Ценорабдитис элегантный
| Ценорабдитис элегантный | |
|---|---|
 | |
| Взрослый -гермафродит C. elegans червь | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Нематоды |
| Сорт: | Хромадория |
| Заказ: | Рабдитида |
| Семья: | Рабдитиды |
| Род: | Ценорхабдитис |
| Разновидность: | К. Элеганс |
| Биномиальное имя | |
| Ценорабдитис элегантный | |
| Подвиды | |
Caenorhabditis elegans ( / ˌ s iː n oʊ r æ b ˈ d aɪ t ə s ˈ ɛ l ə ɡ æ n s / [6] ) — свободноживущая прозрачная нематода длиной около 1 мм. [7] обитающий в почвах умеренного пояса. Это типовой вид своего рода. [8] Название представляет собой смесь греческих слов caeno- (недавний) и рабдитис (палочковидный). [9] и латинское elegans (элегантный). В 1900 году Мопас первоначально назвал его Rhabditides elegans. Оше поместил его в подрод Caenorhabditis в 1952 году, а в 1955 году Догерти повысил Caenorhabditis до статуса рода . [10]
C. elegans представляет собой несегментированный псевдоцеломат , у которого отсутствуют дыхательная и кровеносная системы. [11] Большинство этих нематод — гермафродиты , некоторые — самцы. [12] У самцов есть специализированные хвосты для спаривания, включающие спикулы .
В 1963 году Сидней Бреннер предложил исследовать C. elegans, прежде всего в области развития нейронов. В 1974 году он начал исследования в области молекулярной биологии и биологии развития , C. elegans которая с тех пор широко использовалась в качестве модельного организма . [13] Это был первый многоклеточный организм которого , весь геном был секвенирован , а в 2019 году [14] это был первый организм, у которого был завершен коннектом (нейронная «схема подключения»). [15] [16] [17]
Анатомия
[ редактировать ]
C. elegans несегментированный червеобразный , . , симметричный двусторонне - Он имеет кутикулу (прочный внешний покров, напоминающий экзоскелет ), четыре основных эпидермальных тяжа и заполненный жидкостью псевдоцелом (полость тела). У него также есть некоторые из тех же систем органов, что и у более крупных животных. Примерно одна особь из тысячи — мужчина, остальные — гермафродиты. [18] Базовая анатомия C. elegans включает рот, глотку , кишечник , гонаду и коллагеновую кутикулу. Как и все нематоды, они не имеют ни кровеносной, ни дыхательной системы. Четыре группы мышц, проходящие по всей длине тела, соединены с нервной системой, которая позволяет мышцам перемещать тело животного только в виде дорсального или вентрального изгиба, но не влево или вправо, за исключением головы, где четыре мышцы квадранты подключаются независимо друг от друга. Когда волна дорсальных/вентральных мышечных сокращений распространяется от спины к передней части животного, животное отбрасывается назад. Когда волна сокращений начинается спереди и распространяется назад вдоль тела, животное продвигается вперед. Из-за этого дорсально-вентрального смещения в изгибах тела любой нормальный живой, движущийся человек имеет тенденцию лежать либо на левом, либо на правом боку, когда он наблюдает за пересечением горизонтальной поверхности. Считается, что набор гребней на боковых сторонах кутикулы тела, крыльев, придает животному дополнительное сцепление во время этих изгибающих движений.



Что касается липидного обмена, у C. elegans нет каких-либо специализированных жировых тканей, поджелудочной железы , печени или даже крови для доставки питательных веществ по сравнению с млекопитающими. Вместо этого нейтральные липиды хранятся в кишечнике, эпидермисе и эмбрионах. Эпидермис депо соответствует адипоцитам млекопитающих, поскольку является основным триглицеридов . [19]
Глотка представляет собой мышечный пищевой насос в голове C. elegans , имеющей треугольную форму в поперечном сечении. Это измельчает пищу и транспортирует ее прямо в кишечник. Набор «клапанных клеток» соединяет глотку с кишечником, но как работает этот клапан, непонятно. После пищеварения содержимое кишечника выделяется через прямую кишку, как и у всех других нематод. [20] Между глоткой и выводным каналом, выполняющим функцию выделения жидкой мочи, не существует прямой связи.
У самцов есть однолопастная гонада, семявыносящий проток и хвост, предназначенный для спаривания, со спикулами . Гермафродиты имеют два яичника , яйцеводы и сперматеку , а также одну матку .
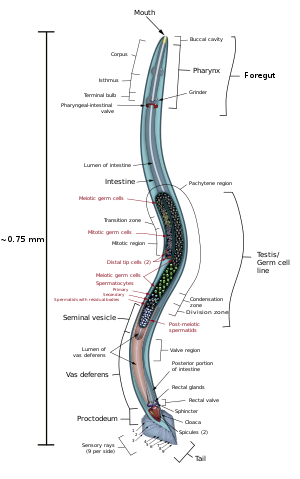
У C. elegans 302 нейрона, что составляет примерно треть всех соматических клеток во всем организме. [21] Многие нейроны содержат дендриты, которые выходят из клетки для получения нейротрансмиттеров или других сигналов, а также отросток , который распространяется на нервное кольцо («мозг») для синаптической связи с другими нейронами. [22] C. elegans имеет возбуждающие холинергические и тормозные ГАМКергические мотонейроны, которые соединяются с мышцами стенок тела и регулируют движение. Кроме того, эти нейроны и другие нейроны, такие как интернейроны, используют различные нейротрансмиттеры для управления поведением. [23]
Хорошие гранулы
[ редактировать ]
В кишечнике C. elegans присутствуют многочисленные кишечные гранулы , функции которых до сих пор полностью не известны, как и многие другие аспекты этой нематоды, несмотря на многие годы ее изучения. Эти кишечные гранулы встречаются во всех отрядах Rhabditida. Они очень похожи на лизосомы в том, что имеют кислую внутреннюю часть и способность к эндоцитозу , но они значительно крупнее, что подтверждает представление о том, что они являются запасающими органеллами.Особенностью гранул является то, что при наблюдении за ними в ультрафиолетовом свете они реагируют, излучая интенсивную синюю флуоресценцию . Другое наблюдаемое явление называется «флуоресценцией смерти». Когда черви умирают, испускается резкий всплеск синей флуоресценции. Эта смертельная флуоресценция обычно имеет место в виде передне-задней волны, которая движется вдоль кишечника, и наблюдается как у молодых, так и у старых червей, независимо от того, подверглись ли они смертельной травме или мирно умирают от старости.
О функциях кишечных гранул было выдвинуто множество теорий, причем более ранние теории были опровергнуты более поздними открытиями. Считается, что одна из их функций — хранение цинка. Недавний химический анализ выявил содержащийся в них синий флуоресцентный материал как гликозилированную форму антраниловой кислоты (АК). Необходимость в больших количествах АК, содержащихся во многих кишечных гранулах, подвергается сомнению. Одна из возможностей заключается в том, что АК обладает антибактериальным действием и используется для защиты от проникновения патогенов. Другая возможность заключается в том, что гранулы обеспечивают фотозащиту; Всплески флуоресценции АА влекут за собой преобразование вредного ультрафиолетового света в относительно безвредный видимый свет. Это рассматривается как возможная связь меланинсодержащими с меланосомами . [24]
Воспроизведение
[ редактировать ]Червь-гермафродит считается специализированной формой самоплодной самки, поскольку его сома женская. Гермафродитная зародышевая линия сначала производит мужские гаметы , а после внутреннего оплодотворения откладывает яйца через матку. Гермафродиты производят всю свою сперму на стадии L4 (150 сперматозоидов на гонадное плечо), а затем производят только ооциты . Гермафродитная гонада действует как яйцеклетка, при этом сперматозоиды хранятся в той же области гонады, что и ооциты, до тех пор, пока первый ооцит не выталкивает сперму в сперматеку ( камеру, в которой ооциты оплодотворяются спермой). [25]
Самец может оплодотворить гермафродита, который предпочтительно будет использовать мужскую сперму (оба типа спермы хранятся в сперматеке).

Сперматозоиды C. elegans амебоидные, без жгутиков и акросом . [27] При самооплодотворении червь дикого типа откладывает около 300 яиц. При оплодотворении самцом число потомков может превышать 1000. Гермафродиты обычно не спариваются с другими гермафродитами. При 20 °C лабораторный штамм C. elegans (N2) имеет среднюю продолжительность жизни около 2–3 недель и время генерации от 3 до 4 дней.
C. elegans имеет пять пар аутосом и одну пару половых хромосом . Пол у C. elegans основан на системе определения пола X0 . Гермафродиты C. elegans имеют совпадающую пару половых хромосом (XX); редкие самцы имеют только одну половую хромосому (X0).
Определение пола
[ редактировать ]C. elegans в основном являются гермафродитными организмами, производящими как сперматозоиды, так и ооциты . [28] Самцы встречаются в популяции примерно с частотой 1 на 200 гермафродитов, но оба пола сильно дифференцированы. [29] [30] Самцы отличаются от своих собратьев-гермафродитов тем, что они меньше по размеру, и их можно идентифицировать по форме хвоста. [30] C.elegans размножаются посредством процесса, называемого андродиоцией . Это означает, что они могут размножаться двумя способами: либо путем самооплодотворения у гермафродитов, либо путем спаривания гермафродитов с самцами. Самцы образуются в результате нерасхождения Х-хромосом во время мейоза. Черви, которые размножаются путем самооплодотворения, подвержены риску высокого неравновесия по сцеплению , что приводит к снижению генетического разнообразия в популяциях и увеличению накопления вредных аллелей. [31] У C. elegans определение соматического пола связано с геном tra-1 . [32] tra -1 представляет собой ген в пути определения пола транскрипционного фактора TRA-1, который регулируется посттранскрипционно и способствует развитию женщин. [32] У гермафродитов (ХХ) наблюдается высокий уровень активности тра-1 , который продуцирует женскую репродуктивную систему и тормозит развитие самцов. [28] [32] В определенный момент жизненного цикла, за день до взросления, гермафродитов можно идентифицировать по добавлению вульвы возле хвоста. У мужчин (XO) наблюдается низкий уровень активности tra-1 , что приводит к нарушению мужской репродуктивной системы. [32] Недавние исследования показали, что существуют три других гена: fem-1, fem-2 и fem-3, которые негативно регулируют путь TRA-1 и действуют как окончательный определяющий пол у C. elegans . [28]
Эволюция
[ редактировать ]Система определения пола у C. elegans — тема, которая уже много лет интересует ученых. [29] Поскольку они используются в качестве модельного организма, любая обнаруженная информация о том, как могла развиваться их система определения пола, может способствовать таким же исследованиям эволюционной биологии на других организмах. После почти 30 лет исследований учёные начали собирать кусочки эволюции такой системы. [29] Они обнаружили, что здесь задействован сложный путь, имеющий несколько уровней регуляции. [29] Близкородственный организм Caenorhabditis briggsae был тщательно изучен, и вся последовательность его генома помогла собрать воедино недостающие части в эволюции определения пола C. elegans . [29] Было обнаружено, что два гена ассимилировались, в результате чего белки XOL-1 и MIX-1 оказали влияние на определение пола у C. elegans . также [29] Мутации в пути XOL-1 приводят к феминизации C. elegans. [33] Известно, что ген mix-1 гипоактивирует Х-хромосому и регулирует морфологию мужского хвоста у C. elegans. [34] Глядя на нематод в целом, можно сказать, что мужской пол и пол гермафродита, вероятно, произошли в результате параллельной эволюции. [29] Параллельная эволюция определяется как сходные черты, развивающиеся от предка в аналогичных условиях; Проще говоря, эти два вида со временем развиваются одинаковым образом. Примером этого могут быть сумчатые и плацентарные млекопитающие. Ученые также выдвинули гипотезу, что бесполое размножение гермафродитов, или «самоопыление», могло эволюционировать конвергентно, изучая виды, похожие на C. elegans. [29] Другие исследования эволюции определения пола показывают, что гены, связанные со спермой, развиваются быстрее, чем женские гены. [35] Однако гены сперматозоидов на Х-хромосоме имеют сниженную скорость эволюции. Гены сперматозоидов имеют короткие кодирующие последовательности, высокую систематическую ошибку кодонов и непропорциональное представительство среди генов-сирот . [35] Эти характеристики генов сперматозоидов могут быть причиной их высоких темпов эволюции, а также могут указывать на то, как гены сперматозоидов развились из червей-гермафродитов. В целом у учёных есть общее представление о пути определения пола у C. elegans , однако эволюция того, как возник этот путь, ещё не чётко определена.
