Асимметричное деление клеток
Асимметричное деление клеток приводит к образованию двух дочерних клеток с разной клеточной судьбой. Это контрастирует с симметричными клеточными делениями , которые дают начало дочерним клеткам с эквивалентной судьбой. Примечательно, что стволовые клетки делятся асимметрично, давая начало двум различным дочерним клеткам: одной копии исходной стволовой клетки, а также второй дочерней клетке, запрограммированной на дифференцировку в судьбу нестволовых клеток. (Во время роста или регенерации стволовые клетки также могут делиться симметрично, образуя две идентичные копии исходной клетки. [1] )
В принципе, существует два механизма, с помощью которых различные свойства могут быть переданы дочерним клеткам делящейся клетки. В одном из них дочерние клетки изначально эквивалентны, но разница индуцируется передачей сигналов между клетками, от окружающих клеток или от клетки-предшественника. Этот механизм известен как внешнее асимметричное деление клеток. При втором механизме предполагаемые дочерние клетки по своей природе различны во время деления материнской клетки. Поскольку этот последний механизм не зависит от взаимодействия клеток друг с другом или с окружающей средой, он должен опираться на внутреннюю асимметрию. Термин «асимметричное деление клеток» обычно относится к таким внутренним асимметричным делениям. [2]
Внутренняя асимметрия
[ редактировать ]Чтобы произошло асимметричное деление, материнская клетка должна быть поляризована , а митотическое веретено должно быть выровнено по оси полярности. Клеточная биология этих событий наиболее изучена на трех животных моделях : мышах , нематоде Caenorhabditis elegans и плодовой мушке Drosophila melanogaster . Позднее внимание было сосредоточено на развитии спирали .
В C. elegans развитии
[ редактировать ]У C. elegans серия асимметричных делений клеток на ранних стадиях эмбриона имеет решающее значение для формирования передней/задней, дорсальной/вентральной и левой/правой осей плана тела. [3] После оплодотворения уже происходят события, в зиготе позволяющие осуществить первое асимметричное деление клеток. Это первое деление производит два совершенно разных бластомера , называемых AB и P1. Когда сперматозоид оплодотворяет яйцеклетку спермия , пронуклеус и центросомы откладываются внутри яйцеклетки, что вызывает поток цитоплазмы, приводящий к движению пронуклеуса и центросом к одному полюсу. [4] Центросомы, отложенные сперматозоидом, отвечают за установление заднего полюса внутри зиготы. [5] Сперматозоиды с мутантными или отсутствующими центросомами не могут установить задний полюс. [6] [7] [8] Установление этой полярности инициирует поляризованное распределение группы белков, присутствующих в зиготе, называемых белками PAR (дефектное разделение), которые представляют собой консервативную группу белков, которые функционируют в установлении полярности клеток во время развития. [9] Эти белки первоначально распределяются равномерно по всей зиготе, а затем поляризуются с образованием заднего полюса. Эта серия событий позволяет одноклеточной зиготе обрести полярность за счет неравномерного распределения множества факторов.
