f-отношение (океанография)
| Часть серии о |
| Планктон |
|---|
 |
В океана биогеохимии f -коэффициент — это доля общего первичного производства, поддерживаемая нитратами (в отличие от других азота, соединений таких как аммоний ). Первоначально это соотношение было определено Ричардом Эппли и Брюсом Петерсоном в одной из первых статей, оценивающих глобальную океаническую продукцию. [1] Первоначально эта фракция считалась значительной, поскольку она, по-видимому, напрямую связана с (выносом) потока органического опусканием морского снега с поверхности океана насосом биологическим . Однако эта интерпретация основывалась на предположении о сильном разделении по глубине параллельного процесса, нитрификации , что поставило под сомнение более поздние измерения. [2]
Обзор
[ редактировать ]Гравитационное погружение организмов (или остатков организмов) переносит твердые частицы органического углерода из поверхностных вод океана в его глубь . Этот процесс известен как биологический насос, и его количественная оценка представляет интерес для ученых, поскольку он является важным аспектом Земли цикла углеродного . По сути, это связано с тем, что углерод, переносимый в глубины океана, изолирован от атмосферы, что позволяет океану выступать в качестве резервуара углерода. Этот биологический механизм сопровождается физико-химическим механизмом, известным как насос растворимости , который также переносит углерод в глубокие недра океана.
Измерение потока тонущего материала (так называемого морского снега) обычно осуществляется путем установки ловушек для отложений , которые перехватывают и сохраняют материал, когда он опускается в толщу воды . Однако это относительно сложный процесс, поскольку ловушки может быть неудобно развертывать или восстанавливать, и их необходимо оставлять на месте в течение длительного периода, чтобы интегрировать опускающийся поток. Более того, известно, что они испытывают смещения и интегрируют как горизонтальные, так и вертикальные потоки из-за водных потоков. [3] [4] По этой причине учёных интересуют свойства океана, которые легче измерить и которые служат показателем опускающегося потока. Коэффициент f является одним из таких показателей.
«Новое» и «возрожденное» производство
[ редактировать ]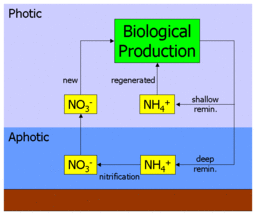
Биодоступный азот встречается в океане в нескольких формах, включая простые ионные формы, такие как нитрат (NO 3 − ), нитрит (NO 2 − ) и аммоний (NH 4 + ) и более сложные органические формы, такие как мочевина ((NH 2 ) 2 CO). Эти формы используются автотрофным фитопланктоном для синтеза органических молекул, таких как аминокислоты (строительные блоки белков ). Поедание фитопланктона зоопланктоном и более крупными организмами переносит этот органический азот вверх по пищевой цепи и по всей морской пищевой сети .
Когда азотистые органические молекулы в конечном итоге метаболизируются организмами, они возвращаются в толщу воды в виде аммония (или более сложных молекул, которые затем метаболизируются до аммония). Это известно как регенерация , поскольку аммоний может использоваться фитопланктоном и снова попадать в пищевую сеть. Таким образом, первичное производство, использующее аммиак, называется регенерированным производством . [5]
Однако аммоний также может быть окислен до нитрата (через нитрит) в процессе нитрификации. Это осуществляется различными бактериями в два этапа:
- NH 3 + O 2 → NO 2 − + 3H + + 2е −
- НЕТ 2 − + H2O → NO3 − +2Ч + + 2е −
Важно отметить, что считается, что этот процесс происходит только в отсутствие света (или как какая-то другая функция глубины). В океане это приводит к вертикальному отделению нитрификации от первичной продукции и ограничивает ее афотической зоной . Это приводит к тому, что любой нитрат в толще воды должен происходить из афотической зоны и происходить из органического материала, перенесенного туда при погружении. Таким образом, первичное производство, подпитываемое нитратами, использует «свежий» источник питательных веществ, а не регенерированный. Таким образом, производство нитратов называется новым производством . [5]
Рисунок в начале этого раздела иллюстрирует это. Нитрат и аммоний поглощаются первичными продуцентами, перерабатываются через пищевую сеть, а затем регенерируются в аммоний. Часть этого обратного потока выбрасывается на поверхность океана (где он снова доступен для поглощения), а часть возвращается на глубину. Аммоний, возвращаемый на глубину, нитрифицируется до нитрата и в конечном итоге смешивается или поднимается в поверхностный океан, чтобы повторить цикл.
Следовательно, значение новой продукции заключается в ее связи с тонущим материалом. В состоянии равновесия экспортный поток органического материала, опускающегося в афотическую зону, уравновешивается восходящим потоком нитратов. Измеряя, сколько нитратов потребляется первичным производством по отношению к количеству регенерированного аммония, можно косвенно оценить поток экспорта.
