Межклеточная связь

Межклеточная коммуникация (ICC) относится к различным способам и структурам, которые биологические клетки используют для общения друг с другом напрямую или через окружающую среду. Часто окружающую среду рассматривают как внеклеточные пространства внутри животного. В более широком смысле клетки могут также общаться с другими животными, принадлежащими к их собственной группе или виду, или с другими видами в более широкой экосистеме. Разные типы клеток используют разные белки и механизмы для связи друг с другом с помощью внеклеточных сигнальных молекул или электрических колебаний, которые можно сравнить с межклеточной сетью Ethernet. [2] Компоненты каждого типа межклеточной коммуникации могут участвовать более чем в одном типе коммуникации. [2] попытки четко разделить перечисленные виды общения несколько тщетны. В общих чертах, межклеточная коммуникация может быть классифицирована как происходящая внутри одного животного или между животным и другими животными в экосистеме, в которой оно живет. В этой статье межклеточная коммуникация была дополнительно сопоставлена по различным областям исследований, а не по функциональным или структурным характеристикам.
Общение внутри организма
[ редактировать ]Передача сигналов ячейки
[ редактировать ]Передача сигналов молекулярных клеток
[ редактировать ]
Одноклеточные организмы будут чувствовать окружающую среду в поисках пищи и могут посылать сигналы другим клеткам, чтобы они вели себя симбиотически или размножались. Классический пример – слизевика . Слизевик демонстрирует, как межклеточная связь с небольшой молекулой, например, циклическим АМФ, позволяет простому организму формироваться из организованного скопления одиночных клеток. [3] Исследования клеточной передачи сигналов изучали рецептор, специфичный для каждого сигнала, или несколько рецепторов, потенциально активируемых одним сигналом. [4] Важно не только наличие или отсутствие сигнала, но и его сила. Использование химического градиента для координации роста и дифференцировки клеток продолжает оставаться важным, поскольку многоклеточные животные и растения становятся все более сложными. Этот тип межклеточной коммуникации внутри организма обычно называют передачей сигналов клетки. Этот тип межклеточной коммуникации характеризуется небольшой сигнальной молекулой, диффундирующей через пространство вокруг клеток. [5] часто полагаясь на диффузионный градиент, являющийся частью сигнального ответа.
Клеточные соединения
[ редактировать ]
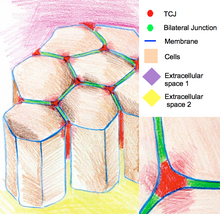
Сложные организмы могут иметь молекулы, удерживающие клетки вместе, которые также могут участвовать в межклеточной коммуникации. Некоторые связывающие молекулы называются внеклеточным матриксом и могут включать более длинные молекулы, такие как целлюлоза клеточной стенки у растений или коллаген у животных. Когда мембраны двух клеток расположены близко, они могут образовывать особые типы клеточных соединений, которые делятся на пять основных типов: адгезионные, десмосомные, щелевые, плотные и трехклеточные соединения. Адгерены, десмосомы, плотные и трехклеточные соединения выполняют структурную роль. Структуры, которые они образуют, также являются частями сложных белковых сигнальных путей. [6] В одном отношении плотные соединения играют общую роль в передаче сигналов клетками, поскольку они могут образовывать плотную застежку вокруг клеток, образуя барьер, предотвращающий проникновение даже небольших нежелательных сигнальных молекул между клетками. [7] В противном случае сигнальные молекулы могут распространиться на другую группу клеток, которые не нуждаются в сигнале или позволят сигнальным молекулам быстро уйти оттуда, где они необходимы.