Теперь отдельная клетка готова к асимметричному делению клеток, однако ориентация, в которой происходит деление, также является важным фактором. Митотическое веретено должно быть правильно ориентировано, чтобы гарантировать правильное распределение детерминант клеточной судьбы среди дочерних клеток. Выравнивание веретена опосредуется белками PAR, которые регулируют положение центросом вдоль оси A/P, а также движение митотического веретена вдоль оси A/P. [3] После этого первого асимметричного деления дочерняя клетка AB делится симметрично, давая начало ABa и ABp, в то время как дочерняя клетка P1 подвергается еще одному асимметричному делению клеток с образованием P2 и EMS. Это деление также зависит от распределения белков PAR. [10]
В дрозофилы развитии нервной системы
[ редактировать ]
У Drosophila melanogaster асимметричное деление клеток играет важную роль в развитии нейронов. Нейробласты — это клетки-предшественники, которые асимметрично делятся, образуя другой нейробласт и материнскую ганглиозную клетку (GMC). Нейробласт неоднократно подвергается асимметричному делению клеток, в то время как GMC продолжает производить пару нейронов. Два белка играют важную роль в создании асимметрии клеточных судеб в нейробластах: Prospero и Numb. Эти белки синтезируются в нейробластах и во время делений разделяются только на GMC. [11] Numb является супрессором Notch, поэтому асимметричная сегрегация Numb в базальной коре смещает ответ дочерних клеток на передачу сигналов Notch, что приводит к двум различным судьбам клеток. [12] Просперо необходим для регуляции генов в ГМК. Он равномерно распределяется по цитоплазме нейробласта, но локализуется в базальной коре, когда нейробласт начинает подвергаться митозу. Как только GMC отпочковывается от базальной коры, Просперо перемещается в ядро GMC и действует как фактор транскрипции. [11]
Другие белки, присутствующие в нейробластах, опосредуют асимметричную локализацию Numb и Prospero. Миранда — это якорный белок, который связывается с Просперо и удерживает его в базальной коре головного мозга. После создания GMC Миранда выпускает Просперо, а затем деградирует. [11] [13] Сегрегация Numb опосредуется Pon (партнером белка Numb). Pon связывается с Numb и колокализуется с ним во время деления клеток нейробластов. [11]
Митотическое веретено также должно располагаться параллельно асимметрично распределенным детерминантам клеточной судьбы, чтобы позволить им разделиться на одну дочернюю клетку, а не на другую. Ориентация митотического веретена опосредована Inscuteable, который отделяется от апикальной коры нейробласта. Без присутствия Inscuteable расположение митотического веретена и детерминант клеточной судьбы по отношению друг к другу становится рандомизированным. Неуязвимые мутанты демонстрируют равномерное распределение Миранды и Онемения в коре головного мозга, а полученные дочерние клетки демонстрируют идентичную судьбу нейронов. [11]
Помимо того, что две дочерние клетки имеют разную судьбу, они имеют разные размеры клеток; образовавшийся нейробласт намного больше, чем GMC. [14] Однако, в отличие от правильной сегрегации детерминант судьбы, асимметричное деление клеток, которое приводит к асимметрии размеров клеток, не зависит от веретена. [15] [16] Вместо этого этот механизм основан на пространственной и временной организации миозина в коре клетки и ее вышестоящих компонентах. Апикальная локализация Pins (партнера Inscuteable) с помощью Inscuteable позволяет Pins-зависимой апикальной локализации протеинкиназы N (Pkn) во время метафазы. Pkn ингибирует Rho-киназу (Rok) , что приводит к своевременной потере миозина и Rok из апикальной коры в начале анафазы. [17] [18] [19] Апикальный миозин течет базально туда, где расположена борозда расщепления. Впоследствии белки Tum и Pav в центральном веретене рекрутируют миозин для увеличения концентрации миозина, генерируя градиент миозина, управляющий апикальным потоком миозина из базальной коры. [19] [20] Этот пространственно-временной контроль локализации миозина приводит к асимметричной потере коркового напряжения, которое обычно противодействует гидростатическому давлению . Другими словами, потеря апикального кортикального миозина позволяет гидростатическому давлению давить на апикальную клеточную мембрану, увеличивая размер апикальной области, которая после деления клеток неизбежно станет более крупным нейробластом. [14] [19] Генерация апикальных и базальных потоков миозина одновременно приводит к симметричному делению клеток, а задержка базальных потоков миозина предотвращает нормальное расширение базальной области делящейся клетки. [14] [19] Хотя этот механизм не зависит от веретена, веретено важно для установления положения борозды расщепления, для доставки миозина в борозду расщепления и для обеспечения очистки базального миозина. [14] [19]
