Энантиорниты
| Энантиорниты | |
|---|---|

| |
| Ископаемый экземпляр бохайорнитида ( Zhouornis hani ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | Авиалае |
| Клэйд : | Орнитотораки |
| Клэйд : | † Энантиорниты Уокер , 1981 год. |
| Подгруппы | |
|
и увидеть текст | |
Энантиорниты (« птиц , также известные в литературе как энантиорнитины или энантиорниты, — группа вымерших авиаланов » в широком смысле), наиболее многочисленная и разнообразная группа, известная с мезозойской эры . [3] [4] [5] Почти все сохранили зубы и когтистые пальцы на каждом крыле, но в остальном внешне очень походили на современных птиц. Было названо более восьмидесяти видов энантиорнитов, но некоторые названия представляют собой только отдельные кости, поэтому вполне вероятно, что не все из них действительны. Энантиорниты вымерли на границе мела и палеогена вместе с гесперорнитами и всеми другими нептичьими динозаврами .
Открытие и присвоение имени
[ редактировать ]Первых обнаруженных энантиорнитов ошибочно отнесли к современным группам птиц. Например, первый известный вид Enantiornithes, Gobipteryx minuta , первоначально считался палеогнатом, родственным страусам и тинаму . [6] Энантиорниты были впервые признаны как отдельная линия или «подкласс» птиц Сирилом А. Уокером в 1981 году. Уокер сделал это открытие на основе некоторых частичных останков позднего мелового периода на территории современной Аргентины , которые он отнес к новый род Enantiornis , давший название всей группе. С 1990-х годов было обнаружено гораздо больше полных экземпляров энантиорнитов, и было установлено, что несколько ранее описанных «птиц» (например, Iberomesornis , Cathayornis и Sinornis ) также были энантиорнитами.
Название «Энантиорниты» означает «противоположные птицы», от древнегреческого энантиос ( ἐνάντιος ) «противоположные» + орниты ( ὄρνιθες ) «птицы». Название было придумано Сирилом Александром Уокером в его знаковом документе, основавшем группу. [7] В своей статье Уокер объяснил, что он имел в виду под «противоположным»:
Возможно, самое фундаментальное и характерное различие между Enantiornithes и всеми другими птицами заключается в характере сочленения лопатки [...] и клювовидного отростка , где «нормальное» состояние полностью противоположное. [7]
Это относится к анатомической особенности – сочленению плечевых костей – которая имеет вогнуто-выпуклое соединение между лопаткой (лопаткой) и клювовидным отростком (основной костью плечевого пояса у позвоночных, кроме млекопитающих), что является обратным у современных птиц. В частности, у Enantiornithes лопатка в этом суставе вогнутая и тарельчатая, а клювовидный отросток выпуклый. [3] : 249–50 У современных птиц клювовидно-лопаточный сустав имеет вогнутый клювовидный отросток и выпуклую лопатку. [8] [9]
Уокер не понял причин, по которым он дал это имя в разделе этимологии своей статьи, и эта двусмысленность привела к некоторой путанице среди более поздних исследователей. Например, Алан Федучча заявил в 1996 году:
Птицы названы так потому, что среди многих отличительных особенностей имеется уникальное строение трехкостного канала и сращение плюсневых костей проксимально с дистально, в отличие от такового у современных птиц. [10]
Точка зрения Федуччиа о предплюсне-плюсневой кости (совмещенной верхней части стопы и голеностопной кости) верна, но Уокер не использовал это рассуждение в своей оригинальной статье. Уокер никогда не описывал слияние предплюсне-плюсневых костей как противоположное, а скорее как «только частичное». Кроме того, нет уверенности в том, что у энантиорнитов были трехкостные каналы, поскольку ни одна окаменелость не сохранила эту особенность. [3]
Как группу, энантиорниты в литературе часто называют «энантиорнитами». Однако ряд учёных отметили, что это неверно, поскольку следование стандартным правилам образования названий групп животных подразумевает отнесение только к подсемейству Enantiornitinae . Следуя соглашениям об именах, используемых для современных птиц, а также для вымерших групп, было отмечено, что правильный термин - «энантиорнит». [11] [12]
Происхождение и ареал
[ редактировать ]Птицы с характеристиками энантиорнитов, обитающие в Австралии , альбе маастрихте Южной и кампане Мексики Америки ( Alexornis [13] ), Монголия и западные окраины доисторической Азии предполагают распространение этой группы по всему миру или, по крайней мере, в относительно теплых регионах. [14] Энантиорниты были обнаружены на всех континентах, кроме Антарктиды . Окаменелости, относящиеся к этой группе, относятся исключительно к меловому периоду, и считается, что энантиорниты вымерли в то же время, что и их нептичьи родственники- динозавры . Самые ранние известные энантиорниты происходят из раннего мела Испании Noguerornis (например, ) и Китая (например, Protopteryx ), а самые поздние из позднего мела Северной и Южной Америки (например, Avisaurus и Enantiornis ). Широкое распространение этой группы позволяет предположить, что по крайней мере некоторые энантиорниты смогли пересечь океаны своим ходом; это первая известная линия птиц, имеющая глобальное распространение.
Описание
[ редактировать ]
Многие окаменелости энантиорнитов очень фрагментарны, а некоторые виды известны только по куску единственной кости. Почти все экземпляры, которые являются целыми, в полном сочленении и с сохранением мягких тканей, известны из Лас-Хойяс в Куэнке , Испания и группы Джехол в Ляонине ( Китай ). Необыкновенные останки Enantiornithes также сохранились в бирманских отложениях янтаря, датируемых 99 миллионами лет назад, и включают детенышей, описанных в 2017 году. [15] и 2018 год, [16] а также отдельные части тела, такие как крылья [17] [18] [19] и ноги. [18] [20] Эти янтарные останки являются одними из наиболее хорошо сохранившихся среди мезозойских динозавров. Окаменелости этой клады были обнаружены как во внутренних, так и в морских отложениях, что позволяет предположить, что они представляли собой экологически разнообразную группу.
Энантиорниты, по-видимому, включали куликов, пловцов, зерноядных, насекомоядных, рыболовов и хищников. Подавляющее большинство энантиорнитов были небольшими, размером от воробья до скворца . [21] однако у некоторых видов наблюдаются значительные различия в размерах. К самым крупным видам этой клады относятся Pengornis houi , [22] Сянгорнис Шенми , [23] Жуорнис Хани , [21] и Мирарсе Еатони , [24] (причем последний вид описан как аналогичный по размеру современным индейкам), хотя, возможно, существовало, по крайней мере, и несколько более крупных видов, в том числе вид, потенциально размером с журавля, известный только по следам в формации Эумералла (и, возможно, также представленный в формации Эумералла). Формирование Вонтхаджи из одной вилочки ). [25] Среди самых мелких описанных экземпляров — безымянные птенцы, хотя голотипические экземпляры Parvavis chuxiongensis [26] и Cratoavis cerensis [27] по размеру сравнимы с синицами или колибри.
Череп
[ редактировать ]
Учитывая широкий диапазон сред обитания и рациона питания, морфология черепа Enantiornithes значительно различалась у разных видов. Черепа энантиорнитов сочетали в себе уникальный набор примитивных и продвинутых особенностей. Как и у более примитивных птиц, таких как археоптерикс , они сохранили несколько отдельных черепных костей, небольшие предчелюстные кости (кости кончика морды), и у большинства видов были зубастые челюсти, а не беззубые клювы. Лишь несколько видов, например Gobipteryx minuta , были полностью беззубыми и имели клювы. У них также были простые квадратные кости , полная перемычка, отделяющая каждую орбиту (глазничное отверстие) от каждого предглазничного отверстия , и зубные кости (основные зубчатые кости нижней челюсти) без раздвоенных задних кончиков. Чешуйчатая кость сохранилась у неопределенного молодого экземпляра, а заглазничная кость сохранилась у Shenqiornis и Pengornis . У современных птиц эти кости ассимилированы с черепом. (отверстия в боковой части головы) могли У некоторых энантиорнитов височные отверстия сливаться с орбитами, как у современных птиц, из-за того, что заглазничные кости либо отсутствовали, либо были недостаточно длинными, чтобы разделить отверстия. [28] , сохранилась квадратноскуловая кость которая у современных птиц срослась со скуловой У Pterygornis . [29] Наличие этих примитивных особенностей черепа сделало энантиорнитов способными лишь к ограниченному краниальному кинезису (способности двигать челюстью независимо от черепа). [30]
Крыло
[ редактировать ]
Будучи очень большой группой птиц, энантиорниты демонстрировали большое разнообразие различных строений тела, основанное на различиях в экологии и питании, что отражалось в равном разнообразии форм крыльев и многих параллельных адаптациях к разному образу жизни, наблюдаемых у современных птиц. В целом, крылья энантиорнитов были развиты по сравнению с более примитивными птицами, такими как археоптерикс , и демонстрировали некоторые особенности, связанные с полетом, аналогичные тем, которые обнаружены в линии, ведущей к современным птицам, орнитуроморфам . Хотя у большинства энантиорнитов были когти по крайней мере на некоторых пальцах, у многих видов были укороченные руки, очень подвижный плечевой сустав и пропорциональные изменения в костях крыльев, как у современных птиц. Как и у современных птиц, у энантиорнитов были алулы , или «ублюдочные крылья», небольшие направленные вперед перья на первом пальце, которые обеспечивали более высокую маневренность в воздухе и помогали точно приземляться. [31]
найдено несколько крыльев с сохранившимися перьями В бирманском янтаре . Это первые полные останки мезозойских динозавров, сохранившиеся таким образом (известно несколько отдельных перьев, не отнесенных к какому-либо виду), и одна из наиболее прекрасно сохранившихся окаменелостей динозавров из известных. [32] Сохранившиеся крылья демонстрируют различия в пигменте перьев и доказывают, что у энантиорнитов были полностью современные перья, включая бородки, усики и крючки, а также современное расположение перьев крыльев, включая длинные маховые, короткие кроющие, большой алула и пуховый подшерсток. [17]
У одной окаменелости энантиорнита на ногах видны крылья, похожие на пучки перьев, похожие на археоптерикса . Перья на ногах также напоминают четырехкрылого динозавра Microraptor , однако отличаются тем, что перья короче, более неорганизованы (они не образуют четкого крыла) и простираются только до щиколотки, а не вдоль ступни. [33]
Хвост
[ редактировать ]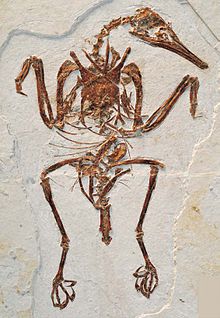
Кларк и др. (2006) исследовали все известные на тот момент окаменелости энантиорнитов и пришли к выводу, что ни у одного из них не сохранились рулевые перья, образующие веер, создающий подъемную силу, как у современных птиц. Они обнаружили, что все авиаланы за пределами Euornithes (клада, которую они называли Ornithurae ) с сохранившимися рулевыми перьями имели только короткие кроющие или удлиненные парные хвостовые перья. Они предположили, что развитие пигостиля у Enantiornithes должно было быть результатом укорочения хвоста, а не развитием современной анатомии рулевых перьев. Эти ученые предположили, что веер из рулевых перьев и связанная с ним мускулатура, необходимая для управления ими, известная как прямокишечная луковица , развилась вместе с коротким треугольным пигостилем, как у современных птиц, а не с длинным, стержнеобразным или кинжаловидным. пигостили у более примитивных авиаланов, таких как Enantiornithes. Вместо веера из перьев у большинства энантиорнитов была пара длинных специализированных перьев, похожих на таковые у вымерших Confuciusornis и некоторых дошедших до нас. райские птицы . [34]
