Словарь птичьих терминов

Ниже приводится словарь общеупотребительных англоязычных терминов, используемых при описании птиц — теплокровных позвоночных животных класса Aves и единственных ныне живущих динозавров . [1] характеризуются перьями , способностью летать у всех, кроме примерно 60 существующих видов нелетающих птиц , беззубыми, клювистыми челюстями , откладкой яиц с твердой скорлупой , высокой скоростью обмена веществ , четырехкамерным сердцем и крепким, но легким скелетом .
Среди других деталей, таких как размер, пропорции и форма, были разработаны термины, определяющие особенности птиц, которые используются для описания особенностей, уникальных для этого класса, особенно эволюционных адаптаций, которые развились для облегчения полета . Существует, например, множество терминов, описывающих сложную структурную структуру перьев (например, бородки , рахиды и лопасти ); типы перьев (например, нитевидные , перистые и перистые перья); а также их рост и потеря (например, изменение цвета , брачного оперения и птерилеза ).
Существуют тысячи терминов, уникальных для изучения птиц. Этот глоссарий не пытается охватить их все, а концентрируется на терминах, которые можно встретить в описаниях множества видов птиц, сделанных любителями птиц и орнитологами . Хотя также включены слова, которые не являются уникальными для птиц, например « спина » или « живот », они определяются в связи с другими уникальными особенностями внешней анатомии птиц , иногда называемыми « топографией ». Как правило, этот глоссарий не содержит отдельных записей ни о одном из примерно 11 000 признанных ныне живущих отдельных видов птиц мира. [2] [3] [а]
А [ править ]
- запутанные яйца
- Also, wind eggs; hypanema.[5] Eggs that are not viable and will not hatch.[6] See related: overbrooding.
- afterfeather
- Any structure projecting from the shaft of the feather at the rim of the superior umbilicus (at the base of the vanes), but typically a small area of downy barbs growing in rows or as tufts.[b] Entirely absent in some birds—notably from many members of the Columbidae family (pigeons and doves)—afterfeathers can significantly increase the insulative attributes of a bird's plumage.[8]
- allopreening
- A form of social grooming among birds, in which one bird preens another or a pair does so mutually. At times it may be used to redirect or sublimate aggression, such as one bird assuming a solicitation posture to indicate its non-aggression and invite allopreening by the aggressive individual.[9]
- alternate plumage
- Also, nuptial plumage; breeding plumage. The plumage of birds during the courtship or breeding season. It results from a prealternate moult that many birds undergo just prior to the season. The alternate plumage is commonly brighter than the basic plumage, for the purposes of sexual display, but may also be cryptic, to hide incubating birds that might be vulnerable on the nest.[10]
- altricial
- Also defined: semi-altricial; altricial-precocial spectrum. Young that, at hatching, have their eyes closed; are naked or only sparsely covered in down feathers (psilopaedic); are not fully able to regulate their body temperature (ectothermic);[11] and are unable to walk or leave the nest for an extended period of time to join their parents in foraging activities (nidicolous), whom they rely on for food.[12] The contrasting state is precocial young, which are born more or less with their eyes open, covered in down, homeothermic, able to leave the nest and ambulate and to participate in foraging.[11][13] The young of many bird species do not precisely fit into either the precocial or altricial category, having some aspects of each and thus fall somewhere on an altricial-precocial spectrum.[14] A defined intermediate state is termed semi-altricial, typified by young that, though born covered in down (ptilopaedic), are unable to leave the nest or walk and are reliant on their parents for food.[15][16]
- alula
- Also, bastard wing; alular digit; alular quills.[17] A small, freely-moving projection on the anterior edge of the wing of modern birds (and a few non-avian dinosaurs)—a bird's "thumb"—the word is Latin and means 'winglet'; it is the diminutive of ala, meaning 'wing'. Alula typically bear three to five small flight feathers, with the exact number depending on the species. The bastard wing normally lies flush against the anterior edge of the wing proper, but can be raised to function in similar manner to the slats of airplane wings that aid in lift by allowing a higher than normal angle of attack. By manipulating the alula structure to create a gap between it and the rest of the wing, a bird can avoid stalling when flying at low speeds or when landing. Feathers on the alular digit are not generally considered to be flight feathers in the strict sense; though they are asymmetrical, they lack the length and stiffness of most true flight feathers. Nevertheless, alula feathers are a distinct aid to slow flight.[18]
- anisodactylous
- Descriptive of tetradactyl (four-toed) birds in which the architecture of the foot consists of three toes projecting forward and one toe projecting backward (the hallux), such as in most passerine species.[19][20]
- anting
- Also defined: passive anting. A self-anointing behaviour during which birds rub insects, usually ants and sometimes millipedes, on their feathers and skin. Anting birds may pick up the insects in their bill and rub them on their bodies, or may simply lie in an area with a high density of the insects and perform dust bathing-like movements. Insects used for anting secrete chemical liquids such as formic acid, which can act as insecticides, miticides, fungicides and bactericides. The practice may also act to supplement a bird's own preen oil. A third purpose may be to render the insects more palatable, by causing removal of distasteful compounds. More than 200 species of bird are known to ant.[21] "Passive anting" refers to when birds simply position themselves so as to allow insects to crawl through their plumage.[6]
- apical spot
- A visible spot near the outer tip of a feather.[22]
- apterylae
- Singular: apteryla. Also, apteria. Regions of a bird's skin, between the pterylae (feather tracts), which are free of contour feathers; filoplumes and down may grow in these areas.[23][24] See related: pterylosis.
- aviculture
- The captive breeding and keeping of birds.[6]
- axilla
- Also, axillar region; "underarm"; "armpit". The "armpit" of a bird, often hosting covert feathers called axillaries.[25]
- axillaries
- Also, axillary feathers; lower humeral coverts; hypopteron. Covert feathers found in the axillar region or "armpit" of a bird, which are typically long, stiff and white in colour.[26]


B[edit]
- back
- The exterior region of a bird's upper parts between the mantle and the rump.[27]
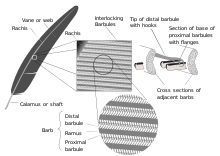
- barb
- Also defined: ramus (plural: rami). The individual structures growing out of the shaft that collectively make up the vanes of the feathers, more or less interconnected by the hooklets of the barbules, extending from each side of the distal part of the feather's shaft known as the rachis. The central axis of a barb is known as the ramus.[8]
- barbules
- Also, radius / radii; tertiary fibres.[28] Also defined: proximal barbules; distal barbules; barbicels; hooklets (hamuli); pennulum; teeth. Just as barbs branch off on parallel sides of the rachis, the barbs in turn have a set of structures called barbules, branching from each side of the ramus. The base cells of the barbule form a plate from which a thinner stalk projects called the pennulum. At one more level of branching, the pennulum hosts small outgrowths from it called barbicels—which when found on pennaceous feathers, vary in structure depending on which side of a barb's ramus they project from. Proximal barbules (on the proximal side of the ramus) have ventral projections near the base called teeth, while growing from the pennulum are cilia—simple pointed structures. At the base of the proximal barbule, the "dorsal edge is recurved into a flange".[29] The distal barbules (on the distal side of the ramus) have a thicker base with more elaborate teeth, and a longer pennulum with hooklets (also called hamuli[30]) at the end, as well as cilia in greater number than on proximal barbules. The hooklets overlap one to four rows of proximal barbules on the next higher barb, locking into their flanges, thereby giving the vane structure, strength, flexibility and stability.[29] See also: friction barbules.
- basic plumage
- Also, winter plumage; non-breeding plumage. Also defined: supplementary plumage. The plumage of birds during the non-breeding season. It results from the prebasic moult that many birds undergo just after the season, and sometimes (rarely) even a second non-breeding season moult (resulting in what's termed "supplementary plumage") prior to the next breeding season.[c] The basic plumage is commonly duller than the alternate or nuptial plumage.[10][31]
- beak
- Also, bill or rostrum. An external anatomical structure of a bird's head, roughly corresponding with the "nose" of a mammal, that is used for eating, grooming, manipulating objects, killing prey, fighting, probing for food, courtship and feeding young. Although beaks vary significantly in size and shape from species to species, their underlying structures follow a similar pattern. All beaks are composed of two jaws, generally known as the upper mandible (or maxilla) and lower mandible (or mandible),[32] covered with a thin, horny sheath of keratin called the rhamphotheca,[33][34] which can be subdivided into the rhinotheca of the upper mandible and the gnathotheca of the lower mandible.[35] The tomia (singular: tomium) are the cutting edges of the two mandibles.[36] Most species of birds have external nares (nostrils) located somewhere on their beak—two holes—circular, oval or slit-like in shape—which lead to the nasal cavities within the bird's skull, and thus to the rest of the respiratory system.[37] Although the word beak was, in the past, generally restricted to describing the sharpened bills of birds of prey,[38] in modern ornithology, the terms beak and bill are generally used synonymously.[33]
- beak colour
- The colour of a bird's beak results from concentrations of pigments—primarily melanins and carotenoids—in the epidermal layers, including the rhamphotheca.[39] In general, beak colour depends on a combination of the bird's hormonal state and diet. Colours are typically brightest as the breeding season approaches and palest after breeding.[40]
- beak trimming
- Also, debeaking and coping. The partial removal of the beak of poultry, especially layer hens and turkeys, although it may also be performed on quail and ducks. Because the beak is a sensitive organ with many sensory receptors, beak trimming or debeaking is "acutely painful"[41] to the birds it is performed on. It is nonetheless routinely done to intensively farmed bird species, because it helps reduce the damage the flocks inflict on themselves due to a number of stress-induced behaviours, including cannibalism, vent pecking and feather pecking. A cauterizing blade or infrared beam is used to cut off about half of the upper beak and about a third of the lower beak. Pain and sensitivity can persist for weeks or months after the procedure, and neuromas can form along the cut edges. Food intake typically decreases for some period after the beak is trimmed. However, studies show that trimmed poultry's adrenal glands weigh less, and their plasma corticosterone levels are lower than those found in untrimmed poultry, indicating that they are less stressed overall.[41] A less radical, separate practice, usually performed by an avian veterinarian or an experienced birdkeeper, involves clipping, filing or sanding the beaks of captive birds for health purposes—in order to correct or temporarily alleviate overgrowths or deformities and better allow the bird to go about its normal feeding and preening activities.[42] "Coping" is the name for this practice amongst raptor keepers.[43]
- belly
- Also, abdomen. The topographical region of a bird's underparts between the posterior end of the breast and the vent.[44]
- billing
- Also, nebbing (chiefly UK). Describes the tendency of mated pairs of many bird species to touch or clasp each other's bills.[45] This behaviour appears to strengthen pair bonding.[46] The amount of contact involved varies among species. Some gently touch only a part of their partner's beak while others clash their beaks vigorously together.[47]
- bill tip organ
- A region found near the tip of the bill in several types of birds that forage particularly by probing. The region has a high density of nerve endings known as the corpuscles of Herbst. These consist of pits in the bill's surface lined with cells that sense pressure changes. The assumption is that these cells allow a bird to perform "remote touch", meaning that it can detect the movement of animals by pressure variations in water, without directly touching the prey. Bird species known to have bill-tip organs include ibises, shorebirds of the family Scolopacidae and kiwis.[48]
- bird ringing
- Also, bird banding. The attachment of small, individually numbered metal or plastic tags to the legs or wings of wild birds to enable individual identification. The practice helps in keeping track of bird movement and life history. Upon capture for ringing, it is common to take measurements and examine the conditions of feather moult, amount of subcutaneous fat, age indications and sex. The subsequent recapture or recovery of banded birds can provide information on migration, longevity, mortality, population, territoriality, feeding behaviour and other aspects that are studied by ornithologists. Bird ringing is the term used in the UK and in some other parts of Europe, while the term bird banding is more often used in the U.S. and Australia.[49]
- bird strike
- The impact of a bird or birds with an airplane in flight.[50]
- body down
- The layer of small, fluffy down feathers that lie underneath the outer contour feathers on a bird's body.[51] Compare: natal down and powder down.
- breast
- The topographical region of a bird's external anatomy between the throat and the belly.[52]
- breeding plumage
- See alternate plumage.
- brood
- The collective term for the offspring of birds, or the act of brooding the eggs.[53]
- brooding
- See egg incubation.
- broodiness
- Also defined: broody. The action or behavioural tendency of a bird to sit on a clutch of eggs to incubate them, often requiring the non-expression of many other behaviours including feeding and drinking.[54] The adjective "broody" is defined as "[b]eing in a state of readiness to brood eggs that is characterized by cessation of laying and by marked changes in behavior and physiology". Example usage: "a broody hen".[55]
- brood patch
- A bare patch of skin that most female birds gain during the nesting season for thermoregulation purposes, by shedding feathers close to the belly, in an area that will be in contact with the eggs during incubation. The patch of bare skin is well supplied with blood vessels at the surface, facilitating heat transfer to the eggs.[56]
- brood parasite
- Birds, such as the common goldeneye, indigobirds, whydahs, honeyguides, cowbirds and New World cuckoos, that lay their eggs in other birds' nests, in order to have their chicks incubated through fledging by the parents of another bird, often of another species.[57][58][59]