Корковые потоки на основе актомиозина управляют реорганизацией плазматической мембраны и клеточной коры нейробласта, которая необходима для создания разницы в размерах между дочерними клетками. [21] [22] [23] [24] В начале митоза корковые потоки собирают мембранные складки и выпячивания вокруг апикального полюса, образуя поляризованный мембранный резервуар. [21] [22] Когда миозин очищается от апикальной коры и проникновение в борозду расщепления приводит к увеличению гидростатического давления, запасы мембраны внутри резервуара используются для расширения апикальной области, которая после деления становится более крупной дочерней клеткой. [21]
В спиральном развитии
[ редактировать ]Спиралии (обычно синонимы lophotrochozoa ) представляют собой разнообразную кладу животных, виды которых составляют основную часть существующих сегодня двусторонних животных. Примеры включают моллюсков , кольчатых червей и энтопрокта . Хотя многое известно на клеточном и молекулярном уровне о других билатеральных кладах ( ecdysozoa и deuterostomia ), исследования процессов, которые управляют спиральным развитием, сравнительно недостаточны. Однако одной объединяющей чертой, общей для всех спиралий, является характер дробления у раннего эмбриона, известный как спиральное дробление . [25]
Механизмы асимметричного деления (см. рисунок, правая панель):
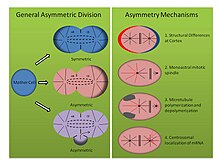
- Tubifex Tubefex: Было показано, что иловый червь Tubifex Tubefex демонстрирует интересное асимметричное деление клеток в момент первого эмбрионального дробления. В отличие от классической идеи о корковых различиях на зиготической мембране, которые определяют асимметрию веретена у эмбриона C. elegans , первое дробление трубочника зависит от количества центросом . [26] Эмбрионы наследуют одну центросому, которая локализуется в проспективной более крупной цитоплазме клеток CD и испускает радиальные микротрубочки во время анафазы, которые вносят вклад как в митотическое веретено, так и в кортикальные астры. Однако центр организации микротрубочек проспективной меньшей AB-клетки выделяет только микротрубочки, которые коммитируются с митотическим веретеном, а не кортикально-связанные астры. Когда эмбрионы сжимаются или деформируются, асимметричные веретена все еще формируются, а окрашивание на гамма-тубулин показывает, что во втором центре организации микротрубочек отсутствует молекулярная подпись центросомы. Более того, когда число центросом удваивается, эмбрионы трубочника делятся симметрично, указывая тем самым, что этот моноастральный механизм асимметричного клеточного деления зависит от центросомы. [26]
- Helobdella Robusta: Пиявка Helobdella Robusta демонстрирует такую же асимметрию в первом эмбриональном делении, что и C. elegans и трубочник , но использует модифицированный механизм. Эксперименты по сжатию эмбрионов робусты не влияют на асимметричное деление, что позволяет предположить, что этот механизм, как и трубочник, использует независимый от коры молекулярный путь. Окрашивание антителами у робусты показывает, что митотическое веретено формируется симметрично до метафазы и происходит из двух смещенных центросом. [27] В начале метафазы асимметрия становится очевидной, поскольку центросома проспективной более крупной клетки CD удлиняет кортикальные звездочки, в то время как астры проспективной меньшей AB-клетки становятся подавленными. Эксперименты с использованием нокодазола и таксола подтверждают это наблюдение. Таксол, который стабилизировал микротрубочки, при использовании в умеренной концентрации заставлял значительное количество эмбрионов расщепляться симметрично. Более того, эмбрионы, обработанные нокодазолом, который изолирует димеры тубулина и способствует деполимеризации микротрубочек, аналогичным образом вызывают симметричное деление у значительного числа эмбрионов. Лечение любым препаратом в этих концентрациях не может нарушить нормальную динамику центросом, указывая тем самым, что баланс полимеризации и деполимеризации микротрубочек представляет собой другой механизм установления асимметричного клеточного деления в развитии спилярий. [27]