Однако дальнейшие открытия показали, что, по крайней мере, у базальных энантиорнитов анатомия хвоста была более сложной, чем считалось ранее. Один род, Shanweiniao , первоначально интерпретировался как имеющий как минимум четыре длинных рулевых пера, которые перекрывали друг друга. [35] и, возможно, образовал поверхность, создающую подъемную силу, подобную хвостовым веерам Евронита . [36] хотя более позднее исследование показывает, что у Shanweiniao , скорее всего, были рулевые перья с преобладанием ости , похожие на перья, присутствующие у Paraprotopteryx . [37] Chiappeavis , примитивный пенгорнитид , имел веер из хвостовых перьев, похожий на веер более примитивных птиц, таких как Sapeornis , что позволяет предположить, что это могло быть предковым состоянием, при этом булавочные перья были особенностью, которая несколько раз развивалась у ранних авиаланов для демонстрационных целей. [37] Другой вид Enantiornithes, Feitianius , также имел сложный веер из рулевых перьев. Что еще более важно, мягкие ткани, сохранившиеся вокруг хвоста, были интерпретированы как остатки луковицы прямой кишки, что позволяет предположить, что эта особенность на самом деле не ограничивалась видами с современными пигостилями, но могла возникнуть гораздо раньше, чем считалось ранее, и присутствовать у многих видов. Энантиорниты. [38] По крайней мере, один род Enantiornithes, Cruralispennia , имел пигостиль современного вида, но не имел хвостового веера. [39]
Биология
[ редактировать ]Диета
[ редактировать ]
Учитывая большое разнообразие форм черепа энантиорнитов, среди этой группы должно было присутствовать множество различных диетических специализаций. У некоторых, таких как Shenqiornis , были большие, крепкие челюсти, подходящие для поедания беспозвоночных с твердым панцирем. Короткие тупые зубы Пенгорниса, вероятно, использовались для питания мягкотелыми членистоногими. [28] Сильно зацепленные когти Bohaiornithidae позволяют предположить, что они были хищниками мелких и средних позвоночных, но их крепкие зубы вместо этого позволяют предположить, что они питались животными с твердым панцирем. [2]
У некоторых экземпляров сохранилось настоящее содержимое желудка. К сожалению, ни один из них не сохранил череп, поэтому прямую корреляцию между их известным рационом питания и формой морды/зубов установить невозможно. Eoalulavis Было обнаружено, что у остатки экзоскелетов водных ракообразных . в пищеварительном тракте сохранились [40] а Энантиофеникс сохранил частицы янтаря среди окаменелых костей, что позволяет предположить, что это животное питалось соком деревьев, как современные сапсосы и другие птицы. Сок окаменел и стал янтарем. [41] Однако совсем недавно было высказано предположение, что сок переместился после смерти и, следовательно, не представляет собой истинное содержимое желудка. В сочетании с предполагаемыми рыбными гранулами Piscivorenantiornis , оказавшимися рыбьими экскрементами, странным содержимым желудка некоторых видов, оказавшимся яичниками , а предполагаемые гастролиты Bohaiornis представляют собой случайные минеральные осадки, только Eoalulavis демонстрирует реальное содержимое желудка. [42]
Исследование пищеварительной системы паравианцев показывает, что известные энантиорниты не имели зоба и желудка, не использовали гастролиты и не выбрасывали гранулы. Считается, что это противоречит большому разнообразию диет, которое подразумевают их разные формы зубов и черепа. [43] хотя некоторые современные птицы потеряли желудок и полагаются исключительно на сильные желудочные кислоты. [44] Был обнаружен пример того, что предположительно было гастролитами, в том, что могло быть желудком окаменелости, что возобновило дискуссию об использовании гастролитов энантиорнитами. Рентгеновское исследование камней и сканирующая микроскопия установили, что это на самом деле кристаллы халцедона, а не гастролиты. [45]
Longipterygidae — наиболее изученное семейство с точки зрения питания из-за довольно необычной ростральной анатомии: длинных челюстей и небольшого количества зубов, расположенных на концах челюстей. Их по-разному интерпретировали как рыбоядных животных. [46] зонды, похожие на куликов [47] и как древесные исследователи коры. [48] Однако исследование 2022 года показало, что они, скорее всего, являются универсальными насекомоядными (возможно, без Shengjingornis из-за его большего размера, плохо сохранившегося черепа и необычной анатомии педалей), поскольку они слишком малы для специализированных плотоядных и травоядных животных; Предполагается, что атипичный рострум не связан с экологией питания. [49]
Хищничество
[ редактировать ]Окаменелость из Испании, о которой сообщили Sanz et al. в 2001 году включали останки четырех вылупившихся скелетов трех разных видов энантиорнитов. Они практически цельные, очень плотно соединены между собой и имеют ямчатую поверхность костей, что указывает на частичное пищеварение. Авторы пришли к выводу, что эта ассоциация представляла собой отрыгнутую гранулу, и, судя по деталям пищеварения и размеру, вылупившиеся птенцы были проглочены целиком птерозавром или небольшим динозавром -тероподом . Это было первое свидетельство того, что мезозойские авиаланы были животными-жертвами и что некоторые мезозойские панавианы отрыгивали пометы, как это делают совы сегодня. [50]
История жизни
[ редактировать ]
Известные окаменелости энантиорнитов включают яйца , [51] [52] эмбрионы , [53] и детеныши . [54] обнаружен эмбрион, все еще свернувшийся в яйце В формации Исянь . [55] Молодые экземпляры можно идентифицировать по совокупности факторов: грубая текстура кончиков костей, указывающая на части, которые на момент смерти все еще состояли из хряща, относительно маленькие грудины, большие черепа и глаза, а также кости, которые еще не срослись друг с другом. . [56] Некоторым вылупившимся экземплярам были даны официальные названия, в том числе « Liaoxiornis delicatus »; однако Луис Кьяппе и его коллеги считали, что практика присвоения названий новым видам на основе молодых особей вредна для изучения Enantiornithes, поскольку практически невозможно определить, к какому взрослому виду принадлежит данный молодой экземпляр, что делает любой вид с голотипом вылупившихся особей nomen dubium. . [56]
Вместе с вылупившимися особями монгольского Gobipteryx. [57] и Гобипипус , [58] [59] эти находки демонстрируют, что только что вылупившиеся энантиорниты имели окостенение скелета, хорошо развитые перья крыльев и большой мозг, что коррелирует с преждевременными или сверхранними паттернами развития современных птиц. Другими словами, энантиорниты, вероятно, вылупились из яйца уже в возрасте нескольких дней, уже хорошо развитыми и готовыми бегать, добывать пищу и, возможно, даже летать. [56]
Результаты показывают, что энантиорниты, особенно зубастые виды, имели более длительный инкубационный период, чем современные птицы. [60] [61]
Для определения темпов роста этих животных был проведен анализ гистологии костей Enantiornites. Исследование костей конкорниса , проведенное в 2006 году , показало характер роста, отличный от современных птиц; хотя рост был быстрым в течение нескольких недель после вылупления, вероятно, до оперения , этот небольшой вид не достигал взрослых размеров в течение длительного времени, вероятно, нескольких лет. [62] Все другие исследования подтвердили мнение о том, что рост до взрослого размера был медленным, как и у ныне живущих скороспелых птиц (в отличие от альтрициальных птиц, которые, как известно, быстро достигают взрослых размеров). [40] Исследования скорости роста костей у различных энантиорнитов показали, что более мелкие виды имеют тенденцию расти быстрее, чем более крупные, что противоположно закономерности, наблюдаемой у более примитивных видов, таких как Jeholornis , и у нептичьих динозавров. [63] Некоторые анализы показали, что гистология костей указывает на то, что энантиорниты, возможно, не обладали полной птичьей эндотермией , а вместо этого имели промежуточную скорость метаболизма . [64] Однако исследование 2021 года отвергает идею о том, что у них был менее эндотермический метаболизм, чем у современных птиц. [65]
Свидетельства колониального гнездования были обнаружены в Энантиорните, в отложениях позднего мела ( маастрихта ) Румынии . [66] Данные с мест гнездования показывают, что энантиорниты закапывали свои яйца, как современные мегаподы , что согласуется с их предполагаемой сверхранней адаптацией. [67]
Исследование перьев молодых особей, проведенное в 2020 году, еще раз подчеркивает онтологическое сходство с современными мегаподами, но предупреждает о некоторых различиях, таких как древесный характер большинства энантиорнитов в отличие от наземного образа жизни мегапод. [68]
Было высказано предположение, что сверхскороспелость энантиорнитов могла помешать им развить особое расположение пальцев ног, наблюдаемое у современных птиц, такое как зигодактилия. [69]
Хотя подавляющее большинство гистологических исследований и известные останки энантиорнитов указывают на то, что сверхскороспелость является нормой, один экземпляр, MPCM-LH-26189, по-видимому, представляет собой альтрициальную молодь, подразумевая, что, как и современные птицы, энантиорниты использовали несколько репродуктивных стратегий. [70]
Полет
[ редактировать ]Поскольку у многих энантиорнитов не было сложных хвостов, а анатомия крыльев радикально отличалась от современных птиц, они стали предметом нескольких исследований, проверяющих их летные способности.
Традиционно их считали плохими летчиками, поскольку анатомия плечевого пояса считалась более примитивной и неспособной поддерживать наземный пусковой механизм. [71] а также из-за отсутствия прямоугольников у многих видов. [34] [36] [72]
Однако несколько исследований показали, что они были эффективными летчиками, как и современные птицы, обладая такой же сложной нервной системой и связками перьев крыльев. Кроме того, отсутствие сложного хвоста, по-видимому, не имело большого значения для полета птиц в целом - у некоторых вымерших птиц, таких как литорниды, также не было сложных рулевых перьев, но они были хорошими летчиками. [73] и, похоже, они были способны к наземному запуску. [74]
Энантиорниты по многим анатомическим особенностям летательного аппарата напоминают орнитуроморфов , но у самых базальных представителей отсутствует грудинный киль, только один базальный таксон, по-видимому, имел трехкостный канал, а их мощный пигостиль, по-видимому, не способен поддерживать мышцы, контролирующие современные рулевые перья, участвующие в полете. [75] Хотя некоторые базальные энантиорниты обладают наследственными летательными аппаратами, к концу мезозоя многие энантиорниты имели несколько особенностей, сходящихся с неорнитами, включая глубоко килеватую грудину, узкую вилку с коротким гипоклеидием и локтевые выступы перьев, которые указывают на повышенные воздушные способности. [76] [77]
По крайней мере, Эльсорнис, похоже, стал вторично нелетающим . [78]
Классификация
[ редактировать ]Некоторые исследователи относят энантиорнитов, наряду с настоящими птицами, к классу Aves . Другие используют более строгое определение группы кроны Aves (которая включает только неорнитов , анатомически современных птиц) и помещают Enantiornites в более инклюзивную группу Avialae . Энантиорниты были более продвинутыми, чем археоптерикс , конфуциусорнис и сапеорнис , но в некоторых отношениях они были более примитивными, чем современные птицы, возможно, следовали промежуточным эволюционным путем.