C[edit]
- calamus
- Plural: calami. The basal part of the quill of pennaceous feathers, which embeds at its proximal tip in the skin of a bird. The calamus is hollow and has pith formed from the dry remains of the feather pulp. The calamus stretches between two openings—at its base is the inferior umbilicus and at its distal end is the superior umbilicus; the rachis of the stem, hosting the vanes, continues above it.[60][61] Calamus derives from the Latin for 'reed' or 'arrow'.[62]
- call
- Specific types: alarm; contact; duet; antiphonal duetting; food begging; flight; mobbing. A type of bird vocalization tending to serve such functions as giving alarm or keeping members of a flock in contact—as opposed to a bird's song, which is longer, more complex and is usually associated with courtship and mating.[63] Individual birds may be sensitive enough to identify each other through their calls. Many birds that nest in colonies can locate their chicks using their calls.[64] Alarm calls are used to sound alarm to other individuals. Food-begging calls are made by baby birds to beg for food, such as the "wah" of infant blue jays.[65] Mobbing calls signal other individuals in mobbing species while harassing a predator. They differ from alarm calls, which alert other species members to allow escape from predators. As an example, the great tit, a European songbird, uses such a signal to call on nearby birds to harass a perched bird of prey, such as an owl. This call occurs in the 4.5kHz range,[66] and carries over long distances. However, when such prey species are in flight, they employ an alarm signal in the 7–8 kHz range. This call is less effective at traveling great distances, but is much more difficult for both owls and hawks to hear (and detect the direction from which the call came).[67] Contact calls are used by birds for the purpose of letting others of their species know their location.[68] Relatedly, flight calls are vocalizations made by birds while flying, which often serve to keep flocks together.[69] These calls are also used for when birds want to alert others that they are taking flight.[70] Many birds engage in duet calls—a call made by two birds at or nearly at the same time. In some cases, the duets are so perfectly timed as to appear almost as one call. This kind of calling is termed antiphonal duetting.[71] Such duetting is noted in a wide range of families including quails,[72] bushshrikes,[73] babblers such as scimitar babblers and some owls[74] and parrots.[75]
- canopy feeding
- Also defined: double-wing feeding. Some herons, such as the black heron, adopt an unusual position while hunting for prey. With their head held down in a hunting position, they sweep their wings forward to meet in front of their head, thereby forming an umbrella shaped canopy. To achieve full canopy closure, the primaries and secondaries touch the water, the nape feathers are erected and the tail is drooped. The bird may take several strides in this position. One theory about the function of this behaviour is that it reduces glare from the water surface, allowing the bird to more easily locate and catch prey. Alternatively, the shade provided by the canopy may attract fish making their capture easier. Some herons adopt a similar behaviour called double-wing feeding in which the wings are swept forward to create an area of shade, though a canopy is not formed.[76]
- carpal bar
- A patch seen on the upperwing of some birds that usually appears as a long stripe or line. It is created by the contrast between the greater coverts and the other wing feathers.[77]
- caruncle
- The collective term for the several fleshy protuberances on the heads and throats of gallinaceous birds, i.e., combs, wattles, ear lobes and nodules. They can be present on the head, neck, throat, cheeks or around the eyes of some birds. Caruncles may be featherless, or present with a small array of scattered feathers. In some species, they may form pendulous structures of erectile tissue, such as the "snood" of the domestic turkey.[78][79] While caruncles are ornamental elements used by males to attract females to breed,[80] it has been proposed that these organs are also associated with genes that encode resistance to disease,[81] and for birds living in tropical regions, that caruncles also play a role in thermoregulation by making the blood cool faster when flowing through them.[82]
- casque
- A horny ridge found on the upper mandible of a bird's bill, especially used in relation to hornbills and cassowaries,[83][84] though other birds may have casques such as common moorhens,[85] tufted puffins[86] and (male) friarbirds.[87] The ridge line on the upper maxilla may extend to a prominent crest on the front of the face and on the head, such as in the "flamboyant" crest of the rhinoceros hornbill.[88] Some hornbill casques contain a hollow space that may act as a resonance chamber, amplifying calls.[89] Similar function has been proposed for cassowary casques, as well as for protection of the head while dense vegetation is traversed, as a sexual ornament and for use as a "shovel" for digging food.[90] Compare: frontal shield.
- cere
- From the Latin cera meaning 'wax', a waxy structure which covers the base of the bills of some bird species from a handful of families—including raptors, owls, skuas, parrots, turkeys and curassows. The cere structure typically contains the nares, except in owls, where the nares are distal to the cere. Although it is sometimes feathered in parrots,[91] the cere is typically bare and often brightly coloured.[92] In raptors, the cere is a sexual signal which indicates the "quality" of a bird; the orangeness of a Montague's harrier's cere, for example, correlates to its body mass and physical condition.[93] The colour or appearance of the cere can be used to distinguish between males and females in some species. For example, the male great curassow has a yellow cere, which the female (and young males) lack,[94] and the male budgerigar's cere is blue, while the female's is pinkish or brown.[95]
- cheek
- Also, malar / malar region. The area of the sides of a bird's head, behind and below the eyes.[96]
- chin
- A small feathered area located just below the base of the bill's lower mandible.[97]
- cloaca
- A multi-purpose opening terminating at the vent at the posterior of a bird: birds expel waste from it; most birds mate by joining cloaca (a "cloacal kiss"); and females lay eggs from it. Birds do not have a urinary bladder or external urethral opening and (with exception of the ostrich) uric acid is excreted from the cloaca, along with faeces, as a semisolid waste.[98][99][100] Additionally, in those few bird species in which males possess a penis (Palaeognathae [with the exception of the kiwis], the Anseriformes [with the exception of screamers] and in rudimentary forms in Galliformes[101][102]) it is hidden within the proctodeum compartment within the cloaca, just inside the vent.[103]
- cloacal kiss
- Most male birds lack a phallus and instead have erectile genital papilla at the terminus of their vas deferens. When male and female birds of such species copulate, they each evert and then press together, or "kiss" their respective proctodeum (the lip of the cloaca). Upon the clocal kiss, the male's sperm spurts into the female's urodoeum (a compartment inside the cloaca), which then make their way into the oviduct.[104][105]
- clutch
- All of the eggs produced by birds often at a single time in a nest. Clutch size differs greatly between species, sometimes even within the same genus. It may also differ intraspecies due to many factors including habitat, health, nutrition, predation pressures and time of year.[106] Average clutch size ranges from one (as in northern gannet[107]) to about 17 (as in grey partridge[108]).
- comb
- Also, cockscomb (coxcomb and other sp. variants). A fleshy growth or crest on the top of the head of gallinaceous birds, such as turkeys, pheasants and domestic chickens. Its alternative name, cockscomb (or coxcomb) reflects that combs are generally larger on males than on females (a male gallinaceous bird is called a cock). Comb shape varies considerably depending on the breed or species of bird. The "comb" most often refers to chickens in which the most common shape is the "single comb" of a rooster from breeds such as the leghorn. Other common comb types are the "rose comb" of, e.g., the eponymous rosecomb; the "pea comb" of, e.g., the brahma and araucana; and others.[109]
- colony
- Also defined: seabird colony; breeding colony; communal roost; heronries; rookery. A large congregation of individuals of one or more species of bird that nest or roost in proximity at a particular location. Many kinds of birds are known to congregate in groups of varying size; a congregation of nesting birds is called a breeding colony. A group of birds congregating for rest is called a communal roost. Approximately 13% of all bird species nest colonially.[110] Nesting colonies are very common among seabirds on cliffs and islands. Nearly 95% of seabirds are colonial,[111] leading to the usage, seabird colony, sometimes called a rookery. Many species of terns nest in colonies on the ground. Herons, egrets, storks and other large waterfowl also nest communally in what are called heronries. Colony nesting may be an evolutionary response to a shortage of safe nesting sites and abundance or unpredictable food sources which are far away from the nest sites.[112]
- colour morph
- See morph.
- commissure
- Depending on usage, may refer to the junction of the upper and lower mandibles,[113] or alternately, to the full-length apposition of the closed mandibles, from the corners of the mouth to the tip of the beak.[114]
- contact call
- A type of call used by birds for the purpose of letting others of their species know their location.[68]
- corpuscles of Herbst
- Nerve-endings similar to the Pacinian corpuscle, found in the mucous membrane of the tongue, in pits on the beak and in other parts of the bodies of birds. They differ from Pacinian corpuscles in being smaller and more elongated, in having thinner and more closely placed capsules and in that the axis-cylinder in the central clear space is encircled by a continuous row of nuclei.[115]
- coverts
- Also, covert feathers; tectrices – singular: tectrix. A layer of non-flight feathers overlaying and protecting the quills of flight feathers. At least one layer of covert feathers appear both above and beneath the flight feathers of the wings as well as above and below the rectrices of the tail.[116] These feathers may vary widely in size. For example, the upper tail tectrices of peacocks—the male peafowl—rather than its rectrices, are what constitute its elaborate and colourful "train".[117] There are a number of types and subtypes of covert feathers—primary, secondary, greater, lesser, marginal, median, etc.—see broadly wing coverts and tail coverts.
- cranial kinesis
- Also defined: prokinesis, amphikinesis and distal rhynchokinesis. Movement of the upper mandible in relation to the front of the skull. There is very little of this movement in birds that feed primarily through grazing and thus do not need to open their bills very widely. This is in contrast to parrots, which use their bills to manipulate food and as a support when climbing trees. There are multiple types of cranial kinesis: prokinesis, where the bill moves only at the craniofacial hinge; amphikinesis, where the whole upper jaw is raised; and distal rhynchokinesis, where the bill flexes somewhere along the length of the bill, compared to just at the base.[118]
- crest feather
- Collectively, the/a crest. Long crest feathers are sometimes called quill feathers.[119] Also defined: recumbent crests and recursive crests. A type of semiplume feather with a long rachis with barbs on either side, that often presents as a prominent tuft on the crown and (or through) the neck and upper back.[120][121] Birds with crests include Victoria crowned pigeons, northern lapwings, macaroni penguins and others, but the most recognizable are cockatoos and cockatiels, which can raise or lower their crests at will and use them to communicate with fellow members of their species, or as a form of defence to frighten away other species that approach too closely, making the bird appear larger when the crest is suddenly and unexpectedly raised.[122] In some species the position of the crest is a threat signal that can be used to predict behaviour. In Steller's jays, for example, a raised crest indicates a likelihood of attack, and a lowered crest indicates a likelihood of retreat.[123] Crests can be recumbent or recursive, depending on the species. The recumbent crest, such as in white cockatoos, has feathers that are straight and lie down essentially flat on the head until fanned out.[124] The recursive crest, such as in sulphur-crested cockatoos and Major Mitchell's cockatoos, is noticeable even when its feathers are not fanned out because they curve upward at the tips even when lying flat, and when standing up, often bend slightly forward toward the front of the head. Some birds, like galahs (also known as the rose-breasted cockatoo), have modified crests that have both recumbent and recursive features.[122]
- crissum
- The feathered area between the vent and the tail. Also, the collective name for the undertail coverts.[125] The crissal thrasher derives its name from the term, having distinctive colouring in the region, in contrast with the balance of its plumage. Other birds having distinctive crissum colouration include Le Conte's thrashers and grey catbirds.[126]
- crop
- An expanded, muscular pouch near the gullet or throat found in some but not all birds. It is a part of the digestive tract, essentially an enlarged part of the esophagus, used for the storage of food prior to digestion. As with most other organisms that have a crop, birds use it to temporarily store food. In adult doves and pigeons, the crop can produce crop milk to feed newly-hatched chicks.[127]
- crop milk
- A secretion from the lining of the crop of parent birds that is regurgitated to young birds. It is found among all pigeons and doves where it is referred to as pigeon milk. An analogue to crop milk is also secreted from the esophagus of flamingos and some penguins.[128][129][130] Crop milk bears little physical resemblance to mammalian milk, the former being a semi-solid substance somewhat like pale yellow cottage cheese. It is extremely high in protein and fat, containing higher levels than cow or human milk[131] and has been shown to also contain antioxidants and immune-enhancing factors.[132]
- crown
- Also defined: occiput / hindhead. The portion of a bird's head found between the forehead—demarcated by an imaginary line drawn from the anterior corners of the eyes—and through the "remainder of the upper part of the head", to the superciliary line. The occiput or hindhead, is the posterior part of the crown.[133]
- cryptic plumage
- Also defined: phaneric plumage. Plumage of a bird that is camouflaging. For example, the white winter plumage of ptarmigans is cryptic as it serves to conceal it in snowy environments.[134] The opposite, "advertising" plumage, is termed "phaneric", such as male birds in colourful nuptial plumage for sexual display, making them stand out to a high degree.[135]
- culmen
- The dorsal ridge of the upper mandible.[136] Likened by ornithologist Elliott Coues to the ridge line of a roof, it is the "highest middle lengthwise line of the bill" and runs from the point where the upper mandible emerges from the forehead's feathers to its tip.[137] The bill's length along the culmen is one of the regular measurements made during bird ringing[138] and is particularly useful in feeding studies.[139] The shape or colour of the culmen can also help with the identification of birds in the field. For example, the culmen of the parrot crossbill is strongly decurved, while that of the very similar appearing red crossbill is more moderately curved.[140]