Кроме того, коэффициент f также может выявить важные аспекты функционирования местной экосистемы. [6] Высокие значения коэффициента f обычно связаны с продуктивными экосистемами, в которых доминирует крупный эукариотический фитопланктон (например, диатомовые водоросли ), который питается крупным зоопланктоном (и, в свою очередь, более крупными организмами, такими как рыбы). Напротив, низкие значения f-коэффициента обычно связаны с низкой биомассой, олиготрофными пищевыми сетями, состоящими из небольшого прокариотического фитопланктона (такого как прохлорококк ), который контролируется микрозоопланктоном. [7] [8]
Предположения
[ редактировать ]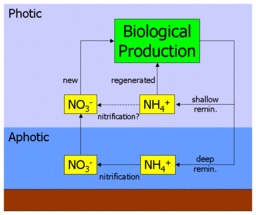
Фундаментальным предположением в этой интерпретации коэффициента f является пространственное разделение первичной продукции и нитрификации. Действительно, в своей оригинальной статье Эппли и Петерсон отметили, что: «Чтобы связать новое производство с экспортом, необходимо, чтобы нитрификация в эвфотической зоне была незначительной». [1] Однако последующие наблюдательные работы по распространению нитрификации показали, что нитрификация может происходить на меньших глубинах и даже внутри фотической зоны. [9] [10] [11]
Как показывает диаграмма рядом, если аммоний действительно нитрифицируется до нитрата в поверхностных водах океана, это, по сути, « короткое замыкание » глубокого пути нитратов. На практике это привело бы к переоценке нового производства и более высокому коэффициенту фтора, поскольку часть якобы нового производства на самом деле будет подпитываться недавно нитрифицированным нитратом, который никогда не покидал поверхность океана. После включения измерений нитрификации в свою параметризацию экосистемная модель региона олиготрофного субтропического круговорота (в частности, участка BATS ) обнаружила, что в годовом исчислении около 40% поверхностных нитратов недавно нитрифицировалось (летом эта цифра возрастает почти до 90%). [12] Дальнейшее исследование, синтезирующее географически разнообразные измерения нитрификации, выявило высокую изменчивость, но не связано с глубиной, и применило это в модели глобального масштаба для оценки того, что до половины поверхностных нитратов поступает за счет поверхностной нитрификации, а не апвеллинга. [2]
Хотя измерения скорости нитрификации все еще относительно редки, они позволяют предположить, что коэффициент f не является таким простым показателем биологического насоса, как когда-то считалось. По этой причине некоторые исследователи предложили различать f-коэффициент и отношение экспорта твердых частиц к первичному производству, которое они называют пер-коэффициентом . [8] Хотя количественно он отличается от f-коэффициента, он демонстрирует аналогичные качественные различия между режимами высокой продуктивности/высокой биомассы/высокого экспорта и режимами низкой продуктивности/низкой биомассы/низкого экспорта.
Кроме того, еще одним процессом, который потенциально усложняет использование коэффициента f для оценки «новой» и «регенерированной» продукции, является диссимиляционное восстановление нитрата до аммония (ДНРА). В средах с низким содержанием кислорода, таких как зоны минимума кислорода и отложения морского дна , хемоорганогетеротрофные микробы используют нитрат в качестве акцептора электронов для дыхания . [13] восстанавливая его до нитрита, затем до аммония. Поскольку, как и нитрификация, DNRA изменяет баланс доступности нитрата и аммония, она может внести неточность в расчетное соотношение f. Однако, поскольку возникновение DNRA ограничивается анаэробными ситуациями, [14] ее значение менее широко распространено, чем нитрификация, хотя она может происходить в связи с первичными производителями. [15] [16]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Эппли, RW; Петерсон, Би Джей (1979). «Поток твердых частиц органического вещества и новая продукция планктона в глубоком океане». Природа . 282 (5740): 677–680. Бибкод : 1979Natur.282..677E . дои : 10.1038/282677a0 . S2CID 42385900 .
- ^ Перейти обратно: а б Юл, А.; Мартин, АП; Фернандес, К.; Кларк, доктор медицинских наук (2007). «Значение нитрификации для новой продукции океана». Природа . 447 (7147): 999–1002. Бибкод : 2007Natur.447..999Y . дои : 10.1038/nature05885 . ПМИД 17581584 . S2CID 4416535 .
- ^ Томас, С.; Ридд, П.В. (2004). «Обзор методов измерения кратковременного накопления отложений». Морская геология . 207 (1–4): 95–114. Бибкод : 2004МГеол.207...95Т . дои : 10.1016/j.margeo.2004.03.011 .
- ^ Бюсселер, КО ; и др. (2007). «Оценка использования ловушек для отложений для оценки потоков частиц в верхних слоях океана» (PDF) . Дж. Мар. Рес . 65 (3): 345–416. дои : 10.1357/002224007781567621 . HDL : 1912/1803 . ISSN 0022-2402 .
