Конодонт
| Конодонты Временной диапазон: | |
|---|---|
 | |
| Элементы конодонта | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Подтип: | Позвоночные животные |
| Сорт: | † Конодонта Пандер , 1856 г. |
| Подгруппы | |
| |
| Синонимы | |
Конодонты ( греч. kōnos , « конус », + odont , « зуб ») — вымершая группа бесчелюстных похожих на , (бесчелюстных) позвоночных животных угрей , отнесенных к классу Conodonta . В течение многих лет они были известны только по окаменелостям их шипастых оральных элементов, которые обычно встречаются изолированно и сейчас называются элементами конодонтов , тогда как знания о мягких тканях остаются ограниченными. Выносливая группа доисторических рыб , конодонты, существовала в Мировом океане более 300 миллионов лет, от кембрия до начала юры . Из-за своего космополитического распространения элементы конодонтов широко используются в качестве индексных окаменелостей , окаменелостей, используемых для определения и идентификации геологических периодов .
Открытие и понимание конодонтов
[ редактировать ]Зубоподобные окаменелости конодонта были впервые обнаружены Хайнцем Кристианом Пандером , а результаты опубликованы в Санкт-Петербурге, Россия , в 1856 году. [2]
Лишь в начале 1980-х годов были найдены первые ископаемые останки животного (см. ниже). В 1990-х годах в Южной Африке были найдены изысканные окаменелости, мягкие ткани которых превратились в глину, сохранив даже мышечные волокна. Наличие мышц для вращения глаз окончательно показало, что животные были примитивными позвоночными. [3]
Номенклатура и таксономический ранг
[ редактировать ]На протяжении всей истории изучения «конодонт» — это термин, который применялся как к отдельным окаменелостям, так и к животным, которым они принадлежали. Первоначальный немецкий термин, использованный Пандером, был «conodonten», который впоследствии был переведен на английский язык как «conodonts», хотя формальное латинизированное название не было предоставлено в течение нескольких десятилетий. Макфарлейн (1923) описал их как отряд Conodontes (греческий перевод), который Хаддл (1934) изменил на латинское написание Conodonta. [4] Несколькими годами ранее Эйхенберг (1930) установил другое название животных, ответственных за окаменелости конодонтов: Conodontophorida («носители конодонтов»). [1] Несколько других научных названий редко и непоследовательно применялись к конодонтам и их предполагаемым близким родственникам в течение 20 века, например Conodontophoridia, Conodontophora, Conodontochordata, Conodontiformes, [5] и конодонтоморфа.
Conodonta и Conodontophorida на сегодняшний день являются наиболее распространенными научными названиями, используемыми для обозначения конодонтов, хотя несоответствия относительно их таксономического ранга все еще сохраняются. Исследование Бенгтсона (1976) по эволюции конодонтов выявило три морфологических уровня ранних конодонтоподобных окаменелостей: протоконодонты , параконодонты и «настоящие конодонты» (эуконодонты). [5] Дальнейшие исследования показали, что протоконодонты, вероятно, были более тесно связаны с хетогнатами (стрелочервями), чем с настоящими конодонтами. С другой стороны, параконодонты по-прежнему считаются вероятным предком или сестринской группой эвконодонтов.
1981 года В томе «Трактата о палеонтологии беспозвоночных» , посвященном конодонтам (пересмотренная часть W, приложение 2), Conodonta упоминается как название как типа , так и класса , а Conodontophorida - как подчиненный отряд для «настоящих конодонтов». Все три ранга были приписаны Эйхенбергу, а Параконодонтида также была включена в состав ордена Конодонта. [6] Этот подход подвергся критике со стороны Фореуса (1983), который утверждал, что он упускает из виду историческую значимость Пандера как основателя и главной фигуры в конодонтологии. Фореус предложил сохранить Conodonta как тип (приписываемый Пандеру) с одним классом Conodontata (Pander) и одним отрядом Conodontophorida (Eichenberg). [4] [7] Последующие авторы продолжали рассматривать Conodonta как тип с постоянно растущим числом подгрупп. [8]
Учитывая все более убедительные доказательства того, что конодонты относятся к типу хордовых, в более поздних исследованиях «настоящие конодонты» обычно относят к классу Conodonta, включающему несколько более мелких отрядов. [9] [10] [11] Параконодонты обычно исключаются из этой группы, хотя по-прежнему считаются близкими родственниками. [9] [10] [11] На практике Conodonta, Conodontophorida и Euconodonta являются эквивалентными терминами и используются как взаимозаменяемые.
Элементы конодонта
[ редактировать ]Одинокие элементы
[ редактировать ]Элементы конодонта состоят из минерализованных зубоподобных структур различной морфологии и сложности. Эволюция минерализованных тканей остается загадкой уже более столетия. Высказано предположение, что первый механизм минерализации тканей хордовых начался либо в ротовом скелете конодонтов, либо в дермальном скелете ранних бесчелюстных .