Консенсус научных анализов показывает, что Enantiornites является одной из двух основных групп внутри более крупной группы Ornithothoraces . Другая группа орнитоторацинов — Euornithes или Ornithuromorpha , которая включает в себя всех ныне живущих птиц как подмножество. Это означает, что энантиорниты были успешной ветвью эволюции птиц, но она развивалась совершенно отдельно от линии, ведущей к современным птицам. [3] Однако одно исследование показало, что общая анатомия грудины была приобретена независимо, и такую взаимосвязь необходимо пересмотреть. [79]
Классификация и таксономия энантиорнитов исторически осложнялись рядом факторов. В 2010 году палеонтологи Джингмай О'Коннор и Гарет Дайк высказали ряд критических замечаний в адрес преобладающей практики, когда ученые не могут описать многие образцы достаточно подробно, чтобы другие могли их тщательно оценить. Некоторые виды были описаны на основе образцов, хранящихся в частных коллекциях, что делает невозможным дальнейшее изучение или анализ предыдущих результатов. Поскольку другим ученым зачастую невозможно лично изучить каждый экземпляр, учитывая всемирное распространение энантиорнитов, а также из-за множества неинформативных описаний, которые были опубликованы для возможно важных образцов, многие из этих образцов становятся «функциональным nomina dubia ». [80] Более того, многие виды были названы на основе крайне фрагментарных экземпляров, которые не имели бы большой научной информативности, даже если бы они были достаточно описаны. Более трети всех названных видов основаны только на фрагменте одной кости. О'Коннор и Дайк утверждали, что, хотя эти образцы могут помочь расширить знания о временном интервале или географическом ареале Энантиорнитов и важно их описать, присвоение названий таким образцам «неоправданно». [80]
Отношения
[ редактировать ]Энантиорниты — сестринская группа Эуорнитов , и вместе они образуют кладу под названием Орнитоторакс (см. выше). Большинство филогенетических исследований выявили энантиорниты как монофилетическую группу, отличную от современных птиц и их ближайших родственников. Однако филогенетический анализ 2002 года, проведенный Кларком и Нореллом, сократил количество аутапоморфий Enantiornites до четырех. [81]
Систематика энантиорнитов весьма условна и, как известно, трудна для изучения из-за их небольшого размера. [27] и тот факт, что энантиорниты имеют тенденцию быть чрезвычайно гомопластичными или очень похожими друг на друга по большинству особенностей скелета из-за конвергентной эволюции, а не общего происхождения. [37] К настоящему времени кажется вполне очевидным, что внутри энантиорнитов существовали подразделения, которые, возможно, включали некоторые второстепенные базальные линии в дополнение к более развитым эуэнантиорнитам. Детали взаимосвязи всех этих линий, а также достоверность большинства из них оспариваются, хотя, например, Avisauridae, похоже, составляют действительную группу. Филогенетические систематики до сих пор очень неохотно предлагали разграничение клад Enantiornithes. [82]
Одно из таких обозначений, названное Euenantiornithes , было определено Чиаппе (2002) как включающее все виды, более близкие к Sinornis, чем к Iberomesornis . Поскольку Iberomesornis часто оказывается самым примитивным или базальным представителем Enantiornites, Euenantiornites может быть чрезвычайно инклюзивной группой, состоящей из всех Enantiornites, за исключением Iberomesornis самого соответствует филогенетической номенклатуре . Несмотря на то, что это определение Euenantiornithes , оно подверглось резкой критике со стороны некоторых исследователей, таких как Пол Серено , который назвал его «плохо определенной кладой [...] хорошим примером плохого выбора в филогенетическом определении». . [82]
была Приведенная ниже кладограмма получена в результате анализа Wang et al. в 2015 году обновлено на основе предыдущего набора данных, созданного Цзинмаем О'Коннором. [29]
| Орнитотораки |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





Кладограмма ниже взята из Wang et al. , 2022 г., включает большинство названных таксонов и обнаруживает несколько ранее названных клад. Буквы на ветвях указывают позиции таксонов с «подстановочными знаками», тех, которые были обнаружены в нескольких несопоставимых позициях. [83]
| Энантиорниты | |
| л |
Ключ к буквам:
б = Болуохия
c = Катайорнис
е = Энантиофеникс
f = Хуорнис
h = Лонгиптерикс
я = Парабохайорнис
j = Птеригорнис
л = Птица
м = Юаньцзяваорнис
n = Юнгаволукрис
Список родов
[ редактировать ]Таксономию Enantiornites трудно оценить, и в результате филогенетический анализ последовательно обнаруживает несколько клад внутри группы. Большинство энантиорнитов не включены ни в одно конкретное семейство и поэтому перечислены здесь. Многие из них считались эуэнантиорнитами, хотя разногласия, связанные с этим названием, означают, что оно не используется последовательно в исследованиях энантиорнитов. [ нужна ссылка ]
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Абаворнис | 1998 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известных только по коракоидам. | ||
| Алетоалаорнис | 2007 | Формация Цзюфотанг ( ранний мел , апт ) | Плохо известен | ||
| Алексорнис | 1976 | Формация Ла Бокана Роха ( поздний мел , кампан ) | Один из первых известных энантиорнитов. Когда-то считался древним родственником валунов и дятлов. | 
| |
| Авимайя | 2019 | Формация Сягоу ( ранний мел , апт ) | Один экземпляр этого рода умер с неотложенным яйцом в теле. | 
| |
| Боксит | 2010 | Формация Чебанья ( поздний мел , сантон ) | Фрагментарный, но уникальный по строению предплюсне-плюсневый сустав. | 
| |
| Бревироструавис | 2021 | Формация Цзюфотанг ( ранний мел , апт ) | Обладал увеличенной подъязычной мышцей , что предполагает специализацию в питании, подобную колибри, медоедам и дятлам. | 
| |
| Кастигноволукрис | 2023 | Безымянное образование ( поздний мел , кампан ) | Возможно, был размером с канадского гуся. | ||
| Катенолеймус | 1998 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известных только по клювовидному отростку. | ||
| Катайорнис | 1992 | Формация Цзюфотанг ( ранний мел , апт ) | Одна из первых описанных энантиорнитов биоты Джехол. Известен от многих видов, хотя некоторые теперь отнесены к отдельным родам. Возможно, имел внешний вид и образ жизни, похожий на питту. | 
| |
| Рога | 1992 | Лас-Хойас ( ранний мел , баррем ) | Один из наиболее полных энантиорнитов Лас-Хойас. | 
| |
| Кратоавис [84] | 2015 | Формация Сантана ( ранний мел , апт ) | Очень хорошо сохранившийся южноамериканский представитель группы с лентообразными рулевыми перьями. | ||
| Круралиспенния [39] | 2017 | Формация Хуацзиин ( ранний мел , готерив ) | Имел необычный пигостиль, похожий на орнитуроморфа, и перья на бедрах, похожие на кисть. Один из древнейших энантиорнитов. | 
| |
| Куспиростризорнис | 1997 | Формация Цзюфотанг ( ранний мел , апт ) | Первоначально ошибочно считалось, что у него был заостренный клюв. | 
| |
| Далингхеорнис | 2006 | Исяньская формация ( ранний мел , апт ) | Был хорошо приспособлен к лазанию благодаря разнодактильным лапам , как у трогона. | ||
| Дуньхуанджа [85] | 2015 | Формация Сягоу ( ранний мел , апт ) | Род Enantiornithes из бассейна Чангма, территории, где необычно доминируют орнитуроморфы. | ||
| Башня Эльбре | 2009 | Формация Лечо ( поздний мел , маастрихт ) | Известен только по костям крыльев. Может быть синонимом другой формации Лечо, Энантиорнитов. | ||
| Электорорнис | 2019 | Бирманский янтарь ( поздний мел , сеноман ) | Известен по сохранившейся в янтаре ступне с удлиненным средним пальцем. | ||
| Энантиорнис | 1981 | Формация Лечо ( поздний мел , маастрихт ) | Хотя этот род известен лишь по нескольким костям, он является тезкой Enantiornites. Это также был один из крупнейших и последних представителей группы до их исчезновения. | ||
| Эоалулавис | 1996 | Лас-Хойас ( ранний мел , баррем ) | Сохраняет перья, включая алулу , особый тип перьев, который контролирует поток воздуха над крылом. | ||
| Эокатайорнис | 2002 | Формация Цзюфотанг ( ранний мел , апт ) | Когда-то считался основным близким родственником Катайорниса , хотя теперь считается более дальним родственником. | ||
| Ээнантиорнис | 1999 | Исяньская формация ( ранний мел , апт ) | Хорошо сохранились, но непоследовательны в филогенетическом размещении. | 
| |
| Евгенавис | 2014 | Илекская свита ( ранний мел , баррем ) | Известен только по предплюсне, которая имеет некоторые общие черты с энантиорнитами. | ||
| Исследовать | 1998 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известных только по коракоидам. | ||
| Фалькатакели | 2020 | Формация Маеварано ( поздний мел , маастрихт ) | Развил массивную морду только с одним зубом, несмотря на сохранение «примитивного» строения черепа в отличие от современных птиц. | 
| |
| Феицианиус [38] | 2015 | Формация Сягоу ( ранний мел , апт ) | Обладал сложным набором рулевых перьев, в отличие от парных лентообразных перьев большинства энантиорнитов. | 
| |
| Флексоморнис | 2010 | Формация Вудбайн ( поздний мел , сеноман ) | Найден один из старейших североамериканских авиаланов, хотя он известен только по фрагментарным останкам. | ||
| Ты был сильным | 2021 | Бирманский янтарь ( поздний мел , сеноман ) | Имел увеличенный внешний палец ноги, который, возможно, был приспособлением для сидения на насесте. | ||
| Фортунгуавис [86] | 2014 | Формация Цзюфотанг ( ранний мел , апт ) | Имел крепкие кости, включая ступни и когти, которые, возможно, были приспособлены для лазания по деревьям. | ||
| Грабауорнис [87] | 2015 | Формация Исянь ( ранний мел , баррем ) | Пропорции крыльев этого рода Enantiornithes, а также наличие алулы позволяют предположить, что это был хороший летчик. | ||
| Грацилорнис | 2011 | Формация Цзюфотанг ( ранний мел , апт ) | Возможный родственник катайорниса с характерно тонкими костями. | ||
| Гурилиния | 1999 | Немегтская свита ( поздний мел , маастрихт ) | Малоизвестный род Enantiornites, но, очевидно, крупный и поздно выживший член группы. | ||
| Голландия [88] | 2010 | Формация Барун-Гойот ( поздний мел , кампан ) | Первоначально идентифицирован как орнитуроморф, но с тех пор переосмыслен как род Enantiornithes, тесно связанный с Lectavis . [89] | 
| |
| Голбоция [90] | 2015 | Андайхудагская свита ( ранний мел , апт ) | Считался небольшим птерозавром с момента его открытия в 1977 году и до получения официального описания в 2015 году. Обладал уникальными шейными позвонками и примитивным небом. | ||
| Уорнис | 1997 | Формация Цзюфотанг ( ранний мел , апт ) | Когда-то считался сомнительным видом Cathayornis , хотя исследование 2015 года показало, что это действительный род. [91] | ||