Паннексины, коннексины, иннексины
[ редактировать ]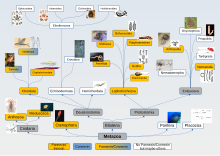
Паннексины , коннексины и иннексины представляют собой трансмембранные белки , названные в честь латинского термина nexus, означающего соединение. Они сгруппированы, поскольку все они имеют схожую структуру из 4 трансмембранных доменов, пересекающих клеточную мембрану одинаковым образом, но не все они имеют достаточную гомологию последовательностей , чтобы можно было считать их прямыми родственниками. [2] [8] Более ранние исследования с участием коннексинов продемонстрировали, что клетки образуют прямую связь друг с другом с использованием групп коннексинов, но не связей с внешней частью клетки. По существу, в то время не считалось, что они участвуют в передаче сигналов внеклеточных клеток. Более поздние исследования показали, что коннексины могут подключаться непосредственно к внешней поверхности клетки, что означает, что они являются каналом для высвобождения и поглощения сигнальных молекул из окружающей среды, внешней по отношению к клетке. [9] Более того, паннексины, по-видимому, делают это в такой степени, что редко, если вообще когда-либо, могут участвовать в прямом межклеточном соединении. [10] Как указано на изображенном дереве паннексинов/иннексинов/коннексинов, многие животные, по-видимому, не имеют паннексинов/иннексинов/коннексинов, что, возможно, указывает на то, что, возможно, еще предстоит открыть другие подобные белки, которые служат для облегчения межклеточной коммуникации у этих животных. [2]
Прямые связи между клетками
[ редактировать ]Септальные поры
[ редактировать ]
У грибов поры, пересекающие клеточные стенки и разделяющие клеточные отсеки, действуют как ICC для перемещения молекул в соседние отсеки. [11]
У большинства красных водорослей в клеточной перегородке могут быть поры , которые разделяют клетку/нить, называемые ямочными соединениями. Остаток митотического деления может закупориться клеткой. Существуют также аналогичные связи между соседними клетками/нитями, которые могут обеспечивать обмен питательными веществами. [12] Клетки разных видов могут инициировать и образовывать ямочную связь с водорослями-хозяевами. [13]
Плазмодесмы у растений
[ редактировать ]
Растительные клетки обычно имеют толстые клеточные стенки, которые необходимо пересекать, чтобы соседние клетки могли напрямую общаться. Плазмодесмы образуют трубку через клеточную стенку, образующую ICC. Трубка имеет еще одну мембранную трубку меньшего размера, концентричную ей, соединяющую эндоплазматический ретикулум двух клеток через трубку, называемую демотубулой . Большая трубка также содержит цитоскелет и другие элементы. Предполагается, что вирусы используют плазмодесмы как путь через клеточные стенки для распространения по растению. [14]
Щелевые контакты у животных
[ редактировать ]Щелевые соединения могут образовывать межклеточные связи, по сути, крошечную «трубу» с прямым регулированием, называемую парой коннексонов, между цитоплазмами двух клеток, образующих соединение. 6 коннексинов образуют коннексон, 2 коннексина образуют пару коннексонов, поэтому 12 белков коннексина образуют каждую крошечную ICC. Этот ICC позволяет двум ячейкам общаться напрямую, будучи изолированными от внешнего мира. [15] Клетки могут образовывать одну или тысячи таких крошечных ICC между собой и другими соседями, потенциально образуя большие сети напрямую связанных ячеек. Пары коннексонов образуют ICC, которые могут транспортировать воду и многие другие молекулы размером примерно до 1000 атомов. [16] и может очень быстро подавать сигнал о включении и выключении по мере необходимости. Эти ICC также передают электрические сигналы, которые можно быстро включать и выключать. Чтобы добавить к их универсальности, существует целый ряд этих типов ICC, поскольку они состоят из более чем 20 различных коннексинов с разными свойствами, которые могут комбинироваться друг с другом различными способами. Разнообразие возможных сигнальных комбинаций огромно. Хорошо изученным примером способности щелевых соединений передавать электрические сигналы являются электрические синапсы, обнаруженные на нервах. [17] [18] [19] В сердечной мышце щелевые соединения выполняют функцию координации сокращений сердца. Добавляя еще больше к их универсальности, щелевые соединения могут также функционировать, образуя прямую связь с внешней частью клетки, параллельно функционированию двоюродного брата белка паннексинов, которые объясняются в другом месте.
Межклеточный мост
[ редактировать ]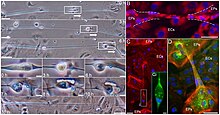
Межклеточные мостики больше, чем ICC щелевых соединений, поэтому они способны обеспечивать движение не только небольших сигнальных молекул, но и крупных молекул ДНК или даже целых клеточных органелл. Они сохраняются между двумя клетками, позволяя им обмениваться цитоплазматическим содержимым, и часто наблюдаются, когда клеткам необходимо тесное общение, например, когда они размножаются. Они обнаружены у прокариот для обмена ДНК, у мелких организмов, таких как Pinnularia , Valonia ventricosa , Volvox , C. elegans. [20] и митоз в целом ( цитокинез ), [21] Блефаризма для полового размножения и во время мейоза , включая сперматоцитогенез, для синхронизации развития половых клеток и оогенеза у более крупных организмов. Было показано, что мосты способствуют миграции клеток, как показано на рисунке рядом. [22] Цитоплазматические мосты также могут быть использованы для атаки на другую клетку, как в случае с Vampirococcus .