D[edit]
- definitive plumage
- Adult plumage sufficiently developed and fixed following the juvenile years such that it no longer changes significantly in appearance with age.[31]
- diastataxis
- The state of lacking a fifth secondary feather on each wing, which occurs in birds in more than 40 non-passerine families. In these birds, the fifth set of secondary covert feathers does not cover any remex. Loons, grebes, pelicans, hawks and eagles, cranes, sandpipers, gulls, parrots and owls are among the families missing this feather.[18]
- dietary classification terms (-vores)
- Birds may be classified by terms related to the types of foods they forage for and eat.[141] The -vore suffix is derived from the Latin vorare, meaning 'to devour'. Equivalent adjectives can be formed through use of the suffix -vorous.[142] For example, granivore (n.) / granivorous (adj.). Generally, classification terms are used based on predominance of food source and/or specialization. There can be much cross-over and mixing between classifications. For example, insectivores and piscivores may at times be described more broadly as types of carnivores, and hummingbirds, though they do eat insects, are often described as nectarivores, rather than insectivores, as nectar is a specialized and predominant food foraging source for that bird family.[143] The feeding strategies of birds is intimately tied to their physiology and evolutionary development. For example, beak shape and structure such as the makeup of the tomia, the presence (or not) of a bill tip organ and myriad other adaptations are tied to a species' feeding strategies.[144] Feeding habits also correlate with aspects of brain development and size. For example, the spatial memory of birds that store food in various locations has been shown to be highly developed and contributes towards success at that feeding tactic.[145]
carnivores (sometimes called faunivores): birds that predominantly forage for the meat of vertebrates—generally hunters as in certain birds of prey—including eagles, owls and shrikes, though piscivores, insectivores and crustacivores may be called specialized types of carnivores.[146] crustacivores: birds that forage for and eat crustaceans, such as crab-plovers and some rails.[146] detritivores: birds that forage for and eat decomposing material, such as vultures. It is usually used as a more general term than "saprovore" (defined below), which often connotes the eating of decaying flesh alone.[147] florivores: birds that forage for and eat plant material in general. Other terms for plant foraging specialization may apply to florivorous species, such as "frugivore" and "granivore".[148] folivores: birds that forage for and eat leaves, such as hoatzin and mousebirds.[141][146] frugivores: birds that forage for and eat fruit, such as turacos, tanagers and birds-of-paradise.[146] granivores: (sometimes called seed-eating): birds that forage for seeds and grains,[149] such as geese, grouse and estrildid finches.[141][146] herbivore: birds that predominantly eat plant material, and mostly do not eat meat; especially of birds that are both granivorous and frugivorous or are grass eaters, such as whistling ducks, ostriches and mute swans.[146][150] insectivores: birds that forage for and eat insects and other arthropods, such as cuckoos, swallows, thrushes, drongos and woodpeckers.[141][146] nectarivores: birds that drink the nectar of flowers, such as hummingbirds, sunbirds and lorikeets.[146] omnivores (sometimes called general feeders): birds that forage for a variety of both plant and meat food sources, such as pheasants, tinamouses and quails. More birds fall under the omnivore classification than any other.[146] piscivores: birds that forage for and eat fish and other sea life, such as darters, loons, pelicans, penguins and storks.[141][146] sanguinivores: birds that forage for and drink blood, such as oxpeckers and sharp-beaked ground finches.[141][148] saprovores: birds that forage for and eat decaying flesh (carrion), such as vultures and crows.[141] However, the term is also used at times synonymously with "detritivore" (defined above), for eaters of any dead matter.[141][147][151] - dimorphism
- See sexual dimorphism.
- diving
- Also defined: surface diving; plunge diving. Some birds dive into the water for food. Two diving strategies are differentiated: surface diving birds dive from the surface of the water and swim actively underwater; and plunge diving birds dive from the air into the water. Plunge diving birds may use the momentum from the plunge to propel themselves underwater, whereas others may swim actively.[152]
- down
- Also, down feathers or plumulaceous feathers. The down of birds—their plumulaceous feathers, as opposed to pennaceous feathers—are a layer of fine, silky feathers found under the tougher exterior feathers, that are often used by humans as a thermal insulator and padding in goods such as jackets, bedding, pillows and sleeping bags. Considered to be the "simplest" of all feather types,[153] down feathers have a short or vestigial rachis (shaft), few barbs and barbules that lack hooks,[154] and (unlike contour feathers) grow from both the pterylae and the apteria.[61] Very young birds are often only clad in down. The loose structure of down feathers traps air, which helps to insulate birds against heat loss[51] and contributes to the buoyancy of waterbirds. Species that experience annual temperature fluctuations typically have more down feathers following their autumn moult.[155] There are three types of down: natal down, body down and powder down.[156]
- drumming
- A form of non-vocal communication engaged in by members of the woodpecker family. It involves the beak striking a hard surface multiple times per second. The drumming pattern, the number of beats per roll and the gap between rolls is specific to each species. Drumming is usually associated with territorial behaviour, with male birds drumming more frequently than females.[157] Drumming may also refer to the sounds produced by the specialised outer tail-feathers of snipe in the course of their courtship display flights.[158]

E[edit]

- ear-coverts
- Small covert feathers located behind a bird's eye, in one to four rows, which cover the ear opening (bird ears have no external features[159]) and may aid in the acuity of bird hearing.[160]
- egg
- Also defined: eggshell; yolk; albumen; chalaza. The organic vessel containing the zygote, in which birds develop until hatching. Eggs are usually oval in shape, and have a base white colour from the predominant calcium carbonate makeup of the outer shell, called the eggshell, though passerine birds especially may have eggs of other colours,[161] such as through deposition of biliverdin and its zinc chelate, which give a green or blue ground colour, and protoporphyrin which produces reds and browns.[162] A viable bird egg (as opposed to a non-viable egg: see addled eggs) consists of a number of structures. The eggshell is 95–97% calcium carbonate crystals, at least in chickens, stabilized by a protein matrix,[163][164][165] without which the crystalline structure would be too brittle to keep its form; the organic matrix is thought to have a role in deposition of calcium during the mineralization process.[166][167][168] The structure and composition of the avian eggshell serves to protect the egg against damage and microbial contamination, prevention of desiccation, regulation of gas and water exchange for the growing embryo and provides calcium for embryogenesis.[164] Inside the eggshell are two shell membranes (inner and outer), and at the center is a yolk—a spherical structure, usually some shade of yellow, to which the fertilized gamete attaches and which the embryonic bird uses as sustenance as it grows. The yolk is suspended in the albumen (also called egg white or glair / glaire) by one or two spiral bands of tissue called the chalazae.[169] The albumen protects the yolk and provides additional nutrition for the embryo's growth,[170] though it is made up of approximately 90% water in most birds.[171] Prior to fertilization, the yolk is a single cell ovum or egg cell; one of the few single cells that can be seen by the naked eye.[172]
- egg binding
- An egg that while traversing the reproductive tract during the process of being laid, becomes stuck near to the opening of the cloaca or further inside the oviduct.[173] The condition may be caused by obesity, nutritional imbalances such as calcium deficiency, environmental stress such as temperature changes, or malformed eggs.[174]
- egg incubation
- Also, brooding. The general care of unhatched eggs by parent birds (more often by females but by birds of both sexes), especially by temperature regulation through sitting on them, crouching or squatting over them, covering them with their wings, providing shade, wetting eggs and related behaviours. The target temperature of most species is 37 °C (99 °F) to 38 °C (100 °F). In monogamous species incubation duties are often shared, whereas in polygamous species one parent is wholly responsible for incubation. Warmth from parents passes to the eggs through brood patches—areas of bare skin on the abdomen or breast of the incubating birds. Incubation can be an energetically demanding process; adult albatrosses, for instance, lose as much as 83 grams (2.9 oz) of body weight per day of incubation.[175][176][177]
- egg tooth
- A small, sharp, calcified projection on the beak that full-term chicks of most bird species have, which they use to chip their way out of their egg.[178] This white spike is located near the tip of the upper mandible in most species (e.g., gulls);[179] near the tip of the lower mandible instead in a minority of others, such as northern lapwings;[179] with a few species, such as Eurasian whimbrels, black-winged stilts and semipalmated sandpipers,[179] having one on each mandible.[180] Despite its name, the projection is not an actual tooth (as the similarly-named projections of some reptiles are); instead, it is part of the integumentary system, as are claws and scales.[181] The hatching chick first uses its egg tooth to break the membrane around an air chamber at the wide end of the egg. Then it pecks at the eggshell while turning slowly within the egg, eventually (over a period of hours or days) creating a series of small circular fractures in the shell.[182] Once it has breached the egg's surface, the chick continues to chip at it until it has made a large hole. The weakened egg eventually shatters under the pressure of the bird's movements.[183] The egg tooth is so critical to a successful escape from the egg that chicks of most species will perish unhatched if they fail to develop one.[180]
- emargination
- A pronounced narrowing at some variable distance along the feather edges at the outermost primaries of large soaring birds, particularly raptors. Whether these narrowings are called notches or emarginations' depends on the degree of their slope.[18] An emargination is a gradual change, and can be found on either side of the feather. A notch is an abrupt change, and is only found on the wider trailing edge of the remige. The presence of notches and emarginations creates gaps at the wingtip; air is forced through these gaps, increasing the generation of lift.[184]
- eye-ring
- Also defined: orbital ring. A visible ring of feathers around a bird's eye; the eye-ring is often paler than the surrounding feathers. By contrast, an orbital ring is bare skin ringing the eye. In some species, such as little ringed plover, the orbital ring may be quite conspicuous.[97]
- eyestripe
- Also, eye line / eyeline. A visible stripe on the feathers of a bird's head, often darker than the surrounding feathers, running through the eye region.[97] Compare supercilium.
 Blue-headed vireo, with a conspicuous eye-ring
Blue-headed vireo, with a conspicuous eye-ring Little ringed plover chick, with a conspicuous orbital ring
Little ringed plover chick, with a conspicuous orbital ring White-eared sibia, with a conspicuous eyestripe
White-eared sibia, with a conspicuous eyestripe Whinchat, with a conspicuous supercilium (eyebrow)
Whinchat, with a conspicuous supercilium (eyebrow)