| Хуошанорнис | 2010 | Формация Цзюфотанг ( ранний мел , апт ) | Возможно, он был очень маневренным летчиком из-за строения руки и грудины. | ||
| Иберомесорнис | 1992 | Лас-Хойас ( ранний мел , баррем ) | Один из первых родов Enantiornites, известных по достойным остаткам. Также один из старейших и наиболее примитивных представителей группы. | 
| |
| Импаравис [92] | 2024 | Формация Цзюфотанг ( ранний мел , апт ) | Самый ранний известный энантиорнитин с беззубым клювом. | ||
| Инколорнис | 1998 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных Bissekty Enantiornithes, известных только по коракоидам . Когда-то считалось, что один вид принадлежит к Enantiornis. | ||
| Юнорнис [93] | 2017 | Исяньская формация ( ранний мел , апт ) | Настолько хорошо сохранившийся, что схему его полета можно было реконструировать по пропорциям перьев и крыльев. | 
| |
| Кызылкумавис | 1984 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известный только по плечевой кости фрагменту . | 
| |
| Большеротый | 1997 | Формация Цзюфотанг ( ранний мел , апт ) | Возможно, связано с Cuspirostrisornis или синонимом Cathayornis. | ||
| Лектавис | 1993 | Формация Лечо ( поздний мел , маастрихт ) | Крупный и длинноногий представитель группы, пропорционально похожий на современных куликов. |  | |
| Ланесорнис | 1996 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известных только по фрагменту синсакрума . Первоначально считалось, что он принадлежал Ихтиорнису. | ||
| Лиаонингорнис | 1996 | Исяньская формация ( ранний мел , апт ) | Первоначально считалось, что это орнитуран, но теперь считается родственником Эоалулависа. | ||
| Лонгченгорнис | 1997 | Формация Цзюфотанг ( ранний мел , апт ) | Возможно, это был синоним Катайорниса. | ||
| Мартинавис | 2007 | Песчаник в формации рептилий , формация Лечо ( поздний мел , маастрихт ) | Хотя этот род известен только из плечевых суставов, он был большим и обитал в широком ареале. | ||
| Микроэнантиорнис | 2017 | Формация Цзюфотанг ( ранний мел , апт ) | Небольшой представитель группы, обладающий некоторыми примитивными и производными чертами по сравнению с другими энантиорнитами. | ||
| Мирусавис | 2020 | Исяньская формация ( ранний мел , от баррема до апта ) | Голотип представлял собой небольшую остеологически незрелую самку с сохранившейся медуллярной костной тканью. | ||
| Моноэнантиорнис [94] | 2016 | Исяньская формация ( ранний мел , апт ) | Известен по молодому экземпляру, который показывает, как различные особенности развивались у энантиорнитов с возрастом. | ||
| Мусивавис | 2022 | Формация Цзюфотанг ( ранний мел , апт ) | Наиболее сходен с бохайорнитидами, но имеет черты других групп энантиорнитов. | ||
| Нананций | 1986 | Формация Тулебук ( ранний мел , альб ) | Фрагментарный, но, возможно, это была морская птица, поскольку останки этого рода были обнаружены в ихтиозавра. содержимом кишечника | ||
| Ногерорнис | 1989 | Эль Монсек ( ранний мел , баррем ) | Сохраняет отпечатки пропатагия — кожного лоскута на плече, который является частью крыла. | ||
| Восточный | 2018 | Формация Хуацзиин ( ранний мел , готерив ) | Многие детали мягких тканей экземпляров этого рода были обнаружены с помощью ультрафиолетового света. | ||
| Отогорнис | 1993 | Формация Иджинхолуо ( ранний мел ) | Плохо известен | ||
| Парапротоптерикс | 2007 | Пачка Цяотоу формации Хуацзиин ( ранний мел , апт ?) | По-видимому, имел четыре лентообразных рулевых пера вместо двух, как у большинства энантиорнитов. | ||
| Парвавис [95] | 2014 | Формация Цзяндихэ ( поздний мел , турон - сантон ) | Маленький, но полностью зрелый на момент смерти. Один из немногих китайских энантиорнитов, датируемых поздним меловым периодом. | 
| |
| Piscivorenantiornis [96] | 2017 | Формация Цзюфотанг ( ранний мел , апт ) | Известен по разъединенному скелету, сохранившемуся над куском содержимого желудка, состоящим из рыбьих костей, которое, возможно, было его последней пищей. | ||
| Протоптерикс | 2000 | Формация Хуацзиин ( ранний мел , готерив ) | Один из старейших и наиболее примитивных представителей группы. | 
| |
| Птеригорнис [29] | 2016 | Формация Цзюфотанг ( ранний мел , апт ) | Один разрозненный скелет этого рода обладает хорошо сохранившимися костями черепа, в том числе квадратно-скуловой костью. | ||
| Килиания | 2011 | Формация Сягоу ( ранний мел , апт ) | Некоторые из останков этого рода включают хорошо сохранившиеся задние конечности. название вида, Q. graffini , названо в честь Грега Граффина из группы Bad Religion. | ||
| Сазавис | 1989 | Биссектинская свита ( поздний мел , турон - коньяк ) | Один из многих фрагментарных энантиорнитов Bissekty, известных только по большеберцовой кости (большеберцовой кости). | 
| |
| Шанъян | 2019 | Формация Цзюфотанг ( ранний мел , апт ) | Необычно то, что предчелюстные кости этого рода были слиты. | ||
| Грешники | 1992 | Формация Цзюфотанг ( ранний мел , апт ) | Одна из первых описанных энантиорнитов биоты Джехол. Похож на Cathayornis , но обычно считается отличным от него. | 
| |
| Ксянгорнис | 2012 | Формация Цзюфотанг ( ранний мел , апт ) | Рука этого рода была похожа на руку орнитуроморфов, вероятно, в результате конвергентной эволюции . Крупный член группы | ||
| Ятенавис [97] | 2022 | Формация Чоррилло ( поздний мел , маастрихт ) | Самый южный известный член Энантиорнита и один из самых молодых членов группы. | ||
| Юаньцзяваорнис [98] | 2015 | Формация Цзюфотанг ( ранний мел , апт ) | Один из крупнейших энантиорнитов, известный по приличным останкам. | ||
| Юнгаволукрис | 1993 | Формация Лечо ( поздний мел , маастрихт ) | Имел большую и необычно широкую предплюсне-плюсневую кость (лодыжку). | 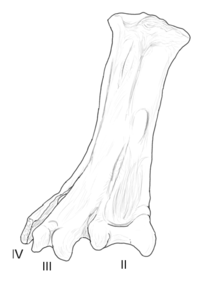
| |
| Юорнис | 2021 | Формация Киупа ( поздний мел , маастрихт ) | Крупный беззубый род Enantiornithes, один из наиболее хорошо сохранившихся представителей позднего мела. | 
|
Лонгиптеригиды
[ редактировать ]Longipterygidae — семейство длиннорылых энантиорнитов раннего мелового периода с зубами только на кончиках морды. Обычно они считаются довольно базальными членами группы. [35]
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Болуохия | 1995 | Формация Цзюфотанг ( ранний мел , апт ) | Первоначально ошибочно полагали, что у него был крючковатый клюв. | ||
| Камптодонторнис | 2010 | Формация Цзюфотанг ( ранний мел , апт ) | Первоначально назывался Camptodontus , хотя это название рода занимает жук . | ||
| Дапинфангорнис | 2006 | Формация Цзюфотанг ( ранний мел , апт ) | Возможно, на лбу у него было шипоподобное образование. | ||
| Лонгиптерикс | 2001 | Формация Цзюфотанг ( ранний мел , апт ) | Самый распространенный и известный представитель семейства. | 
| |
| Лонгиростравис | 2004 | Исяньская формация ( ранний мел , апт ) | Как и другие лонгиптеригиды, он обладал тонкой мордой, которую, возможно, использовали для поиска беспозвоночных в грязи или коре. | 
| |
| Рапаксавис | 2009 | Формация Цзюфотанг ( ранний мел , апт ) | Специализируется на сидении благодаря строению ног. | 
| |
| Шанвэйняо | 2009 | Исяньская формация ( ранний мел , апт ) | Приобретено несколько рулевых перьев, которые, возможно, способны создавать подъемную силу, как у современных птиц. | 
| |
| Шэнджингорнис | 2012 | Формация Цзюфотанг ( ранний мел , апт ) | Крупный член семьи |
Пенгорнитиды
[ редактировать ]Pengornithidae . — семейство крупных ранних энантиорнитов У них было множество мелких зубов и множество примитивных черт, которые потеряны у большинства других энантиорнитов. [1] В основном известен из раннего мела Китая, с предполагаемыми таксонами позднего мела с Мадагаскара. [99]
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Кьяппеавис [37] | 2015 | Формация Цзюфотанг ( ранний мел , апт ) | Обладал веерообразным хвостом, состоящим из множества перьев. | 
| |
| Эопенгорнис | 2014 | Формация Хуацзиин ( ранний мел , готерив ) | Самый старый известный член семейства и один из старейших известных известных предполагаемых родов Enantiornithes. Имелись очень хорошо сохранившиеся хвостовые ленты. | ||
| Парапенгорнис [100] | 2015 | Формация Цзюфотанг ( ранний мел , апт ) | Предложено вести образ жизни, подобный дятлу, из-за особенностей стопы и хвоста. | 
| |
| Гарнишер | 2008 | Формация Цзюфотанг ( ранний мел , апт ) | Первый обнаруженный пенгорнитид, а также один из крупнейших представителей энантиорнитов, известных по приличным останкам. | ||
| Юаньчуавис | 2021 | Формация Цзюфотанг ( ранний мел , апт ) | Имеет сложный хвостовой веер, длиннее его тела, который, возможно, имел функцию дисплея. | 
|
Бохайорнитиды
[ редактировать ]Бохайорнитиды были крупными, но геологически недолговечными ранними энантиорнитами, с длинными крючковатыми когтями и крепкими зубами с изогнутыми кончиками. Возможно, они были эквивалентны хищным птицам, хотя эта интерпретация вызывает много споров. [2] Монофилия эволюционная этой группы сомнительна, и на самом деле это может быть ступень . [101]
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Бейгуорнис | 2022 | Формация Лунцзян ( ранний мел , апт ) | Имел короткое, но надежное руководство по нагвалению. | ||
| Бохайорнис | 2011 | Формация Цзюфотанг ( ранний мел , апт ) | Первоначально считалось, что они сохранились с гастролитами , хотя позже было обнаружено, что это минеральные конкременты. | 
| |
| Гретчениао | 2019 | Формация Исянь ( ранний мел , баррем ) | Приспособлен для взмахивания крыльями, а не для парящего полета. Его описатели предполагают парафилию или полифилию Bohaiornithidae. | 
| |
| Линьорнис [102] | 2016 | Формация Цзюфотанг ( ранний мел , апт ) | Возможный член семейства, известный по хорошо сохранившемуся скелету со структурами, предположительно развивающими яйца. | ||
| Длинноязычный | 2014 | Формация Цзюфотанг ( ранний мел , апт ) | Довольно типичный представитель семейства | 
| |
| Парабохайорнис | 2014 | Формация Цзюфотанг ( ранний мел , апт ) | Близкий родственник Бохайорниса. | ||
| Шенькёрнис | 2010 | Пачка Цяотоу формации Хуацзиин ( ранний мел , апт ?) | Первый известный член семьи, хотя близким родственником Бохайорниса считался лишь несколько лет спустя. Сохраняет большую заглазничную кость. | ||
| Ты бороздишь | 2013 | Исяньская формация ( ранний мел , апт ) | Близкий родственник Shenqiornis с рифленой эмалью на зубах, уникальный среди ископаемых птиц. | 
| |
| Жуорнис | 2013 | Формация Цзюфотанг ( ранний мел , апт ) | Крупный член семейства с хорошо сохранившейся черепной коробкой. | 
|
Гобиптеригиды
[ редактировать ]Некоторые представители группы малоизвестны или плохо описаны и могут быть синонимами типового вида Gobipteryx minuta .