Массив элементов представлял собой кормовой аппарат, радикально отличающийся от челюстей современных животных. Теперь их называют «элементами конодонтов», чтобы избежать путаницы. Три формы зубов: конусообразные шишки, разветвленные бруски и гребенчатые платформы, вероятно, выполняли разные функции.
В течение многих лет конодонты были известны только по загадочным зубообразным микроокаменелостям (длиной от 200 микрометров до 5 миллиметров). [12] ), которые обычно, но не всегда, встречаются изолированно и не связаны ни с какими другими ископаемыми. До начала 1980-х годов зубы конодонтов не были обнаружены в сочетании с окаменелостями организма-хозяина в консервационных лагерштетте . [13] Это связано с тем, что у конодонта было мягкое тело, поэтому все, кроме зубов, было непригодно для сохранения при нормальных обстоятельствах.
Эти микроокаменелости состоят из гидроксиапатита (фосфатного минерала). [14] Элементы конодонтов можно извлечь из горных пород с использованием подходящих растворителей. [15] [16] [17]
Они широко используются в биостратиграфии . Элементы конодонта также используются в качестве палеотермометров , индикатора термических изменений во вмещающей породе, поскольку при более высоких температурах фосфат претерпевает предсказуемые и постоянные изменения цвета, измеряемые с помощью индекса изменения конодонта . Это сделало их полезными для разведки нефти там, где они известны, в породах, датируемых периодом от кембрия до позднего триаса .
Полный аппарат
[ редактировать ]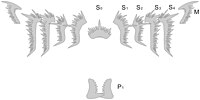 Полный набор элементов конодонта Hindeodus parvus
Полный набор элементов конодонта Hindeodus parvus Сохранившаяся сочлененная ассоциация элементов конодонтов вида Archeognathus primus (ордовик, Северная Америка)
Сохранившаяся сочлененная ассоциация элементов конодонтов вида Archeognathus primus (ордовик, Северная Америка)


Конодонтный аппарат может состоять из ряда дискретных элементов, в том числе спатогнатиформной, озаркодиноформной, трихоноделлиформной, неоприониодиформной и других форм. [18]
В 1930-х годах концепцию комплексов конодонтов описал Герман Шмидт. [19] и Гарольда В. Скотта в 1934 году. [20] [21] [22] [23]
Элементы озаркодинид
[ редактировать ]Питательный аппарат озаркодинид состоит из осевого элемента Sa спереди, фланкированного двумя группами из четырех близко расположенных удлиненных элементов Sb и Sc, наклоненных наклонно внутрь и вперед. Над этими элементами лежала пара дугообразных и направленных внутрь (макелатных) элементов М. За массивом СМ лежали поперечно ориентированные и билатерально противоположные (пектинообразные, т.е. гребенчатые) элементы Pb и Pa. [24]
Конодонт животное
[ редактировать ] Восстановление жизни прекрасного обещания
Восстановление жизни прекрасного обещания Восстановление Panderodus unicostatus
Восстановление Panderodus unicostatus Ископаемое тело Panderodus unicostatus.
Ископаемое тело Panderodus unicostatus. Сравнение размеров трех видов конодонтов с сохранившимися окаменелостями тел.
Сравнение размеров трех видов конодонтов с сохранившимися окаменелостями тел. Окаменелости Typhloesus , когда-то считавшиеся первым ископаемым телом конодонта.
Окаменелости Typhloesus , когда-то считавшиеся первым ископаемым телом конодонта.




