и микроинженерия». ЦРК Пресс. стр. 17–1–17–41. дои : 10.1201/9781315221670-17 . ISBN 978-0-8493-8528-5 . S2CID 199021983 .
- ^ Иванников, М.; и др. (2013). «Экзоцитоз синаптических пузырьков в синаптосомах гиппокампа напрямую коррелирует с общим объемом митохондрий» . Дж. Мол. Неврология. 49 (1): 223–230. дои : 10.1007/s12031-012-9848-8 . ПМЦ 3488359 . ПМИД 22772899 .
- ^ Бор, В.Ф. и Булпап, Э.Л. (2012), Медицинская физиология. Клеточный и молекулярный подход , вып. 2, Филадельфия: Эльзевир [ постоянная мертвая ссылка ]
Внешние ссылки
[ редактировать ]- Экзоцитоз Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)