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Гобиптерикс | 1974 | Формация Барун-Гойот ( поздний мел , кампан ) | Беззубый продвинутый род Enantiornites, обладающий крепким клювом, который эволюционировал конвергентно с клювом современных птиц. | ||
| Джибейния | 1997 | Пачка Цяотоу формации Хуацзиин ( ранний мел , апт ?) | Плохо известен и описан по скелету, который сейчас утерян. Возможно, было синонимом Вескорниса. | ||
| Вескорнис | 2004 | Пачка Цяотоу формации Хуацзиин ( ранний мел , апт ?) | Небольшой и короткорылый род Enantiornithes, который может быть синонимом Jibeinia. |
Авизавриды
[ редактировать ]Avisauridae подпадает под два разных определения разной степени инклюзивности. Здесь используется более широкое определение, которое следует из Cau & Arduini (2008). Авизавриды представляли собой древнее и широко распространенное семейство энантиорнитов, которые в основном отличались особенностями предплюсне-плюсневых костей (костей лодыжки). Самые крупные и развитые представители группы сохранились в Северной и Южной Америке вплоть до конца мелового периода, однако они очень фрагментарны по сравнению с некоторыми более ранними таксонами.
| Имя | Год | Формирование | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Авизавр | 1985 | Формация Хелл-Крик ( поздний мел , маастрихт ) | Одноименный авизаврид, а также один из крупнейших представителей семейства. Первоначально считался нептичьим динозавром. | 
| |
| Эльсорнис | 2007 | Формация Джадохта ( поздний мел , кампан ) | Хотя его скелет и неполный, он сохранился в трехмерном виде. Возможно, нелетающий из-за пропорций крыльев. | ||
| Энантиофеникс | 2008 | Формация Уади-аль-Габур ( поздний мел , сеноман ) | Когда-то считалось, что он питался древесным соком, поскольку он сохранялся вместе с янтарными бусинами, однако позже это было определено как артефакт сохранности, а не показатель диеты. | ||
| Геттия | 2018 | Две формации медицины ( поздний мел , кампан ) | Новый род во славу Авизавра | ||
| Галиморнис | 2002 | Меловая формация Мурвилля ( поздний мел , кампан ) | Жил бы в прибрежной среде | ||
| Интиорнис | 2010 | Формация Лас Куртьембрес ( поздний мел , кампан ) | Хотя представители этого рода были тесно связаны с некоторыми из крупнейших авизаврид, они были очень маленькими птицами. | 
| |
| Мирарсе | 2018 | Кайпаровицкая свита ( поздний мел , кампан ) | Самый полный известный североамериканский авизаврид. | 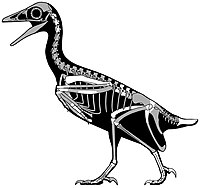
| |
| Мистиорнис | 2011 | Илекская свита ( ранний мел , баррем - апт ) | Обладает множеством черт различных групп Паравеса , хотя наиболее близко напоминает авизаврид среди отобранных групп. | ||
| Неукенорнис | 1994 | Формация Бахо-де-ла-Карпа ( поздний мел , сантон ) | Обладал длинными крыльями и обратным большим пальцем, что указывает на хорошие способности летать и садиться. | ||
| Сороавизавр | 1993 | Формация Лечо ( поздний мел , маастрихт ) | Очень близкий родственник Авизавра. |
Сомнительные роды и примечательные безымянные экземпляры.
[ редактировать ]- Gobipipus reshetovi : описан в 2013 году по образцам эмбрионов в яичной скорлупе из формации Барун Гойот в Монголии . Эти экземпляры были очень похожи на эмбриональные экземпляры Gobipteryx , хотя описатели Gobipipus (группа палеонтологов, вызывающих споры, включая Евгения Курочкина и Санкара Чаттерджи ) считают их отличными. [103]
- Hebeiornis fengningensis : синоним Vescornis , поскольку он был описан по одному и тому же экземпляру. Несмотря на то, что оно было описано в 1999 году, за 5 лет до описания Вескорниса , оно было настолько плохим по сравнению с описанием Вескорниса , что большинство авторов считают последнее название приоритетным. В результате имя Hebeiornis считается nomen nudum («обнаженное имя»).
- « Проорнис » — птица с неофициальным названием из Северной Кореи. Возможно, он не принадлежит к Энантиорнитам.
- Liaoxiornis delicatus : Описан в 1999 году по экземпляру Enantiornithes, найденному в формации Исянь . Первоначально этот экземпляр считался крошечной взрослой особью, но позже выяснилось, что это птенец. Отныне к этому роду были отнесены и другие экземпляры. Из-за отсутствия отличительных признаков многие палеонтологи считали этот род недиагностическим nomen dubium .
- «Васаибпанчи»: предполагаемый член Энантиорнита из Пакистана; описывающая статья имеет сомнительный статус.
- LP-4450 : Молодь неопределенного экземпляра Enantiornithes из формации Эль-Монсек в Испании . В его описании 2006 года изучалась гистология скелета, а в более поздних исследованиях сообщалось о наличии чешуйчатой кости в образце, но неизвестной у других энантиорнитов.
- IVPP V 13939 : Кратко описанный в 2004 году, этот экземпляр Yixian Enantiornites имел развитые перистые перья на ногах, похожие на (хотя и короче) перья других паравианов, таких как Microraptor и Anchiornis . [33]
- DIP-V-15100 и DIP-V-15101 : два разных крыла вылупившихся особей, описанных в 2015 году. После своего описания они привлекли значительное внимание средств массовой информации. Они сохранились в исключительных деталях, поскольку находились в ловушке в бирманском янтаре примерно 99 миллионов лет. [17]
- HPG-15-1 : частичный труп детеныша энантиорнита, также сохранившийся в бирманском янтаре. Несмотря на неопределенность, он привлек еще больше внимания средств массовой информации, чем два крыла после его описания в 2017 году. [15]
- CUGB P1202 : Неопределенный молодой бохайорнитид из формации Цзюфотанг . Анализ его оперения, проведенный в 2016 году, выявил предполагаемые удлиненные меланосомы , что позволяет предположить, что большая часть его оперения была радужной .
- DIP-V-15102 : Еще один труп неопределенного детеныша, сохранившийся в бирманском янтаре. Описан в начале 2018 года. [16]
- MPCM-LH-26189 a/b : Частичный скелет детеныша из Лас-Хойяс в Испании, включая компоненты плиты и контрплиты. Его описание в 2018 году показало, как различные особенности развивались у Энантиорнитов по мере их старения. К таким особенностям относятся оссификация грудины из различных более мелких костей и слияние хвостовых позвонков в пигостиль .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Ван, X.; О'Коннор, Дж. К.; Чжэн, X.; Ван, М.; Хм.; Чжоу, З. (2014). «Понимание эволюции хвостовых перьев доминировало на основе нового базального энантиорнитина (Aves: Ornithothoraces)» . Биологический журнал Линнеевского общества . 113 (3): 805–819. дои : 10.1111/bij.12313 .
- ^ Перейти обратно: а б с Ван, Мин; Чжоу, Чжун-Хе; О'Коннор, Цзинмай К.; Зеленков, Никита В. (2014). «Новое разнообразное семейство энантиорнитинов (Bohaiornithidae fam. nov.) из нижнего мела Китая с информацией о двух новых видах» (PDF) . Позвоночные Палазиатские . 52 (1): 31–76.
- ^ Перейти обратно: а б с д Кьяппе, Луис М.; Уокер, Сирил А. (2002). «Морфология скелета и систематика меловых эуэнантиорнитов (Ornithothoraces: Enantiornithes)» . В Кьяппе, Луис М.; Уитмер, Лоуренс М. (ред.). Мезозойские птицы: над головами динозавров . Издательство Калифорнийского университета. стр. 240–67. ISBN 978-0-520-20094-4 .
- ^ Кьяппе, Луис М. (2007). Прославленные динозавры: происхождение и ранняя эволюция птиц . Хобокен, Нью-Джерси: Джон Уайли и сыновья. ISBN 978-0-471-24723-4 . [ нужна страница ]
- ^ О'Коннор, Цзинмай К.; Кьяппе, Луис М.; Гао, Чуньлин; Чжао, Бо (сентябрь 2011 г.). «Анатомия раннемеловой энантиорнитиновой птицы Rapaxavis pani» . Acta Palaeontologica Polonica . 56 (3): 463–475. дои : 10.4202/app.2010.0047 . S2CID 55311115 .
- ^ Эльжановский, Анджей (1974). «Предварительная заметка о палеогнатной птице верхнего мела Монголии» (PDF) . Палеонтология Полоника . 29 : 103–9. S2CID 53487062 .
- ^ Перейти обратно: а б Уокер, Калифорния (1981). «Новый подкласс птиц мела Южной Америки». Природа . 292 (5818): 51–3. Бибкод : 1981Natur.292...51W . дои : 10.1038/292051a0 . S2CID 4340858 .
- ^ Надежда, Сильвия (2002). «Мезозойская радиация неорнитов». В Кьяппе, Луис М.; Уитмер, Лоуренс М. (ред.). Мезозойские птицы: над головами динозавров . Издательство Калифорнийского университета. стр. 339–88. ISBN 978-0-520-20094-4 .
- ^ Майр, Г. (2021). «Ключично-лопаточный сустав неорнитиновых птиц - обширная гомоплазия широко игнорируемой суставной поверхности грудного пояса птиц и ее возможные функциональные корреляты» . Зооморфология . 140 (2): 217–28. дои : 10.1007/s00435-021-00528-2 . S2CID 236397491 .
- ^ Федучча, Алан (1996). Происхождение и эволюция птиц . Нью-Хейвен: Издательство Йельского университета. ISBN 978-0-300-06460-5 . [ нужна страница ]
- ^ Харрис, Джеральд Д.; Ламанна, Мэтью С.; Ты, Хай-лу; Цзи, Шу-ан; Цзи, Цян (18 мая 2006 г.). «Второе крыло энантиорнита (Aves: Ornithothoraces) из раннемеловой формации Сягоу недалеко от Чангмы, провинция Ганьсу, Китайская Народная Республика». Канадский журнал наук о Земле . 43 (5): 547–554. Бибкод : 2006CaJES..43..547H . дои : 10.1139/e06-007 .
- ^ Ты, Хай-лу; Ламанна, Мэтью С.; Харрис, Джеральд Д.; Кьяппе, Луис М.; О'Коннор, Цзинмай; Цзи, Шу-ан; Лю, Цзюнь-чан; Юань, Чун-си; Ли, Да-цин; Чжан, Син; Лаковара, Кеннет Дж.; Додсон, Питер; Цзи, Цян (16 июня 2006 г.). «Почти современная птица-амфибия из раннего мела Северо-Западного Китая». Наука . 312 (5780): 1640–1643. Бибкод : 2006Sci...312.1640Y . дои : 10.1126/science.1126377 . ПМИД 16778053 . S2CID 42723583 .
- ^ Пирс Бродкорб (1976). Обнаружение птицы мелового периода, очевидно, предка отрядов Coraciiformes и Piciformes (Aves: Carinatae) [размер файла 70 МБ] (PDF) . Том. 27. Вклад Смитсоновского института в палеобиологию. стр. 67–73. Архивировано из оригинала (PDF) 19 октября 2014 года.
- ^ L. A. Nesov, A. A. Yarkov (1989). Новые птицы мел-палеогена СССР и некоторые замечания по истории возникновения и эволюции класса (New Cretaceous-Paleogene birds of USSR and some remarks about history of the origin and evolution of class) (PDF) (in Russian). Vol. 197. USSR Academy of scienses. p. 82. Archived (PDF) from the original on 27 March 2019.
