Аутапоморфия
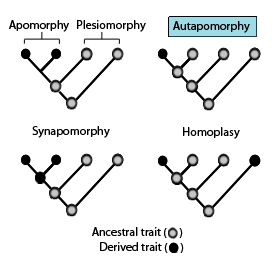
В филогенетике аутапоморфия — это отличительный признак, известный как производный признак, уникальный для данного таксона. То есть он встречается только в одном таксоне , но не встречается ни в каких других или чужих таксонах , даже в тех, которые наиболее тесно связаны с очаговым таксоном (который может быть видом , семейством или вообще какой-либо кладой). [2] Поэтому это можно считать апоморфией по отношению к одному таксону. [3] Слово аутапоморфия , введенное в 1950 году немецким энтомологом Вилли Хеннигом , происходит от греческих слов αὐτός, autos «сам»; ἀπό, апо «вдали от»; и μορφή, morphḗ = «форма». [4]
Обсуждение
[ редактировать ]Поскольку аутапоморфии присутствуют только в одном таксоне, они не передают информацию о родстве. Следовательно, аутапоморфии бесполезны для вывода о филогенетических отношениях. Однако аутапоморфия, как синапоморфия и плезиоморфия, является понятием относительным, зависящим от рассматриваемого таксона. Аутапоморфия на данном уровне вполне может быть синапоморфией на менее инклюзивном уровне. [5] Пример аутапоморфии можно описать у современных змей. Змеи утратили две пары ног, характерные для всех четвероногих , а наиболее близкие к Ophidia таксоны , как и их общие предки, имеют по две пары ног. Таким образом, таксон Ophidia представляет собой аутапоморфию в отношении отсутствия ног. [3]
Концепция аутапоморфных видов — один из многих методов, которые ученые могут использовать для определения и различения видов друг от друга. Это определение распределяет виды на основе степени дивергенции, связанной с репродуктивной несовместимостью, которая измеряется по существу количеством аутапоморфий. [6] Этот метод группировки часто называют «концепцией монофилетического вида» или концепцией «филовида», и он был популяризирован Д. Е. Розеном в 1979 году. В рамках этого определения вид рассматривается как «наименее инклюзивная монофилетическая группа, определяемая хотя бы одной аутапоморфией». ". [7] Хотя эта модель видообразования полезна тем, что позволяет избежать немонофилетических группировок, она также имеет свои критические замечания. Н. И. Платник, например, считает концепцию аутапоморфного вида неадекватной, поскольку она допускает возможность репродуктивной изоляции и видообразования при отмене «видового» статуса материнской популяции. Другими словами, если периферийная популяция отделится и станет репродуктивно изолированной, ей, вероятно, потребуется развить хотя бы одну аутапоморфию, чтобы ее признали другим видом. Если это может произойти без того, чтобы более крупная материнская популяция также не развила новую аутапоморфию, то материнская популяция не может оставаться видом в соответствии с концепцией аутапоморфного вида: у нее больше не будет никаких апоморфий, которые не свойственны также дочерним видам. [8]
Филогенетические сходства: эти филогенетические термины используются для описания различных моделей наследственных и производных состояний характера или черт, как указано на диаграмме выше, в связи с синапоморфиями. [1]
- Гомоплазия в биологической систематике — это когда признак приобретался или утрачивался независимо в отдельных линиях в ходе эволюции. Эта конвергентная эволюция приводит к тому, что виды независимо разделяют признак, отличающийся от признака, который, как предполагалось, присутствовал у их общего предка. [9] [10] [11]
- Параллельная гомоплазия – производный признак, присутствующий у двух групп или видов без общего предка в результате конвергентной эволюции . [12]
- Обратная гомоплазия – признак, присутствующий у предка, но не у прямых потомков, но вновь появляющийся у более поздних потомков. [13]
- Апоморфия – производный признак. Апоморфия, присущая двум или более таксонам и унаследованная от общего предка, называется синапоморфией. Апоморфия, уникальная для данного таксона, является аутапоморфией. [14] [15] [16] [17]
- Синапоморфия / Гомология – производный признак, который встречается в некоторых или всех терминальных группах клады и унаследован от общего предка, для которого он был аутапоморфией (т.е. не присутствовал у его непосредственного предка).
- Базовая синапоморфия - синапоморфия, которая снова утрачена у многих представителей клады. Если потеряно все, кроме одного, его будет трудно отличить от аутапоморфии.
- Аутапоморфия – отличительный производный признак, уникальный для данного таксона или группы. [3]
- Симплезиоморфия – наследственный признак, общий для двух или более таксонов.
- Плезиоморфия - симплезиоморфия, обсуждаемая в отношении более производного состояния.
- Псевдоплезиоморфия – признак, который нельзя идентифицировать ни как плезиоморфию, ни как апоморфию, являющуюся инверсией. [18]
- Реверсия – это утрата производного признака, присутствующего у предка, и восстановление плезиоморфного признака.