Хотя элементы конодонтов широко распространены в летописи окаменелостей, окаменелости, сохранившие мягкие ткани конодонтов, известны лишь из нескольких месторождений в мире. Одними из первых возможных окаменелостей конодонта были окаменелости Typhloesus , загадочного животного, известного из известняка Медвежьего ущелья в Монтане . [25] Эта возможная идентификация была основана на присутствии элементов конодонта в окаменелостях Typhloesus . зоне существа Однако это утверждение было опровергнуто, поскольку элементы конодонтов на самом деле находились в пищеварительной . [26] Это животное теперь рассматривается как возможный моллюск , родственный брюхоногим моллюскам . [26] По состоянию на 2023 год существует только три описанных вида конодонтов, у которых сохранились окаменелости туловища: Clydagnathuswindsorensis из каменноугольного возраста пласта креветок Грантон в Шотландии , Promissum pulchrum из ордовикского возраста сланцев Сум в Южной Африке и Panderodus unicostatus из силурийского возраста Уакеша. Биота в Висконсине . [9] [27] [28] Есть и другие примеры конодонтов, у которых сохранилась только область головы, включая глаза, у животных, известных из стоянки Эрамоза силурийского возраста в Онтарио и триасового возраста секции Аккамори в Японии . [29] [30]
Судя по этим окаменелостям, у конодонтов были большие глаза, плавники с плавниковыми лучами, шевронообразные мышцы и осевая линия, которые интерпретировались как хорда или спинной нервный канатик . [27] [31] В то время как Clydagnathus и Panderodus имели длину всего 4–5 см (1,6–2,0 дюйма), Promissum , по оценкам, достигал 40 см (16 дюймов) в длину, если бы он имел те же пропорции, что и Clydagnathus . [27] [28]
Экология
[ редактировать ]
«Зубы» некоторых конодонтов интерпретируются как аппараты-фильтраторы, фильтрующие планктон из воды и пропускающие его в глотку. [32] Другие интерпретировались как «схватывающий и сокрушающий массив». [28] Износ некоторых элементов конодонта позволяет предположить, что они функционировали как зубы: следы износа, вероятно, образовались в результате еды, а также в результате окклюзии другими элементами. [33] Исследования пришли к выводу, что таксоны конодонтов занимали как пелагические (открытый океан), так и нектобентические (плавающие над поверхностью отложений) ниши. [33] Сохранившаяся мускулатура позволяет предположить, что некоторые конодонты ( по крайней мере, Промиссум ) были эффективными крейсерами, но неспособными развивать максимальную скорость. [28] На основании изотопных данных было высказано предположение, что некоторые девонские конодонты были потребителями низкого уровня и питались зоопланктоном . [33]
исследование динамики популяций Alternognathus Опубликовано . Среди прочего, это демонстрирует, что, по крайней мере, этот таксон имел короткую продолжительность жизни, продолжавшуюся около месяца. [34] Исследование соотношений Sr / Ca и Ba /Ca в популяции конодонтов карбонатной платформы силура Швеции показало, что разные виды и роды конодонтов, вероятно, занимали разные трофические ниши . [33]
Некоторые виды рода Panderodus считаются ядовитыми на основании борозд, обнаруженных на некоторых элементах. [35]
Классификация и филогения
[ редактировать ]Родство
[ редактировать ]По состоянию на 2012 год [update]Ученые относят конодонтов к типу Хордовые на основании их плавников с плавниковыми лучами, шевронообразных мышц и хорды . [36]
Милсом и Ригби представляют их как позвоночных, внешне похожих на современных миксин и миног. [37] и филогенетический анализ предполагает, что они более производны , чем любая из этих групп. [9] Однако этот анализ имеет одну оговорку: самые ранние окаменелости, похожие на конодонтов, протоконодонты , по-видимому, образуют отдельную кладу от более поздних параконодонтов и эвконодонтов . Протоконодонты, вероятно, не являются родственниками настоящих конодонтов, но, вероятно, представляют собой стволовую группу Chaetognatha , неродственного типа, включающего стрелковых червей. [38]
Более того, некоторые анализы не рассматривают конодонтов ни как позвоночных , ни как краниатов , поскольку у них отсутствуют основные характеристики этих групп. [39] Совсем недавно было высказано предположение, что конодонты могут быть стеблевыми круглоротыми , более близкими родственниками миксинам и миногам, чем челюстным позвоночным животным . [40]
Внутригрупповые отношения
[ редактировать ]Отдельные элементы конодонтов трудно последовательно классифицировать, но в настоящее время известно все большее число видов конодонтов из многоэлементных комплексов, которые дают больше данных, позволяющих сделать вывод о том, как различные линии конодонтов связаны друг с другом. Ниже приводится упрощенная кладограмма, основанная на Свите и Донохью (2001): [10] который суммировал предыдущую работу Sweet (1988). [8] и Донохью и др. (2000): [9]
Лишь несколько исследований подходят к вопросу внутригрупповых отношений конодонтов с кладистической точки зрения, о чем свидетельствуют данные филогенетического анализа . Одним из самых широких исследований такого рода был анализ Donoghue et al. (2008), в которых основное внимание уделялось «сложным» конодонтам (Prioniodontida и другим группам потомков): [11]
Эволюционная история
[ редактировать ]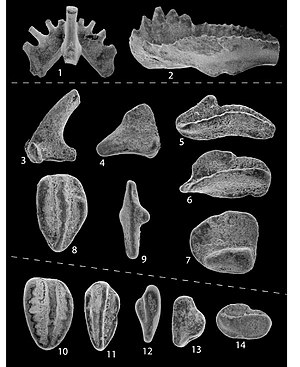
Самые ранние окаменелости конодонтов известны из кембрийского периода. Конодонты значительно разнообразились в раннем ордовике, достигнув пика разнообразия в средней части периода, и испытали резкое снижение в позднем ордовике и силуре, прежде чем достичь нового пика разнообразия в середине позднего девона. Разнообразие конодонтов сократилось в каменноугольном периоде с вымиранием в конце среднего турне. [41] и длительный период значительной потери разнообразия в Пенсильванском периоде . [42] [43] В пермском периоде присутствовало лишь несколько родов конодонтов, хотя разнообразие увеличилось после вымирания PT в раннем триасе.