- ^ Перейти обратно: а б Син, Лида; О'Коннор, Цзинмай К.; МакКеллар, Райан С.; Кьяппе, Луис М.; Ценг, Куовэй; Ли, Банда; Бай, Мин (сентябрь 2017 г.). «Детеныш энантиорнитина (Aves) среднего мела, сохранившийся в бирманском янтаре, с необычным оперением». Исследования Гондваны . 49 : 264–277. Бибкод : 2017GondR..49..264X . дои : 10.1016/j.gr.2017.06.001 .
- ^ Перейти обратно: а б Син, Лида; О'Коннор, Цзинмай К.; МакКеллар, Райан С.; Кьяппе, Луис М.; Бай, Мин; Ценг, Куовей; Чжан, Цзе; Ян, Хайдун; Фанг, Джун; Ли, Банда (февраль 2018 г.). «Плоский энантиорнитин в бирманском янтаре среднего мела: морфология и сохранность» . Научный вестник . 63 (4): 235–243. Бибкод : 2018SciBu..63..235X . дои : 10.1016/j.scib.2018.01.019 . ПМИД 36659012 .
- ^ Перейти обратно: а б с Син, Лида; МакКеллар, Райан С.; Ван, Мин; Бай, Мин; О'Коннор, Цзинмай К.; Бентон, Майкл Дж.; Чжан, Цзяньпин; Ван, Ян; Ценг, Куовэй; Локли, Мартин Г.; Ли, Банда; Чжан, Вэйвэй; Сюй, Син (28 июня 2016 г.). «Мумифицированные ранние птичьи крылья в бирманском янтаре среднего мела» . Природные коммуникации . 7 (1): 12089. Бибкод : 2016NatCo...712089X . дои : 10.1038/ncomms12089 . ПМЦ 4931330 . ПМИД 27352215 .
- ^ Перейти обратно: а б Син, Лида; МакКеллар, Райан С.; О'Коннор, Цзинмай К.; Бай, Мин; Ценг, Куовей; Кьяппе, Луис М. (30 января 2019 г.). «Полностью оперенный энантиорнитиновый фрагмент стопы и крыла, сохранившийся в бирманском янтаре середины мела» . Научные отчеты . 9 (1): 927. Бибкод : 2019NatSR...9..927X . дои : 10.1038/s41598-018-37427-4 . ПМК 6353931 . ПМИД 30700773 .
- ^ Син, Лида; МакКеллар, Райан С.; О'Коннор, Цзинмай К. (июнь 2020 г.). «Необычайно большое птичье крыло из бирманского янтаря середины мела». Меловые исследования . 110 : 104412. Бибкод : 2020CrRes.11004412X . дои : 10.1016/j.cretres.2020.104412 . S2CID 213510021 .
- ^ Син, Лида; О'Коннор, Цзинмай К.; Кьяппе, Луис М.; МакКеллар, Райан С.; Кэрролл, Натан; Ху, Хан; Бай, Мин; Лей, Фумин (22 июля 2019 г.). «Новая энантиорнитиновая птица с необычными пропорциями педалей, найденная в янтаре» . Современная биология . 29 (14): 2396–2401.e2. дои : 10.1016/j.cub.2019.05.077 . ISSN 0960-9822 . ПМИД 31303484 . S2CID 195887085 .
- ^ Перейти обратно: а б Чжан, Цзихуэй; Кьяппе, Луис М.; Хан, Банда; Чинсами, Анусуя (2013). «Крупная птица из раннего мела Китая: новые сведения о черепе энантиорнитинов». Журнал палеонтологии позвоночных . 33 (5): 1176–89. Бибкод : 2013JVPal..33.1176Z . дои : 10.1080/02724634.2013.762708 . S2CID 84677039 .
- ^ Чжоу, Чжунхэ; Кларк, Джулия; Чжан, Фучэн (май 2008 г.). «Понимание разнообразия, размера тела и морфологической эволюции крупнейшей энантиорнитиновой птицы раннего мела» . Журнал анатомии . 212 (5): 565–77. дои : 10.1111/j.1469-7580.2008.00880.x . ПМК 2409080 . ПМИД 18397240 .
- ^ Ху, Дунъюй; Сюй, Син; Хоу, Ляньхай; Салливан, Корвин (2012). «Новая энантиорнитиновая птица из нижнего мела Западного Ляонина, Китай, и ее значение для ранней эволюции птиц». Журнал палеонтологии позвоночных . 32 (3): 639–45. Бибкод : 2012JVPal..32..639H . дои : 10.1080/02724634.2012.652321 . S2CID 85942925 .
- ^ Аттерхольт, Джесси; Хатчисон, Дж. Ховард; О'Коннор, Цзинмай К. (13 ноября 2018 г.). «Наиболее полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae» . ПерДж . 6 : е5910. дои : 10.7717/peerj.5910 . ПМК 6238772 . ПМИД 30479894 .
- ^ Мартин, Энтони Дж.; Викерс-Рич, Патрисия; Рич, Томас Х.; Холл, Майкл; Ангельчик, Кеннет (январь 2014 г.). «Самые старые известные следы птиц из Австралии: формация Эумералла (Альб), бухта динозавров, Виктория» . Палеонтология . 57 (1): 7–19. Бибкод : 2014Palgy..57....7M . дои : 10.1111/пала.12082 .
- ^ Ван, Мин; Чжоу, Чжунхэ; Сюй, Гуанхуэй (01 января 2014 г.). «Первая энантиорнитиновая птица из верхнего мела Китая». Журнал палеонтологии позвоночных . 34 (1): 135–145. Бибкод : 2014JVPal..34..135W . дои : 10.1080/02724634.2013.794814 . ISSN 0272-4634 . S2CID 85255564 .
- ^ Перейти обратно: а б Карвальо, Исмар; НОВАС, Фернандо; Аньолин, Федерико; Исаси, Марсело; Фрейтас, Франциско; АНДРАДЕ, Хосе (5 июня 2015 г.). «Новый род и вид энантиорнитиновых птиц из раннего мела Бразилии» . Бразильский геологический журнал . 45 (2): 161–171. дои : 10.1590/23174889201500020001 .
- ^ Перейти обратно: а б О'Коннор, Цзинмай К.; Кьяппе, Луис М. (28 февраля 2011 г.). «Пересмотр морфологии черепа энантиорнитина (Aves: Ornithothoraces)». Журнал систематической палеонтологии . 9 (1): 135–157. Бибкод : 2011JSPal...9..135O . дои : 10.1080/14772019.2010.526639 . S2CID 86503357 .
- ^ Перейти обратно: а б с Ван, Мин; Ху, Хан; Ли, Чжихэн (21 августа 2015 г.). «Новая маленькая энантиорнитиновая птица из биоты Джехол, имеющая значение для ранней эволюции морфологии черепа птиц». Журнал систематической палеонтологии . 14 (6): 481–497. дои : 10.1080/14772019.2015.1073801 . S2CID 83603202 .
- ^ Ван, Мин; Ху, Хан (январь 2017 г.). «Сравнительное морфологическое исследование скуловых и квадратно-скуловых костей у ранних птиц и их родственников-динозавров» . Анатомическая запись . 300 (1): 62–75. дои : 10.1002/ar.23446 . ПМИД 28000410 . S2CID 3649504 .
- ^ Кьяппе, Луис М. (2009). «Уменьшенные динозавры: эволюционный переход к современным птицам» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 248–56. дои : 10.1007/s12052-009-0133-4 .
- ^ Беккер, Рэйчел (28 июня 2016 г.). «Птичьи крылья, заключённые в янтаре, — это окаменелость, появившаяся еще в эпоху динозавров». Природа . дои : 10.1038/nature.2016.20162 . S2CID 88601510 .
- ^ Перейти обратно: а б Чжан, Фучэн; Чжоу, Чжунхэ (октябрь 2004 г.). «Палеонтология: перья на ногах птиц раннего мела» . Природа . 431 (7011): 925. Бибкод : 2004Natur.431..925Z . дои : 10.1038/431925a . ПМИД 15496911 . S2CID 4322054 .
- ^ Перейти обратно: а б Кларк, Джулия А.; Чжоу, Чжунхэ; Чжан, Фучэн (март 2006 г.). «Взгляд на эволюцию полета птиц из новой клады раннемеловых орнитуринов из Китая и морфологию Yixianornis grabaui » . Журнал анатомии . 208 (3): 287–308. дои : 10.1111/j.1469-7580.2006.00534.x . ПМК 2100246 . ПМИД 16533313 .
- ^ Перейти обратно: а б О'Коннор, Цзинмай К.; Ван, Сюри; Кьяппе, Луис М.; Гао, Чуньлин; Мэн, Цинцзинь; Ченг, Сяодун; Лю, Цзиньюань (12 марта 2009 г.). «Филогенетическая поддержка специализированной клады энантиорнитиновых птиц мелового периода информацией от нового вида». Журнал палеонтологии позвоночных . 29 (1): 188–204. Бибкод : 2009JVPal..29..188O . дои : 10.1080/02724634.2009.10010371 . S2CID 196607241 .
- ^ Перейти обратно: а б Кьяппе, Луис М.; Бо, Чжао; О'Коннор, Цзинмай К.; Чуньлин, Гао; Сюри, Ван; Хабиб, Майкл; Маруган-Лобон, Хесус; Цинцзинь, Мэн; Сяодун, Ченг (2014). «Новый экземпляр раннемеловой птицы Hongshanornis longicresta : понимание аэродинамики и диеты базального орнитуроморфа» . ПерДж . 2 : е234. дои : 10.7717/peerj.234 . ПМЦ 3898307 . ПМИД 24482756 .
- ^ Перейти обратно: а б с д О'Коннор, Цзинмай К.; Ван, Сяоли; Чжэн, Сяотин; Ху, Хан; Чжан, Сяомэй; Чжоу, Чжунхэ (январь 2016 г.). «Энантиорнитин с веерообразным хвостом и эволюция ретрициального комплекса у ранних пташек» . Современная биология . 26 (1): 114–119. дои : 10.1016/j.cub.2015.11.036 . ПМИД 26748849 .
- ^ Перейти обратно: а б О'Коннор, Цзинмай К.; Ли, Да-Цин; Ламанна, Мэтью С.; Ван, Мин; Харрис, Джеральд Д.; Аттерхольт, Джесси; Ю, Хай-Лу (30 декабря 2015 г.). «Новый энантиорнитин раннего мела (Aves, Ornithothoraces) из северо-западного Китая со сложным орнаментом на хвосте». Журнал палеонтологии позвоночных . 36 (1): e1054035. дои : 10.1080/02724634.2015.1054035 . S2CID 85800831 .
- ^ Перейти обратно: а б Ван, Мин; О'Коннор, Цзинмай К.; Пан, Яньхун; Чжоу, Чжунхэ (31 января 2017 г.). «Причудливая энантиорнитиновая птица раннего мела с уникальными перьями на голенях и орнитуроморфным пигостилем в форме плуга» . Природные коммуникации . 8 : 14141. Бибкод : 2017NatCo...814141W . дои : 10.1038/ncomms14141 . ПМК 5290326 . ПМИД 28139644 .
- ^ Перейти обратно: а б Санс, Хосе Л.; Кьяппе, Луис М.; Бускалиони, Анжела Д. (1995). «Остеология Concornis lacustris (Aves: Enantiornithes) из нижнего мела Испании и пересмотр его филогенетических связей». Новитаты Американского музея (3133): 1–23. HDL : 2246/3667 .
- ^ Далла Веккья, Фабио М.; Кьяппе, Луис М. (2003). «Первый птичий скелет из мезозоя северной Гондваны». Журнал палеонтологии позвоночных . 22 (4): 856–60. doi : 10.1671/0272-4634(2002)022[0856:FASFTM]2.0.CO;2 . JSTOR 4524284 . S2CID 130143737 .