Слияние клеток
[ редактировать ]Клетки, которым требуется более постоянная и обширная цитоплазматическая связь, во многих случаях могут сливаться друг с другом в различной степени, образуя одну большую клетку или синцитий. Это происходит во время развития скелетных мышц, образующих крупные мышечные волокна . Позже это было подтверждено и в других тканях, например, в хрусталике глаза. Хотя в обоих случаях участвуют клеточные волокна, в случае глазного хрусталика слияние клеток более ограничено, что приводит к менее обширному слиянию стратифицированного синцития. [23]
Везикулы
[ редактировать ]
Связанные с липидными мембранами везикулы самых разных размеров обнаруживаются внутри и снаружи клеток и содержат огромное количество вещей: от пищи до вторгающихся организмов, от воды до сигнальных молекул. Использование электрического нервного импульса от нейрона нервно -мышечного соединения для стимуляции сокращения мышцы является примером очень малого [24] (около 0,05 мкм) везикулы, принимающие непосредственное участие в регуляции межклеточной коммуникации. Нейрон производит тысячи крошечных пузырьков, каждый из которых содержит тысячи сигнальных молекул. Один везикула высвобождается вблизи мышцы примерно каждую секунду во время отдыха. При активации нервным импульсом одновременно высвободятся более 100 везикул, сотни тысяч сигнальных молекул, вызывающих значительное сокращение мышечного волокна. Все это происходит за небольшую долю секунды.
Обычно небольшие пузырьки, используемые для транспортировки сигнальных молекул, высвобождаемых из клетки, называются экзосомами. [25] [26] [27] или просто внеклеточные везикулы (ВВ), [28] и помимо их важности для организма они также важны для биосенсоров . [24] Внеклеточные везикулы могут выделяться из злокачественных раковых клеток. Было показано, что эти внеклеточные везикулы содержат белки щелевых соединений, сверхэкспрессируемые в злокачественных клетках, которые распространяются на нераковые клетки, что, по-видимому, усиливает распространение злокачественного новообразования. [29] Везикулы также связаны с транспортировкой материалов за пределы клетки, обеспечивая рост и восстановление тканей во внеклеточном матриксе. [30] [31] В подобных ситуациях им могут быть присвоены специальные обозначения, например матричные везикулы (МВ).
Примеры более крупных везикул находятся в регуляторных секреторных путях в эндокринных , экзокринных тканях, [32] трансцитоз [33] [34] и везикуло-вакуолярная органелла (ВВО) в эндотелиальных и, возможно, других типах клеток. [35] Другая форма переноса кусочков мембраны вокруг соединений называется трансэндоцитозом. [36] Некоторые крупные межклеточные везикулы также остаются интактными, поскольку они транспортируют свое содержимое из одной части ткани в другую и включают в себя бляшки щелевых соединений. [37]
Коммуникация в нервной системе
[ редактировать ]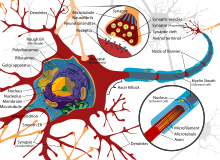

Когда мы думаем о межклеточной коммуникации, мы часто используем нашу нервную систему как точку отсчета. Нервы, состоящие из множества клеток у позвоночных, обычно высокоспециализированы по форме и функциям и обычно являются наиболее сложными в мозге . Они обеспечивают быструю и точную направленную связь между клетками на больших расстояниях, например, от мозга к руке. Нервные клетки можно рассматривать как посредников, которые не столько общаются друг с другом, сколько передают сообщения от одной соседней клетки к другой. Будучи «вспомогательными» клетками, передающими сообщение, они требуют дополнительного пространства и могут потреблять много энергии внутри организма. [38]
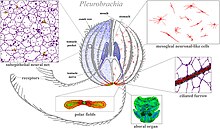
У более простых организмов, таких как губки и плакозойи, часто меньше пищи и, следовательно, меньше энергии. Их нервная система менее специализирована, и клетки, входящие в ее состав, должны выполнять и другие функции. [39]
Эфаптическая связь
[ редактировать ]Когда группы нервных клеток образуют другой тип межклеточной коммуникации, называемый эфаптической связью . Впервые он был определен количественно Кацем в 1940 году. [40] но было трудно связать какую-либо структуру или «эфапс» с этой формой коммуникации. Существуют редукционистские попытки связать определенные группы нервных клеток, демонстрирующих эфаптическую связь, с определенными функциями мозга. [41] Пока еще нет исследований простейших нейронных систем, таких как полярные тельца гребневиков, чтобы выяснить, может ли эфаптическая связь объяснить некоторые из их более сложных моделей поведения. [39]
Экосистемная межклеточная связь
[ редактировать ]

Определение биологической коммуникации непростое. [42] Ранние исследования в области клеточной биологии проводились на уровне клеток и организмов. Трудно было проследить, как отдельные клетки одного организма могут влиять на клетки другого, и это не представляло первостепенной важности. Если межклеточная коммуникация включает в себя передачу одной клеткой сигнала другой, чтобы вызвать ответ, межклеточная коммуникация не ограничивается клетками одного организма. Сообщается о межцарственной связи у растений на небольших расстояниях. [11] Размножение в воде часто включает в себя обширный синхронизированный выпуск гамет , называемый нерестом . [43] На больших расстояниях клетки одного растения будут взаимодействовать с клетками другого растения того же вида и других видов, выпуская в воздух сигналы, такие как летучие вещества зеленых листьев , которые могут, среди прочего, заранее предупреждать соседей о травоядных животных или в случае Газ этилен сигнал запускает созревание плодов. Межклеточная передача сигналов у растений также может происходить под землей с помощью микоризной сети , которая может связывать большие площади растений через грибковые сети, позволяя перераспределять ресурсы окружающей среды.