Разработка
[ редактировать ]
Эмбриональное развитие
[ редактировать ]Оплодотворенная зигота подвергается ротационному голобластическому дроблению .
Попадая сперматозоида в ооцит, начинается формирование передне-задней оси. [36] спермы Центр организации микротрубочек направляет движение пронуклеуса спермия к будущему заднему полюсу эмбриона, а также стимулирует движение белков PAR , группы факторов цитоплазматической детерминации, в соответствующие места. [37] В результате разницы в распределении белков PAR первое деление клеток является крайне асимметричным . [38] C. elegans Эмбриогенез является одним из наиболее изученных примеров асимметричного деления клеток. [39]
Все клетки зародышевой линии возникают из единственной первичной зародышевой клетки , называемой клеткой P4 , возникшей на ранних стадиях эмбриогенеза . [40] [41] Эта первичная клетка делится, образуя два предшественника зародышевой линии, которые делятся дальше только после вылупления. [41]
Формирование оси
[ редактировать ]Образующиеся дочерние клетки первого клеточного деления называются клеткой AB (содержащей PAR-6 и PAR-3) и клеткой P1 (содержащей PAR-1 и PAR-2). Второе деление клеток производит клетки ABp и ABa из клетки AB, а также клетки EMS и P2 из клетки P1. Это деление устанавливает дорсально-вентральную ось, при этом клетка ABp образует дорсальную сторону, а клетка EMS маркирует вентральную сторону. [42] Посредством передачи сигналов Wnt клетка P2 дает команду EMS-клетке делиться вдоль передне-задней оси. [43] Посредством передачи сигналов Notch клетка P2 дифференциально специфицирует клетки ABp и ABa, что дополнительно определяет дорсально-вентральную ось. Ось лево-право также становится очевидной на ранних стадиях эмбриогенеза, хотя неясно, когда именно эта ось определяется. Однако большинство теорий развития LR-оси предполагают некоторые различия в клетках, происходящих из AB-клетки. [44]
Гаструляция
[ редактировать ]Гаструляция происходит после того, как эмбрион достигает стадии 24 клеток. [45] C. elegans — разновидность протостомов , поэтому бластопор в конечном итоге образует рот. Инволюция в бластопор начинается с перемещения клеток энтодермы и последующего формирования кишечника, за которым следуют предшественники зародышевой линии P4 и, наконец, клетки мезодермы , включая клетки, которые в конечном итоге формируют глотку. Гаструляция заканчивается, когда эпиболия гипобластов закрывает бластопор. [46]
Постэмбриональное развитие
[ редактировать ]

условиях окружающей среды В благоприятных для размножения вылупившиеся личинки развиваются через четыре личиночные стадии — L1, L2, L3 и L4 — всего за 3 дня при 20 °C. В стрессовых условиях, например, при нехватке пищи, чрезмерной плотности популяции или высокой температуре, C. elegans может перейти в альтернативную третью личиночную стадию, L2d, называемую стадией дауера ( Dauer в переводе с немецкого означает «постоянный»). Вхождение в состояние дауэра регулируется особым феромоном дауэра. Этот феромон состоит из аналогичных производных 3,6-дидезоксисахара, аскарилозы . Аскарозиды, названные в честь основания аскарилозы, участвуют во многих половых и социальных формах поведения. [47] Таким образом, они составляют химический язык, который C. elegans использует для модуляции различных фенотипов. Личинки Дауэра стрессоустойчивы; они тонкие, их рот закрыт характерной твёрдой кутикулой и не может принимать пищу. Они могут оставаться на этой стадии в течение нескольких месяцев. [48] [49] Стадия заканчивается, когда условия улучшаются, благоприятствуя дальнейшему росту личинки, которая теперь переходит на стадию L4, хотя развитие гонад останавливается на стадии L2. [50]
Каждый переход между стадиями отмечен линькой прозрачной кутикулы червя. Переходы через эти стадии контролируются генами гетерохронного пути — эволюционно консервативным набором регуляторных факторов. [51] Многие гетерохронные гены кодируют микроРНК , которые подавляют экспрессию гетерохронных факторов транскрипции и других гетерохронных микроРНК. [52] микроРНК были первоначально обнаружены у C. elegans. [53] Важные события развития, контролируемые гетерохронными генами, включают деление и возможное синцитиальное слияние клеток подкожного шва и их последующую секрецию крыльев у молодых взрослых. Считается, что гетерохронный путь представляет собой эволюционно консервативного предшественника циркадных часов . [54]
Некоторые нематоды имеют фиксированное, генетически детерминированное количество клеток — явление, известное как эвтелия . Взрослый гермафродит C. elegans имеет 959 соматических клеток, а самец - 1033 клетки. [55] [56] [57] хотя было высказано предположение, что количество их кишечных клеток может увеличиться от одной до трех в ответ на кишечные микробы, с которыми сталкиваются матери. [58] В большей части литературы число клеток у мужчин описывается как 1031, но открытие пары левого и правого нейронов MCM увеличило это число на два в 2015 году. [57] Число клеток не меняется после прекращения клеточного деления в конце личиночного периода, а последующий рост происходит исключительно за счет увеличения размеров отдельных клеток. [59]
Экология
[ редактировать ]Различные виды Caenorhabditis обитают в различных средах, богатых питательными веществами и бактериями. Они питаются бактериями, развивающимися в разлагающихся органических веществах ( микробиворы ). Они обладают хемосенсорными рецепторами, которые позволяют обнаруживать бактерии и секретируемые бактериями метаболиты (такие как сидерофоры железа), чтобы они могли мигрировать к своей бактериальной добыче. [60] В почве недостаточно органического вещества для поддержания самоподдерживающихся популяций. C. elegans может выжить на диете, состоящей из множества бактерий, но его дикая экология в значительной степени неизвестна. Большинство лабораторных штаммов были взяты из искусственных сред, таких как сады и компостные кучи . Совсем недавно было обнаружено, что C. elegans процветает в других видах органических веществ, особенно в гниющих фруктах. [61] C. elegans также может поглощать загрязняющие вещества, особенно крошечные нанопластики, что может способствовать ассоциации с устойчивыми к антибиотикам бактериями, что приводит к распространению нанопластиков и устойчивых к антибиотикам бактерий C. elegans по почве. [62]
C. elegans также может использовать различные виды дрожжей , включая Cryptococcus laurentii и C. kuetzingii , в качестве единственного источника пищи. [63] Несмотря на то, что является бактериофагом , C. elegans он может быть уничтожен рядом патогенных бактерий, включая патогены человека, такие как Staphylococcus aureus , [64] синегнойная палочка , [65] Salmonella enterica или Enterococcus faecalis . [66] Патогенные бактерии также могут образовывать биопленки, липкая экзополимерная матрица которых может препятствовать C. elegans. подвижности [67] и скрывает хемоаттрактанты, чувствительные к бактериальному кворуму, от обнаружения хищниками. [68]
Беспозвоночные, такие как многоножки , насекомые , равноногие и брюхоногие моллюски , могут переносить личинок дауэра в различные подходящие места. Также было замечено, что личинки питаются своими хозяевами после смерти. [69]
Нематоды могут пережить высыхание , и у C. elegans механизм этой способности, как было показано, заключается в обилии белков позднего эмбриогенеза . [70]
C. elegans , как и другие нематоды, могут поедать хищные нематоды и другие всеядные животные, включая некоторых насекомых. [71]
Вирус Орсе — вирус, поражающий C. elegans , а также вирус Caenorhabditis elegans Cer1. [72] и вирус Caenorhabditis elegans Cer13 .
- Взаимодействие с грибами
Дикие изоляты Caenorhabditis elegans регулярно обнаруживаются при инфекциях грибами Microsporidia . Один из таких видов, Nematocida parisii , размножается в кишечнике C. elegans . [73]
Arthrobotrys oligospora — модельный организм для взаимодействия грибов и нематод. [74] Это наиболее распространенный и распространенный гриб, захватывающий нематод.
Использование в качестве модельного организма
[ редактировать ]В 1963 году Сидней Бреннер предложил использовать C. elegans в качестве модельного организма для изучения, в первую очередь, развития нервной системы у животных. Это один из простейших организмов, имеющих нервную систему . Нейроны не запускают потенциалы действия и не экспрессируют потенциалзависимые натриевые каналы . [75] У гермафродита эта система состоит из 302 нейронов. [76] структура которого была тщательно нанесена на карту, [14] в так называемом коннектоме , [77] и показано, что это сеть маленького мира . [78]
Исследования изучили нервные и молекулярные механизмы, которые контролируют некоторые виды поведения C. elegans , включая хемотаксис , термотаксис , механотрансдукцию , обучение , память и поведение при спаривании . [79] В 2019 году коннектом самца был опубликован с использованием техники, отличной от той, которая использовалась для гермафродита. В той же статье новая техника использовалась для переделки коннектома гермафродита, обнаружив 1500 новых синапсов. [80]
Его использовали в качестве модельного организма для изучения молекулярных механизмов метаболических заболеваний. [81] Бреннер также выбрал его, поскольку его легко выращивать в больших популяциях и он удобен для генетического анализа. [82] Это многоклеточный эукариотический организм, но достаточно простой, чтобы его можно было изучить очень подробно. Прозрачность C. elegans облегчает изучение клеточной дифференцировки и других процессов развития в интактном организме. Спикулы у самца четко отличают самцов от самок. Штаммы дешевы в разведении и могут быть заморожены. При последующем размораживании они остаются жизнеспособными, что позволяет хранить их в течение длительного времени. [13] Обслуживание проще по сравнению с другими многоклеточными модельными организмами. Несколько сотен нематод можно содержать на одной чашке с агаром и подходящей питательной средой. Бреннер описал использование мутанта кишечной палочки – OP50. OP50 — это организм, нуждающийся в урациле , и его дефицит в пластинке предотвращает чрезмерный рост бактерий, которые могут скрыть червей. [83] Использование OP50 не требует каких-либо серьезных мер лабораторной безопасности, поскольку он непатогенен и легко выращивается в среде Лурия-Бертани (LB) в течение ночи. [84]
Картирование клеточного происхождения
[ редактировать ]судьба развития каждой отдельной соматической клетки (959 у взрослого гермафродита; 1031 у взрослого мужчины). Была картирована [85] [86] Эти паттерны клеточного происхождения в значительной степени инвариантны у разных особей, тогда как у млекопитающих развитие клеток в большей степени зависит от клеточных сигналов эмбриона.