- ^ О'Коннор, Цзинмай К. (1 января 2019 г.). «Трофические привычки ранних птиц». Палеогеография, Палеоклиматология, Палеоэкология . 513 : 178–195. Бибкод : 2019PPP...513..178O . дои : 10.1016/j.palaeo.2018.03.006 . S2CID 133781513 .
- ^ О'Коннор, Цзинмай К.; Чжоу, Чжунхэ; Смит, Эндрю (6 ноября 2019 г.). «Эволюция пищеварительной системы современных птиц: выводы из паравианских окаменелостей из биоты Яньляо и Джехоль» . Палеонтология . 63 (1): 13–27. дои : 10.1111/пала.12453 .
- ^ Хьюстон, Дэвид С.; Копси, Дж. А. (1994). «Костное пищеварение и морфология кишечника бородача» . Журнал исследований хищников . 28 (2): 73–78.
- ^ «Короткий кристалл: кварц и окаменевшая птица | Георедкости» . 12 марта 2021 г. Проверено 6 мая 2021 г.
- ^ О'Коннор, Цзинмай К.; Чжоу, Чжунхэ; Чжан, Фучэн (28 февраля 2011 г.). «Переоценка Boluochia zhengi (Aves: Enantiornithes) и обсуждение внутрикладового разнообразия орнитофауны Джехол, Китай». Журнал систематической палеонтологии. 9 (1): 51–63. дои: 10.1080/14772019.2010.512614. S2CID 84817636.
- ^ Ляньхай Хоу; Луис М. Кьяппе; Фучэн Чжан; Ченг-Минг Чуонг (2004). «Новые окаменелости раннего мела из Китая документируют новую трофическую специализацию мезозойских птиц». Naturwissenschaften. 91 (1): 22–25. Бибкод: 2004NW.....91...22H. doi: 10.1007/s00114-003-0489-1. ПМЦ 4382005. ПМИД 14740099.
- ^ Моршхаузер, Э.М.; Варриккио, диджей; Гао, К.; Лю, Дж.; Ван, З.; Ченг, X. и Мэн, Q. (2009). «Анатомия раннемеловой птицы Rapaxavis pani, нового вида из провинции Ляонин, Китай». Журнал палеонтологии позвоночных. 29 (2): 545–554. дои: 10.1671/039.029.0210. S2CID 84643293.
- ^ Миллер, Кейс Винсент; Питтман, Майкл; Ван, Сяоли; Чжэн, Сяотин; Брайт, Джен А. (2022). «Рацион мезозойских зубатых птиц (Longipterygidae), полученный на основе количественного анализа существующих показателей рациона птиц» . БМК Биология . 20 (1): 101. дои : 10.1186/s12915-022-01294-3 . ПМК 9097364 . ПМИД 35550084 .
- ^ Санс, Хосе Л.; Кьяппе, Луис М.; Фернадес-Халво, Иоланда; Ортега, Франциско; Санчес-Шильон, Бегонья; Пойато-Ариса1, Франциско Дж.; Перес-Морено, Бернардино П. (февраль 2001 г.). «Раннемеловая окатушка». Природа . 409 (6823): 998–1000. Бибкод : 2001Natur.409..998S . дои : 10.1038/35059172 . ПМИД 11234054 . S2CID 663531 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Михайлов, Константин Евгеньевич (1991). «Классификация ископаемых яичных скорлупок амниотических позвоночных» (PDF) . Acta Palaeontologica Polonica . 36 (2): 193–238.
- ^ Михайлов, Константин Евгеньевич (1996). «Новые роды ископаемых яиц из верхнего мела Монголии». Палеонтологический журнал . 30 (2): 246–8.
- ^ Эльжановский, Анджей (1981). «Эмбриональные скелеты птиц из позднего мела Монголии» (PDF) . Палеонтология Полоника . 42 : 147–79.
- ^ Санс, Хосе Л.; Кьяппе, Луис М.; Перес-Морено, Бернардино П.; Мораталла, Хосе Дж.; Эрнандес-Карраскилья, Франциско; Бускалиони, Анжела Д.; Ортега, Франциско; Пойато-Ариса, Франциско Дж.; Раскин-Гутман, Диего; Мартинес-Дельклос, Ксавье (6 июня 1997 г.). «Птенец из нижнего мела Испании: значение для эволюции черепа и шеи птиц». Наука . 276 (5318): 1543–6. дои : 10.1126/science.276.5318.1543 .
- ^ Чжоу, Чжунхэ; Чжан, Фучэн (22 октября 2004 г.). «Преждевременный птичий эмбрион из нижнего мела Китая». Наука . 306 (5696): 653. doi : 10.1126/science.1100000 . ПМИД 15499011 . S2CID 34504916 .
- ^ Перейти обратно: а б с Кьяппе, Луис М.; Шуан, Цзи; Цян, Цзи (2007). «Молодые птицы раннего мела Китая: значение для онтогенеза энантиорнитина» . Новитаты Американского музея (3594): 1–46. doi : 10.1206/0003-0082(2007)3594[1:JBFTEC]2.0.CO;2 . hdl : 2246/5890 . S2CID 85871695 .
- ^ Эльжановский, Анджей (1995). «Меловые птицы и филогения птиц». Курьерский научно-исследовательский институт Зенкенберга . 181 : 37–53.
- ^ Курочкин Е.Н.; Чаттерджи, С.; Михайлов, К.Э. (декабрь 2013 г.). «Эмбриональная энантиорнитиновая птица и связанные с ней яйца из мелового периода Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Бибкод : 2013PalJ...47.1252K . дои : 10.1134/S0031030113110087 . S2CID 86747842 .
- ^ Курочкин Е.Н.; Чаттерджи, С.; Михайлов, К.Е. (19 декабря 2013 г.). «Эмбриональная энантиорнитиновая птица и связанные с ней яйца из мелового периода Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Бибкод : 2013PalJ...47.1252K . дои : 10.1134/S0031030113110087 . S2CID 86747842 .
- ^ Инкубационные периоды динозавров, определенные непосредственно по подсчету линий роста в эмбриональных зубах, демонстрируют развитие уровня рептилий - PNAS.
- ^ Происхождение птичьего клюва: новые сведения об инкубационных периодах динозавров.
- ^ Камбра-Му, Оскар; Бускалиони, Анхела Дельгадо; Кубо, Хорхе; Кастанье, Жак; Лот, Мари-Мадлен; де Маржери, Эммануэль; де Риклес, Арманд (2006). «Гистологические наблюдения энантиорнитиновой кости (Saurischia, Aves) из нижнего мела Лас-Хойяс (Испания)». Аккаунты Палевол . 5 (5): 685–91. Бибкод : 2006CRPal...5..685C . дои : 10.1016/j.crpv.2005.12.018 .
- ^ О'Коннор, Цзинмай К.; Ван, Мин; Чжэн, Сяо-Тин; Ван, Сяо-Ли; Чжоу, Чжун-Хе (2014). «Гистология двух самок раннемеловых птиц» (PDF) . Позвоночные Палазиатские . 52 (1): 112–28.
- ^ Кьяппе, LM (1995). «Филогенетическое положение меловых птиц Аргентины: Enantiornithes и Patagopteryx deferrariisi ». Курьер Форшунгсинститута Зенкенберга . 181 : 55–63.
- ^ Кубо, Хорхе; Бускалиони, Анжела Д.; Лежандр, Лукас Дж.; Бурдон, Эстель; Санс, Хосе Л.; Рикле, Арман (2021). «Палеогистологические выводы о скорости метаболизма в покое у Concornis и Iberomesornis (Enantiornithes, Ornithothoraces) из нижнего мела Лас-Хойяс (Испания)» . Палеонтология . 65 . дои : 10.1111/пала.12583 . S2CID 245082389 .
- ^ Дайк, Гарет; Времир, Матьяш; Кайзер, Гэри; Нэйш, Даррен (июнь 2012 г.). «Затонувшая мезозойская гнездовая колония птиц из позднего мела Трансильвании». Die Naturwissenschaften . 99 (6): 435–42. Бибкод : 2012NW.....99..435D . CiteSeerX 10.1.1.394.9006 . дои : 10.1007/s00114-012-0917-1 . ПМИД 22575918 . S2CID 1396792 .
- ^ Фернандес, Мариэла С.; Гарсия, Родольфо А.; Фиорелли, Лукас; Сколаро, Алехандро; Сальвадор, Родриго Б.; Котаро, Карлос Н.; Кайзер, Гэри В.; Дайк, Гарет Дж.; Фарке, Эндрю А. (17 апреля 2013 г.). «Большое скопление птичьих яиц из позднего мела Патагонии (Аргентина) раскрывает новую стратегию гнездования мезозойских птиц» . ПЛОС ОДИН . 8 (4): е61030. Бибкод : 2013PLoSO...861030F . дои : 10.1371/journal.pone.0061030 . ПМК 3629076 . ПМИД 23613776 .
- ^ О'Коннор, Цзинмай К.; Фальк, Аманда; Ван, Мин; Чжэн, Сяо-Тин (2020). «Первое сообщение о незрелых перьях молодых энантиорнитов из орнитофауны Джехол раннего мела». Позвоночные Палазиатские . 58 : 24–44. doi : 10.19615/j.cnki.1000-3118.190823 .
- ^ Кларк, Александр Д.; О'Коннор, Цзинмай К. (15 июня 2021 г.). «Изучение экоморфологии двух меловых энантиорнитинов с уникальной морфологией педалей» . Границы экологии и эволюции . 9 : 654156. дои : 10.3389/fevo.2021.654156 .
- ^ Кэй, Томас Г.; Питтман, Майкл; Маруган-Вольф, Иисус; Мартин-Эббот, Хью; Санс, Джозеф Луи; Бускалиони, Анджела Д. (21 марта 2019 г.). «Полноценный детеныш энантиорнитина, обнаруженный с помощью лазерно-стимулированной флуоресценции, поддерживает раннее гнездовое поведение» . Научные отчеты 9 (1): 5006. Бибкод : 2019НатСР... 9.5006К дои : 10.1038/ s41598-019-41423-7 ПМК 6428842 . ПМИД 30899080 .
- ^ Падиан, Кевин; Кьяппе, Луис М. (11 января 2007 г.). «Происхождение и ранняя эволюция птиц» (PDF) . Биологические обзоры . 73 (1): 1–42. дои : 10.1111/j.1469-185x.1997.tb00024.x . S2CID 86007060 .
- ^ Чжоу, Шуан; Чжоу, Чжун-Хе; О'Коннор, Цзинмай К. (2012). «Новая орнитуриновая птица с базальным клювом из нижнего мела Западного Ляонина, Китай» (PDF) . Позвоночные Палазиатские . 50 (1): 9–24.
- «Новая базальноклювая птица-орнитурин, найденная в нижнем меловом периоде Западного Ляонина, Китай» . Физика.орг . 13 февраля 2012 г.
- ^ Хоуд, Питер В. (1988). «Палеогнатные птицы раннего третичного периода Северного полушария». Публикации Орнитологического клуба Наттолла (Кембридж, Массачусетс, США: Орнитологический клуб Наттолла) 22
- ^ Навалон, Уильям; Маруган-Лобон, Хесус; Кьяппе, Луис М.; Луис Санс, Хосе; Бускалиони, Анжела Д. (6 октября 2015 г.). «Система мягких тканей и кожи в крыле раннемеловой птицы: значение для эволюции птичьего полета» . Научные отчеты . 5 (1): 14864. Бибкод : 2015NatSR...514864N . дои : 10.1038/srep14864 . ПМЦ 4594305 . ПМИД 26440221 .