- Ilyanasa obsoleta: Третий, менее традиционный механизм, способствующий асимметричному делению клеток при спиральном развитии, был обнаружен у моллюска Ilyanasa obsoleta . Эксперименты по гибридизации in situ и иммунофлуоресценции показывают, что транскрипты мРНК совместно локализуются с центросомами во время раннего расщепления. [28] Следовательно, эти транскрипты наследуются стереотипным образом в отдельные клетки. Все исследованные транскрипты мРНК были вовлечены в формирование паттерна оси тела, а гибридизация in situ транскриптов, связанных с другими функциями, не выявила такой локализации. Более того, нарушение полимеризации микротрубочек нокодазолом и полимеризации актина цитохализином B показывает, что цитоскелет также важен в этой асимметрии. Похоже, что микротрубочки не необходимы для рекрутирования мРНК в центросому, а актин необходим для прикрепления центросомы к коре головного мозга. Наконец, введение нескольких центросом в одну клетку путем ингибирования цитокинеза показывает, что мРНК надежно локализуется на правильной центросоме, что указывает на внутренние различия между каждой композицией центросом. Важно отметить, что эти результаты отражают эксперименты, проведенные после первых двух делений, но все же демонстрируют разные молекулярные способы установления асимметрии в делящейся клетке. [28]
В стволовых клетках и предшественниках
[ редактировать ]Животные состоят из огромного количества различных типов клеток . В ходе развития зигота претерпевает множество делений клеток, которые дают начало различным типам клеток, включая эмбриональные стволовые клетки. Асимметричное деление этих эмбриональных клеток приводит к появлению одной клетки одинаковой активности ( самообновления ), а другой может иметь такую же активность или стимулироваться для дальнейшей дифференцировки в специализированные типы клеток, такие как нейроны. Эта стимулированная дифференциация возникает под действием многих факторов, которые можно разделить на две широкие категории: внутренние и внешние. Внутренние факторы обычно включают различное количество детерминант клеточной судьбы, распределяемых в каждой дочерней клетке. Внешние факторы включают взаимодействие с соседними клетками, а также микро- и макроокружением клетки-предшественника. [29]
В дополнение к вышеупомянутому примеру с нейронами дрозофилы, было высказано предположение, что макросенсорные органы дрозофилы, в частности глиальные клетки, также возникают в результате сходного набора асимметричных делений единственной клетки-предшественника посредством регуляции сигнального пути Notch и факторов транскрипции . [30] Примером того, как внешние факторы вызывают это явление, является физическое вытеснение одной из дочерних клеток из исходной ниши стволовых клеток, подвергая ее воздействию сигнальных молекул, таких как хондроитинсульфат . [31] Таким образом, дочерняя клетка вынуждена взаимодействовать с сильно сульфатированными молекулами, которые стимулируют ее дифференцировку, в то время как другая дочерняя клетка остается в исходной нише в состоянии покоя.
Роль в болезни
[ редактировать ]В нормальных стволовых клетках и клетках -предшественниках асимметричное деление клеток уравновешивает пролиферацию и самообновление с выходом из клеточного цикла и дифференцировкой. Нарушение асимметричного деления клеток приводит к аберрантному самообновлению и нарушению дифференцировки и, следовательно, может представлять собой раннюю стадию туморогенной трансформации стволовых клеток и клеток-предшественников. В нормальных неопухолевых стволовых клетках описан ряд генов, отвечающих за плюрипотентность, таких как Bmi-1 , Wnt и Notch . Эти гены были обнаружены также в случае раковых стволовых клеток и показывают, что их аберрантная экспрессия важна для формирования массы опухолевых клеток. [32] Например, было показано, что рак желудочно-кишечного тракта содержит редкую субпопуляцию раковых стволовых клеток, способных делиться асимметрично. Асимметричное деление в этих клетках регулируется раковой нишей (микроокружением) и путем Wnt. Блокирование пути Wnt с помощью IWP2 (антагониста WNT) или siRNA-TCF4 приводило к сильному подавлению асимметричного деления клеток. [33]
Другая мутация асимметричного деления клеток, которая участвует в росте опухоли, — это мутации потери функции. Первое предположение о том, что потеря асимметричного деления клеток может быть вовлечена в онкогенез, появилось в исследованиях дрозофилы . Исследования мутаций потери функции в ключевых регуляторах асимметричного деления клеток, включая lgl, aurA, polo, numb и brat, выявили гиперпролиферативные фенотипы in situ. У этих мутантов клетки делятся более симметрично и генерируют неправильно определенное потомство, которое не может выйти из клеточного цикла и дифференцироваться, а вместо этого непрерывно пролиферирует и образует массу опухолевых клеток. [34]
Ссылки
[ редактировать ]- ^ Моррисон С.Дж., Кимбл Дж. (июнь 2006 г.). «Асимметричные и симметричные деления стволовых клеток в развитии и раке». Природа . 441 (7097): 1068–1074. Бибкод : 2006Natur.441.1068M . дои : 10.1038/nature04956 . hdl : 2027.42/62868 . ПМИД 16810241 . S2CID 715049 .