Как упоминалось ранее, первые клеточные деления раннего эмбриогенеза у C. elegans являются одними из наиболее изученных примеров асимметричных клеточных делений , а червь является очень популярной модельной системой для изучения биологии развития. [39]
Запрограммированная гибель клеток
[ редактировать ]Запрограммированная гибель клеток ( апоптоз ) уничтожает множество дополнительных клеток (131 у гермафродита, большинство из которых в противном случае стали бы нейронами ); эта «предсказуемость апоптоза» способствовала выяснению некоторых апоптозных генов . Были идентифицированы гены, способствующие гибели клеток, и один ингибитор клеточной гибели. [87]
РНК-интерференция и молчание генов
[ редактировать ]
РНК-интерференция (РНКи) — относительно простой метод нарушения функции определенных генов. Замалчивание функции гена иногда может позволить исследователю сделать вывод о его возможной функции. Нематоду можно замачивать, инъецировать, [88] или кормят генетически трансформированными бактериями, которые экспрессируют интересующую двухцепочечную РНК, последовательность которой дополняет последовательность гена, который исследователь хочет отключить. [89] РНКи стала мощным инструментом в изучении функциональной геномики. C. elegans использовался для анализа функций генов и обещал будущие открытия в области систематических генетических взаимодействий. [90]
Поглощение РНКи из окружающей среды намного хуже у других видов червей рода Caenorhabditis . Хотя инъекция РНК в полость тела животного вызывает молчание генов у большинства видов, только C. elegans и несколько других отдаленно родственных нематод могут поглощать РНК из бактерий, которые они едят, для получения РНКи. [91] Эта способность была связана с единственным геном, sid-2 , который, будучи вставленным в виде трансгена в другие виды, позволяет им поглощать РНК для РНКи, как это делает C. elegans . [92]
Деление клеток и клеточный цикл
[ редактировать ]Исследование мейоза значительно упростилось, поскольку каждое ядро зародышевой клетки находится в одном и том же заданном положении при движении вниз по гонаде, а значит, находится на одной и той же стадии мейоза. На ранней фазе мейоза ооциты становятся чрезвычайно устойчивыми к радиации, и эта устойчивость зависит от экспрессии генов rad51 и atm , которые играют ключевую роль в рекомбинационной репарации. [93] [94] Ген mre-11 также играет решающую роль в рекомбинационном восстановлении повреждений ДНК во время мейоза. [95] Исследование частоты ауткроссинга в природных популяциях показало, что самоопыление является преобладающим способом размножения C. elegans , но нечастые случаи ауткроссинга происходят с частотой около 1%. [96] Мейозы, которые приводят к самоопылению, вряд ли будут вносить значительный вклад в полезную генетическую изменчивость, но эти мейозы могут обеспечить адаптивное преимущество рекомбинационного восстановления повреждений ДНК, которые возникают, особенно в стрессовых условиях. [97]
Злоупотребление наркотиками и зависимость
[ редактировать ]Никотиновую зависимость также можно изучать с помощью C. elegans, поскольку она демонстрирует поведенческие реакции на никотин, аналогичные реакциям млекопитающих. Эти реакции включают острую реакцию, толерантность, абстинентность и сенсибилизацию. [98]
Биологические базы данных
[ редактировать ]Что касается большинства модельных организмов, ученые, работающие в этой области, создают специальную онлайн-базу данных, а WormBase — это база данных C. elegans . WormBase пытается сопоставить всю опубликованную информацию о C. elegans и других родственных нематодах. Информация о C. elegans включена в данные о других модельных организмах Альянса геномных ресурсов. [99] .
Старение
[ редактировать ]C. elegans был модельным организмом для исследования старения ; например, было показано, что ингибирование сигнального пути инсулиноподобного фактора роста увеличивает продолжительность жизни взрослых в три раза; [100] [101] в то время как кормление глюкозой способствует окислительному стрессу и сокращает продолжительность жизни взрослых вдвое. [81] Аналогично, индуцированная деградация рецептора инсулина/IGF-1 в позднем возрасте значительно увеличила продолжительность жизни червей. [102] Было продемонстрировано, что долгоживущие мутанты C. elegans устойчивы к окислительному стрессу и УФ-излучению . [103] Эти долгоживущие мутанты обладали более высокой способностью к репарации ДНК, дикого типа чем C. elegans . [103] Нокдаун гена эксцизионной репарации нуклеотидов Xpa-1 увеличил чувствительность к ультрафиолету и сократил продолжительность жизни долгоживущих мутантов. Эти данные показывают, что способность к репарации ДНК лежит в основе долголетия .
Способность восстанавливать повреждения ДНК путем эксцизионной репарации нуклеотидов снижается с возрастом. [104]
C. elegans , подвергшийся воздействию 5 мМ хлорида лития (LiCl), продемонстрировал увеличение продолжительности жизни. [105] При воздействии 10 мкМ LiCl наблюдалось снижение смертности, но не при 1 мкМ. [106]
C. elegans сыграл важную роль в идентификации функций генов, вовлеченных в болезнь Альцгеймера , таких как пресенилин . [107] Более того, обширные исследования C. elegans выявили, что РНК-связывающие белки являются важными факторами во время зародышевого и раннего эмбрионального развития. [108]
Теломеры , длина которых, как было показано, коррелирует с увеличением продолжительности жизни и задержкой начала старения у множества организмов, от C. elegans. [109] [110] людям, [111] демонстрируют интересное поведение у C. elegans. В то время как C. elegans поддерживает свои теломеры каноническим образом, подобно другим эукариотам, Drosophila melanogaster, напротив , примечательна тем, что использует ретротранспозоны для поддержания своих теломер. [112] при нокауте каталитической субъединицы теломеразы ( trt-1 ) C. elegans может приобретать способность альтернативного удлинения теломер (АЛТ). C. elegans был первым эукариотом, получившим функциональность АЛТ после отключения канонического теломеразного пути. [113] АЛТ также наблюдается примерно в 10-15% всех клинических случаев рака. [114] Таким образом, C. elegans является главным кандидатом для исследования АЛТ. [115] [116] [117] Баят и др. показали парадоксальное укорочение теломер во время trt-1 сверхэкспрессии , что привело к почти бесплодию , в то время как у червей даже наблюдалось небольшое увеличение продолжительности жизни, несмотря на укорочение теломер. [118]
Спать
[ редактировать ]C. elegans известен в исследованиях сна на животных как самый примитивный организм, демонстрирующий состояния, подобные сну. У C. elegans фаза летаргуса каждой наступает незадолго до линьки . [119] Также было продемонстрировано, что C. elegans засыпает после воздействия физического стресса, включая тепловой шок, ультрафиолетовое излучение и бактериальные токсины. [120]
Сенсорная биология
[ редактировать ]Хотя у червя нет глаз, было обнаружено, что он чувствителен к свету из-за третьего типа светочувствительного животного белка-фоторецептора , LITE-1 , который в 10–100 раз более эффективно поглощает свет, чем два других типа. фотопигменты ( опсины и криптохромы ), встречающиеся в животном мире. [121]
C. elegans замечательно переносит ускорение. он может выдержать 400 000 g По данным генетиков из Университета Сан-Паулу в Бразилии, . В ходе эксперимента 96% из них остались живы без каких-либо побочных эффектов после часа пребывания в ультрацентрифуге. [122]
Космические исследования
[ редактировать ]C. elegans попал в новости, когда было обнаружено, что экземпляры пережили космического корабля «Колумбия» катастрофу в феврале 2003 года. [123] Позже, в январе 2009 года, было объявлено, что живые образцы C. elegans из Ноттингемского университета проведут две недели на Международной космической станции в октябре того же года в рамках проекта космических исследований по изучению влияния невесомости на развитие мышц и физиологию. Исследование в первую очередь касалось генетической основы мышечной атрофии , которая связана с космическим полетом или прикованностью к постели, гериатрическим заболеванием или диабетом . [124] Потомки червей на борту «Колумбии» в 2003 году были запущены в космос на корабле «Индевор» для миссии STS-134 . [125] Дополнительные эксперименты по мышечной дистрофии во время космического полета проводились на борту МКС начиная с 2018 года. [126] Было показано, что гены, влияющие на прикрепление мышц, меньше экспрессируются в космосе. Однако еще предстоит выяснить, влияет ли это на мышечную силу.
Генетика
[ редактировать ]Геном
[ редактировать ] | |
| NCBI Идентификатор генома | 41 |
|---|---|
| Плоидность | диплоидный |
| Размер генома | 101,169 Мб (гаплоидный) |
| Количество хромосом | 5 пар аутосом (I, II, III, IV и V) + 1 или 2 половые хромосомы (X [127] ) |
| Год завершения | 1998 |
| Секвенированная органелла | митохондрия |
| Размер органеллы | 0,01 Мб |
C. elegans был первым многоклеточным организмом, весь геном которого секвенирован . Этот эпизод был опубликован в 1998 году. [128] хотя присутствовали некоторые небольшие пробелы; последний разрыв завершился к октябрю 2002 года. [ нужна ссылка ] В преддверии полного генома Консорциум секвенирования C. elegans / Проект генома C. elegans опубликовал несколько частичных сканирований, включая Wilson et al. 1994. [129] [130] [131]
Размер и содержание генов
[ редактировать ]Геном C. elegans имеет длину около 100 миллионов пар оснований и состоит из шести пар хромосом у гермафродитов или пяти пар аутосом с ХО-хромосомой у самцов C. elegans и митохондриального генома . Плотность его генов составляет примерно один ген на пять пар тысяч оснований . Интроны составляют 26%, а межгенные области — 47% генома. Многие гены расположены в кластерах, и сколько из них оперонов . неизвестно, [132] C. elegans и другие нематоды относятся к числу немногих известных в настоящее время эукариот, имеющих опероны; к ним относятся трипаносомы , плоские черви (особенно трематода Schistosoma mansoni ) и примитивная хордовая оболочница Oikopleura dioica . Вероятно, будет показано, что многие другие организмы имеют эти опероны. [133]
Геном содержит около 20 470 белок кодирующих генов, . [134] Около 35% генов C. elegans имеют человеческие гомологи . Примечательно, что неоднократно было показано, что человеческие гены заменяют своих гомологов C. elegans при введении в C. elegans . И наоборот, многие гены C. elegans могут функционировать аналогично генам млекопитающих. [48]
Число известных генов РНК в геноме значительно увеличилось благодаря открытию в 2006 году нового класса, названного генами 21U-РНК . [135] Сейчас считается, что геном содержит более 16 000 генов РНК, по сравнению с всего лишь 1300 в 2005 году. [136]
Научные кураторы продолжают оценивать набор известных генов; Продолжают добавляться новые модели генов, а неправильные модифицируются или удаляются.