F[edit]

- feather
- Epidermal growths that form the distinctive outer covering, or plumage, on birds. They are considered the most complex integumentary structures found in vertebrates,[185][186] and, indeed, a premier example of a complex evolutionary novelty.[187] Feathers are among the characteristics that distinguish the extant birds from other living groups.[188] Although feathers cover most parts of the body of birds, they arise only from certain well-defined tracts on the skin. They aid in flight, thermal insulation and waterproofing, with their colouration helping in communication and protection.[189] Although there are many subdivisions of feathers, at the broadest levels, feathers are either classified as i) vaned feathers, which cover the exterior of the body and include pennaceous feathers, or ii) down feathers, which grow underneath the vaned feathers. A third rarer type of feather, the filoplume, is hairlike and (if present in a bird; they are entirely absent in ratites[190]) grows alongside the contour feathers.[185] A typical vaned feather features a main shaft called the quill with an upper section called the rachis. Fused to the rachis are a series of branches, or barbs; the barbs in turn have barbules branching off them, and they in turn branch yet again with a series of growths called barbicels, some of which have minute hooks called hooklets for cross-attachment. Down feathers are fluffy because they lack barbicels, so the barbules float free of each other, allowing the down to trap air and provide excellent thermal insulation. At the base of the feather, the rachis expands to form the hollow tubular calamus which inserts into a follicle in the skin. The basal part of the calamus is without vanes. This part is embedded within the skin follicle and has an opening at the base (proximal umbilicus) and a small opening on the side (distal umbilicus).[191]
- feather pecking
- A behavioural problem in which one bird repeatedly pecks at the feathers of another, that occurs most frequently amongst domestic hens reared for egg production,[192][193] although it is seen in other poultry such as pheasants,[194] turkeys,[195] ducks[196] and sometimes in farmed ostriches.[197] Two levels of severity are recognised: "gentle" and "severe".[198]
- feather-plucking
- Also feather-picking, feather damaging behaviour or pterotillomania.[199] A maladaptive, behavioural disorder commonly seen in captive birds which chew, bite or pluck their own feathers with their beak, resulting in damage to the feathers and occasionally the skin.[200][201] It is especially common among Psittaciformes, with an estimated 10% of captive parrots exhibiting the disorder.[202] The chief areas of the body that are pecked or plucked are the more accessible regions such as the neck, chest, flank, inner thigh and ventral wing area. Contour and down feathers are generally identified as the main targets, although in some cases, tail and flight feathers are affected. Although feather-plucking shares characteristics with feather pecking, commonly seen in commercial poultry, the two behaviours are currently considered to be distinct, as in the latter, the birds peck at and pull out the feathers of other individuals.
- feather tract
- See: pterylae.
- fecal sac
- Also, faecal sac. A mucous membrane, generally white or clear with a dark end,[203] that surrounds the feces of some species of nestling birds,[204] and allows parent birds to more easily remove fecal material from the nest. The nestling usually produces a fecal sac within seconds of being fed; if not, a waiting adult may prod around the youngster's cloaca to stimulate excretion.[205] Young birds of some species adopt specific postures or engage in specific behaviours to signal that they are producing fecal sacs.[206] For example, nestling curve-billed thrashers raise their posteriors in the air, while young cactus wrens shake their bodies.[207] Other species deposit the sacs on the rim of the nest, where they are likely to be seen (and removed) by parent birds.[206] Not all species generate fecal sacs. They are most prevalent in passerines and their near relatives, which have young that remain in the nest for longer periods.[205]
- filoplume
- Also, filoplume feather; hair feather, thread feather. A hairlike type of feather that, if present in a bird (they are entirely absent in ratites[190]) grows alongside the contour feathers.[185] The typical filoplume is silky in appearance, lacks pith and a superior umbilicous opening, has a very slender, straight shaft lacking differentiation into calamus and rachis, and is naked or has only a few barbs (that lack cross-attachment) at the distal end. They are closely associated with contour feathers and often entirely hidden by them, with one or two filoplumes attached and sprouting from near the same point of the skin as each contour feather, at least on a bird's head, neck and trunk.[208][209] Filoplume feathers host a cluster of sensory corpuscles at their base,[210] that serve to detect air currents that affect contour and flight feathers.[92] Filoplumes are one of the three major classes of feathers, the others being pennaceous and plumulaceous feathers.
- flange
- Also, recurved margin. "The thickened dorsal edge of the bases of pennaceous barbules, generally recurved in proximal barbules, and frequently so in distal barbules also"[211] which anchor the hooklets.[29] (See diagram; refer to figures 3 [in which the flange is referred to as the "folded edge"] and 6 [in which the mechanism of interlocking between a flange and hooklet is shown].)
- flanks
- The topographical region of the underparts[d] sketched "between the posterior half of the abdomen and the rump".[96]
- fledge
- Also, fledging. The stage in a young bird's life when the feathers and wing muscles are sufficiently developed for flight, or describing the act of a chick's parents in raising it to that time threshold.[212]
- fledgling
- A juvenile bird during the period it is venturing from or has left the nest and is learning to run and fly; a young bird during the period immediately after fledging, when it is still dependent upon parental care and feeding.[213]
- flight
- Most birds can fly, which distinguishes them from almost all other vertebrate classes (cf. bats and pterosaurs). Flight is the primary means of locomotion for most bird species and is used for breeding, feeding and predator avoidance and escape. Birds have various adaptations for flight, including a lightweight skeleton, two large flight muscles, the pectoralis (which accounts for 15% of the total mass of the bird) and the supracoracoideus, as well as modified forelimbs (wings) that serve as aerofoils.[214] Wing shape and size generally determine a bird species' type of flight; many birds combine powered, flapping flight with less energy-intensive soaring flight. About 60 extant bird species are flightless, as were many extinct birds.[215] Flightlessness often arises in birds on isolated islands, probably due to limited resources and the absence of land predators.[216] Though flightless, penguins use similar musculature and movements to "fly" through the water, as do auks, shearwaters and dippers.[217]
- flight feather
- Also, Pennae volatus.[218] The long, stiff, asymmetrically shaped, but symmetrically paired pennaceous feathers on the tail or wings of a bird; those on the tail are called rectrices (singular: rectrix), while those on the wings are called remiges (singular: remex). Based on their location, remiges are subdivided into primaries, secondaries and tertiaries.[219]
- foot paddling
- A foraging behaviour of gulls in which individuals stand at a location, often in shallow water, and perform rapid stepping actions that are thought to make subterranean worms or other food rise to the surface.[220]
- forehead
- The portion of a bird's head extending "up and back from the bill to an imaginary line joining the anterior corners of the eyes".[133]
- fovea
- Plural: foveas or foveae. A small cavity in the retina of the eye that hosts a large number of light receptors; more than anywhere else on the retina. About one half of bird species with fovea have a single one, but uniquely in birds,[221] some, such as terns, kingfishers and hummingbirds, have a second fovea,[222] called the temporal fovea, that assists in judging speed and distance and increases visual acuity. Birds that do not have a second fovea will sometimes bob their head to improve their visual field.[223]
- friction barbules
- A specialized type of barbule located on the distal part of the inner vane of primary feathers on the wings of most flighted birds. Friction barbules support lobe-shaped ("lobular") barbicels that are broader than the typical barbicel hosted by other vaned feathers, and which in turn support more hooklets. The theory is that the augmented surface area and other adaptations significantly increases grip through friction when the outer web of barbs of one primary feather come into contact and rub against the inner web of barbs of another primary feather it overlays, thereby preventing slippage during the rigors of flight.[224][225] Friction barbules are found only on those parts of primary feathers that are in "zones of overlap" with neighboring primaries. The theory (and the use of "friction" in the title of the defined phrase) has been criticized. In Avian Flight (2005), the author notes that "most birds open and close their wings during every wing beat cycle", and proposes that the energetic cost to overcome friction during the "wing extension and flexion" of each beat cycle would be prohibitive. Offered instead is the theory that the function of these specialized barbules is to lock the primary feathers together on the wings' downstroke, during which high pressure from below the wings' surface would otherwise tend to cause the feathers to spread.[226]
- frontal shield
- Also facial shield; face shield; frontal plate. A hard or fleshy plate extending from the base of the upper mandible over the forehead of several bird species including some water birds in the rail family, especially the gallinules and moorhens, swamphens and coots, as well as in jacana.[227] While most face shields are made up of fatty tissues, some birds, such as certain turacos, e.g., the red-crested turaco, have face shields that are hard extensions of the mandible.[228] The size, shape and colour may exhibit testosterone-dependent variation in either sex during the year.[229] Functionality appears to relate to protection of the face while feeding in or moving through dense vegetation, as well as to courtship display and territorial defence.[230] Compare: casque.
- furcula
- Also, wishbone; merry-thought. From the Latin for "little fork", the furcula is a forked bone, also found in some dinosaurs, located below the neck and formed by the fusion of the two clavicles. Its primary function is in the strengthening of the thoracic skeleton to withstand the rigors of flight. It works as a strut between a bird's shoulders, and articulates to each of a bird's scapulae. In conjunction with the coracoid and the scapula, it forms a unique structure called the triosseal canal, which houses a strong tendon that connects the supracoracoideus muscles to the humerus. This system is responsible for lifting the wings during a recovery stroke. As the thorax is compressed by the flight muscles during a downstroke, the upper ends of the furcula spread apart, expanding by as much as 50% of its resting width, and then contract. Furcula may also aid in respiration, by helping to pump air through the air sacs.[231][232]



G[edit]
- gape
- The interior of the open mouth of a bird.[233] The width of the gape can be a factor in the choice of food.[234]
- gape flange
- The region where the upper and lower mandibles join together at the base of the beak.[233] When born, the chick's gape flanges are fleshy. As it grows into a fledgling, the gape flanges remain somewhat swollen and can thus be used to recognize that a particular bird is young.[235] Gape flanges can serve as a target for food for parents, and when touched, stimulate the nestling to open its mouth to eat.[236]
- gizzard
- Also, ventriculus; gastric mill; gigerium. A specialized stomach organ found in the digestive tract of some birds constructed of thick muscular walls that is used for grinding up food, often aided by particles of stone or grit. Food, after going through the crop and proventriculus, passes into the gizzard where it can be ground with previously swallowed stones and passed back to the proventriculus, and vice versa. Bird gizzards are lined with a tough layer made of a carbohydrate-protein complex called koilin, that protects the muscles in the gizzard.[237][238]
- gleaning
- Specialized cases: foliage gleaning; hover-gleaning; crevice-gleaning. The strategy of gleaning over surfaces by birds to catch invertebrate prey—chiefly insects and other arthropods—by plucking them from foliage or the ground, from crevices such as of rock faces and under the eaves of houses, or even, as in the case of ticks and lice, from living animals. Gleaning the leaves and branches of trees and shrubs is called "foliage gleaning", which can involve a variety of styles and maneuvers. Some birds, such as the common chiffchaff[239] of Eurasia and the Wilson's warbler of North America, feed actively and appear energetic. Some will even hover in the air near a twig while gleaning from it; this behaviour is called "hover-gleaning". Other birds are more methodical in their approach to gleaning, even seeming lethargic as they perch upon and deliberately pick over foliage. This behaviour is characteristic of the bay-breasted warbler[240] and many vireos. Another tactic is to hang upside-down from the tips of branches to glean the undersides of leaves. Tits such as the familiar black-capped chickadee are often observed feeding in this manner. Some birds, like the ruby-crowned kinglet, use a combination of these tactics. "Crevice-gleaning" is a niche particular to dry and rocky habitats. Gleaning birds are typically small with compact bodies and have small, sharply pointed beaks. Birds often specialize in a particular niche, such as a particular stratum of forest or type of vegetation.[241]
- gnathotheca
- The rhamphotheca (thin horny sheath of keratin) covering the lower mandible.[33][34][35]
- gonys
- The ventral ridge of the lower mandible, created by the junction of the bone's two rami, or lateral plates.[242]
- gonydeal angle
- Also, gonydeal expansion. The proximal end of the junction created by the gonys two rami, or lateral plates—the place where the two plates separate. The size and shape of the gonydeal angle can be useful in identifying between otherwise similar species.[40]
- gonydeal spot
- The spot near the gonydeal expansion, in adults of many bird species but especially in gulls, usually reddish or orangish in colour, that triggers begging behaviour. Gull chicks, for example, peck at the spot on its parent's bill, which in turn stimulates the parent to regurgitate food.[40][243]
- gorget
- A patch of coloured feathers found on the throat or upper breast of some species of birds.[242] It is a feature found on many male hummingbirds, particularly those found in North America, whose gorgets are typically iridescent.[244] Other species having gorgets include the purple-throated fruitcrow[245] and the chukar partridge.[246] The term is derived from the gorget used in military armor to protect the throat.[247]
- grooming
- See entry for preening.
- gular region
- The posterior part of the underside of a bird's head, described as "a continuation of the chin to an imaginary line drawn between the angles of the jaw".[8] See also: gular skin.
- gular skin
- Also defined, gular sac / throat sac; gular pouch; gular flutter. Describes the gular region when it is featherless. In many species, the gular skin forms a flap, or gular pouch, which is generally used to store fish and other prey while hunting. In many others, such as frigatebirds, the gular skin may overlie a sac—the gular sac or throat sac—that may be dramatically inflated by males during courtship display. In some species the gular region is used for thermoregulation, by the fluttering of the hyoid bone and surrounding muscles, vibrating as many as 735 times per minute, which causes an increase in heat dissipation from the gular skin.[248][249][250]