Глядя на колонии насекомых, таких как пчелы и муравьи, мы обнаружили феромоны. [44] Высвобождаемые из клеток одного организма в клетки другого организма, они могут координировать колонии, напоминая слизевики . Передача сигналов от клетки к клетке с использованием «феромонов» также была обнаружена у более сложных животных. По мере увеличения сложности растет и влияние сигналов. «Феромоны» у более сложных животных, таких как позвоночные, теперь правильнее называть «хемосигналами». [45] [46] [47] в том числе между видами. [48]

Идея о том, что межклеточная коммуникация между клетками внутри организма так же схожа, как и между клетками между разными организмами, даже добычей, продемонстрирована виннексином . [49] Этот белок представляет собой модифицированную форму белка иннексина, обнаруженного в гусенице. То есть виннексин очень похож на собственный иннексин гусеницы и мог быть получен только из невирусного иннексина каким-то неясным способом. Иннексин гусеницы образует нормальные межклеточные связи внутри гусеницы как часть иммунного ответа гусеницы на яйцо, имплантированное паразитической осой. Иннексин помогает обеспечить нейтрализацию яйца осы, спасая гусеницу от паразита. Так что же делает виннексин и как? Эволюция привела к появлению вируса, который общается с осами таким образом, что уклоняется от противовирусных реакций ос, позволяя вирусу жить и размножаться в яичниках ос. Когда оса впрыскивает свое яйцо в гусеницу-хозяина, в нее также попадает много вирусов из яичника осы. Вирусные частицы не реплицируются в клетках гусениц, а скорее взаимодействуют с генетическим механизмом гусениц, производя белок виннексин. Белок виннексин внедряется в клетки гусеницы, изменяя коммуникацию в гусенице, поэтому гусеница продолжает жить, но с измененным иммунным ответом. Виннексины способны смешиваться с обычными иннексинами, чтобы изменить общение внутри гусеницы, и, вероятно, так и происходит. Измененная коммуникация внутри гусеницы предотвращает отторжение защитными силами гусеницы яйца осы. В результате яйцо осы вылупляется, поглощает гусеницу и вирус от матери личинки осы и цикл повторяется. Видно, что вирус и оса необходимы друг другу и хорошо взаимодействуют друг с другом, позволяя вирусу жить и размножаться, но только неразрушающим образом внутри яичника осы. Вирус вводится оса в гусеницу, но вирус не размножается в гусенице, вирус только связывается с гусеницей, чтобы модифицировать ее несмертельным способом. Затем личинки осы будут медленно поедать гусеницу, не останавливаясь, при этом снова общаясь с вирусом, чтобы гарантировать, что у осы есть место в яичнике для повторного размножения. Коннексины/иннексины/виннексины, которые когда-то считались участвующими только в обеспечении пути для сигнальных молекул или электрических сигналов, теперь показали, что сами действуют как сигнальные молекулы.
Ссылки
[ редактировать ]- ^ Гук, Роберт (1665). Микрография: или некоторые физиологические описания мельчайших тел, сделанные с помощью лупы, с последующими наблюдениями и исследованиями . Королевское общество. п. 113.
- ^ Перейти обратно: а б с д Сливко-Колчик Георгий Александрович; Кузнецов Виктор П.; Панчин, Юрий В. (февраль 2019 г.). «Существуют ли щелевые соединения без коннексинов или паннексинов?» . Эволюционная биология BMC . 19 (S1): 46. дои : 10.1186/s12862-019-1369-4 . ПМК 6391747 . ПМИД 30813901 .