Эталонная последовательность генома C. elegans продолжает меняться, поскольку новые данные выявляют ошибки в исходном секвенировании. Большинство изменений незначительны: добавляются или удаляются лишь несколько пар оснований ДНК. Например, в выпуске WormBase WS202 (апрель 2009 г.) к последовательности генома были добавлены две пары оснований. [137] Иногда вносятся более обширные изменения, как отмечено в выпуске WS197 от декабря 2008 г., в котором к последовательности добавлена область длиной более 4300 п.н. [138] [139]
C. elegans» Wilson et al. из проекта «Геном . 1994 год – основан ЦелВав. [131] и фактора А фон Виллебранда домен [129] и с Уилсоном и др. 1998 г. предоставило первые достоверные доказательства существования арилуглеводородного рецептора (AHR) гомолога за пределами позвоночных. [130] 2
Родственные геномы
[ редактировать ]В 2003 году также была определена последовательность генома родственной нематоды C. briggsae , что позволило исследователям изучить сравнительную геномику этих двух организмов. [140] Последовательности генома других нематод того же рода , например, C. remanei , [141] К. японская [142] и C. brenneri (названный в честь Бреннера) также изучались с использованием метода дробового секвенирования . [143] Эти последовательности теперь завершены. [144] [145]
Другие генетические исследования
[ редактировать ]
По состоянию на 2014 год C. elegans является самым базальным видом в группе «Elegans» (10 видов) супергруппы «Elegans» (17 видов) в филогенетических исследованиях. Он образует отдельную ветвь, отличную от любого другого вида группы. [146]
Транспозон Tc1 представляет собой ДНК-транспозон, активный у C. elegans .
Научное сообщество
[ редактировать ]В 2002 году Нобелевская премия по физиологии и медицине была присуждена Сидни Бреннеру , Х. Роберту Хорвицу и Джону Салстону за работу по генетике развития органов и запрограммированной гибели клеток у C. elegans . 2006 года Нобелевская премия по физиологии и медицине была присуждена Эндрю Файру и Крейгу К. Мелло за открытие РНК-интерференции у C. elegans . [147] В 2008 году Мартин Чалфи получил Нобелевскую премию по химии за работу над зеленым флуоресцентным белком ; некоторые исследования включали использование C. elegans .
Многие ученые, исследующие C. elegans, тесно связаны с Сидни Бреннером, с которым в 1970-х годах начались почти все исследования в этой области; они работали либо в качестве докторанта , либо в аспирантуре в лаборатории Бреннера или в лаборатории кого-то, кто ранее работал с Бреннером. Большинство из тех, кто работал в его лаборатории, позже основали свои собственные лаборатории по исследованию червей, тем самым создав довольно хорошо документированную «линию» ученых C. elegans , которая была WormBase на Международной встрече червей 2003 года. довольно подробно записана в базе данных [148]
См. также
[ редактировать ]- Испытания на животных на беспозвоночных
- Биолюминесценция
- Эйлин Саутгейт
- Интронератор
- OpenWorm
- Червячная Книга
Ссылки
[ редактировать ]- ^ Мопас, Э (1900). «Способы и формы размножения нематод». Архив экспериментальной и общей зоологии . 8 : 463–624.
- ^ Нигон V (1949). «Особенности размножения и половой детерминизм у некоторых свободноживущих нематод». Энн. наук. Нат. Зоол. Биол. Анимация . 11 :1–132.
- ^ Моерман Д.Г., Уотерстон Р.Х. (декабрь 1984 г.). «Спонтанные нестабильные мутации unc-22 IV у C. elegans var. Bergerac» . Генетика . 108 (4): 859–77. дои : 10.1093/генетика/108.4.859 . ПМК 1224270 . ПМИД 6096205 .
- ^ Бэбити Дж.М., Starr TV, Rose AM (июнь 1990 г.). «Транспозиция Tc1 и мутаторная активность у бристольского штамма Caenorhabditis elegans ». Молекулярная и общая генетика . 222 (1): 65–70. дои : 10.1007/bf00283024 . ПМИД 1978238 . S2CID 11275388 .
- ^ Харрис Л.Дж., Роуз А.М. (июль 1989 г.). «Структурный анализ элементов Tc1 у Caenorhabditis elegans var. Bristol (штамм N2)». Плазмида . 22 (1): 10–21. дои : 10.1016/0147-619x(89)90031-0 . ПМИД 2550981 .
- ^ «Ценорабдитис» . Словарь Merriam-Webster.com .
- ^ Вуд, ВБ (1988). Нематода Caenorhabditis elegans . Лабораторный пресс Колд-Спринг-Харбор . п. 1. ISBN 978-0-87969-433-3 .
- ^ Судхаус В., Кионтке К. (2009). «Филогения подрода Caenorhabditis Rhabditis ( Rhabditidae, Nematoda)» . Журнал зоологической систематики и эволюционных исследований . 34 (4): 217–233. дои : 10.1111/j.1439-0469.1996.tb00827.x .
- ^ καινός (caenos) = новый, недавний; ῥάβδος (рабдос) = жезл, жезл.
- ^ Феррис, Х. (30 ноября 2013 г.). « Каенорхабдитис элегантный » . Калифорнийский университет в Дэвисе . Архивировано из оригинала 9 декабря 2013 года . Проверено 19 ноября 2013 г.
- ^ Уоллес Р.Л., Риччи С., Мелоне Дж. (1996). «Кладистический анализ морфологии псевдоцеломата (ашельминта)». Биология беспозвоночных . 115 (2): 104–112. дои : 10.2307/3227041 . JSTOR 3227041 .
- ^ «Введение в определение пола» . www.wormbook.org . Проверено 15 марта 2017 г.
- ^ Перейти обратно: а б Бреннер С. (май 1974 г.). «Генетика Caenorhabditis elegans » . Генетика . 77 (1): 71–94. дои : 10.1093/генетика/77.1.71 . ПМЦ 1213120 . ПМИД 4366476 .
- ^ Перейти обратно: а б Кук, С.Дж.; Джаррелл, штат Техас; Бриттин, Калифорния; Ван, Ю; Блониарз, А.Е.; Яковлев, М.А.; Нгуен, KCQ; Тан, лейтенант-H; Байер, Э.А.; Дюрр, Дж.С.; Бюлов, HE; Хоберт, О; Холл, ДХ; Эммонс, Юго-Запад (3 декабря 2019 г.). «Цельные коннектомы животных обоих полов Caenorhabditis elegans » . Природа . 571 (7763). Национальная медицинская библиотека США, Национальные институты здравоохранения: 63–71. Бибкод : 2019Natur.571...63C . дои : 10.1038/s41586-019-1352-7 . ПМК 6889226 . ПМИД 31270481 .
- ^ Уайт Дж.Г., Саутгейт Э., Томсон Дж.Н., Бреннер С. (ноябрь 1986 г.). «Строение нервной системы нематоды Caenorhabditis elegans » . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 314 (1165): 1–340. Бибкод : 1986RSPTB.314....1W . дои : 10.1098/rstb.1986.0056 . ПМИД 22462104 .
- ^ Уайт JG (июнь 2013 г.). «Проникновение в разум червя – личный взгляд» . Червячная книга : 1–10. дои : 10.1895/wormbook.1.158.1 . ПМЦ 4781474 . ПМИД 23801597 .
- ^ Джабр Ф (2 октября 2012 г.). «Дебаты о коннектоме: стоит ли картировать разум червя?» . Научный американец . Проверено 18 января 2014 г.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2007). Молекулярная биология клетки (5-е изд.). Гирляндная наука . п. 1321. ИСБН 978-0-8153-4105-5 .
- ^ Лемье Г.А., Ашрафи К. (август 2016 г.). «Исследование связи между метаболизмом, долголетием и поведением Caenorhabditis elegans» . Тенденции в эндокринологии и обмене веществ . 27 (8): 586–596. дои : 10.1016/j.tem.2016.05.004 . ПМЦ 4958586 . ПМИД 27289335 .
- ^ « Глотка C. elegans : модель органогенеза» . www.wormbook.org . Проверено 15 марта 2017 г.
- ^ Соарес Ф.А., Фагундес Д.А., Авила Д.С. (2017). «Нейродегенерация, вызванная металлами у Caenorhabditis elegans». Нейротоксичность металлов . Достижения нейробиологии. Том. 18. стр. 55–383. дои : 10.1007/978-3-319-60189-2_18 . ISBN 978-3-319-60188-5 . ПМИД 28889277 .
- ^ Нонет, М. (2004) О нематоде Caenorhabditis elegans .
- ^ Хоберт, Оливер (2005). «Спецификация нервной системы» . Червячная книга : 1–19. дои : 10.1895/wormbook.1.12.1 . ПМЦ 4781215 . ПМИД 18050401 .
- ^ Коберн С., Джемс Д. (2013). «Таинственный случай кишечных гранул C. elegans : смертельная флуоресценция, антраниловая кислота и путь кинуренина» . Границы генетики . 4 : 151. дои : 10.3389/fgene.2013.00151 . ПМЦ 3735983 . ПМИД 23967012 .
- ^ Наяк С., Гори Дж., Шедл Т. (январь 2005 г.). «Туман-2 и эволюция самоплодного гермафродитизма у Caenorhabditis » . ПЛОС Биология . 3 (1): e6. doi : 10.1371/journal.pbio.0030006 . ПМК 539060 . ПМИД 15630478 .
- ^ Лоер CM, Кеньон CJ (декабрь 1993 г.). «Мутанты с дефицитом серотонина и брачное поведение самцов нематоды Caenorhabditis elegans » . Журнал неврологии . 13 (12): 5407–17. doi : 10.1523/jneurosci.13-12-05407.1993 . ПМК 6576401 . ПМИД 8254383 .
- ^ Ма Х, Чжао Ю, Сунь В, Симабукуро К, Мяо Л (октябрь 2012 г.). «Трансформация: как сперматозоиды нематод активируются и ползают?» . Белок и клетка . 3 (10): 755–61. дои : 10.1007/s13238-012-2936-2 . ПМЦ 4875351 . ПМИД 22903434 .