- ^ Хокинс Н., Гаррига Дж. (декабрь 1998 г.). «Асимметричное деление клеток: от А до Я» . Гены и развитие . 12 (23): 3625–3638. дои : 10.1101/gad.12.23.3625 . ПМИД 9851969 .
- ^ Перейти обратно: а б Генчи П., Роуз Л.С. (октябрь 2005 г.). «Асимметричное деление клеток и формирование оси у эмбриона» . Червячная книга : 1–20. дои : 10.1895/wormbook.1.30.1 . ПМЦ 4780927 . ПМИД 18050411 .
- ^ Гольдштейн Б., Хирд С.Н. (май 1996 г.). «Характеристика передне-задней оси Caenorhabditis elegans». Разработка . 122 (5): 1467–74. дои : 10.1242/dev.122.5.1467 . ПМИД 8625834 .
- ^ Коуэн Ч.Р., Хайман А.А. (сентябрь 2004 г.). «Центросомы управляют полярностью клеток независимо от сборки микротрубочек у эмбрионов C. elegans». Природа . 431 (7004): 92–96. Бибкод : 2004Natur.431...92C . дои : 10.1038/nature02825 . ПМИД 15343338 . S2CID 4422297 .
- ^ О'Коннелл К.Ф., Максвелл К.Н., Уайт Дж.Г. (июнь 2000 г.). «Ген spd-2 необходим для поляризации переднезадней оси и образования звезд сперматозоидов в зиготе Caenorhabditis elegans» . Биология развития . 222 (1): 55–70. дои : 10.1006/dbio.2000.9714 . ПМИД 10885746 .
- ^ Хэмилл Д.Р., Северсон А.Ф., Картер Дж.К., Бауэрман Б. (ноябрь 2002 г.). «Для созревания центросом и сборки митотического веретена у C. elegans требуется SPD-5, белок с несколькими спиральными доменами» . Развивающая клетка . 3 (5): 673–684. дои : 10.1016/s1534-5807(02)00327-1 . ПМИД 12431374 .
- ^ Сэдлер П.Л., Шейкс, округ Колумбия (январь 2000 г.). «Безъядерные сперматозоиды Caenorhabditis elegans могут ползать, оплодотворять ооциты и направлять передне-заднюю поляризацию одноклеточного эмбриона». Разработка . 127 (2): 355–366. дои : 10.1242/dev.127.2.355 . ПМИД 10603352 .
- ^ Чикс Р.Дж., Кэнман Дж.К., Габриэль В.Н., Мейер Н., Строум С., Гольдштейн Б. (май 2004 г.). «Белки PAR C. elegans функционируют путем мобилизации и стабилизации асимметрично локализованных белковых комплексов» . Современная биология . 14 (10): 851–862. Бибкод : 2004CBio...14..851C . дои : 10.1016/j.cub.2004.05.022 . ПМИД 15186741 .
- ^ Шнайдер С.К., Бауэрман Б. (2003). «Полярность клеток и цитоскелет в зиготе Caenorhabditis elegans». Ежегодный обзор генетики . 37 : 221–249. дои : 10.1146/annurev.genet.37.110801.142443 . ПМИД 14616061 .
- ^ Перейти обратно: а б с д и Мацузаки Ф (февраль 2000 г.). «Асимметричное деление нервных стволовых клеток дрозофилы: основа нервного разнообразия». Современное мнение в нейробиологии . 10 (1): 38–44. дои : 10.1016/s0959-4388(99)00052-5 . ПМИД 10679433 . S2CID 187054 .
- ^ Го М, Ян Л.Ю., Ян Ю.Н. (июль 1996 г.). «Контроль судеб дочерних клеток во время асимметричного деления: взаимодействие Numb и Notch» . Нейрон . 17 (1): 27–41. дои : 10.1016/s0896-6273(00)80278-0 . ПМИД 8755476 .
- ^ Икешима-Катаока Х., Скит Дж.Б., Набешима Ю., Доу CQ, Мацузаки Ф (декабрь 1997 г.). «Миранда направляет Просперо к дочерней клетке во время асимметричных делений дрозофилы». Природа . 390 (6660): 625–629. Бибкод : 1997Natur.390..625I . дои : 10.1038/37641 . ПМИД 9403694 . S2CID 4423032 .