Разнообразие продолжало сокращаться в течение среднего и позднего триаса, что привело к их исчезновению вскоре после границы триаса и юры. Большая часть их разнообразия в палеозое, вероятно, контролировалась уровнем моря и температурой, причем основное снижение произошло в позднем ордовике и позднем карбоне из-за более низких температур, особенно ледниковых явлений и связанных с ними морских регрессий , которые сократили площадь континентального шельфа . Однако их окончательная гибель, скорее всего, связана с биотическими взаимодействиями , возможно, конкуренцией с новыми мезозойскими таксонами. [44]
Таксономия
[ редактировать ]Таксономия Conodonta основана на Sweet (1988), [8] Свит и Донохью (2001), [10] и Филогенический архив Микко. [45] [ нужны разъяснения ]
- Класс Conodonta Pander, 1856 [ Conodontophorida Eichenberg, 1930; «эуконодонты» Бенгтсон, 1976 г. ]
- Кавидонти Сладкий, 1988 год.
- Заказать Белоделлиду ? Сладкий, 1988 год.
- Анселлиды ? Фореус и Хантер, 1985 г.
- Belodellidae Ходалевич и Черних, 1973 г.
- Дапсилодонтиды ? Сладкий, 1988 год.
- Отряд Proconodontida Sweet, 1988 г.
- Cordylodontidae Линдстрём, 1970 г.
- Fryxellodontidae Миллер, 1981 г.
- Псевдоонеотодиды ? Ван и Олдридж, 2010 г.
- Проконодонтиды Линдстрём, 1970 г.
- Пигодонтиды ? Бергстрем, 1981 г.
- Заказать Белоделлиду ? Сладкий, 1988 год.
- Конодонти Пандер, 1856 г., не Брэнсон, 1938 г.
- Отряд Protopanderodontida Sweet, 1988 [Panderodontida Sweet, 1988 ]
- Акантодонтиды Линдстрём, 1970 г.
- Клавохамулиды Линдстрём, 1970 г.
- Дрепаноистодонтиды ? Fåhraeus, 1978 [Distacodontidae Bassler, 1925 ]
- Пандеродонтиды Линдстрём, 1970 г.
- Protopanderodontidae Lindström, 1970 [Scolopodontidae Bergström, 1981 ; Oneotodontidae Miller, 1981 ; Теридонтиды Миллер, 1981 ]
- Серратогнатиды ? Чжэнь и др., 2009 г.
- Страханогнатиды ? Бергстрем, 1981 [Cornuodontidae Fåhraeus, 1966 ]
- Отряд Prioniodontida Dzik, 1976 (парафилетический)
- Акодонтиды ? Кабан, 1993 г. [Tripodontinae Sweet, 1988 ]
- Кахабаннатиды ? Стоудж и Баньоли, 1999 г.
- Дистакодонтиды ? Басслер, ред. 1925 г. Ульрих и Басслер, 1926 г. [Drepanodontinae Fåhraeus & Nowlan, 1978 ; Лончодининае Хасс, 1959 ]
- Гамахигнатиды ? Ван и Олдридж, 2010 г.
- Яблоннодонтиды ? Кабан, 2006 г.
- Нурреллиды ? Помешано-Черки, 1967 г.
- Паракордилодонтиды ? Бергстрем, 1981 г.
- Плейфордииды ? Кабан, 2002 г.
- Ульриходиниды ? Бергстрем, 1981 г.
- Россодус Репецкий и Этингтон, 1983 г.
- Multioistodontidae Harris, 1964 [Dischidognathidae]
- Oistodontidae Lindström, 1970 [Juanognathidae Bergström, 1981 ]
- Периодонтиды Линдстрём, 1970 г.
- Rhipidognathidae Lindström, 1970 в смысле Sweet, 1988
- Приониодонтиды Басслер, 1925 г.
- Phragmodontidae Bergström, 1981 [Cyrtoniodontinae Hass, 1959 ]
- Плектодиниды Сладкие, 1988 г.
- Пигодонтиды ? Бергстрем, 1981 г.
- Икриодонтацея
- Балогнатиды (Хасс, 1959)
- Polyplacognathidae Bergström, 1981 г.
- Дистомодонтиды Клаппер, 1981 г.
- Icriodellidae Sweet, 1988 г.
- Icriodontidae Мюллер и Мюллер, 1957 г.
- Отряд Prioniodinida Sweet, 1988 г.
- Оэпикодонтиды ? Бергстрем, 1981 г.
- Ксаниогнатиды ? Сладкое, 1981 г.
- Chirognathidae Брэнсон и Мель, 1944 г.
- Prioniodinidae Bassler, 1925 [Hibbardellidae Mueller, 1956 ]
- Бактрогнатиды Линдстрем, 1970 г.
- Эллисонииды Кларк, 1972 г.
- Гондоллиды Линдстрём, 1970 г.
- Отряд Озаркодинида Дзик, 1976 [Polygnathida]
- Анчингнатодонтиды ? Кларк, 1972 год.