- ^ Перейти обратно: а б с Старостина Н.Г., Лим Дж.М., Шварцштейн М., Уэллс Л., Спенс А.М., Кипреос Э.Т. (июль 2007 г.). «Убиквитинлигаза CUL-2, содержащая три белка FEM, разрушает TRA-1, регулируя определение пола C. elegans» . Развивающая клетка . 13 (1): 127–39. дои : 10.1016/j.devcel.2007.05.008 . ПМК 2064902 . ПМИД 17609115 .
- ^ Перейти обратно: а б с д и ж г час Хааг ES (декабрь 2005 г.). «Эволюция определения пола нематод: C. elegans как ориентир для сравнительной биологии» . WormBook: Интернет-обзор биологии C. Elegans : 1–14. дои : 10.1895/wormbook.1.120.1 . ПМЦ 4781019 . ПМИД 18050417 .
- ^ Перейти обратно: а б «Справочник – Мужское знакомство» . www.wormatlas.org . Проверено 30 марта 2021 г.
- ^ Фрезал Л., Феликс М.А. (март 2015 г.). «C. elegans вне чашки Петри» . электронная жизнь . 4 : e05849. doi : 10.7554/eLife.05849 . ПМЦ 4373675 . ПМИД 25822066 .
- ^ Перейти обратно: а б с д Конрад, Барбара; Хорвиц, Х. Роберт (6 августа 1999 г.). «Белок определения пола TRA-1A C. elegans регулирует гибель половых диморфных клеток путем репрессии гена-активатора гибели клеток egl-1» . Клетка . 98 (3): 317–327. дои : 10.1016/S0092-8674(00)81961-3 . ISSN 0092-8674 . ПМИД 10458607 . S2CID 14951719 .
- ^ Миллер Л.М., Пленефиш Дж.Д., Кассон Л.П., Мейер Б.Дж. (октябрь 1988 г.). «xol-1: ген, который контролирует мужские способы определения пола и компенсацию дозы Х-хромосомы у C. elegans». Клетка . 55 (1): 167–83. дои : 10.1016/0092-8674(88)90019-0 . ПМИД 3167975 . S2CID 5005906 .
- ^ «микс-1 (ген) — WormBase: Информационный ресурс по нематодам» . wormbase.org . Проверено 23 апреля 2021 г.
- ^ Перейти обратно: а б Каттер А.Д., Уорд С. (январь 2005 г.). «Половая и временная динамика молекулярной эволюции в развитии C. elegans» . Молекулярная биология и эволюция . 22 (1): 178–88. дои : 10.1093/molbev/msh267 . ПМИД 15371532 .
- ^ Гольдштейн Б., Хирд С.Н. (май 1996 г.). «Характеристика переднезадней оси Caenorhabditis elegans» . Разработка . 122 (5): 1467–74. дои : 10.1242/dev.122.5.1467 . ПМИД 8625834 .
- ^ Гилберт С.Ф. (2016). Биология развития (11-е изд.). Синауэр. п. 268. ИСБН 9781605354705 .
- ^ Го С., Кемфуес К.Дж. (май 1995 г.). «par-1, ген, необходимый для установления полярности у эмбрионов C. elegans , кодирует предполагаемую киназу Ser/Thr, которая распределена асимметрично» . Клетка . 81 (4): 611–20. дои : 10.1016/0092-8674(95)90082-9 . ПМИД 7758115 .
- ^ Перейти обратно: а б Генчи П., Роуз Л.С. (октябрь 2005 г.). «Асимметричное деление клеток и формирование оси у эмбриона» . Червячная книга : 1–20. дои : 10.1895/wormbook.1.30.1 . ПМЦ 4780927 . ПМИД 18050411 .
- ^ Кимбл Дж., Криттенден С.Л. Пролиферация зародышевой линии и ее контроль. 15 августа 2005 г. В: WormBook: Интернет-обзор биологии C. elegans [Интернет]. Пасадена (Калифорния): WormBook; 2005-. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK19769/.
- ^ Перейти обратно: а б «WBbt:0006773 (анатомический термин)». WormBase (изд. WS242). 14 мая 2014 г. WBbt:0006773.
- ^ Гилберт С.Ф. (2016). Биология развития (11-е изд.). Синауэр. п. 272. ИСБН 9781605354705 .
- ^ Торп С.Дж., Шлезингер А., Картер Дж.К., Бауэрман Б. (август 1997 г.). «Передача сигналов Wnt поляризует ранний бластомер C. elegans , чтобы отличить энтодерму от мезодермы» . Клетка . 90 (4): 695–705. дои : 10.1016/s0092-8674(00)80530-9 . ПМИД 9288749 .
- ^ Пол С., Бао З. (сентябрь 2010 г.). «Киральные силы организуют формирование левого и правого паттерна у C. elegans путем разъединения средней линии и переднезадней оси» . Развивающая клетка . 19 (3): 402–12. дои : 10.1016/j.devcel.2010.08.014 . ПМЦ 2952354 . ПМИД 20833362 . Вилларес Х.К., Карлини Э.А. (1988). «[Количественная оценка сальной экскреции у добровольцев: влияние хронологического возраста, пола и расы]». Мед Кутан Иберо Лат Ам . 16 (6): 439–44. ПМИД 3073266 . Гилберт С.Ф. (2016). Биология развития (11-е изд.). Синауэр. п. 269. ИСБН 9781605354705 .
- ^ Скиба Ф, Ширенберг Э (июнь 1992 г.). «Клеточные линии, сроки развития и формирование пространственной структуры у эмбрионов свободноживущих почвенных нематод» . Биология развития . 151 (2): 597–610. дои : 10.1016/0012-1606(92)90197-о . ПМИД 1601187 .
- ^ Гилберт С.Ф. (2016). Биология развития (11-е изд.). Синауэр. п. 273. ИСБН 9781605354705 .
- ^ Людвиг, Андреас Х.; Шредер, Фрэнк К. (18 января 2013 г.). «Передача сигналов аскарозида у C. elegans» . Червячная книга : 1–22. дои : 10.1895/wormbook.1.155.1 . ISSN 1551-8507 . ПМЦ 3758900 . ПМИД 23355522 .
- ^ Перейти обратно: а б «Знакомство с C. Elegans » . К. Элеганс как модельный организм . Университет Рутгерса. Архивировано из оригинала 18 августа 2002 г. Проверено 15 августа 2014 г.
- ^ «Справочник – Введение» .
- ^ «Дауэр» . www.wormbook.org . Проверено 27 сентября 2018 г.
- ^ Резник Т.Д., Маккалок К.А., Ругви А.Е. (май 2010 г.). «МиРНК дают червям время их жизни: малые РНК и временной контроль у Caenorhabditis elegans » . Динамика развития . 239 (5): 1477–89. дои : 10.1002/dvdy.22260 . ПМЦ 4698981 . ПМИД 20232378 .
- ^ Ругви А.Э., Мосс Э.Г. (2013). «Переходы в развитии на личиночных стадиях C. Elegans». Переходы в развитии на C. elegans личиночных стадиях . Сроки развития. Том. 105. Академическая пресса. стр. 153–80. дои : 10.1016/B978-0-12-396968-2.00006-3 . ISBN 9780123969682 . ПМИД 23962842 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Ли Р.К., Фейнбаум Р.Л., Амброс В. (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК с антисмысловой комплементарностью к lin-14» . Клетка . 75 (5): 843–54. дои : 10.1016/0092-8674(93)90529-у . ПМИД 8252621 .
- ^ Банерджи Д., Квок А., Лин С.Ю., Слэк Ф.Дж. (февраль 2005 г.). «Время развития C. elegans регулируется kin-20 и tim-1, гомологами основных генов циркадных часов» . Развивающая клетка . 8 (2): 287–95. дои : 10.1016/j.devcel.2004.12.006 . ПМИД 15691769 .
- ^ Салстон, Дж. Э.; Хорвиц, HR (март 1977 г.). «Постэмбриональные клеточные линии нематоды Caenorhabditis elegans» . Биология развития . 56 (1): 110–156. дои : 10.1016/0012-1606(77)90158-0 . ПМИД 838129 .
- ^ Салстон, Дж. Э.; Ширенберг, Э.; Уайт, Дж. Г.; Томсон, Дж. Н. (ноябрь 1983 г.). «Эмбриональная клеточная линия нематоды Caenorhabditis elegans» . Биология развития . 100 (1): 64–119. дои : 10.1016/0012-1606(83)90201-4 . ПМИД 6684600 .
- ^ Перейти обратно: а б Саммут, Мишель; Кук, Стивен Дж.; Нгуен, Кен CQ; Фелтон, Терри; Холл, Дэвид Х.; Эммонс, Скотт В.; Пул, Ричард Дж.; Барриос, Арантса (октябрь 2015 г.). «Нейроны, происходящие из глии, необходимы для обучения C. elegans в зависимости от пола» . Природа . 526 (7573): 385–390. Бибкод : 2015Natur.526..385S . дои : 10.1038/nature15700 . ISSN 0028-0836 . ПМК 4650210 . ПМИД 26469050 .
- ^ Оно, Хаяо; Бао, Чжижун (14 ноября 2020 г.). «Маленькие РНК соединяют программы эмбрионального развития с кишечными микробами» . биоRxiv . дои : 10.1101/2020.11.13.381830 . S2CID 227060212 .
- ^ Руперт Э.Э., Фокс Р.С., Барнс Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Cengage Обучение. п. 753. ИСБН 978-81-315-0104-7 .
- ^ Ху, Минци; Ма, Йепин; Чуа, Сун Линь (16 января 2024 г.). «Бактериоядные нематоды расшифровывают микробные железные сидерофоры как сигнал добычи во взаимодействиях хищник-жертва» . Труды Национальной академии наук . 121 (3): e2314077121. Бибкод : 2024PNAS..12114077H . дои : 10.1073/pnas.2314077121 . ISSN 0027-8424 . ПМК 10801909 . ПМИД 38190542 .
- ^ Феликс М.А., Брэндл С. (ноябрь 2010 г.). «Естественная история Caenorhabditis elegans » . Современная биология . 20 (22): R965–9. Бибкод : 2010CBio...20.R965F . дои : 10.1016/j.cub.2010.09.050 . ПМИД 21093785 .
- ^ Чан, пастырь Юэнь; Лю, Сильвия Ян; Ву, Ронгбен; Вэй, Вэй; Фанг, Джеймс Кар-Хей; Чуа, Сун Линь (2 июня 2023 г.). «Одновременное распространение нанопластиков и устойчивости к антибиотикам нематодными курьерами» . Экологические науки и технологии . 57 (23): 8719–8727. Бибкод : 2023EnST...57.8719C . дои : 10.1021/acs.est.2c07129 . ISSN 0013-936X . ПМИД 37267481 . S2CID 259047038 .
- ^ Милонакис Э., Осубель Ф.М., Перфект Дж.Р., Хейтман Дж., Колдервуд С.Б. (ноябрь 2002 г.). «Уничтожение Caenorhabditis elegans Cryptococcus neoformans как модель дрожжевого патогенеза» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (24): 15675–80. Бибкод : 2002PNAS...9915675M . дои : 10.1073/pnas.232568599 . ПМЦ 137775 . ПМИД 12438649 .