- ^ Нестле, Мэрион; Сассман, Морис (август 1972 г.). «Влияние циклического АМФ на морфогенез и накопление ферментов у Dictyostelium discoideum». Биология развития . 28 (4): 545–554. дои : 10.1016/0012-1606(72)90002-4 . ПМИД 4340352 .
- ^ Де Мейтс, Пьер; Рот, Джесси; Невилл, Дэвид М.; Гэвин, Джеймс Р.; Лесняк, Максин А. (ноябрь 1973 г.). «Взаимодействие инсулина с его рецепторами: экспериментальные доказательства отрицательной кооперативности». Связь с биохимическими и биофизическими исследованиями . 55 (1): 154–161. дои : 10.1016/S0006-291X(73)80072-5 . ПМИД 4361269 .
- ^ Галл, В. Эйнар; Эдельман, Джеральд М. (21 августа 1981 г.). «Боковая диффузия поверхностных молекул в клетках и тканях животных». Наука . 213 (4510): 903–905. Бибкод : 1981Sci...213..903G . дои : 10.1126/science.7196087 . ПМИД 7196087 .
- ^ Мендоса, Кристофер; Нагиди, Сай Харша; Коллетт, Кьетил; Макелл, Джейкоб; Мизрахи, Дарио (январь 2022 г.). «Кальций регулирует взаимодействие между плотным соединением и эпителиально-адгезионным соединением на плазматической мембране» . Письма ФЭБС . 596 (2): 219–231. дои : 10.1002/1873-3468.14252 . ПМИД 34882783 . S2CID 245028289 .
- ^ Фаркуар, Мэрилин Г.; Паладе, Джордж Э. (1 мая 1963 г.). «Соединительные комплексы в различных эпителиях» . Журнал клеточной биологии . 17 (2): 375–412. дои : 10.1083/jcb.17.2.375 . ПМК 2106201 . ПМИД 13944428 .
- ^ Д'ондт, Кэтлин; Понсартс, Раф; Де Смедт, Гумберт; Бултинк, Герт; Химпенс, Бернард (сентябрь 2009 г.). «Паннексины, дальние родственники семейства коннексинов со специфическими клеточными функциями?» . Биоэссе . 31 (9): 953–974. doi : 10.1002/bies.200800236 . ПМИД 19644918 . S2CID 10733461 .
- ^ Стар, Клаудия М.; Прието-Виллалобос, Джон; Марамбио-Руис, Люк; Балмазабаль, Ксавье; Альвеар, Танья Ф.; Вега, Матиас; Барра, Паола; Ретамал, Морис А.; Орельяна, Джон А.; Гомес, Гонсало И. (14 декабря 2022 г.). «Гипертоническая нефропатия: раскрытие возможного участия полуканалов и паннексонов» . Международный журнал молекулярных наук . 23 (24):15936.doi : 10.3390 /ijms232415936 . ПМЦ 9785367 . PMID 36555574 .
- ^ Боасса, Даниэла; Амбрози, Чинция; Цю, Фэн; Даль, Герхард; Гайетта, Гвидо; Сосинский, Джина (октябрь 2007 г.). «Каналы паннексина 1 содержат сайт гликозилирования, который направляет гексамер к плазматической мембране» . Журнал биологической химии . 282 (43): 31733–31743. дои : 10.1074/jbc.M702422200 . ПМИД 17715132 .
- ^ Перейти обратно: а б Ван, Мэнъин; Дин, Ральф А. (апрель 2020 г.). «Движение малых РНК внутри растений и грибов и между ними» . Молекулярная патология растений . 21 (4): 589–601. дои : 10.1111/mpp.12911 . ПМК 7060135 . ПМИД 32027079 .
- ^ Дауэс, Клинтон Дж.; Скотт, Флора М.; Боулер, Э. (ноябрь 1961 г.). «Световое и электронно-микроскопическое исследование клеточных стенок водорослей. I. Phaeophyta и Rhodophyta». Американский журнал ботаники . 48 (10): 925–934. дои : 10.2307/2439535 . JSTOR 2439535 .
- ^ Уэтерби, Р.; Квирк, HM (октябрь 1982 г.). «Тонкая структура образования вторичной ямочной связи между аллопаразитом красных водорослей Holmsella australis и его хозяином из красных водорослей Gracilaria Furcellata». Протоплазма . 110 (3): 166–176. дои : 10.1007/BF01283319 . S2CID 21177509 .