- ^ Перейти обратно: а б с д Фам Т.Т., Моннард А., Хелениус Дж., Лунд Э., Ли Н., Мюллер DJ, Кабернард С. (март 2019 г.). «Пространственно-временно-контролируемая релокализация миозина и внутреннее давление вызывают асимметрию размеров родственных клеток» . iScience . 13 :9–19. Бибкод : 2019iSci...13....9P . дои : 10.1016/j.isci.2019.02.002 . ПМК 6383127 . ПМИД 30785031 .
- ^ Кабернар С., Прехода К.Е., Доу CQ (сентябрь 2010 г.). «Независимый от шпинделя путь позиционирования борозды расщепления» . Природа . 467 (7311): 91–94. Бибкод : 2010Natur.467...91C . дои : 10.1038/nature09334 . ПМК 4028831 . ПМИД 20811457 .
- ^ Коннелл М., Кабернард К., Рикетсон Д., Доу CQ, Прехода К.Е. (ноябрь 2011 г.). «Асимметричное расширение коры меняет положение борозды расщепления в нейробластах дрозофилы» . Молекулярная биология клетки . 22 (22): 4220–4226. дои : 10.1091/mbc.e11-02-0173 . ПМК 3216648 . ПМИД 21937716 .
- ^ Хомем CC, Кноблих Я.А. (декабрь 2012 г.). «Нейробласты дрозофилы: модель биологии стволовых клеток» . Разработка . 139 (23): 4297–4310. дои : 10.1242/dev.080515 . ПМИД 23132240 . S2CID 14960710 .
- ^ Цанкова А, Фам Т.Т., Гарсия Д.С., Отте Ф., Кабернар К. (июль 2017 г.). «Полярность клеток регулирует смещенную активность и динамику миозина во время асимметричного деления клеток с помощью Rho-киназы дрозофилы и протеинкиназы N» . Развивающая клетка . 42 (2): 143–155.e5. дои : 10.1016/j.devcel.2017.06.012 . ПМИД 28712722 .
- ^ Перейти обратно: а б с д и Рубине С., Цанкова А., Фам Т.Т., Моннар А., Коссинус Е., Аффольтер М., Кабернар С. (ноябрь 2017 г.). «Пространственно-временное разделение корковых потоков и геометрия веретена создают физическую асимметрию в нервных стволовых клетках мух» . Природные коммуникации . 8 (1): 1383. Бибкод : 2017NatCo...8.1383R . дои : 10.1038/s41467-017-01391-w . ПМЦ 5680339 . ПМИД 29123099 .
- ^ Майер М., Депкен М., Буа Дж.С., Юлихер Ф., Grill SW (сентябрь 2010 г.). «Анизотропия коркового напряжения раскрывает физическую основу поляризации корковых потоков» . Природа . 467 (7315): 617–621. Бибкод : 2010Natur.467..617M . дои : 10.1038/nature09376 . ПМИД 20852613 . S2CID 4378520 .
- ^ Перейти обратно: а б с ЛаФойя, Брайс; Прехода, Кеннет Э. (апрель 2023 г.). «Потребление поляризованного мембранного резервуара приводит к асимметричному расширению мембраны во время неравных делений нервных стволовых клеток» . Развивающая клетка . 58 (11): 993–1003.e3. дои : 10.1016/j.devcel.2023.04.006 . ПМЦ 10247545 . ПМИД 37116487 .
- ^ Перейти обратно: а б ЛаФойя, Брайс; Прехода, Кеннет Э. (май 2021 г.). «Актин-зависимая поляризация мембраны раскрывает механическую природу цикла полярности нейробластов» . Отчеты по ячейкам . 35 (7): 109146. doi : 10.1016/j.celrep.2021.109146 . ПМК 8174105 . ПМИД 34010656 .
- ^ Ун, Чет Хуан; Прехода, Кеннет Э (15 ноября 2021 г.). «Фазы динамики коркового актомиозина, связанные с циклом полярности нейробластов» . электронная жизнь . 10 : е66574. дои : 10.7554/eLife.66574 . ISSN 2050-084X . ПМК 8641948 . ПМИД 34779402 .