H[edit]
- hallux
- Also, hind toe; first digit. Specialized types: incumbent and elevated. A bird's usually rear-facing toe; its hind toe. When it is attached near the base of the metatarsus it is termed incumbent, and when projecting from a higher portion of the metatarsus (as in rails), it is termed elevated. The hallux is a bird's first digit, in many birds being the sole rear-facing toe (including in most passerine species that have anisodactylous feet) and is homologous to the human big toe.[251][252][253]
- home range
- Also defined: territory. The area of land in which a bird performs most of its activities. When the area is guarded (in whole or in part) from individuals of the same species it is called a territory.[254] These territories may be temporary or permanent, breeding or non-breeding. Breeding territories may be of four types: i) an all-purpose territory where all activities are conducted; ii) a separate breeding and feeding territory (or just a breeding territory, with foraging conducted outside of it); iii) a territory surrounding the nest and a very small area outside of it; iv) a small territory used solely for display within a lek.[255] The size of the home range can be affected by what food a bird eats and how much the bird weighs. Birds that weigh more usually have larger ranges, and carnivores on average have larger ranges than non-carnivores.[256]
- Humphrey–Parkes terminology
- A system of nomenclature for the plumage of birds proposed in 1959 by Philip S. Humphrey and Kenneth C. Parkes[10] to make the terminology for describing bird plumages more uniform.[257] Examples of Humphrey–Parkes terminology versus traditional terminology may be seen in the entries for prealternate moult and prebasic moult.
- hyoid apparatus
- The system of bones to which the tongue is attached. It usually includes the tongue bone, to which the tongue is actually attached,[258] the basihyal, behind the tongue bone, the urohyal, itself behind the basihyal, a pair of ceratobranchial bones, and a pair of epibranchial.[259] The latter two bones form the hyoid horns, which are contained in a pair of fascia vaginalis. This allows the tongue to slide out smoothly. The hyoid apparatus is attached to the larynx.[258]
I[edit]
- inferior umbilicus
- Also, proximal umbilicus. A small opening located at the bottom tip of a feather's shaft, i.e., at the base of the calamus, that is embedded within the skin of a bird. The growth of the feather is fed by the flow through the inferior umbilicus of a nutrient (or nutritive) pulp of highly vascular dermal cells (sometimes called the nutritive dermis; a part of what is termed the Malpighian layer), that in turn produce a nourishing plasma. In mature feathers the opening is sometimes sealed over by a keratinous plate.[189][260][261] Compare: superior umbilicus.
- inner wing
- Also defined: outer wing. The inner wing of a bird is that portion of the wing stretching from its connection to the body and through the "wrist" joint. The outer wing stretches from the wrist to the wingtip.[262]
- iris
- The coloured outer ring that surrounds a bird's pupil. Though brown predominates, the iris may be of or include a variety of colours—red, yellow, grey, blue, etc.—and the colouration may vary according to the age, sex and species.[159]
J[edit]
- jizz
- Also, gestalt,[263] of which the term may be a corruption[264] (or possibly deriving from the air force acronym GISS for "General Impression of Size and Shape (of an aircraft)", associated with World War II lingo; though this derivation may by anachronistic as jizz was first used in birdwatching as early as 1922),[265] it describes the overall impression or appearance of a bird—"the indefinable quality of a particular species, the 'vibe' it gives off"[266]—garnered from such features as shape, posture, flying style or other habitual movements, size and colouration combined with voice, habitat and location.[267][268][269] Example use: "It had the jizz of a thrush, but I couldn't make out what kind."
K[edit]
- keel
- Also, carina. An extension of the sternum (breastbone) which runs axially along the midline of the sternum and extends outward, perpendicular to the plane of the ribs. The keel provides an anchor to which a bird's wing muscles attach, thereby providing adequate leverage for flight. Keels do not exist on all birds; in particular, some flightless birds lack a keel structure. Historically, the presence or absence of a pronounced keel structure was used as a broad classification of birds into two orders: Carinatae (from Latin carina, 'keel'), having a pronounced keel; and ratites (from Latin ratis, 'raft'—referring to the flatness of the sternum), having a subtle keel structure or lacking one entirely. However, this classification has fallen into disuse as evolutionary studies have shown that many flightless birds have evolved from flighted birds.[270][271]
- kleptoparasitism
- A bird's propensity to steal food from other birds, either opportunistically, or on a regular basis.[272]
L[edit]
- lateral throat-stripe
- A usually dark stripe of colour bordering the throat below both the moustachial stripe and the submoustachial stripe, if any.[27]
- lek
- Also defined: lekking; dispersed lek. An aggregation of male birds gathered to engage in competitive displays (known as lekking) that may entice visiting females that are assessing prospective partners for copulation.[273] These males are usually in sight of each other, but when they are only in earshot, it is called a dispersed lek or an exploded lek. Mating usually occurs on the display area.[274]
- lore
- Adj. form: loreal. The topographical region of a bird's head between the eye and the beak on the sides of the head.[96]
- lower mandible
- Also, mandible. The lower part of a bird's bill or beak, roughly corresponding to the lower jaw of mammals, it is supported by a bone known as the inferior maxillary bone—a compound bone composed of two distinct ossified pieces. These ossified plates (or rami), which can be U-shaped or V-shaped,[32] join distally (the exact location of the joint depends on the species) but are separated proximally, attaching on either side of the head to the quadrate bone. The jaw muscles, which allow the bird to close its beak, attach to the proximal end of the lower mandible and to the bird's skull.[34] The muscles that depress the lower mandible are usually weak, except in a few birds such as the starlings (and the extinct Huia), which have well-developed digastric muscles that aid in foraging by prying or gaping actions.[275] In most birds, these muscles are relatively small as compared to the jaw muscles of similarly sized mammals.[276] The outer surface is covered in thin horny sheath of keratin called rhamphotheca,[33][34] specially called gnathotheca in the lower mandible.[35] Compare: upper mandible.
M[edit]
- maintenance behaviour
- Also, comfort behaviour. The name given to any behaviour or activity which a bird uses to maintain its plumage and soft parts: preening, bathing, anting, sunbathing, stretching, scratching, and other such activities.[277][278] Roosting is also often considered to be a comfort behaviour.[279] The phrase is sometimes used more inclusively, to describe all life-sustaining activities, such as feeding, drinking, predator avoidance, and locomotion,[280] and even involuntary biological processes, like moulting.[281]
- mandible
- In birds, the word mandible, alone, usually refers to the lower mandible.
- mantle
- The forward area of a bird's upper side sandwiched between the nape and the start of the back. However, in gulls and terns, the term is often used to refer to much of the upper surface below the nape.[27]
- migration
- Also defined: partial migration. The regular seasonal movement, often north and south, undertaken by many species of birds. Bird movements include those made in response to changes in food availability, habitat, or weather. Sometimes journeys are not termed "true migration" because they are irregular (nomadism, invasions, irruptions) or only in one direction (dispersal, movement of young away from natal areas). Migration is marked by its annual seasonality.[282][283] Non-migratory birds are said to be resident or sedentary. Approximately 1,800 of the world's bird species are long-distance migrants.[284][285] Many bird populations migrate long distances along a flyway. The most common pattern involves flying north in the northern spring to breed in the temperate or Arctic summer, and returning in the autumn to wintering grounds in warmer regions to the south. In the southern hemisphere the directions are reversed, but there is less land area in the far south to support long-distance migration.[286] Not all populations within a species may be migratory; this is known as "partial migration". Partial migration is very common in birds of the southern continents. For example, in Australia, 44% of non-passerine birds and 32% of passerine species are partially migratory.[287] Many, if not most birds migrate in flocks. For larger birds, flying in flocks reduces the energy cost, e.g., geese in a V-formation may conserve 12–20% of the energy they would otherwise need if flying alone.[288][289]
- morph
- Also, colour/color morph. Polymorphic variance in the colouring of the plumage between individuals of the same species, unrelated to age, sex or season. For example, the snow goose has two plumage morphs, white (snow) or grey/blue (blue), thus the common description of individuals as either "snows" or "blues". At one time this colour morph resulted in the "blue goose" being classified as a separate species.[290][291]
- moult
- Also, molt (chiefly U.S.) and moulting / molting. The periodic replacement of feathers by shedding old feathers while producing new ones. Feathers are dead structures at maturity which are gradually abraded and need to be replaced. Adult birds moult at least once a year, although many moult twice and a few three times each year. It is generally a slow process, as birds rarely shed all their feathers at any one time; the bird must retain sufficient feathers to regulate its body temperature and repel moisture. The number and area of feathers that are shed varies. In some moulting periods, a bird may renew only the feathers on the head and body, shedding the wing and tail feathers during a later moulting period. Some species of bird become flightless during an annual "wing moult" and must seek a protected habitat with a reliable food supply during that time. Typically, a bird begins to shed some old feathers, then pin feathers grow in to replace the old feathers. As the pin feathers become full feathers, other feathers are shed. This is a cyclical process that occurs in many phases. It is usually symmetrical, with feather loss equal on each side of the body. Because feathers make up 4–12% of a bird's body weight, it takes a large amount of energy to replace them. For this reason, moults often occur immediately after the breeding season, but while food is still abundant. The plumage produced during this time is called postnuptial plumage.[292]
- moult strategy
- Specialized types defined: simple basic strategy; simple alternate strategy; complex basic strategy; complex alternate strategy. The pattern of moults that occurs regularly based on the season and time. There are four main moulting strategies: i) simple basic strategy, in which there is one moult per year, every year; ii) simple alternate strategy, in which there are two moults per year, every year; iii) complex basic strategy, in which there is one moult per year, except in the first year of life, where there is an additional moult; and iv) complex alternate strategy, in which there are two moults per year, except in the first year of life, where there is an additional moult.[293]
- moustachial stripe
- Also, mustache stripe; whisker stripe; malar stripe. A dark feather colouration line running "obliquely from the gape down and along the lower border of the ear-coverts". Compare: submoustachial stripe and lateral throat-stripe.[27] Birds with moustachial stripes include some of the pipits as well as buntings.[27]