- ^ Цитовский, Виталий; Замбриски, Партиция (август 1991 г.). «Как нуклеиновые кислоты растительных вирусов перемещаются по межклеточным связям?». Биоэссе . 13 (8): 373–379. дои : 10.1002/bies.950130802 . ПМИД 1953699 . S2CID 11349048 .
- ^ Декер, Роберт С.; Друг, Дэниел С. (1 июля 1974 г.). «Сборка щелевых соединений во время нейруляции амфибий» . Журнал клеточной биологии . 62 (1): 32–47. дои : 10.1083/jcb.62.1.32 . ПМК 2109180 . ПМИД 4135001 .
- ^ Левенштейн, WR (14 июля 1966 г.). «Проницаемость мембранных соединений». Анналы Нью-Йоркской академии наук . 137 (2): 441–72. Бибкод : 1966NYASA.137..441L . дои : 10.1111/j.1749-6632.1966.tb50175.x . ПМИД 5229810 . S2CID 22820528 .
- ^ Фуршпан, Э.Дж.; Поттер, Д.Д. (август 1957 г.). «Механизм передачи нервного импульса в синапсах рака». Природа . 180 (4581): 342–343. Бибкод : 1957Natur.180..342F . дои : 10.1038/180342a0 . ПМИД 13464833 . S2CID 4216387 .
- ^ Бэйлор, округ Колумбия; Николлс, Дж. Г. (1 августа 1969 г.). «Химические и электрические синаптические связи между кожными механорецепторными нейронами центральной нервной системы пиявки» . Журнал физиологии . 203 (3): 591–609. doi : 10.1113/jphysicalol.1969.sp008881 . ПМЦ 1351532 . ПМИД 4319015 .
- ^ Харрис, Эндрю Л. (3 декабря 2018 г.). «Электрическая муфта и ее каналы» . Журнал общей физиологии . 150 (12): 1606–1639. дои : 10.1085/jgp.201812203 . ПМК 6279368 . ПМИД 30389716 .
- ^ Ван, Сянчуань; Ху, Бойи; Чжао, Чжунъин; Цзе, Ю Чунг (июль 2022 г.). «От первичных половых клеток к сперматидам Caenorhabditis elegans» . Семинары по клеточной биологии и биологии развития . 127 : 110–120. дои : 10.1016/j.semcdb.2021.12.005 . ПМИД 34930663 . S2CID 245291439 .
- ^ Андраде, Вирджиния; Эшар, Арно (24 ноября 2022 г.). «Механика и регуляция цитокинетического отторжения» . Границы клеточной биологии и биологии развития . 10 : 1046617. doi : 10.3389/fcell.2022.1046617 . ПМЦ 9730121 . ПМИД 36506096 .
- ^ Зани, Бретт Г.; Индольфи, Лаура; Эдельман, Элазер Р. (28 января 2010 г.). «Тубулярные мосты для миграции и связи бронхиальных эпителиальных клеток» . ПЛОС ОДИН . 5 (1): е8930. Бибкод : 2010PLoSO...5.8930Z . дои : 10.1371/journal.pone.0008930 . ПМК 2812493 . ПМИД 20126618 .
- ^ Ши, Янжун; Бартон, Келли; Де Мария, Алисия; Петраш, Дж. Марк; Шилс, Алан; Басснетт, Стивен (15 мая 2009 г.). «Многослойный синцитий хрусталика позвоночных» . Журнал клеточной науки . 122 (10): 1607–1615. дои : 10.1242/jcs.045203 . ПМК 2680101 . ПМИД 19401333 .
- ^ Перейти обратно: а б Хатамие, Амир; Он, Сюлань; Чжан, Синь-Вэй; Оомен, Питер Э.; Юинг, Эндрю Г. (январь 2023 г.). «Достижения в области нано/микромасштабных электрохимических датчиков и биосенсоров для анализа одиночных везикул, ключевых наноразмерных органелл в клеточной коммуникации» . Биосенсоры и биоэлектроника . 220 : 114899. дои : 10.1016/j.bios.2022.114899 . ПМИД 36399941 . S2CID 253476056 .
- ^ Дензер, К.; Клеймер, МЮ; Хейнен, ХФ; Стурфогель, В.; Гёз, HJ (1 октября 2000 г.). «Экзосома: от внутреннего пузырька мультивезикулярного тела к межклеточному сигнальному устройству». Журнал клеточной науки . 113 (19): 3365–3374. дои : 10.1242/jcs.113.19.3365 . ПМИД 10984428 .