- Археогнатиды ? Миллер, 1969 год.
- Белодонтиды ? Хаддл, 1934 год.
- Колеодонтиды ? Branson & Mehl, 1944 [Hibbardellidae Müller, 1956 ; Локсодонтиды]
- Эогнатодонтиды ? Бардашев, Веддидж и Зиглер, 2002 г.
- Франкодиниды ? Кабан, 2006 г.
- Гладигондолеллиды ? (Хирш, 1994) [Sephardiellinae Plasencia, Hirsch & Márquez-Aliaga, 2007 ; Neogondolellinae Hirsch, 1994 ; Сад Корнудинин , 2005 г .; Epigondolellinae Orchard, 2005 ; Marquezellinae Plasencia et al. , 2018 ; Сад Paragondolellinae , 2005 г .; Pseudofurnishiidae Ramovs, 1977 ]
- Иовагнатиды ? Лю и др. , 2017
- Новиспатодонтиды ? (Фруктовый сад, 2005)
- Трухерогнатиды ? Брэнсон и Мель, 1944 год.
- Вьяловогнатиды ? Шен, Юань и Хендерсон, 2015 г.
- Вапитиодонтиды ? Фруктовый сад, 2005 г.
- Cryptotaxidae Клэппер и Филип, 1971 г.
- Spathognathodontidae Hass, 1959 [ Ozarkodinidae Dzik, 1976 ]
- Pterospathodontidae Cooper, 1977 [Carniodontidae]
- Kockelellidae Klapper, 1981 [Caenodontontidae]
- Polygnathidae Bassler, 1925 [?Eopolygnathidae Bardachev, Weddige & Ziegler, 2002 ]
- Пальматолепиды сладкие, 1988 г.
- Хиндеодонтиды (Хасс, 1959)
- Elictognathidae Остин и Родос, 1981 г.
- Gnathodontidae Sweet, 1988 г.
- Idiognathodontidae Харрис и Холлингсворт, 1933 г.
- Mestognathidae Остин и Родос, 1981 г.
- Cavusgnathidae Остин и Родос, 1981 г.
- Sweetognathidae Риттер, 1986 г.
- Отряд Protopanderodontida Sweet, 1988 [Panderodontida Sweet, 1988 ]
- Кавидонти Сладкий, 1988 год.
См. также
[ редактировать ]- Хронология эволюционной истории жизни
- Микропалеонтология
- Список родов конодонтов
- Биостратиграфия конодонтов
- Индекс изменения конодонта
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Эйхенберг, В. (1930). «Конодонты из вершины Гарца». Палеонтологический журнал . 12 (3–4): 177–182. дои : 10.1007/BF03044446 . S2CID 129519805 .
- ^ Мило, Уолтер К.; Купер, Барри Дж. (декабрь 2008 г.). «Знакомство Ч. Пандера с конодонтами, 1856 г.» . Проверено 3 января 2019 г.
- ^ Ян Заласевич и Сара Габботт (5 июня 1999 г.). «Быстрый и мертвый» . Новый учёный .
- ^ Перейти обратно: а б Фореус, Ларс Э. (1983). «Тип Conodonta Pander, 1856 и номенклатурный приоритет» . Систематическая зоология . 32 (4): 455–459. дои : 10.2307/2413175 . JSTOR 2413175 .
- ^ Перейти обратно: а б Бенгтсон, Стефан (1976). «Структура некоторых конодонтов среднего кембрия, а также ранняя эволюция структуры и функций конодонтов» . Летайя . 9 (2): 185–206. дои : 10.1111/j.1502-3931.1976.tb00966.x . ISSN 0024-1164 .
- ^ Кларк, Дэвид Л. (1981). «Глава 3: Систематические описания» . В Муре, Раймонд К.; Робисон, Р.А. (ред.). Часть W, Разное, Приложение 2: Conodonta . Трактат по палеонтологии беспозвоночных . Боулдер, Колорадо; Лоуренс, Канзас: Геологическое общество Америки; Университет Канзаса. стр. 111–180. ISBN 0-8137-3028-7 .
- ^ Фореус, Ларс Э. (1984). «Критический взгляд на классификацию семейных групп Конодонты в Трактате: упражнение в эклектизме» . Летайя . 17 (4): 293–305. дои : 10.1111/j.1502-3931.1984.tb00675.x . ISSN 0024-1164 .
- ^ Перейти обратно: а б с Сладкий, туалет (1988). «Conodonta: морфология, таксономия, палеоэкология и история эволюции давно вымершего типа животных» . Оксфордские монографии по геологии и геофизике (10): 1–211. ISBN 978-0-19-504352-5 .
- ^ Перейти обратно: а б с д и Донохью, PCJ; Фори, Польша; Олдридж, Р.Дж. (2000). «Сродство конодонтов и филогения хордовых» . Биологические обзоры . 75 (2): 191–251. дои : 10.1111/j.1469-185X.1999.tb00045.x . ПМИД 10881388 . S2CID 22803015 .