N[edit]
- nail
- A plate of hard horny tissue at the tip of the beak of all birds in the family Anatidae (ducks, geese and swans).[294] This shield-shaped structure, which sometimes spans the entire width of the beak, is often bent at the tip to form a hook.[295] It serves different purposes depending on the bird's primary food source. Most species use their nails to dig seeds out of mud or vegetation,[296] while diving ducks use theirs to pry molluscs from rocks.[297] There is evidence that the nail may help a bird to grasp things; species which use strong grasping motions to secure their food (such as when catching and holding onto a large squirming frog) have very wide nails.[298] Certain types of mechanoreceptors, nerve cells that are sensitive to pressure, vibration or touch, are located under the nail.[299]
- nares
- The two holes—circular, oval or slit-like in shape—which lead to the nasal cavities within the bird's skull, and thus to the rest of the respiratory system.[37] In most bird species, the nares are located in the basal third of the upper mandible. Kiwis are a notable exception; their nares are located at the tip of their bills.[92] A handful of species have no external nares. Cormorants and darters have primitive external nares as nestlings, but these close soon after the birds fledge; adults of these species (and gannets and boobies of all ages, which also lack external nostrils) breathe through their mouths. There is typically a septum made of bone or cartilage that separates the two nares, but in some families (including gulls, cranes and New World vultures), the septum is missing. While the nares are uncovered in most species, they are covered with feathers in a few groups of birds, including grouse and ptarmigans, crows and some woodpeckers.[37]
- nasal canthus
- The area where the eyelids come together at the anterior corner of the eye, on the side of the nares, as opposed to the temporal canthus.[133]
- natal down
- Also, neossoptiles. Also defined: protoptiles; mesoptiles. The layer of down feathers that cover most birds at some point in their early development. Precocial nestlings are already covered with a layer of down when they hatch, while altricial nestlings develop their down layer within days or weeks of hatching. Megapode hatchlings are the sole exception; they are already covered with contour feathers when they hatch.[300] The natal down coat is usually lost within a week or two of hatching.[301] There are two different kinds of natal down feathers; protoptiles and mesoptiles.[302] The former appears first[303] and the latter appears second.[304] Hathchlings born naked or with a diffuse coat of scant natal down feathers are called psilopaedic, while those born covered with a dense fuzz of natal down are termed ptilopaedic.[301] Compare: body down and powder down.
- nest
- Also, bird nest. Specialized types: burrow; cavity; cup (adherent cup; statant cup); dome; dormitory; eyries (or aeries); hanging; ledge; mound; pendant; platform; roost (or winter-nest); scrape; saucer or plate; and sphere. The spot in which a bird lays and incubates its eggs and raises its young. Although the term popularly refers to a specific structure made by the bird itself—such as the grassy cup nest of the American robin or Eurasian blackbird, or the elaborately woven hanging nest of the Montezuma oropendola or the village weaver—for some species, a nest is simply a shallow depression made in sand; for others, it is the knot-hole left by a broken branch, a burrow dug into the ground, a chamber drilled into a tree, an enormous rotting pile of vegetation and earth, a shelf made of dried saliva or a mud dome with an entrance tunnel. The smallest bird nests are those of some hummingbirds, tiny cups which can be a mere 2 cm (0.79 in) across and 2–3 cm (0.79–1.18 in) high.[305] At the other extreme, some nest mounds built by the dusky scrubfowl measure more than 11 m (36 ft) in diameter and stand nearly 5 m (16 ft) tall.[306] Not all bird species build nests. Some species lay their eggs directly on the ground or rocky ledges, while brood parasites lay theirs in the nests of other birds, letting unwitting "foster parents" do all the work of rearing the young. Although nests are primarily used for breeding, they may also be reused in the non-breeding season for roosting and some species build special dormitory nests or roost nests (or winter-nest) that are used only for roosting.[307] Most birds build a new nest each year, though some refurbish their old nests.[308] The large eyries (or aeries) of some eagles are platform nests that have been used and refurbished for several years. In most species, the female does most or all of the nest construction, though the male often helps.[309] The nest may form a part of the courtship display, such as in weaver birds.[274]
- nictitating membrane
- Also, third eyelid. A transparent or translucent third eyelid that is drawn across the eye for protection, to moisten it while maintaining vision.[310] The nictitating membrane also covers the eye and acts as a contact lens in many aquatic birds.[311] With the exception of pigeons and a few other species, most birds blink only with their nictitating membrane, and when not sleeping, use the eyelids chiefly only when the eye is threatened with some foreign matter.[312]
- nidicolous
- Young that remain in their nest after hatching for an extended period of time and thus must be fed by their parents.[16] Contrast: nidifugous.
- nidifugous
- Young that leave the nest soon after hatching, joining their parents in foraging.[16] Contrast: nidicolous.
- non-breeding plumage
- See basic plumage.
- notch
- A pronounced narrowing at some variable distance along the feather edges at the outermost primaries of large soaring birds, particularly raptors. Whether these narrowings are called notches or emarginations depends on the degree of their slope.[18] An emargination is a gradual change, and can be found on either side of the feather. A notch is an abrupt change, and is only found on the wider trailing edge of the remiges. The presence of notches and emarginations creates gaps at the wingtip; air is forced through these gaps, increasing the generation of lift.[184]
- nuptial plumage
- See alternate plumage.



O[edit]
- oology
- The scientific study of eggs.[313]
- operculum
- Plural: opercula. A membraneous, horny or cartilaginous flap covering the nares of some birds.[314][315] For example, in diving birds, the operculum keeps water out of the nasal cavity;[314] when the birds dive, the impact force of the water closes the operculum.[316]
- overbrooding
- The not uncommon phenomenon of birds continuing to brood eggs that are not viable and will not hatch, sometimes for lengthy periods of time beyond the normal incubation period.[317]
P[edit]

- passerine
- Also, perching bird. Any bird of the order Passeriformes, which includes more than half of all bird species. A notable feature of passerines compared to other orders of Aves is the arrangement of their toes—three pointing forward and one back—which facilitates perching. Sometimes known as perching birds or, less accurately, as songbirds, the passerines form one of the most diverse terrestrial vertebrate orders, with over 5,000 identified species.[318] The order has roughly twice as many species as Rodentia, the largest order of mammals. There are more than 110 families of passerine birds, the second-most of any tetrapods (after Squamata, the scaled reptiles).
- pectinate claw
- Also, feather comb.[251] A claw on the middle toe of some birds, such as nightjars, herons, and barn owls,[319] with a serrated edge. The degree of serration varies from fine to coarse, depending on the species. It is used in preening, functioning similar to a comb. Some birds may use it to straighten the rictal bristles.[320] It may also play an important role in parasite removal. For example, a study of barn owls found a significant correlation between the number of serrated teeth on an individual's pectinate claw, and the prevalence of its lice infestation.[321]
- pectoral tuft
- Elongated feathers, often brightly coloured, which arise from the sides of the chest of some species of birds, including many sunbirds, spiderhunters and flowerpeckers.[322] They are typically hidden when the wing is folded on a perched bird, but prominent during courtship and territorial displays.[323] In sunbirds and spiderhunters, these tufts may be yellow, red or orange; in flowerpeckers, they are usually white.[322] The size of the tufts in some species can be an indicator of a male's fitness and status; males with larger tufts defend larger territories and have higher reproductive success.[324]
- pennaceous feather
- Also, contour feather. A type of feather present in most modern birds and in some other species of maniraptoriform dinosaurs.[325] Pennaceous feathers have a stalk called a quill, with the basal part called a calamus, that is embedded in the skin exclusively at pterylae (feather tracts).[61] The calamus is hollow and has pith formed from the dry remains of the feather pulp. The calamus stretches between two openings—at its base is the inferior umbilicus and at its distal end is the superior umbilicus; the rachis of the main stem, hosting the vanes, continues above it.[60] The rachides of contour feathers have an umbilical groove on their underside, with the vanes or vexilla, spreading to either side. The vanes comprise many flattened barbs, that are connected to one another by hooklets that are anchored into flanges on proximal barbules on neighboring barbs. Some specialized feathers are considered "modified" contour feathers, such as ear coverts and bird eyelashes called rictal bristles.[61] Contrast: plumulaceous (down) feathers and filoplumes.
- philopatry
- The habit of a bird returning to the area it first bred.[326]
- pigeon milk
- The term for crop milk when found among all pigeons and doves.[129]
- pileum
- As defined in the treatise Manual of Ornithology: Avian Structure and Function, the "entire top of the head, including the forehead, crown and occipital regions."[121]
- pin feather
- Also, blood feather. A developing feather encased in a feather sheath that, unlike a fully developed feather, has a blood supply flowing through it. As such, damage to pin feathers can cause significant bleeding. Pin feathers may be the first feather growth during a bird's infancy, or those growing during moulting at any stage of a bird's life. As pin feather growth progresses, the blood supply is concentrated only in the base of the calamus. Pin feathers begin to develop after the feather bud invaginates a cylinder of epidermal tissue around the base of the dermal papilla, forming the feather follicle. At the base of the feather follicle, epithelial cells proliferate to grow the epidermal collar or cylinder. As the epidermal cylinder extends through dermis, it differentiates into a protective peripheral sheath, longitudinal barb ridges and growth plates. Over time these barb ridges grow in a helical manner, branching to create barbs and barbules, and fusing to form the central feather shaft. Moreover, the barb plate further differentiates into hooklets and cilia, while the marginal and axial plate die to form the intervening space within the feather structure.[327][328][329]
- pinioning
- Also defined: pinion joint. The act of surgically removing one pinion joint—the joint of a bird's wing farthest from the body—thereby stopping the growth of the primary feathers, to prevent flight. The animal welfare impact of pinioning is subject to increasing debate. For example, it is known that the operation, which is often performed without pain relief, is just as painful in young birds as in mature birds, if not more so.[330] Evidence also suggests that pinioning may cause a phantom limb syndrome similar to what is observed in human amputees.[330]
- pinions
- The outermost primaries—those connected to the phalanges.[331]
- plumage
- Plumage (Latin: plūma 'feather') refers both to the layer of feathers that cover a bird and the pattern, colour and arrangement of those feathers.[332] The pattern and colours of plumage differ between species and subspecies, often vary with age and may vary sharply between males and females of the same species that exhibit sexual dimorphism. Within species there can be different colour morphs. Despite the great variance, as broad observations: 1) among many bird species adult males are more brightly coloured than are females, especially when in alternate or nuptial plumage, as opposed to basic or winter plumage; 2) when male birds undergo the prebasic moult and attain their non-breeding plumage, they much more closely resemble females; and 3) the plumage of juvenile birds (of both sexes) tends to be relatively dull and inconspicuous and typically resembles that of adult females.[333]
- plumology
- Also, plumage science. The name for the science that is associated with the study of feathers.[334][335]
- podotheca
- Also, foot sheath. Also defined: boot; reticulate; scutellate; booted. The skin that covers the bare feet and legs. It usually consists of many small scales, called scutes. This type of podotheca is described as being scutellate. When there are no scutes, and the podotheca is a smooth sheath, it is called a boot, and those with podotheca like this are called booted. The podotheca may also be reticulated with small and irregularly raised plates. This arrangement is considered to be reticulate.[336]
- powder down
- Also, pulviplumes; feather dust. A special type of down that occurs in a few groups of apparently unrelated birds. In some species, the tips of the barbules on powder down feathers disintegrate, forming fine particles of keratin, which appear as a powder, or feather dust, among the feathers. These feathers grow continuously and are not moulted.[337] In other species, the powder grains come from cells that surround the barbules of growing feathers.[338] These specialized feathers are typically scattered among ordinary down feathers, though in some species, they occur in clusters.[154] All parrots have powder down, with some species (including the mealy parrot) producing copious amounts.[339] It is also found in tinamous and herons.[154] The dust produced from powder down feathers is a known allergen in humans.[340] Compare: body down & natal down.
- postjuvenal moult
- See prebasic moult.
- postnuptial moult
- See prebasic moult.
- prealternate moult
- Also, prenuptial moult (sp. variation: in the U.S., sometimes molt). The moult before the breeding season that most birds undergo, during which parts of the basic plumage are shed and nuptial plumage is grown. "Prealternate moult" is used in Humphrey–Parkes terminology, and "prenuptial moult" is the traditional term. In either system of nomenclature, in juveniles this type of moult is numbered, 1st, 2nd, 3rd (etc.), until the definitive plumage is attained, after which the numbering is dropped.[341] Unlike the prebasic moult, in the prealternate moult the major flight feathers and primary coverts are not replaced in most species, and the feathers involved typically are "only head and body feathers, wing coverts and sometimes tertials and rectrices".[342]
- prebasic moult
- Also, postnuptial moult (sp. variation: in the U.S., sometimes molt). Also defined: postjuvenal moult. The moult after the breeding season that most birds undergo, during which nuptial plumage is shed and basic or winter plumage is grown. In Humphrey–Parkes terminology, prebasic moults in juveniles, prior to attaining definitive plumage, are numbered the 1st, 2nd, 3rd (etc.) prebasic moult. In traditional terminology, the first is called the postjuvenal moult, and thereafter the 1st, 2nd, 3rd (etc.) postnuptial moult. In both nomenclature systems, after definitive plumage is reached, the numbering is dropped.[341]
- precocial
- Also defined: semi-precocial; altricial-precocial spectrum. Young that, at hatching, have their eyes open; are covered in down feathers (ptilopaedic); are homeothermic;[11] and are able to leave the nest soon after hatching and join their parents in foraging activities (nidifugous). The contrasting state is altricial young, which are born "helpless"—more or less naked, blind, ectothermic and unable to leave the nest.[13] The young of many bird species do not precisely fit into either the precocial or altricial category, having some aspects of each and thus fall somewhere on an altricial-precocial spectrum.[14] A defined intermediate state is termed semi-precocial, typified by young born covered in down and with open eyes; that are usually able to walk shortly after hatching; but which remain mostly confined to the nest, relying on their parents for food (nidicolous).[16][343]
- preening
- Also, grooming. Feathers require maintenance and birds preen or groom them daily, spending an average of around 9% of their daily time on this activity.[344] The bill is used to brush away foreign particles and to apply waxy secretions from the uropygial gland; these secretions protect the feathers' flexibility and act as an antimicrobial agent, inhibiting the growth of feather-degrading bacteria.[345] This may be supplemented with the secretions of formic acid from ants, which birds receive through a behaviour known as anting, to remove feather parasites.[346]
- prenuptial moult
- See prealternate moult.
- primaries
- Also, primary feathers; primary remiges. A type of remex flight feather, they are connected to the manus (the bird's "hand", composed of carpometacarpus and phalanges); these are the longest and narrowest of the remiges (particularly those attached to the phalanges), and they can be individually rotated. These feathers are especially important for flapping flight, as they are the principal source of thrust, moving the bird forward through the air. Most thrust is generated on the downstroke of flapping flight. However, on the upstroke (when the bird often draws its wing in close to its body), the primaries are separated and rotated, reducing air resistance while still helping to provide some thrust.[347] The flexibility of the remiges on the wingtips of large soaring birds also allows for the spreading of those feathers, which helps to reduce the creation of wingtip vortices, thereby reducing drag.[348] In most flighted birds, some of the distal barbules on the inner vane of these feathers, called friction barbules, are specialized, with large lobular barbicels that are thought to help grip and prevent slippage of overlying feathers.[349] Species vary in the number of primaries they possess. The number in non-passerines generally varies between 9 and 11,[350] but grebes, storks and flamingos have 12 and ostriches have 16.[351] While most modern passerines have ten primaries,[350] some have only nine. Those with nine are missing the most distal primary, sometimes called the remicle, which is typically very small and sometimes rudimentary in passerines.[351]
- primary projection
- Also, primary extension. The distance that a bird's longest primaries extend beyond its longest secondaries (or tertials) when its wings are folded.[352] As with wing formulae, this measurement is useful for distinguishing between similarly plumaged birds; however, unlike wing formulae, it is not necessary to have the bird in-hand to make the measurement. Rather, this is a useful relative measurement—some species have long primary extensions, while others have shorter ones. Among the Empidonax flycatchers of the Americas, for example, the dusky flycatcher has a much shorter primary extension than does the very similarly plumaged Hammond's flycatcher.[352] Europe's common skylark has a long primary projection, while that of the near-lookalike Oriental skylark is very short.[353] As a general rule, species which are long distance migrants will have longer primary projection than similar species which do not migrate or migrate shorter distances.[354]
- proventriculus
- The first part of the stomach of a bird. The inside is lined with gastric glands, which secrete gastric juices containing enzymes and hydrochloric acid. In some birds, such as petrels, the proventriculus is expandable. This allows these birds to either digest their food later or carry it back to their young. The proventriculus leads into the gizzard.[355]
- psilopaedic
- Young that are born naked or with only a small amount of down feather coverage, as opposed to birds that are ptilopaedic.[16]
- pterylae
- Singular: pteryla. The feather tracts of a bird's skin from which the contour feathers grow, often in sharply defined and dense clusters—as opposed to the apterylae.[23] See related: pterylosis.
- pterylosis
- Also, pterylography. The arrangement of feather tracts, as seen in a bird's pterylae and apterylae, which varies across bird families and has been used in the past as a means for determining the evolutionary relationships among them.[356][357]
- ptilopaedic
- Young that are born covered with down feathers, as opposed to birds that are psilopaedic.[16]
- pupil
- The dark disk at the centre of a bird's eye through which light enters, surrounded by the coloured outer ring of the iris.[358]
- pygostyle
- Also, tail bone. A skeletal formation, plowshare-shaped in birds, in which the final few caudal vertebrae are fused into a single ossification, supporting the tail feathers and musculature. In modern birds, the rectrices attach to these. The pygostyle is the main component of the uropygium.[359][360]