- ^ ван Ниль, Гийом; Порту-Каррейро, Изабель; Симоэс, Сабрина; Рапосо, Граса (1 июля 2006 г.). «Экзосомы: общий путь выполнения специализированной функции» . Журнал биохимии . 140 (1): 13–21. дои : 10.1093/jb/mvj128 . ПМИД 16877764 .
- ^ Дондт, Берт; Руссо, Квентин; Де Вевер, Оливье; Хендрикс, Ан (1 сентября 2016 г.). «Функция внеклеточных везикуло-ассоциированных микроРНК при метастазировании» . Исследования клеток и тканей . 365 (3): 621–641. дои : 10.1007/s00441-016-2430-x . ПМИД 27289232 . S2CID 253969773 .
- ^ Дехганбанадаки, Ходжат; Форузанфар, Катаюн; Какаи, Ардешир; Зейди, Самане; Салехи, Негар; Арджманд, Бабак; Рази, Фариде; Хашеми, Эхсан (1 апреля 2022 г.). «Роль мРНК CDH2 и MCP-1 внеклеточных везикул крови в прогнозировании ранней стадии диабетической нефропатии» . ПЛОС ОДИН . 17 (4): e0265619. Бибкод : 2022PLoSO..1765619D . дои : 10.1371/journal.pone.0265619 . ПМЦ 8975111 . ПМИД 35363774 .
- ^ Акунья, Родриго А.; Варас-Годой, Мануэль; Берту, Вивиана М.; Альфаро, Иван Евгеньевич; Ретамал, Маурисио А. (28 апреля 2020 г.). «Коннексин-46, содержащийся во внеклеточных везикулах, усиливает признаки злокачественности клеток рака молочной железы» . Биомолекулы . 10 (5): 676. doi : 10.3390/biom10050676 . ПМЦ 7277863 . ПМИД 32353936 .
- ^ Нараускайте, Д; Видмантайте, Г; Рустейкайте, Ю; Сампат, Р; Рудайтете, А; Сташите, Г; Апарисио Кальвенте, Мичиган; Екабсоне, А. (18 августа 2021 г.). «Внеклеточные везикулы в заживлении кожных ран» . Фармацевтика (Базель, Швейцария) . 14 (8): 811. doi : 10.3390/ph14080811 . ПМЦ 8400229 . ПМИД 34451909 .
- ^ Андерсон, ХК; Сесил, Р; Сайдера, SW (май 1975 г.). «Кальцификация рахитического хряща крысы in vitro везикулами внеклеточного матрикса» . Американский журнал патологии . 79 (2): 237–54. ЧВК 1912651 . ПМИД 1146961 .
- ^ Идзуми, Тетсуро; Гоми, Хироши; Касаи, Кадзуо; Мизутани, Шин; Тории, Сейджи (2003). «Роль Rab27 и его эффекторов в регулируемых секреторных путях» . Структура и функции клеток . 28 (5): 465–474. дои : 10.1247/csf.28.465 . ПМИД 14745138 .
- ^ Симионеску, Майя; Гафенку, Анка; Антоэ, Фелисия (1 июня 2002 г.). «Транцитоз макромолекул плазмы в эндотелиальных клетках: клеточный биологический обзор». Микроскопические исследования и техника . 57 (5): 269–288. дои : 10.1002/jemt.10086 . ПМИД 12112439 . S2CID 28337130 .
- ^ Геберт, А; Гоке, М; Роткоттер, Х.Дж.; Дитрих, Chr F (октябрь 2000 г.). «Механизмы захвата антигена в тонком и толстом кишечнике: роль М-клеток в инициации иммунных ответов*». Журнал гастроэнтерологии . 38 (10): 855–872. дои : 10.1055/с-2000-10001 . ПМИД 11089271 . S2CID 83474242 .
- ^ Дворжак, Энн М.; Фэн, Диан (апрель 2001 г.). «Везикуло-вакуолярная органелла (ВВО): новая органелла, проницаемая для эндотелиальных клеток» . Журнал гистохимии и цитохимии . 49 (4): 419–431. дои : 10.1177/002215540104900401 . ПМИД 11259444 . S2CID 29246235 .
- ^ Сакураи, Такаши; Вуллс, Мелисса Дж.; Джин, Сук-Вон; Мураками, Масахиро; Саймонс, Майкл (6 марта 2014 г.). «Межклеточный обмен клеточными компонентами посредством VE-кадгерин-зависимого трансэндоцитоза» . ПЛОС ОДИН . 9 (3): е90736. Бибкод : 2014PLoSO...990736S . дои : 10.1371/journal.pone.0090736 . ПМЦ 3946293 . ПМИД 24603875 .