- Конвергенция – независимая эволюция сходного признака у двух и более таксонов.
- гемиплазия [19] [20]
Ссылки
[ редактировать ]- ^ Jump up to: а б Пейдж РД, Холмс ЕС (14 июля 2009 г.). Молекулярная эволюция: филогенетический подход . Джон Уайли и сыновья. ISBN 978-1-4443-1336-9 . OCLC 609843839 .
- ^ Футуйма диджей (1998). Эволюционная биология (3-е изд.). Sinauer Associates, Inc. с. 95.
- ^ Jump up to: а б с Аппель Р.Д., Фейтманс Э. (2009). «Глава 3: Введение в филогенетику и ее молекулярные аспекты». Биоинформатика: швейцарская перспектива (1-е изд.). Мировое научное издательство.
- ^ Калоу П.П. (2009). Энциклопедия экологии и природопользования . Джон Уайли и сыновья. ISBN 978-1-4443-1324-6 . OCLC 1039167559 .
- ^ Фори П.Л. (1997). История целакантов (1-е изд.). Спринтер.
- ^ Ховард DJ, Берлохер С.Х. (1998). Бесконечные формы: виды и видообразование (1-е изд.). США: Издательство Оксфордского университета. ISBN 978-0-19-510901-6 . ОСЛК 60181901 .
- ^ Булл АТ (2004). Микробное разнообразие и биоразведка . АСМ Пресс.
- ^ Платник Н.И. (2001). « От кладограмм к классификациям: путь к дефилокоду » . (PDF) . Ассоциация систематиков.
- ^ Гаугер А. (17 апреля 2012 г.). «Сходство случается! Проблема гомоплазии» . Эволюция сегодня и новости науки .
- ^ Сандерсон М.Дж., Хаффорд Л. (21 октября 1996 г.). Гомоплазия: повторение сходства в эволюции . Эльзевир. ISBN 978-0-08-053411-4 . ОСЛК 173520205 .
- ^ Брэндли MC, Уоррен Д.Л., Личе А.Д., Макгуайр Дж.А. (апрель 2009 г.). «Гомоплазия и поддержка клад» . Систематическая биология . 58 (2): 184–98. дои : 10.1093/sysbio/syp019 . ПМИД 20525577 .
- ^ Арчи Дж.В. (сентябрь 1989 г.). «Коэффициенты избытка гомоплазии: новые индексы для измерения уровней гомоплазии в филогенетической систематике и критика индекса согласованности». Систематическая зоология . 38 (3): 253–269. дои : 10.2307/2992286 . JSTOR 2992286 .
- ^ Wake DB, Wake MH, Specht CD (февраль 2011 г.). «Гомоплазия: от обнаружения закономерностей к определению процесса и механизма эволюции». Наука . 331 (6020): 1032–5. Бибкод : 2011Sci...331.1032W . дои : 10.1126/science.1188545 . ПМИД 21350170 . S2CID 26845473 .
- «Гомоплазия: хорошая нить, которую можно потянуть, чтобы понять клубок эволюционной нити» . ScienceDaily (пресс-релиз). 25 февраля 2011 г.
- ^ Симпсон М.Г. (9 августа 2011 г.). Систематика растений . Амстердам: Эльзевир. ISBN 9780080514048 .
- ^ Рассел П.Дж., Герц П.Е., Макмиллан Б. (2013). Биология: динамическая наука . Cengage Обучение. ISBN 978-1-285-41534-5 .
- ^ Липскомб Д. (1998). «Основы кладистического анализа» (PDF) . Вашингтон, округ Колумбия: Университет Джорджа Вашингтона.
- ^ Чоудхури С (9 мая 2014 г.). Биоинформатика для начинающих: гены, геномы, молекулярная эволюция, базы данных и аналитические инструменты (1-е изд.). Академическая пресса. п. 51. ИСБН 978-0-12-410471-6 . OCLC 950546876 .
- ^ Уильямс Д., Шмитт М., Уиллер К. (21 июля 2016 г.). Будущее филогенетической систематики: наследие Вилли Хеннига . ISBN 978-1-107-11764-8 . OCLC 951563305 .
- ^ Avise JC, Робинсон TJ (июнь 2008 г.). «Гемиплазия: новый термин в лексиконе филогенетики» . Систематическая биология . 57 (3): 503–7. дои : 10.1080/10635150802164587 . ПМИД 18570042 .
- ^ Копетти Д., Буркес А., Бустаманте Е., Шарбоно Дж.Л., Чайлдс К.Л., Эгиарте Л.Е. и др. (ноябрь 2017 г.). «Обширное несоответствие генного дерева и гемиплазия сформировали геномы колонновидных кактусов Северной Америки» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (45): 12003–12008. дои : 10.1073/pnas.1706367114 . ПМЦ 5692538 . ПМИД 29078296 .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||