- ^ Перейти обратно: а б с д Милый, туалет; Донохью, PCJ (2001). «Конодонты: прошлое, настоящее, будущее» (PDF) . Журнал палеонтологии . 75 (6): 1174–1184. doi : 10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 . S2CID 53395896 . Архивировано (PDF) из оригинала 30 октября 2022 г.
- ^ Перейти обратно: а б с Донохью, Филип CJ; Пурнелл, Марк А.; Олдридж, Ричард Дж.; Чжан, Шуньсинь (1 января 2008 г.). «Взаимоотношения «сложных» конодонтов (Vertebrata)» . Журнал систематической палеонтологии . 6 (2): 119–153. дои : 10.1017/S1477201907002234 . ISSN 1477-2019 .
- ^ ЧУДО. «Конодонты» . Проверено 26 августа 2014 г.
- ^ Бриггс, DEG; Кларксон, ЭНК; Олдридж, Р.Дж. (1983). «Конодонтное животное». Летайя . 16 (1): 1–14. дои : 10.1111/j.1502-3931.1983.tb01993.x .
- ^ Троттер, Джули А. (2006). «Химическая систематика апатита конодонта, определенная методом лазерной абляции ICPMS». Химическая геология . 233 (3–4): 196–216. Бибкод : 2006ЧГео.233..196Т . doi : 10.1016/j.chemgeo.2006.03.004 .
- ^ Джеппссон, Леннарт; Анехус, Рикард (1995). «Техника буферной муравьиной кислоты для извлечения конодонтов». Журнал палеонтологии . 69 (4): 790–794. дои : 10.1017/s0022336000035319 . JSTOR 1306313 . S2CID 131850219 .
- ^ Грин, Оуэн Р. (2001). «Методы экстракции фосфатных ископаемых». Руководство по практическим лабораторным и полевым методам палеобиологии . стр. 318–330. дои : 10.1007/978-94-017-0581-3_27 . ISBN 978-90-481-4013-8 .
- ^ Куинтон, Пейдж С. (2016). «Влияние протоколов экстракции на изотопный состав кислорода элементов конодонтов». Химическая геология . 431 : 36–43. Бибкод : 2016ЧГео.431...36Q . doi : 10.1016/j.chemgeo.2016.03.023 .
- ^ Бергстрем, С.М.; Карнс, Дж.Б.; Этингтон, РЛ; Вотав, РБ; Вигли, П.Б. (1974). «Appalachignathus, новый многоэлементный род конодонтов из среднего ордовика Северной Америки». Журнал палеонтологии . 48 (2): 227–235. doi : 10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 . JSTOR 1303249 . S2CID 53395896 .
- ^ Шмидт, Герман (1934). «Находки Конодонта в исходном контексте». Палеонтологический журнал . 16 (1–2): 76–85. дои : 10.1007/BF03041668 . S2CID 128496416 .
- ^ Гарольд В. Скотт, «Зоологические взаимоотношения конодонтов. Палеонтологический журнал» , том 8, № 4 (декабрь 1934 г.), страницы 448-455 ( стабильный URL-адрес )
- ^ Скотт, Гарольд В. (1942). «Комплексы конодонтов из формации Хит, Монтана». Журнал палеонтологии . 16 (3): 293–300. JSTOR 1298905 .
- ^ Данн, Дэвид Л. (1965). «Поздние конодонты Миссисипи из формации Берд-Спринг в Неваде» . Журнал палеонтологии . 39 :6. Архивировано из оригинала 18 августа 2016 г. Проверено 15 июля 2016 г.
- ^ Барнс, Кристофер Р. (1967). «Сомнительный природный комплекс конодонтов из известняка среднего ордовика, Оттава, Канада». Журнал палеонтологии . 41 (6): 1557–1560. JSTOR 1302203 .
- ^ Пурнелл, Массачусетс; Донохью, ПК (1997). «Архитектура и функциональная морфология скелетного аппарата конодонтов озаркодинид» . Философские труды Королевского общества B: Биологические науки . 352 (1361): 1545–1564. Бибкод : 1997РСТБ.352.1545П . дои : 10.1098/rstb.1997.0141 . ПМК 1692076 .
- ^ Конвей Моррис, Саймон (12 апреля 1990 г.). « Typhloesus wellsi (Melton and Scott, 1973), причудливое многоклеточное животное из каменноугольного периода Монтаны, США» . Философские труды Лондонского королевского общества. Б. Биологические науки . 327 (1242): 595–624. Бибкод : 1990РСТБ.327..595М . дои : 10.1098/rstb.1990.0102 .
- ^ Перейти обратно: а б Конвей Моррис, Саймон; Карон, Жан-Бернар (2022). «Возможный дом для странного каменноугольного животного: является ли Typhloesus пелагическим брюхоногим моллюском?» . Письма по биологии . 18 (9). дои : 10.1098/rsbl.2022.0179 . ПМЦ 9489302 . ПМИД 36126687 .