- ^ Sifri CD, Begun J, Ausubel FM, Calderwood SB (апрель 2003 г.). «Caenorhabditis elegans как модельный хозяин патогенеза Staphylococcus aureus» . Инфекция и иммунитет . 71 (4): 2208–17. дои : 10.1128/IAI.71.4.2208-2217.2003 . ПМК 152095 . ПМИД 12654843 .
- ^ Тан М.В., Махаджан-Миклош С., Аусубель FM (январь 1999 г.). «Уничтожение Caenorhabditis elegans Pseudomonas aeruginosa, используемое для моделирования бактериального патогенеза млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (2): 715–20. Бибкод : 1999PNAS...96..715T . дои : 10.1073/pnas.96.2.715 . ПМЦ 15202 . ПМИД 9892699 .
- ^ Гарсин Д.А., Вильянуэва Дж.М., Бегун Дж., Ким Д.Х., Сифри С.Д., Колдервуд С.Б. и др. (июнь 2003 г.). «Долгоживущие мутанты C. elegans daf-2 устойчивы к бактериальным патогенам». Наука . 300 (5627): 1921. doi : 10.1126/science.1080147 . ПМИД 12817143 . S2CID 37703980 .
- ^ Чан, пастырь Юэнь; Лю, Сильвия Ян; Сенг, Цзыцзин; Чуа, Сун Линь (январь 2021 г.). «Матрица биопленки нарушает подвижность и хищническое поведение нематод» . Журнал ISME . 15 (1): 260–269. Бибкод : 2021ISMEJ..15..260C . дои : 10.1038/s41396-020-00779-9 . ISSN 1751-7370 . ПМЦ 7852553 . ПМИД 32958848 .
- ^ Ли, Шаоян; Лю, Сильвия Ян; Чан, пастырь Юэнь; Чуа, Сун Линь (май 2022 г.). «Матрица биопленки скрывает бактериальный кворум, чувствительный к хемоаттрактантам, от обнаружения хищниками» . Журнал ISME . 16 (5): 1388–1396. Бибкод : 2022ISMEJ..16.1388L . дои : 10.1038/s41396-022-01190-2 . ISSN 1751-7370 . ПМЦ 9038794 . ПМИД 35034106 .
- ^ Кионтке К., Судхаус В. (январь 2006 г.). «Экология видов Caenorhabditis» . Червячная книга : 1–14. дои : 10.1895/wormbook.1.37.1 . ПМК 4780885 . ПМИД 18050464 .
- ^ Гал Т.З., Глейзер И., Колтай Х. (ноябрь 2004 г.). «Член семьи LEA группы 3 участвует в выживании C. elegans во время воздействия стресса» . Письма ФЭБС . 577 (1–2): 21–6. Бибкод : 2004FEBSL.577...21G . дои : 10.1016/j.febslet.2004.09.049 . ПМИД 15527756 . S2CID 21960486 .
- ^ Элейн Р. Ингхэм Учебник по биологии почвы , Министерство сельского хозяйства США
- ^ Боуэн, Нью-Джерси (1999). «Геномный анализ Caenorhabditis elegans выявил древние семейства ретровирусоподобных элементов» . Геномные исследования . 9 (10): 924–935. дои : 10.1101/гр.9.10.924 . ПМИД 10523521 .
- ^ Куомо Калифорния, Дежарден Калифорния, Баковски М.А., Голдберг Дж., Ма А.Т., Бекнель Дж.Дж., Дидье Э.С., Фан Л., Хейман Д.И., Левин Дж.З., Янг С., Зенг К., Тремель Э.Р. (декабрь 2012 г.). «Анализ генома микроспоридий выявляет эволюционные стратегии обязательного внутриклеточного роста» . Геномные исследования . 22 (12): 2478–88. дои : 10.1101/гр.142802.112 . ПМЦ 3514677 . ПМИД 22813931 .
- ^ Ню XM, Чжан К.К. (2011). « Arthrobotrys oligospora : модельный организм для понимания взаимодействия между грибами и нематодами» . Микология . 2 (2): 59–78. дои : 10.1080/21501203.2011.562559 .
- ^ Клэр Дж.Дж., Тейт С.Н., Ноббс М., Романос М.А. (ноябрь 2000 г.). «Натриевые каналы, управляемые напряжением, как терапевтические мишени». Открытие наркотиков сегодня . 5 (11): 506–520. дои : 10.1016/S1359-6446(00)01570-1 . ПМИД 11084387 .
- ^ Косинский Р.А., Заремба М (2007). «Динамика модели нейронной сети Caenorhabditis Elegans». Акта Физика Полоника Б. 38 (6): 2201. Бибкод : 2007AcPPB..38.2201K .
- ^ Бруйетт, Моник (21 апреля 2022 г.). «Картирование мозга, чтобы понять разум» . Познаваемый журнал | Ежегодные обзоры . doi : 10.1146/knowable-042122-1 .
- ^ Уоттс-ди-джей, Строгац С.Х. (июнь 1998 г.). «Коллективная динамика сетей «маленького мира». Природа . 393 (6684): 440–2. Бибкод : 1998Natur.393..440W . дои : 10.1038/30918 . ПМИД 9623998 . S2CID 4429113 .
- ^ Шафер В.Р. (сентябрь 2005 г.). «Расшифровка нейронных и молекулярных механизмов поведения C. elegans» . Современная биология . 15 (17): R723–9. Бибкод : 2005CBio...15.R723S . дои : 10.1016/j.cub.2005.08.020 . ПМИД 16139205 .
- ^ Кук С.Дж., Джаррелл Т.А., Бриттин К.А., Ван Ю., Блониарц А.Е., Яковлев М.А. и др. (июль 2019 г.). «Цельные коннектомы животных обоих полов Caenorhabditis elegans» . Природа . 571 (7763): 63–71. Бибкод : 2019Natur.571...63C . дои : 10.1038/s41586-019-1352-7 . ПМК 6889226 . ПМИД 31270481 .
- ^ Перейти обратно: а б Алькантар-Фернандес Х, Наварро Р.Э., Саласар-Мартинес А.М., Перес-Андраде М.Э., Миранда-Риос Х. (2018). «Caenorhabditis elegans реагирует на диету с высоким содержанием глюкозы через сеть стресс-чувствительных факторов транскрипции» . ПЛОС ОДИН . 13 (7): e0199888. Бибкод : 2018PLoSO..1399888A . дои : 10.1371/journal.pone.0199888 . ПМК 6039004 . ПМИД 29990370 .
- ^ Эйвери, Л. «Сидни Бреннер» . Юго-Западный медицинский центр . Архивировано из оригинала 15 августа 2011 года . Альтернативный вариант. URL-адрес, заархивированный 8 декабря 2013 г. на Wayback Machine.
- ^ Бреннер, С. (1974). «Генетика Caenorhabditis elegans» . Генетика . 77 (1): 71–94. дои : 10.1093/генетика/77.1.71 . ПМЦ 1213120 . ПМИД 4366476 .
- ^ "Поведение" . www.wormbook.org . Проверено 26 сентября 2018 г.
- ^ Салстон Дж. Э., Хорвиц Х. Р. (март 1977 г.). «Постэмбриональные клеточные линии нематоды Caenorhabditis elegans». Биология развития . 56 (1): 110–56. дои : 10.1016/0012-1606(77)90158-0 . ПМИД 838129 .
- ^ Кимбл Дж., Хирш Д. (июнь 1979 г.). «Постэмбриональные клеточные линии гермафродитных и мужских гонад Caenorhabditis elegans». Биология развития . 70 (2): 396–417. дои : 10.1016/0012-1606(79)90035-6 . ПМИД 478167 .
- ^ Педен Э., Киллиан DJ, Сюэ Д. (август 2008 г.). «Спецификация гибели клеток C. elegans» . Клеточный цикл . 7 (16): 2479–84. дои : 10.4161/cc.7.16.6479 . ПМЦ 2651394 . ПМИД 18719375 .
- ^ NIDDK, Национальный институт диабета, заболеваний органов пищеварения и почек (5 марта 2015 г.). «Инъекция гонад C. elegans» . Ютуб . Архивировано из оригинала 17 ноября 2021 г. Проверено 21 марта 2020 г.
- ^ Камат Р.С., Фрейзер А.Г., Донг Й., Пулин Г., Дурбин Р., Готта М., Канапин А., Ле Бот Н., Морено С., Сорманн М., Уэлчман Д.П., Зипперлен П., Арингер Дж. (январь 2003 г.). «Систематический функциональный анализ генома Caenorhabditis elegans с использованием РНКи». Природа . 421 (6920): 231–7. Бибкод : 2003Natur.421..231K . дои : 10.1038/nature01278 . hdl : 10261/63159 . ПМИД 12529635 . S2CID 15745225 .
- ^ Фортунато А., Fraser AG (2005). «Раскройте генетические взаимодействия у Caenorhabditis elegans с помощью РНК-интерференции». Отчеты по биологическим наукам . 25 (5–6): 299–307. дои : 10.1007/s10540-005-2892-7 . ПМИД 16307378 . S2CID 6983519 .
- ^ Феликс М.А. (ноябрь 2008 г.). «РНК-интерференция в нематодах и шанс, который благоприятствовал Сидни Бреннеру» . Журнал биологии . 7 (9): 34. дои : 10.1186/jbiol97 . ПМЦ 2776389 . ПМИД 19014674 .
- ^ Уинстон В.М., Сазерлин М., Райт А.Дж., Фейнберг Э.Х., Хантер С.П. (июнь 2007 г.). «Caenorhabditis elegans SID-2 необходим для вмешательства в РНК окружающей среды» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (25): 10565–70. Бибкод : 2007PNAS..10410565W . дои : 10.1073/pnas.0611282104 . ЧВК 1965553 . ПМИД 17563372 .
- ^ Таканами Т., Мори А., Такахаши Х., Хигаситани А. (ноябрь 2000 г.). «Гиперрезистентность мейотических клеток к радиации из-за сильной экспрессии одного recA-подобного гена у Caenorhabditis elegans» . Исследования нуклеиновых кислот . 28 (21): 4232–6. дои : 10.1093/нар/28.21.4232 . ПМЦ 113154 . ПМИД 11058122 .
- ^ Таканами Т., Чжан Ю., Аоки Х., Абэ Т., Ёсида С., Такахаши Х., Хориучи С., Хигаситани А. (сентябрь 2003 г.). «Эффективное восстановление повреждений ДНК, вызванных частицами тяжелых ионов в ядрах профазы I мейоза Caenorhabditis elegans» . Журнал радиационных исследований . 44 (3): 271–6. Бибкод : 2003JRadR..44..271T . дои : 10.1269/млр.44.271 . ПМИД 14646232 .