- ^ Ун, Чет Хуан; Прехода, Кеннет Э (08 мая 2019 г.). «Асимметричное рекрутирование и актин-зависимые корковые потоки управляют циклом полярности нейробластов» . электронная жизнь . 8 : е45815. doi : 10.7554/eLife.45815 . ISSN 2050-084X . ПМК 6524966 . ПМИД 31066675 .
- ^ Генри Джей-Джей, Мартиндейл MQ (1999). «Сохранение и инновации в спиральном развитии». Репродуктивные стратегии и закономерности развития кольчатых червей . Том. 402. стр. 255–65. дои : 10.1007/978-94-017-2887-4_15 . ISBN 978-90-481-5340-4 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Перейти обратно: а б Симидзу Т., Исии Р., Такахаши Х. (июнь 1998 г.). «Неравномерное дробление раннего эмбриона трубочника». Развитие, рост и дифференциация . 40 (3): 257–266. дои : 10.1046/j.1440-169x.1998.00001.x . ПМИД 9639353 . S2CID 23026919 .
- ^ Перейти обратно: а б Рен Икс, Вейсблат Д.А. (апрель 2006 г.). «Асимметризация первого расщепления за счет временной разборки одного веретена полюсной астры у пиявки Helobdellarobusta» . Биология развития . 292 (1): 103–115. дои : 10.1016/j.ydbio.2005.12.049 . ПМИД 16458880 .
- ^ Перейти обратно: а б Ламберт Дж. Д., Надь Л. М. (декабрь 2002 г.). «Асимметричное наследование центросомально локализованных мРНК во время эмбрионального дробления». Природа . 420 (6916): 682–686. Бибкод : 2002Natur.420..682L . дои : 10.1038/nature01241 . ПМИД 12478296 . S2CID 4383189 .
- ^ Ян Ю.Н., Ян Л.И. (апрель 1998 г.). «Асимметричное деление клеток». Природа . 392 (6678): 775–778. Бибкод : 1998Natur.392..775J . дои : 10.1038/33854 . ПМИД 9572136 . S2CID 4392481 .
- ^ Го М., Беллаиш Ю., Швайсгут Ф. (август 1999 г.). «Возвращаясь к линии микрохет Drosophila: новое по сути асимметричное деление клеток приводит к образованию глиальных клеток». Разработка . 126 (16): 3573–84. дои : 10.1242/dev.126.16.3573 . ПМИД 10409503 .
- ^ Хейс А.Дж., Тюдор Д., Ноуэлл М.А., Катерсон Б., Хьюз К.Э. (февраль 2008 г.). «Мотивы сульфатирования хондроитина как предполагаемые биомаркеры для выделения клеток-предшественников суставного хряща» . Журнал гистохимии и цитохимии . 56 (2): 125–138. дои : 10.1369/jhc.7a7320.2007 . ПМК 2324172 . ПМИД 17938280 .
- ^ Гомес-Лопес С., Лернер Р.Г., Петрич С. (февраль 2014 г.). «Асимметричное деление стволовых клеток и клеток-предшественников во время гомеостаза и рака» . Клеточные и молекулярные науки о жизни . 71 (4): 575–597. дои : 10.1007/s00018-013-1386-1 . ПМК 3901929 . ПМИД 23771628 .
- ^ Синь Х.В., Амбе С.М., Рэй С., Ким Б.К., Коидзуми Т., Виганд Г.В. и др. (2013). «Wnt и раковая ниша: паракринные взаимодействия с раковыми клетками желудочно-кишечного тракта, подвергающимися асимметричному клеточному делению» . Журнал рака . 4 (6): 447–457. дои : 10.7150/jca.6896 . ПМЦ 3726705 . ПМИД 23901343 .
- ^ Гонсалес С. (июнь 2007 г.). «Ориентация веретена, асимметричное деление и подавление опухоли в стволовых клетках дрозофилы». Обзоры природы. Генетика . 8 (6): 462–472. дои : 10.1038/nrg2103 . ПМИД 17510666 . S2CID 22558696 .
Дальнейшее чтение
[ редактировать ]- Масиейра-Коэльо А, изд. (2007). Асимметричное деление клеток . Прогресс молекулярной и субклеточной биологии. Том. 45. Берлин, Гейдельберг, Нью-Йорк: Springer Verlag. ISBN 978-3-540-69160-0 .