- ^ Перейти обратно: а б Дагдейл, Колорадо; Геринг, Джей-Джей (1967). «Поглощение новых и регенерированных форм азота в первичной продуктивности» . Лимнол. Океаногр . 12 (2): 196–206. Бибкод : 1967LimOc..12..196D . дои : 10.4319/lo.1967.12.2.0196 .
- ^ Аллен, А.Е.; Ховард-Джонс, Миннесота; Бут, МГ; Фришер, Мэн; Верити, П.Г.; Бронк, округ Колумбия; Сандерсон, член парламента (2002). «Значение гетеротрофной бактериальной ассимиляции аммония и нитратов в Баренцевом море летом». Журнал морских систем . 38 (1–2): 93–108. Бибкод : 2002JMS....38...93A . дои : 10.1016/s0924-7963(02)00171-9 .
- ^ Законы, Э.А.; Фальковски, П.Г.; Смит, Висконсин; Даклоу, Х.; Маккарти, Джей-Джей (2000). «Влияние температуры на экспортную продукцию в открытом океане» . Глобальные биогеохимические циклы . 14 (4): 1231–1246. Бибкод : 2000GBioC..14.1231L . дои : 10.1029/1999GB001229 .
- ^ Перейти обратно: а б Данн, JP; Армстронг, РА; Гнанадэсикан, А.; Сармьенто, JL (2005). «Эмпирические и механистические модели коэффициента экспорта частиц» . Глобальные биогеохимические циклы . 19 (4): GB4026. Бибкод : 2005GBioC..19.4026D . дои : 10.1029/2004GB002390 . S2CID 9019774 .
- ^ Доре, Дж. Э.; Карл, DM (1996). «Нитрификация в эвфотической зоне как источник нитритов, нитратов и закиси азота на станции АЛОХА» . Лимнол. Океаногр . 41 (8): 1619–1628. Бибкод : 1996LimOc..41.1619D . дои : 10.4319/lo.1996.41.8.1619 . JSTOR 00243590 .
- ^ Рембо, П.; Славик, Г.; Буджелал, Б.; Коатаноан, К.; Конан, П.; Косте, Б.; Гарсия, Н.; Мутен, Т.; Пужо-Пэй, М. (1999). «Поглощение и экспорт углерода и азота в экваториальной части Тихого океана на 150° з.д.: свидетельства эффективного регенерированного производственного цикла» . Дж. Геофиз. Рез . 104 (С2): 3341–3356. Бибкод : 1999JGR...104.3341R . дои : 10.1029/1998JC900004 . S2CID 55579865 .
- ^ Диас, Ф.; Рэмбо, П. (2000). «Регенерация азота и выброс растворенного органического азота весной в прибрежной зоне северо-запада Средиземноморья (Львиный залив): значение для оценки нового производства» . Мар Экол. Прог. Сер . 197 : 51–65. Бибкод : 2000MEPS..197...51D . дои : 10.3354/meps197051 .
- ^ Мартин, АП; Пондавен, П. (2006). «Новое первичное производство и нитрификация в западной субтропической части Северной Атлантики: моделирование» . Глобальные биогеохимические циклы . 20 (4): н/д. Бибкод : 2006GBioC..20.4014M . дои : 10.1029/2005GB002608 .
- ^ Крафт, Б. Страус, М. и Тегетмейер, Х.Э. (2011). «Микробное нитратное дыхание - Гены, ферменты и распространение в окружающей среде». Журнал биотехнологии . 155 (1): 104–117. doi : 10.1016/j.jbiotec.2010.12.025 . ПМИД 21219945 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лам, Филлис и Кайперс, Марсель М.М. (2011). «Микробные азотные процессы в зонах минимума кислорода». Ежегодный обзор морской науки . 3 : 317–345. Бибкод : 2011ARMS....3..317L . doi : 10.1146/annurev-marine-120709-142814 . hdl : 21.11116/0000-0001-CA25-2 . ПМИД 21329208 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Камп, Аня; Пиво, Дирк де; Нитч, Яна Л.; Лавик, Гауте; Стиф, Питер (5 апреля 2011 г.). «Диатомовые водоросли дышат нитратами, чтобы выжить в темноте и бескислородных условиях» . Труды Национальной академии наук . 108 (14): 5649–5654. Бибкод : 2011PNAS..108.5649K . дои : 10.1073/pnas.1015744108 . ISSN 0027-8424 . ПМК 3078364 . ПМИД 21402908 .
- ^ Камп, Аня; Стиф, Питер; Кнаппе, Ян; Пиво, Дирк де (2 декабря 2013 г.). «Реакция вездесущей пелагической диатомовой водоросли Thalassiosira weissflogii на темноту и аноксию» . ПЛОС ОДИН . 8 (12): е82605. Бибкод : 2013PLoSO...882605K . дои : 10.1371/journal.pone.0082605 . ISSN 1932-6203 . ПМЦ 3846789 . ПМИД 24312664 .