- ^ Чин Г.М., Вильнёв А.М. (март 2001 г.). «C. elegans mre-11 необходим для мейотической рекомбинации и восстановления ДНК, но не является обязательным для контрольной точки мейотического повреждения ДНК G(2)» . Гены и развитие . 15 (5): 522–34. дои : 10.1101/gad.864101 . ПМК 312651 . ПМИД 11238374 .
- ^ Барьер А., Феликс М.А. (июль 2005 г.). «Высокое местное генетическое разнообразие и низкий уровень ауткроссинга в природных популяциях Caenorhabditis elegans». Современная биология . 15 (13): 1176–84. arXiv : q-bio/0508003 . Бибкод : 2005CBio...15.1176B . дои : 10.1016/j.cub.2005.06.022 . ПМИД 16005289 . S2CID 2229622 .
- ^ Бернштейн Х., Бернштейн С. (июль 2010 г.) «Эволюционное происхождение рекомбинации во время мейоза», BioScience 60 (7), 498-505. https://doi.org/10.1525/bio.2010.60.7.5
- ^ Фэн З., Ли В., Уорд А., Пигготт Б.Дж., Ларкспер Э.Р., Штернберг П.В., Сюй XZ (ноябрь 2006 г.). «Модель никотинзависимого поведения C. elegans: регуляция каналами семейства TRP» . Клетка . 127 (3): 621–33. дои : 10.1016/j.cell.2006.09.035 . ПМК 2859215 . ПМИД 17081982 .
- ^ «Форум сообщества Альянса геномных ресурсов» . Форум сообщества Альянса геномных ресурсов . Проверено 1 августа 2024 г.
- ^ Волков К.А., Кимура К.Д., Ли М.С., Рувкун Г. (октябрь 2000 г.). «Регуляция продолжительности жизни C. elegans посредством инсулиноподобной передачи сигналов в нервной системе». Наука . 290 (5489): 147–50. Бибкод : 2000Sci...290..147W . дои : 10.1126/science.290.5489.147 . ПМИД 11021802 .
- ^ Эвальд С.И., Лэндис Дж.Н., Портер Абате Дж., Мерфи Коннектикут, Блэквелл Т.К. (март 2015 г.). «Дауэр-независимая передача сигналов инсулина/IGF-1 способствует ремоделированию коллагена в долголетии» . Природа . 519 (7541): 97–101. Бибкод : 2015Natur.519...97E . дои : 10.1038/nature14021 . ПМЦ 4352135 . ПМИД 25517099 .
- ^ Венц, Ричард; Пекеч, Тина; Катич, «Искра»; Циоск, Рафаль; Эвальд, Коллен Ив (10 сентября 2021 г.). Лейзер, Скотт Ф; Каберлейн, Мэтт; Альседо, Джой (ред.). «Направленная деградация инсулина DAF-2/рецептора IGF-1 в конце жизни способствует долголетию без патологий, связанных с ростом» . электронная жизнь . 10 : е71335. дои : 10.7554/eLife.71335 . ISSN 2050-084X . ПМК 8492056 . ПМИД 34505574 .
- ^ Перейти обратно: а б Хён, Мунджон; Ли, Джихён; Ли, Кёнджин; Мэй, Альфред; Бор, Вильгельм А.; Ан, Бёнчан (2008). «Долголетие и устойчивость к стрессу коррелируют со способностью восстановления ДНК у Caenorhabditis elegans» . Исследования нуклеиновых кислот . 36 (4): 1380–1389. дои : 10.1093/нар/gkm1161 . ПМК 2275101 . ПМИД 18203746 .
- ^ Мейер, Джоэл Н.; Бойд, Винди А.; Аззам, Грегори А.; Хауген, Астрид К.; Фридман, Джонатан Х.; Ван Хаутен, Беннетт (2007). «Снижение способности эксцизионной репарации нуклеотидов у стареющего Caenorhabditis elegans» . Геномная биология . 8 (5): 70 рандов. дои : 10.1186/gb-2007-8-5-r70 . ЧВК 1929140 . ПМИД 17472752 .
- ^ Макколл Г., Киллили Д.В., Хаббард А.Е., Вантипалли М.С., Мелов С., Литгоу Г.Дж. (январь 2008 г.). «Фармакогенетический анализ замедленного старения, вызванного литием, у Caenorhabditis elegans» . Журнал биологической химии . 283 (1): 350–7. дои : 10.1074/jbc.M705028200 . ПМЦ 2739662 . ПМИД 17959600 .
- ^ Зарсе К., Терао Т., Тиан Дж., Ивата Н., Исии Н., Ристоу М. (август 2011 г.). «Низкие дозы лития способствуют долголетию у людей и многоклеточных животных» . Европейский журнал питания . 50 (5): 387–9. дои : 10.1007/s00394-011-0171-x . ПМК 3151375 . ПМИД 21301855 .
- ^ Эвальд С.И., Ли С. (март 2010 г.). «Понимание молекулярной основы болезни Альцгеймера с использованием модельной системы Caenorhabditis elegans» . Структура и функции мозга . 214 (2–3): 263–83. дои : 10.1007/s00429-009-0235-3 . ПМК 3902020 . ПМИД 20012092 .
- ^ Ханадзава М., Йонетани М., Сугимото А. (март 2011 г.). «Белки PGL самостоятельно связываются и связываются с RNP, опосредуя сборку зародышевых гранул у C. elegans» . Журнал клеточной биологии . 192 (6): 929–37. дои : 10.1083/jcb.201010106 . ПМК 3063142 . ПМИД 21402787 .
- ^ Куттс, Фиона; Палмос, Алиш Б.; Дуарте, Родриго Р.Р.; де Йонг, Симона; Льюис, Кэтрин М.; Дима, Данай; Пауэлл, Тимоти Р. (март 2019 г.). «Полигенная природа длины теломер и антивозрастные свойства лития» . Нейропсихофармакология . 44 (4): 757–765. дои : 10.1038/s41386-018-0289-0 . ISSN 1740-634X . ПМК 6372618 . ПМИД 30559463 .
- ^ Райс, Марсела; Маруяма, Хьюго; Диллин, Эндрю; Карлседер, Ян (сентябрь 2005 г.). «Развязка продолжительности жизни и длины теломер у C. elegans» . ПЛОС Генетика . 1 (3): е30. дои : 10.1371/journal.pgen.0010030 . ISSN 1553-7404 . ПМК 1200426 . ПМИД 16151516 .
- ^ Люлькевич, М.; Байсерт, Дж.; Копчинский, П.; Барчак, В.; Рубис, Б. (сентябрь 2020 г.). «Длина теломер: какое значение имеет длина» . Отчеты по молекулярной биологии . 47 (9): 7181–7188. дои : 10.1007/s11033-020-05551-y . ISSN 1573-4978 . ПМЦ 7561533 . ПМИД 32876842 .
- ^ Пардью, Мэри-Лу; ДеБарише, разыгрывающий (20 декабря 2011 г.). «Ретротранспозоны, поддерживающие концы хромосом» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (51): 20317–20324. дои : 10.1073/pnas.1100278108 . ISSN 1091-6490 . ПМК 3251079 . ПМИД 21821789 .
- ^ Мейер, Беттина; Клеян, Юваль; Лю, Ян; Лоуден, Миа; Гартнер, Антон; Ходжкин, Джонатан; Ахмед, Шон (февраль 2006 г.). «trt-1 представляет собой каталитическую субъединицу теломеразы Caenorhabditis elegans» . ПЛОС Генетика . 2 (2): е18. дои : 10.1371/journal.pgen.0020018 . ISSN 1553-7404 . ПМЦ 1361356 . ПМИД 16477310 .
- ^ Чезаре, Энтони Дж.; Реддел, Роджер Р. (май 2010 г.). «Альтернативное удлинение теломер: модели, механизмы и последствия» . Обзоры природы. Генетика . 11 (5): 319–330. дои : 10.1038/nrg2763 . ISSN 1471-0064 . ПМИД 20351727 . S2CID 19224032 .
- ^ Иджомоне, Омамуёвви М.; Миа, Махфузур Р.; Перес, Танара В.; Нвоха, Поликарп У.; Ашнер, Майкл (декабрь 2016 г.). «Нулевые аллельные мутанты trt-1, каталитической субъединицы теломеразы Caenorhabditis elegans, менее чувствительны к Mn-индуцированной токсичности и DAergic дегенерации» . Нейротоксикология . 57 : 54–60. Бибкод : 2016NeuTx..57...54I . дои : 10.1016/j.neuro.2016.08.016 . ISSN 1872-9711 . ПМИД 27593554 .
- ^ Штессель, Людмила; Лоуден, Миа Рошель; Ченг, Чен; Саймон, Мэтт; Ван, Кайл; Ахмед, Шон (февраль 2013 г.). «Caenorhabditis elegans POT-1 и POT-2 подавляют пути поддержания теломер» . G3: Гены, геномы, генетика . 3 (2): 305–313. дои : 10.1534/g3.112.004440 . ISSN 2160-1836 . ПМЦ 3564990 . ПМИД 23390606 .
- ^ Квон, Ми-Сун; Мин, Джэвон; Чон, Хи-Ён; Хван, Кванву; Ким, Чуна; Ли, Чуно; Йонг, Дже-Гун; Пак, Вунг-Янг; Ли, Хёнсук (октябрь 2016 г.). «Парадоксальная задержка старения при истощении BRCA2 у червей с дефицитом теломеразы» . Открытая биография FEBS . 6 (10): 1016–1024. дои : 10.1002/2211-5463.12109 . ISSN 2211-5463 . ПМК 5055038 . ПМИД 27761361 .
- ^ Баят, Мелих; Тэнни, Робин Э.; Ван, Е; Херден, Карла; Дэниел, Йенс; Андерсен, Эрик С.; Либау, Ева; Вашк, Дэниел Э.Дж. (30 марта 2020 г.). «Эффекты сверхэкспрессии теломеразы в модельном организме Caenorhabditis elegans» . Джин . 732 : 144367. doi : 10.1016/j.gene.2020.144367 . ISSN 1879-0038 . ПМИД 31954861 . S2CID 210829489 .
- ^ Иванир С., Трамм Н., Надь С., Райт С., Иш Д., Бирон Д. (март 2013 г.). «Микроархитектура поведения C. elegans во время летаргуса: гомеостатическая динамика приступа, типичное положение тела и регуляция центральным нейроном» . Спать . 36 (3): 385–95. дои : 10.5665/Sleep.2456 . ПМЦ 3571756 . ПМИД 23449971 .
- ^ Хилл А.Дж., Мэнсфилд Р., Лопес Дж.М., Райзен Д.М., Ван Баскирк С. (октябрь 2014 г.). «Клеточный стресс вызывает у C. elegans защитное состояние, подобное сну» . Современная биология . 24 (20): 2399–405. Бибкод : 2014CBio...24.2399H . дои : 10.1016/j.cub.2014.08.040 . ПМК 4254280 . ПМИД 25264259 .
- ^ У маленьких безглазых червей есть совершенно новые светочувствительные клетки.
- ^ Scientific American, август 2018 г., стр. 14.