- ^ Перейти обратно: а б с Мердок, Дункан Дж. Э.; Смит, М. Пол (2021). Сансом, Роберт (ред.). «Пандеродус из Ваукеша Лагерштетте в Висконсине, США: примитивный позвоночный хищник-макрофаг» . Статьи по палеонтологии . 7 (4): 1977–1993. дои : 10.1002/spp2.1389 . ISSN 2056-2799 . S2CID 237769553 .
- ^ Перейти обратно: а б с д Габботт, SE; Р. Дж. Олдридж; Дж. Н. Терон (1995). «Гигантский конодонт с сохранившейся мышечной тканью из верхнего ордовика Южной Африки». Природа . 374 (6525): 800–803. Бибкод : 1995Natur.374..800G . дои : 10.1038/374800a0 . S2CID 4342260 .
- ^ фон Биттер, Питер Х.; Пурнелл, Марк А.; Тетро, Денис К.; Стотт, Кристофер А. (2007). «Eramosa Lagerstätte — исключительно сохранившаяся мягкотелая биота с мелководными ракушками и биотурбирующими организмами (силурийский период, Онтарио, Канада)» . Геология . 35 (10): 879. Бибкод : 2007Гео....35..879В . дои : 10.1130/g23894a.1 . ISSN 0091-7613 .
- ^ Такахаши, Сатоши; Ямакита, Сатоши; Сузуки, Норитоши (15 июня 2019 г.). «Природные комплексы конодонта Clarkina в глубоководных черных аргиллитах нижнего триаса из северо-восточной Японии с вероятными отпечатками мягких тканей» . Палеогеография, Палеоклиматология, Палеоэкология . 524 : 212–229. Бибкод : 2019PPP...524..212T . дои : 10.1016/j.palaeo.2019.03.034 . ISSN 0031-0182 . S2CID 134664744 .
- ^ Фостер, Джон (6 июня 2014 г.). Кембрийский океанский мир: древняя морская жизнь Северной Америки . Издательство Университета Индианы. стр. 300–301. ISBN 978-0-253-01188-6 .
- ^ Пурнелл, Марк А. (1 апреля 1993 г.). «Механизмы питания конодонтов и функции твердых тканей ранних позвоночных» . Геология . 21 (4): 375–377. Бибкод : 1993Geo....21..375P . doi : 10.1130/0091-7613(1993)021<0375:FMICAT>2.3.CO;2 . Проверено 15 июля 2021 г.
- ^ Перейти обратно: а б с д Террилл, Дэвид Ф.; Яроховска, Эмилия; Хендерсон, Чарльз М.; Ширли, Брайан; Бремер, Оскар (08 апреля 2022 г.). «Соотношения Sr/Ca и Ba/Ca поддерживают трофическое разделение внутри силурийского сообщества конодонтов из Готланда, Швеция» . Палеобиология . 48 (4): 601–621. дои : 10.1017/pab.2022.9 . ISSN 0094-8373 . S2CID 248062641 .
- ^ Свис, Пшемыслав (2019). «Динамика численности позднедевонского конодонта Alternognathus в днях». Историческая биология: Международный журнал палеобиологии : 1–9. дои : 10.1080/08912963.2018.1427088 . S2CID 89835464 .
- ^ Санявский, Хуберт (декабрь 2009 г.). «Самые ранние известные ядовитые животные, обнаруженные среди конодонтов» . Acta Palaeontologica Polonica . 54 (4): 669–676. дои : 10.4202/app.2009.0045 .
- ^ Бриггс, Д. (май 1992 г.). «Конодонты: основная вымершая группа, добавленная к позвоночным». Наука . 256 (5061): 1285–1286. Бибкод : 1992Sci...256.1285B . дои : 10.1126/science.1598571 . ПМИД 1598571 .
- ^ Милсом, Клэр; Ригби, Сью (2004). «Позвоночные». Окаменелости с первого взгляда . Виктория, Австралия: Blackwell Publishing. п. 88. ИСБН 978-0-632-06047-4 .
- ^ Санявский, Х. (2002). «Новые доказательства протоконодонтного происхождения щетинистых» (PDF) . Acta Palaeontologica Polonica . 47 (3): 405.