Q[edit]
- quartering
- A hunting technique where a bird flies slowly just above the water or ground in open habitats.[361]
- quill
- Also, primary quill; main stem; scapus. The main stem of a feather from which all structures branch, if any. The proximal portion is called the calamus or shaft, and the distal portion is called the rachis, with the demarcation point between them usually defined as the superior umbilicus.[61][362] However, some authorities define quill very differently, using it as a synonym for the calamus.[363] The contradiction between sources on this issue is quite sharp. For example, in Asa Chandler's well known treatise, A study of the structure of feathers, with reference to their taxonomic significance, "quill" is defined at page 250 as: "The main stem of a feather, including both shaft and calamus (Coues, 1884; Beebe, 1906, et al.). Synonyms: main stem (Nitzsche, 1867)".[61] By contrast, in Frank B. Gill's well known treatise, Ornithology, he writes at page 80: "The hollow base of the shaft—the calamus, or quill—anchors the feather in a follicle below the surface of the skin".[363] In this glossary, the former definition will be used, treating quill as denoting the "main stem" of a feather.
R[edit]
- rachis
- Plural: rachises; rachides. Also, shaft. Variable sp.: rhachis.[364] Also defined: medulla. The distal or upper section of the quill, above the calamus, stretching from the superior umbilicus to the tip of the feather, upon which the vanes and other structures are anchored. The rachis is flexible from side to side toward the blade projections, but far stiffer inward and outward from the bird's body. This arrangement aids in allowing the flight feather to act as a "resilient airfoil".[362] The rachis is composed of two layers: a pithy, opaque material makes up the core layer called the medulla, which is covered by a thin and translucent outer layer or cortex that features longitudinal internal ridges on the dorsal side and external ridges on the ventral side.[8]
- rectrices
- Singular: rectrix. From the Latin for 'helmsman', they are the long, stiff, asymmetrically shaped, but symmetrically paired pennaceous feathers on the tail of a bird, which help it to brake and steer in flight. They lie in a single horizontal row on the rear margin of the anatomic tail. The vast majority of species having six pairs. They are absent in grebes and some ratites, and greatly reduced in size in penguins.[351][365][366] Many grouse species have more than 12 rectrices. Some species (including ruffed grouse, hazel grouse and common snipe) have a number that varies among individuals.[367] Domestic pigeons have a highly variable number, due to centuries of selective breeding.[368]
- remiges
- Singular: remex. Also defined: postpatagium. From the Latin for 'oarsman', they are the long, stiff, asymmetrically shaped, but symmetrically paired pennaceous feathers on the wings of a bird. They are located on the posterior side of the wing. Ligaments attach the long calami firmly to the wing bones, and a thick, strong band of tendinous tissue known as the postpatagium helps to hold and support the remiges in place.[369] Corresponding remiges on individual birds are symmetrical between the two wings, matching to a large extent in size and shape (except in the case of mutation or damage), though not necessarily in pattern.[370][371] They are given different names depending on their position along the wing.
- remicle
- Also, little remex. The most distal primary feather. The remicle is very small and usually non-functional—interpreted by Christian Ludwig Nitzsch as "an aborted first primary". It was named by Richard S. Wray in 1887, who similarly thought of it as "representing an originally functional primary".[372]
- resident
- Also, permanent resident; sedentary. Non-migratory; a bird that stays year-round and breeds in one geographic area or habitat.[373] Both permanent resident and sedentary are common with the defined usage. Although resident is used in this sense, it is often used as well for birds that are "migratory in part of their breeding range and resident elsewhere",[374] e.g., "Eurasian blackcaps (Sylvia atricapilla) are summer visitors to Eastern and Northern Europe, resident in Western Europe and winter visitors to Africa."
- rictal bristles
- Also, bristles; bristle feathers; eyelashes. Stiff, hair-like, tapering feathers with a large rachis but few barbs found around the eyes and the base of the beak of some bird species.[375] They are common among insectivorous birds, but are also found in some non-insectivorous species.[376] They may serve a similar purpose to eyelashes and vibrissae in mammals. Although there is as yet no clear evidence, it has been suggested that rictal bristles have sensory functions and may help insectivorous birds to capture prey.[375][377] There is some experimental evidence to suggest that bristles may prevent particles from striking the eyes if, for example, a prey item is missed or breaks apart upon contact.[376]
- rosette
- Also, gape rosette. A fleshy rosette found at the corners of the beaks of some birds, such as the puffin.[378] In the puffin, this is grown as part of its display plumage.[379]
- rhamphotheca
- The outer surface of the beak consisting of a thin horny sheath of keratin,[33][34] which can be subdivided into the rhinotheca of the upper mandible and the gnathotheca of the lower mandible.[35] This covering arises from the Malpighian layer of the bird's epidermis,[35] growing from plates at the base of each mandible.[380] There is a vascular layer between the rhamphotheca and the deeper layers of the dermis, which is attached directly to the periosteum of the bones of the beak.[381] The rhamphotheca grows continuously in most birds, and in some species, the colour varies seasonally.[382]
- rhinotheca
- The rhamphotheca (thin horny sheath of keratin) covering the upper mandible.[33][34][35]
- rump
- Also, uropygium; uropygial region; parson's nose, pope's nose; sultan's nose. Topographically, the region of a bird's upperparts between the end of the back and the base of the tail.[383] Anatomically, the fleshy protuberance visible at the posterior end of a bird. Its swollen appearance results from it housing the uropygial gland.[384] See related: pygostyle.