- ^ «Черви пережили катастрофу в Колумбии» . Новости Би-би-си . 1 мая 2003 года . Проверено 11 июля 2008 г.
- ^ «Университет отправляет червей в космос» . Новости Би-би-си . 17 января 2009 года . Проверено 9 июля 2009 г.
- ^ Клотц, I (16 мая 2011 г.). «Наследие космических червей, летающих на шаттле» . Новости Дискавери . Архивировано из оригинала 16 июня 2012 г. Проверено 17 мая 2011 г.
- ^ Сони, Пурушоттам; Анупом, Таслим; Лесанпезешки, Лейла; Рахман, Мизанур; Хьюитт, Дженнифер Э.; Веллоне, Мэтью; Стодик, Луи; Блавдзевич, Ежи; Шевчик, Натаниэль Дж.; Ванапалли, Шива А. (07 ноября 2022 г.). «Интегрированное в микрофлюидику космическое оборудование для измерения мышечной силы Caenorhabditis elegans на Международной космической станции» . npj Микрогравитация . 8 (1): 50. Бибкод : 2022npjMG...8...50S . дои : 10.1038/s41526-022-00241-4 . ISSN 2373-8065 . ПМЦ 9640571 . ПМИД 36344513 .
- ^ Стром С., Келли В.Г., Эркан С., Либ Дж.Д. (март 2014 г.). «Регуляция Х-хромосом у Caenorhabditis elegans » . Перспективы Колд-Спринг-Харбор в биологии . 6 (3): а018366. doi : 10.1101/cshperspect.a018366 . ПМЦ 3942922 . ПМИД 24591522 .
- ^ Консорциум секвенирования C. elegans (декабрь 1998 г.). «Последовательность генома нематоды C. elegans : платформа для изучения биологии». Наука . 282 (5396): 2012–8. Бибкод : 1998Наука...282.2012. . дои : 10.1126/science.282.5396.2012 . ПМИД 9851916 .
- ^ Перейти обратно: а б Сэдлер, Дж. Эван (1998). «Биохимия и генетика фактора Виллебранда» . Ежегодный обзор биохимии . 67 (1). Годовые обзоры : 395–424. doi : 10.1146/annurev.biochem.67.1.395 . ISSN 0066-4154 . ПМИД 9759493 .
- ^ Перейти обратно: а б Хан, Марк Э. (2002). «Арильные углеводородные рецепторы: разнообразие и эволюция». Химико-биологические взаимодействия . 141 (1–2). Эльзевир : 131–160. Бибкод : 2002CBI...141..131H . дои : 10.1016/s0009-2797(02)00070-4 . ISSN 0009-2797 . ПМИД 12213389 .
- ^ Перейти обратно: а б Бустело, Хосе Р. (2000). «Регуляторные и сигнальные свойства семейства вав» . Молекулярная и клеточная биология . 20 (5). Американское общество микробиологии : 1461–1477. дои : 10.1128/mcb.20.5.1461-1477.2000 . ISSN 0270-7306 . ПМЦ 85310 . ПМИД 10669724 .
- ^ Блюменталь Т., Эванс Д., Линк CD, Гуффанти А., Лоусон Д., Тьерри-Миг Дж., Тьерри-Миг Д., Чиу В.Л., Дьюк К., Кирали М., Ким СК (июнь 2002 г.). «Глобальный анализ оперонов Caenorhabditis elegans ». Природа . 417 (6891): 851–4. Бибкод : 2002Natur.417..851B . дои : 10.1038/nature00831 . ПМИД 12075352 . S2CID 4351788 .
- ^ Блюменталь Т. (ноябрь 2004 г.). «Опероны у эукариот» . Брифинги по функциональной геномике и протеомике . 3 (3): 199–211. дои : 10.1093/bfgp/3.3.199 . ПМИД 15642184 .
- ^ «Письмо о выпуске WS227» . ВормБейс . 10 августа 2011 года. Архивировано из оригинала 28 ноября 2013 года . Проверено 19 ноября 2013 г.
- ^ Руби Дж.Г., Ян С., Игрок С, Акстелл М.Дж., Ли В., Нусбаум С., Ге Х., Бартель Д.П. (декабрь 2006 г.). «Крупномасштабное секвенирование выявляет 21U-РНК, а также дополнительные микроРНК и эндогенные миРНК у C. elegans » . Клетка . 127 (6): 1193–207. дои : 10.1016/j.cell.2006.10.040 . ПМИД 17174894 .
- ^ Стриклин С.Л., Гриффитс-Джонс С., Эдди С.Р. (июнь 2005 г.). « C. elegans Гены некодирующей РНК » . Червячная книга : 1–7. дои : 10.1895/wormbook.1.1.1 . ПМЦ 4781554 . ПМИД 18023116 .
- ^ «Письмо о выпуске WS202» . ВормБейс . 29 мая 2009 года. Архивировано из оригинала 1 декабря 2013 года . Проверено 19 ноября 2013 г.
- ^ «Письмо о выпуске WS197» . ВормБейс . 27 ноября 2008 г. Архивировано из оригинала 17 октября 2019 г. Проверено 19 ноября 2013 г.
- ^ «Изменения последовательности генома» . ВормБейс . 15 июня 2011 г. Архивировано из оригинала 17 октября 2019 г. . Проверено 13 августа 2011 г.
- ^ Стейн Л.Д., Бао З., Блазиар Д., Блюменталь Т., Брент М.Р., Чен Н., Чинвалла А., Кларк Л., Кли С., Коглан А., Коулсон А., Д'Эстахио П., Фитч Д.Х., Фултон Л.А., Фултон Р.Э., Гриффитс-Джонс С., Харрис Т.В., Хиллиер Л.В., Камат Р., Кувабара П.Е., Мардис Э.Р., Марра М.А., Майнер Т.Л., Минкс П., Малликин Дж.К., Пламб Р.В., Роджерс Дж., Шейн Дж.Е., Сорманн М., Спит Дж., Стажич Дж.Е., Вэй С., Уилли Д., Уилсон Р.К., Дурбин Р., Уотерстон Р.Х. (ноябрь 2003 г.). «Последовательность генома Caenorhabditis briggsae : платформа для сравнительной геномики» . ПЛОС Биология . 1 (2): Е45. doi : 10.1371/journal.pbio.0000045 . ПМК 261899 . ПМИД 14624247 .
- ^ Центр секвенирования генома. « Caenorhabditis remanei : Предыстория» . Медицинский факультет Вашингтонского университета . Архивировано из оригинала 16 июня 2008 г. Проверено 11 июля 2008 г.
- ^ Центр секвенирования генома. « Caenorhabditis japonica : Предыстория» . Медицинский факультет Вашингтонского университета . Архивировано из оригинала 26 июня 2008 г. Проверено 11 июля 2008 г.
- ^ Стаден Р. (июнь 1979 г.). «Стратегия секвенирования ДНК с использованием компьютерных программ» . Исследования нуклеиновых кислот . 6 (7): 2601–10. дои : 10.1093/нар/6.7.2601 . ПМК 327874 . ПМИД 461197 .
- ^ «Браузер генома UCSC» . Проверено 8 июля 2014 г.
- ^ Кун Р.М., Карольчик Д., Цвейг А.С., Ван Т., Смит К.Е., Розенблум К.Р., Рхед Б., Рэйни Б.Дж., Пол А., Фазан М., Мейер Л., Сюй Ф., Хинрикс А.С., Харт Р.А., Джардин Б., Фудзита П., Дикханс М. , Дрессер Т., Клоусон Х., Барбер Г.П., Хаусслер Д., Кент В.Дж. (январь 2009 г.). «База данных браузера генома UCSC: обновление 2009 г.» . Исследования нуклеиновых кислот . 37 (Проблема с базой данных): D755–61. дои : 10.1093/нар/gkn875 . ПМЦ 2686463 . ПМИД 18996895 .
- ^ Феликс М.А., Брэндл С., Каттер А.Д. (2014). «Упрощенная система диагностики видов Caenorhabditis (Nematoda: Rhabditidae) с обозначениями названий для 15 различных биологических видов» . ПЛОС ОДИН . 9 (4): е94723. Бибкод : 2014PLoSO...994723F . дои : 10.1371/journal.pone.0094723 . ПМЦ 3984244 . ПМИД 24727800 .
- ^ Файр А, Сюй С, Монтгомери МК, Костас С.А., Драйвер С.Е., Мелло CC (февраль 1998 г.). «Мощное и специфическое генетическое вмешательство двухцепочечной РНК в Caenorhabditis elegans» . Природа . 391 (6669): 806–11. Бибкод : 1998Natur.391..806F . дои : 10.1038/35888 . ПМИД 9486653 . S2CID 4355692 .
- ^ Харрис Т.В., Антошечкин И., Биери Т., Блазиар Д., Чан Дж., Чен В.Дж. и др. (январь 2010 г.). «WormBase: комплексный ресурс по исследованию нематод» . Исследования нуклеиновых кислот . 38 (Проблема с базой данных): D463-7. дои : 10.1093/нар/gkp952 . ПМК 2808986 . ПМИД 19910365 .
Дальнейшее чтение
[ редактировать ]- Берд Дж, Берд AC (1991). Строение нематод . Академическая пресса . стр. 1, 69–70, 152–153, 165, 224–225. ISBN 978-0-12-099651-3 .
- Надежда, И.А. (1999). C. elegans : практический подход . Издательство Оксфордского университета . стр. 1–6. ISBN 978-0-19-963738-6 .
- Риддл Д.Л., Блюменталь Т., Мейер Р.Дж., Присс Дж.Р. (1997). К. Элеганс II . Лабораторный пресс Колд-Спринг-Харбор . стр. 1–4, 679–683. ISBN 978-0-87969-532-3 . ПМИД 21413221 .
Внешние ссылки
[ редактировать ]- Бреннер С. (2002) Дар природы науке. В. http://nobelprize.org/nobel_prizes/medicine/laureates/2002/brenner-lecture.pdf (также лекции Хорвица и Салстона)
- WormBase - обширная онлайн-база данных, охватывающая биологию и геномику C. elegans и других нематод.
- WormAtlas - онлайн-база данных по всем аспектам анатомии C. elegans с подробными пояснениями и высококачественными изображениями.
- WormBook - онлайн-обзор C. elegans биологии
- AceView WormGenes — еще одна база данных генома C. elegans , поддерживаемая NCBI.
- C. elegans II – бесплатный онлайн-учебник.
- Нейронная сеть WormWeb – онлайн-инструмент для визуализации и навигации по коннектому C. elegans.
- о C. elegans Фильмы – визуальное представление о C. elegans
- Просмотрите сборку генома ce11 в браузере генома UCSC .
- Caenorhabditis elegans на сайте eppo.int ( код ЕОКЗР CAEOEL)
- Цилиндр, Дрю (2022). «Использование C. elegans для изучения анестезии» . doi : 10.6084/m9.figshare.20963590.v1 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь )
| Базы данных органов управления : Национальные |
|---|