- ^ Тернер С., Берроу С.Дж., Шульце Х.П., Блик А., Рейф В.Е., Рексроуд С.Б., Бултынк П., Ноулан Г.С.; Нора; Шульце; Блик; Рейф; Рексроуд; Бултынк; Ноулан (2010). «Вставные зубы: новый взгляд на филогенетические взаимоотношения конодонтов и позвоночных» (PDF) . Геодиверситас . 32 (4): 545–594. дои : 10.5252/g2010n4a1 . S2CID 86599352 . Архивировано из оригинала (PDF) 19 марта 2012 г. Проверено 11 февраля 2011 г.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мияшита, Тецуто; Коутс, Майкл И.; Фаррар, Роберт; Ларсон, Питер; Мэннинг, Филип Л.; Вогелиус, Рой А.; Эдвардс, Николас П.; Энн, Дженнифер; Бергманн, Уве; Палмер, А. Ричард; Карри, Филип Дж. (05 февраля 2019 г.). «Мексика из мелового моря Тетис и урегулирование морфолого-молекулярного конфликта в ранней филогении позвоночных» . Труды Национальной академии наук . 116 (6): 2146–2151. Бибкод : 2019PNAS..116.2146M . дои : 10.1073/pnas.1814794116 . ISSN 0027-8424 . ПМК 6369785 . ПМИД 30670644 .
- ^ Журавлёв Андрей Владимирович; Плотицын, Артем Н. (18 января 2022 г.). «Средний-поздний турнейский кризис разнообразия конодонтов: сравнение Северо-Восточной Лавруссии и Северо-Восточной Сибири» . Палеомир . 31 (4): 633–645. дои : 10.1016/j.palwor.2022.01.001 . S2CID 246060690 . Проверено 16 октября 2022 г.
- ^ Ши, Юкун; Ван, Сяндун; Фань, Цзюньсюань; Хуан, Хао; Сюй, Хуэйцин; Чжао, Инъин; Шен, Шучжун (сентябрь 2021 г.). «Карбон — самое раннее пермское событие морского биоразнообразия (CPBE) во время позднепалеозойского ледникового периода» . Обзоры наук о Земле . 220 : 103699. Бибкод : 2021ESRv..22003699S . doi : 10.1016/j.earscirev.2021.103699 . Проверено 4 сентября 2022 г.
- ^ Сепкоски, Джей Джей (2002). «Сборник родов ископаемых морских животных». Бюллетени американской палеонтологии . 363 : 1–560.
- ^ Гино, Самуэль; Гудеманд, Николя (декабрь 2020 г.). «Глобальные изменения климата объясняют основные тенденции разнообразия конодонтов, но не их окончательную гибель» . Глобальные и планетарные изменения . 195 : 103325. Бибкод : 2020GPC...19503325G . дои : 10.1016/j.gloplacha.2020.103325 . S2CID 225005180 .
- ^ Архив филогении Микко [1] Хаарамо, Микко (2007). « Конодонта-конодонты » . Проверено 30 декабря 2015 г.
Дальнейшее чтение
[ редактировать ]- Олдридж, Р.Дж.; Бриггс, DEG; Смит, М. Пол ; Кларксон, ЭНК; Кларк, НДЛ (1993). «Анатомия конодонтов». Философские труды Лондонского королевского общества, серия B. 340 (1294): 405–421. дои : 10.1098/rstb.1993.0082 .
- Олдридж, Р.Дж.; Пурнелл, Массачусетс (1996). «Споры о конодонтах». Тенденции экологии и эволюции . 11 (11): 463–468. дои : 10.1016/0169-5347(96)10048-3 . ПМИД 21237922 .
- Донохью, PCJ; Фори, Польша; Олдридж, Р.Дж. (2000). «Сродство конодонтов и филогения хордовых». Биологические обзоры . 75 (2): 191–251. дои : 10.1111/j.1469-185X.1999.tb00045.x . ПМИД 10881388 . S2CID 22803015 .
- Гулд, Стивен Джей (1985). «Сокращающие загадки». В «Улыбке фламинго» , 245–260. Нью-Йорк, WW Norton and Company. ISBN 0-393-30375-6 .
- Жанвье, П. (1997). «Эуконодонта» . Веб-проект «Древо жизни» . Проверено 5 сентября 2007 г.
- Нелл, Саймон Дж. Великая загадка ископаемых: поиск конодонта (издательство Университета Индианы; 2012) 440 страниц
- Сладкий, Уолтер (1988). Conodonta: морфология, таксономия, палеоэкология и история эволюции давно вымершего типа животных. Оксфорд, Кларендон Пресс .
- Милый, туалет; Донохью, PCJ (2001). «Конодонты: прошлое, настоящее и будущее» (PDF) . Журнал палеонтологии . 75 (6): 1174–1184. doi : 10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 . ISSN 0022-3360 . S2CID 53395896 . Архивировано (PDF) из оригинала 30 октября 2022 г.
- Линдстрем, Мауриц (1970). «Супрародовая таксономия конодонтов». Летайя . 3 (4): 427–445. дои : 10.1111/j.1502-3931.1970.tb00834.x .
Внешние ссылки
[ редактировать ]- Марк Пернелл . «Наклонный вид спереди модели аппарата пенсильванского конодонта Idiognathodus » .
- « « Челюсти, которые ловят »: введение в конодонту» . Палеос . Проверено 1 июля 2013 г.
- Джим Дэвисон (15 октября 2002 г.). «Ордовикские конодонты» . Проверено 7 июля 2009 г.