- ^ Грюйтерс, В. (2003). «Участвуют ли мембранные бляшки щелевых соединений в межклеточном переносе везикул?». Международная клеточная биология . 27 (9): 711–717. дои : 10.1016/s1065-6995(03)00140-9 . ПМИД 12972275 . S2CID 37315556 .
- ^ Альборг, Гунвор; Варен, Дж. (январь 1972 г.). «Использование субстрата мозга во время длительных упражнений». Скандинавский журнал клинических и лабораторных исследований . 29 (4): 397–402. дои : 10.3109/00365517209080256 . ПМИД 21488407 .
- ^ Перейти обратно: а б Мороз Леонид Л.; Романова, Дарья Ю. (23 декабря 2022 г.). «Альтернативные нейронные системы: Что такое нейрон? (гребневики, губки и плакозойи)» . Границы клеточной биологии и биологии развития . 10 : 1071961. дои : 10.3389/fcell.2022.1071961 . ПМЦ 9816575 . ПМИД 36619868 .
- ^ Кац, Бернхард; Шмитт, Отто Х. (14 февраля 1940 г.). «Электрическое взаимодействие между двумя соседними нервными волокнами» . Журнал физиологии . 97 (4): 471–488. дои : 10.1113/jphysicalol.1940.sp003823 . ПМЦ 1393925 . ПМИД 16995178 .
- ^ Мартинес-Банаклоча, Маркос (февраль 2020 г.). «Астроглиальная изопотенциальность и влияние биомагнитного поля, связанного с кальцием, на соединение кортикальных нейронов» . Клетки . 9 (2): 439. doi : 10.3390/cells9020439 . ПМК 7073214 . ПМИД 32069981 .
- ^ Скотт-Филлипс, TC (март 2008 г.). «Определение биологической связи» . Журнал эволюционной биологии . 21 (2): 387–395. дои : 10.1111/j.1420-9101.2007.01497.x . ПМИД 18205776 . S2CID 5014169 .
- ^ Харрисон, Польша; Бэбкок, Колорадо; Булл, Грузия; Оливер, Дж. К.; Уоллес, CC; Уиллис, БЛ (16 марта 1984 г.). «Массовое нерест в кораллах тропических рифов». Наука . 223 (4641): 1186–9. Бибкод : 1984Sci...223.1186H . дои : 10.1126/science.223.4641.1186 . ПМИД 17742935 . S2CID 31244527 .
- ^ Ренье, FE; Ло, Дж. Х. (сентябрь 1968 г.). «Феромоны насекомых» . Журнал исследований липидов . 9 (5): 541–51. дои : 10.1016/S0022-2275(20)42699-9 . ПМИД 4882034 .
- ^ Доти, Ричард Л. (2010). Великий миф о феромонах . Балтимор, Мэриленд, США: Издательство Университета Джона Хопкинса.
- ^ Ридделл, П; Париж, MCJ; Джуне, CJ; Пейджет, П; Париж, DBBP (27 мая 2021 г.). «Умиротворение феромонов для управления стрессом и агрессией во время сохранения диких псовых: может ли решение быть прямо у нас под носом?» . Животные . 11 (6): 1574. дои : 10.3390/ani11061574 . ПМК 8230031 . ПМИД 34072227 .
- ^ Йе, Ютинг; Лу, Чжунхуа; Чжоу, Вэнь (2021). «Влияние феромонов на гипоталамус человека в зависимости от сексуальной ориентации и пола». Гипоталамус человека: нервно-психические расстройства . Справочник по клинической неврологии. Том. 182. стр. 293–306. дои : 10.1016/B978-0-12-819973-2.00021-6 . ISBN 9780128199732 . ПМИД 34266600 . S2CID 235962401 .
- ^ Кальви, Э; Квассоло, Ю; Массайя, М; Скандурра, А; Д'Аньелло, Б; Д'Амелио, П. (май 2020 г.). «Запах эмоций: систематический обзор внутри- и межвидовой химической коммуникации эмоций человека» . Мозг и поведение . 10 (5): e01585. дои : 10.1002/brb3.1585 . ПМЦ 7218249 . ПМИД 32212329 .
- ^ Хасегава, Дания; Тернбулл, Миссури (17 апреля 2014 г.). «Последние открытия в эволюции и функциях иннексинов насекомых». Письма ФЭБС . 588 (8): 1403–10. дои : 10.1016/j.febslet.2014.03.006 . ПМИД 24631533 . S2CID 25970503 .