- ^ Брусатте, Стивен Л.; О'Коннор, Цзинмай К.; Джарвис, Эрих Д. (5 октября 2015 г.). «Происхождение и разнообразие птиц» . Современная биология . 25 (19): 888–898 рэндов. Бибкод : 2015CBio...25.R888B . дои : 10.1016/j.cub.2015.08.003 . hdl : 10161/11144 . ISSN 1879-0445 . ПМИД 26439352 . S2CID 3099017 .
- ^ Аттерхольт, Джесси; Хатчисон, Дж. Ховард; О'Коннор, Цзинмай К. (2018). «Наиболее полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae» . ПерДж . 6 : е5910. дои : 10.7717/peerj.5910 . ISSN 2167-8359 . ПМК 6238772 . ПМИД 30479894 .
- ^ Ван, Ся; Макгоуэн, Алистер Дж.; Дайк, Гарет Дж.; Терви, Сэмюэл Т. (7 декабря 2011 г.). «Пропорции птичьих крыльев и стили полета: первый шаг к прогнозированию режимов полета мезозойских птиц» . ПЛОС ОДИН . 6 (12): e28672. Бибкод : 2011PLoSO...628672W . дои : 10.1371/journal.pone.0028672 . ПМЦ 3233598 . ПМИД 22163324 .
- ^ Кьяппе, Луис М.; Сузуки, Сигэру; Дайк, Гарет Дж.; Ватабе, Махито; Цогтбаатар, К.; Барсболд, Ринчен (январь 2007 г.). «Новая энантиорнитиновая птица из позднего мела пустыни Гоби». Журнал систематической палеонтологии . 5 (2): 193–208. Бибкод : 2007JSPal...5..193C . дои : 10.1017/S1477201906001969 . S2CID 85391743 .
- ^ Чжэн, Сяотин; Ван, Сяоли; О'Коннор, Цзинмай; Чжоу, Чжунхэ (9 октября 2012 г.). «Понимание ранней эволюции птичьей грудины у молодых энантиорнитинов» . Природные коммуникации . 3 (1): 1116. Бибкод : 2012NatCo...3.1116Z . дои : 10.1038/ncomms2104 . ПМИД 23047674 .
- ^ Перейти обратно: а б О'Коннор, Цзинмай; Дайк, Гарет (2010). «Переоценка Sinornis santensis и Cathayornis пишет (Aves: Enantiornithes)» . Записи Австралийского музея . 62 : 7–20. дои : 10.3853/j.0067-1975.62.2010.1540 .
- ^ Кларк, Джулия А.; Норелл, Марк А. (2002). «Морфология и филогенетическое положение апсарависа ухааны из позднего мела Монголии». Новитаты Американского музея (3387): 1–46. CiteSeerX 10.1.1.693.8475 . doi : 10.1206/0003-0082(2002)387<0001:TMAPPO>2.0.CO;2 . S2CID 52971055 .
- ^ Перейти обратно: а б Серено, ПК (2005) TaxonSearch: Stem Archosauria . Архивировано 19 февраля 2007 г. в Wayback Machine . Версия 1.0, 7 ноября 2005 г. Получено 2 октября 2006 г.
- ^ Ван, Сюри; Кау, Андреа; Ло, Сяолин; Кундрат, Мартин; У, Вэньшэн; Ю, Шубин; Го, Чжэнь; Лю, Ичуань; Цзи, Цян (11 февраля 2022 г.). «Новая бохайорнитидоподобная птица из нижнего мела Китая заполняет пробел в энантиорнитиновом неравенстве» . Журнал палеонтологии . 96 (4): 961–976. Бибкод : 2022JPal...96..961W . дои : 10.1017/jpa.2022.12 . ISSN 0022-3360 . S2CID 247432530 .
- ^ Карвальо; Новас; Аньолин; Исаси; Фрейтас; Андраде (2015). «Новый род и вид энантиорнитиновых птиц из раннего мела Бразилии» . Бразильский геологический журнал . 45 (2): 161–171. дои : 10.1590/23174889201500020001 .
- ^ Ван, Ли; О'Коннор, Чжоу; Ты (2015). «Второй вид энантиорнитиновых птиц из нижнего мелового бассейна Чангмы, северо-западный Китай, имеющий значение для таксономического разнообразия орнитофауны Чангмы». Меловые исследования . 55 : 56–65. Бибкод : 2015CrRes..55...56W . дои : 10.1016/j.cretres.2015.01.008 .
- ^ Ван, М.; О'Коннор, Дж. К.; Чжоу, З. (2014). «Новая крепкая энантиорнитиновая птица из нижнего мела Китая со сканирующими адаптациями». Журнал палеонтологии позвоночных . 34 (3): 657–671. Бибкод : 2014JVPal..34..657W . дои : 10.1080/02724634.2013.812101 . S2CID 85313872 .
- ^ Далсетт, Дж.; Эриксон, П.Г.; Чжоу, З. (2015). «Новый энантиорнит (Aves) из раннего мела Китая». Акта Геологика Синика . 86 (2): 801–807. дои : 10.1111/1755-6724.12270 . S2CID 131170912 .
- ^ Белл, Алисса К.; Кьяппе, Луис М.; Эриксон, Грегори М.; Сузуки, Сигэру; Ватабе, Махито; Барсболд, Ринчен; Цогтбаатар, К. (февраль 2010 г.). «Описание и экологический анализ Hollanda luceria, позднемеловой птицы из пустыни Гоби (Монголия)». Меловые исследования . 31 (1): 16–26. Бибкод : 2010CrRes..31...16B . дои : 10.1016/j.cretres.2009.09.001 .
- ^ Хартман, Скотт; Мортимер, Микки; Уол, Уильям Р.; Ломакс, Дин Р.; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ПМК 6626525 . ПМИД 31333906 .
- ^ Зеленков Никита В.; Аверьянов, Александр О. (13 июня 2015 г.). «Исторический экземпляр энантиорнитиновой птицы из раннего мела Монголии, представляющий новый таксон со специальной морфологией шеи». Журнал систематической палеонтологии . 14 (4): 319–338. дои : 10.1080/14772019.2015.1051146 . S2CID 85784633 .
- ^ Ван, М.; Лю, Д. (2015). «Таксономическая переоценка Cathayornithidae (Aves: Enantiornithes)». Журнал систематической палеонтологии . 14 : 1–19. дои : 10.1080/14772019.2014.994087 . S2CID 86665059 .
- ^ Ван, Сяоли; Кларк, Александр Д.; О'Коннор, Цзинмай К.; Чжан, Сянъюй; Ван, Син; Чжэн, Сяотин; Чжоу, Чжунхэ (27 февраля 2024 г.). «Первый беззубый энантиорнитин (Aves: Ornithothoraces) из нижнемеловой авифауны Джехол» . Меловые исследования . 159 (в печати): 105867. doi : 10.1016/j.cretres.2024.105867 . ISSN 0195-6671 .
- ^ Лю, Ди; Кьяппе, Луис М.; Серрано, Франциско; Хабиб, Майкл; Чжан, Югуан; Мэн, Циньцзин; Шоки, Мэтью (11 октября 2017 г.). «Аэродинамика полета энантиорнитинов: информация от новой китайской птицы раннего мела» . ПЛОС ОДИН . 12 (10): e0184637. Бибкод : 2017PLoSO..1284637L . дои : 10.1371/journal.pone.0184637 . ПМК 5636078 . ПМИД 29020077 .
- ^ Ху, Хан; О'Коннор, Цзинмай К. (14 ноября 2016 г.). «Первый вид энантиорнитов из Сихеданга проясняет развитие скелета энантиорнитов раннего мела». Журнал систематической палеонтологии . 15 (11): 909–926. дои : 10.1080/14772019.2016.1246111 . S2CID 89551799 .
- ^ Ван, Мин; Чжоу, Чжунхэ; Сюй, Гуанхуэй (7 января 2014 г.). «Первая энантиорнитиновая птица из верхнего мела Китая». Журнал палеонтологии позвоночных . 34 (1): 135–145. Бибкод : 2014JVPal..34..135W . дои : 10.1080/02724634.2013.794814 . S2CID 85255564 .
- ^ Ван, Мин; Чжоу, Чжунхэ (12 апреля 2017 г.). «Морфологическое исследование первой известной рыбоядной энантиорнитиновой птицы из раннего мела Китая». Журнал палеонтологии позвоночных . 37 (2): e1278702. Бибкод : 2017JVPal..37E8702W . дои : 10.1080/02724634.2017.1278702 . S2CID 89858642 .
- ^ Ху, Дунъюй; Лю, Ин; Ли, Цзиньхуа; Сюй, Син; Хоу, Ляньхай (июль 2015 г.). «Yuanjiawaornis viriosus, gen. et sp. nov., крупная энантиорнитиновая птица из нижнего мела западного Ляонина, Китай». Меловые исследования . 55 : 210–219. Бибкод : 2015CrRes..55..210H . дои : 10.1016/j.cretres.2015.02.013 .
- ^ О'Коннор, Патрик М.; Тернер, Алан Х.; Гроенке, Джозеф Р.; Феличе, Райан Н.; Роджерс, Раймонд Р.; Краузе, Дэвид В.; Рахантарисоа, Лидия Дж. (10 декабря 2020 г.). «Птица позднемелового периода с Мадагаскара демонстрирует уникальное развитие клюва». Природа . 588 (7837): 272–276. Бибкод : 2020Natur.588..272O . дои : 10.1038/s41586-020-2945-x . ПМИД 33239782 . S2CID 227174405 .
- ^ Ху, Хан; О'Коннор, Цзинмай К.; Чжоу, Чжунхэ; Фарке, Эндрю А. (3 июня 2015 г.). «Новый вид Pengornithidae (Aves: Enantiornithes) из нижнего мела Китая предполагает наличие специализированной скансорной среды обитания, ранее неизвестной у ранних птиц» . ПЛОС ОДИН . 10 (6): e0126791. Бибкод : 2015PLoSO..1026791H . дои : 10.1371/journal.pone.0126791 . ПМЦ 4454694 . ПМИД 26039693 .
- ^ Кьяппе, Луис М.; Цинцзинь, Мэн; Серрано, Франциско; Сигурдсен, Тронд; Мин, Ван; Белл, Алисса; Ди, Лю (25 октября 2019 г.). «Новая бохайорнисоподобная птица из раннего мела Китая: энантиорнитиновые взаимоотношения и летные качества» . ПерДж . 7 : е7846. дои : 10.7717/peerj.7846 . ПМК 6816414 . ПМИД 31667014 .
- ^ Ван, Ян; Ван, Мин; О'Коннор, Цзинмай К.; Ван, Сяоли; Чжэн, Сяотин; Чжан, Сяомэй (11 января 2016 г.). «Новая энантиорнитиновая птица Джехол с трехмерной сохранностью и фолликулами яичников». Журнал палеонтологии позвоночных . 36 (2): e1054496. Бибкод : 2016JVPal..36E4496W . дои : 10.1080/02724634.2015.1054496 . S2CID 85807045 .
- ^ Курочкин Е.Н.; Чаттерджи, С.; Михайлов, К.Е. (19 декабря 2013 г.). «Эмбриональная энантиорнитиновая птица и связанные с ней яйца из мелового периода Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Бибкод : 2013PalJ...47.1252K . дои : 10.1134/s0031030113110087 . S2CID 86747842 .
Внешние ссылки
[ редактировать ]![]() СМИ, связанные с Энантиорнитом, на Викискладе?
СМИ, связанные с Энантиорнитом, на Викискладе?