Resident year-round
Winter visitor
Range map for Eurasian blackcaps—corresponding to the example use of "resident" in its definition at the left

S[edit]
- scales
- Specialized types: cancella; scutella; scutes. Also defined: reticulae; acrometatarsium / acrotarsium. The scales of birds are composed of keratin like their beaks and claws, and are found mainly on the toes and metatarsus, though they may appear further up on the ankle. They do not overlap significantly, except in the cases of kingfishers and woodpeckers. The scales and scutes of birds were originally thought to be homologous to those of reptiles and mammals;[385] later research, however, suggests that scales in birds re-evolved after the evolution of feathers.[386][387] Bird embryos begin development with smooth skin. On the feet, the corneum, or outermost layer of this skin may keratinize, thicken and form scales. Bird scales can be organized into:
i) cancella—minute scales which are really just a thickening and hardening of the skin, crisscrossed with shallow grooves;[388]
ii) scutella—scales that are not quite as large as scutes, such as those found on the caudal, or hind part, of the chicken metatarsus;[388] and
iii) scutes—the largest scales, usually on the anterior surface of the metatarsus and dorsal surface of the toes. The rows of scutes on the anterior of the metatarsus is sometimes termed an "acrometatarsium" or "acrotarsium".[388]
Reticulae, by contrast, are small, keeled scale-like structures covering the bottoms of bird feet, but are not true scales. Evolutionary developmental studies on reticulae found that they are composed entirely of alpha-keratin (whereas, true epidermal scales are composed of a mix of alpha and beta keratin).[387] This, along with their unique structure, has led to the suggestion that reticulae are actually feather buds that were arrested early in development.[92][387][389] See also: podotheca. - scapulars
- Also, humeral region. Feathers covering a bird's scapula "at the base of the dorsal wing".[390]
- secondaries
- Also, secondary feathers; secondary remiges. A type of remex flight feather, they are connected to the ulna. In some but not all bird species, the ligaments that bind secondaries to the bone connect to small, rounded projections that are called quill knobs. Secondary feathers remain close together in flight (they cannot be individually separated like the primaries can) and help to provide lift by creating the airfoil shape of the bird's wing. Secondaries tend to be shorter and broader than primaries, with blunter ends. They vary in number from six in hummingbirds, to as many as 40 in some species of albatross. In general, larger and longer-winged species have a larger number of secondaries.[51] Birds in more than 40 non-passerine families are missing the fifth secondary feather on each wing; a state known as diastataxis.[18]
- semiplume
- Also, semiplume feather. A type of feather that has features straddling the differences between true down and contour feathers, such as by lacking cross-attachment between their barbs (i.e., without a flange–hooklet anchoring system), but having a central, full-length shaft as in contour feathers. They are usually found on the margins of the pennaceous feather tracts and, like down, provide insulative qualities and help fill out the plumage. Crest feathers are a type of semiplume.[160][391][392]
- sexual dimorphism
- The common phenomenon amongst birds in which males and females of the same species exhibit different characteristics beyond the differences in their sexual organs. It can manifest in size or plumage differences. Sexual size dimorphism varies among taxa with males typically being larger, though this is not always the case, i.e., in birds of prey, hummingbirds and some species of flightless birds.[393][394] Plumage dimorphism, in the form of ornamentation or colouration, also varies, though males are typically the more ornamented or brightly coloured sex.[395] Such differences have been attributed to the unequal reproductive contributions of the sexes.[396]
- shaft
- See entry for rachis.
- sides
- The topographical area between the breast and the back, stretching from the axilla (underarm) to the Iliac crest.[397]
- song
- A type of bird vocalization associated with courtship and mating, tending to be longer and more complex than a bird's call, which serve such functions as giving alarm or keeping members of a flock in contact.[63]
- speculum
- Also, speculum feathers; mirror. A patch of usually brightly coloured feathers, often iridescent, on the inner secondary remiges in a number of duck species,[398] though other birds may have them, such as the bright red or orange wing speculum of several parrots from the genus Amazona.[399]
- spur
- Outgrowths of bone covered in a sheath of horn which are found on some birds. Spurs are most commonly found on the feet or legs, though some birds possess spurs on the leading edge of the wings.[400]
- sternum
- Plural: sterna. The breastbone of a bird. There are two types: i) carinate sterna—appearing in flighted birds, in which the ventral surface is keel-shaped, which provides ample surface area for attachment of wing muscles used for flight; and ii) ratite sterna—appearing in flightless birds, such as rhea, in which the ventral surface is flattened.[401]
- submoustachial stripe
- A usually light stripe of colour between the moustachial stripe above, and the lateral throat-stripe below.[27]
- supercilium
- Plural: supercilia. Also, superciliary line; eyebrow / eye brow. A pale line appearing above the eye of a bird.[97] Compare eye stripe.
- superior umbilicus
- Also, distal umbilicus. A very small opening on the ventral side (facing toward the bird's skin) of a feather's shaft, marking the distal end of the calamus and the start of the rachis. It is a remnant of the epithelial tube used by a bird's body to construct the feather.[189] Compare: inferior umbilicus.
- supraloral
- Where a stripe is present only above the lores, and does not continue behind the eye, it is called a supraloral stripe or simply supraloral, rather than the supercilium.[402]
- syrinx
- From the Greek word for pan pipes, σύριγξ, it is the vocal organ of birds. Located at the base of a bird's trachea, it produces sounds without the vocal folds of mammals.[403] As air flows through the syrinx, sound is produced by the vibration of some or all of the membrana tympaniformis (the walls of the syrinx) and a bar of cartilage connecting the dorsal and ventral extremities, known as the pessulus. This sets up a self-oscillating system that regulates the airflow used to create vocalizations. Sound modulation is achieved by varying the tension of muscles connected to the membranes and bronchial openings involved.[404] The syrinx enables some species of birds (such as parrots, crows and mynas) to mimic human speech. Unlike the mammalian larynx, the syrinx is located where the trachea forks into the lungs. Thus, lateralization of bird song is possible, and some songbirds can produce more than one sound at a time.[405]



T[edit]
- tail
- While the underlying structure of the tail is made up of bone (the pygostyle[406]) and flesh, the term "tail" is most commonly used to refer to the feathers growing from the region and the shape they form—that is, undertail and uppertail coverts and rectrices, commonly radiating in a fan configuration.[125]
- tail coverts
- See: undertail coverts, uppertail coverts and crissum.
- tail streamers
- The elongated, narrow tips of the tail seen in some birds.[407] They generally function as a sexual ornament.[408] In the barn swallow, this has resulted in tail streamers about 12 mm (0.47 in) longer than is aerodynamically optimal.[409]
- talon
- The claw of a bird of prey; its primary hunting tool.[410] The talons are very important; without them, most birds of prey would not be able to catch their food. Some birds also use claws for defensive purposes. Cassowaries use claws on their inner toe (digit II) for defence, and have been known to disembowel people. All birds, however, have claws, which are used as general holdfasts and protection for the tip of the digits. The hoatzin and turaco are unique among extant birds in having functional claws on the thumb and index finger (digit I and II) on the forelimbs as chicks, allowing them to climb trees until the adult plumage with flight feathers develop.[411][412] However, several birds have a claw- or nail-like structure hidden under the feathers at the end of the hand digits, notably ostriches, emus, ducks, geese and kiwis.[413]
- tarsus
- Also defined: tarsometatarsus. The third and most conspicuous portion of the bird's leg, from which the toes spring; the foot.[414] In birds, the true tarsus has disappeared, with the proximal tarsals having fused with the tibia, the centralia having disappeared, and the distal bones having fused with the metatarsals to form a single tarsometatarsus bone, effectively giving the leg a third segment.[415]
- teleoptiles
- The collective term for all of the feathers of an adult bird.[416]
- temporal canthus
- The area where the eyelids come together at the posterior corner of the eye (toward the sides of the head), as opposed to the nasal canthus.[133]
- tertiaries
- Also, tertials; tertiary feathers; humeral feathers. A type of feather arising in the brachial region, i.e., "proximal to the innermost secondaries", usually growing in a grouping of three to four feathers.[417] They are not considered true remiges as they are not supported by attachment to the corresponding bone—in this case the humerus. These elongated "true" tertials act as a protective cover for all or part of the folded primaries and secondaries, and do not qualify as flight feathers as such.[418] However, many authorities use the term tertials to refer to the shorter, more symmetrical innermost secondaries of passerines (arising from the olecranon and performing the same function as true tertials) in an effort to distinguish them from other secondaries. The term humeral is sometimes used for birds such as the albatrosses and pelicans that have a long humerus.[419][420]
- throat
- The triangular area of a bird's external anatomy, typically feathered, located between the chin and the upper part of the breast.[97]
- thigh
- The topographical area between the "knee" and the trunk of the body.[421]
- tibia
- The usually feathered part of a bird's leg extending above the foot (tarsus).[422]
- tomia
- Singular: tomium. The cutting edges of the upper and lower mandibles.[36] In most birds, these range from rounded to slightly sharp, but some species have evolved structural modifications that allow them to handle their typical food sources better.[92] Granivorous (seed-eating) birds, for example, have ridges in their tomia, which help the bird to slice through a seed's outer hull.[423] Birds in roughly 30 families have tomia lined with tight bunches of very short bristles along their entire length. Most of these species are either insectivores (preferring hard-shelled prey) or snail eaters, and the brush-like projections may help to increase the coefficient of friction between the mandibles, thereby improving the bird's ability to hold hard prey items.[424] Serrations on hummingbird bills, found in 23% of all hummingbird genera, may perform a similar function, allowing the birds to effectively hold insect prey. They may also allow shorter-billed hummingbirds to function as nectar thieves, as they can more effectively hold and cut through long or waxy flower corollas.[425] In some cases, the colour of a bird's tomia can help to distinguish between similar species. The snow goose, for example, has a reddish-pink bill with black tomia, while the whole beak of the similar Ross's goose is pinkish-red, without darker tomia.[426]
- topography
- The mapping of external features of bird anatomy.[383]
- torpor
- Also, hypometabolism. An energy-conserving strategy used by some small birds [under 100 g (3.5 oz)] in which they become inactive and unreactive to external stimuli, and reduce their metabolism below their normal respiration and heart rate, causing decreases in body temperature by an average of 4–35 °C (39–95 °F). Two types of torpor are recognized. In one type, such as in hummingbirds, torpor may be entered on a daily basis and last only a few hours; other behaviours, such as foraging, continue during non-torpor periods. For other species, torpor may only be entered during cold conditions or when food becomes limited, but may persist for weeks or even months. The extent of unresponsiveness during torpor can be pronounced. A hummingbird in deep torpor, for example, with a body temperature of 18 °C (64 °F), will not respond to a variety of external stimuli, such as attempts to push it from a perch; only the locking reflex of the feet stops the bird from falling. Torpor has been reported in eight orders of birds.[427][428]



U[edit]
- undertail coverts
- Covert feathers covering the base of the tail on its underside.[125] Compare: uppertail coverts; contrast wing coverts.
- underwing
- The bottom side of a bird's wing; the hidden surface of the wing when it is folded.[429] Compare: upperwing.
- upper mandible
- Also, maxilla. The upper part of a bird's bill or beak, roughly corresponding to the upper jaw of mammals, it is supported by a three-pronged bone called the intermaxillary. The upper prong of this bone is embedded into the forehead, while the two lower prongs attach to the sides of the skull. At the base of the upper mandible a thin sheet of nasal bones is attached to the skull at the nasofrontal hinge, which gives mobility to the upper mandible, allowing it to move upwards and downwards.[33] The outer surface is covered in a thin, horny sheath of keratin called rhamphotheca[33][34] specially called rhinotheca in the upper mandible.[35] Compare: lower mandible.
- upper parts
- А также верхние части и верхние части . Топографические спину глазами птицы (обычно не включая глазах , если таковая имеется) и всю верхнюю поверхность птицы, включая « макушку , и полосу области над , третичные на внутренние кроющие крыла », [430] хотя этот термин иногда используется более широко, для всех особенностей, от клюва до хвоста. [431] Сравните: под частями .
- под частями
- А также нижние части ; нижняя часть тела ; нижние части ; нижние части ; нижние части . Все топографические области на нижней стороне или «под поверхностью» птицы, [431] т. е. от подбородка до нижнего конца хвоста (на туловище и, следовательно, не включая ноги). Сравните верхние части .
- надхвостовые кроющие
- Скрытые перья покрывают основание хвоста с верхней стороны. [125] Иногда эти кроющие более специализированы. «Хвост» павлина на самом деле представляет собой очень удлиненные надхвостовые кроющие. [117] Сравните кроющие под хвостом ; контрастные кроющие крыльев .
- верхнее крыло
- Верхняя или верхняя сторона птичьего крыла. [432] Сравните: подкрылок .
- уропигиальная железа
- Кроме того, чистите железу ; сальная железа . Железа, расположенная на крупе , у основания хвоста , которая производит восковой секрет, состоящий из масел, жирных кислот, жиров и воды, который большинство птиц используют для ежедневной чистки своих перьев. Хотя перья периодически линяют , они представляют собой инертные структуры без системы питания и быстро портятся без такого применения во время чистки. Нанесенный восковой секрет способствует водонепроницаемости, помогает поддерживать влажность перьев, предохраняя их от ломкости и гибкости, а также, как считается, способствует росту непатогенных грибов, отпугивает перьевых вшей и некоторых форм грибков и бактерий, поедающих кератин. Железа представляет собой двудольную структуру и обычно окружена пучком пушистых перьев, которые могут действовать как фитиль для выделяемого масла. [433]
- уропигий
- См . крупу .

V [ edit ]
- бродяга
- Описание птицы, обитающей за пределами обычного ареала или ареала миграции ее вида или подвида. [434]
- у них есть
- Кроме того, vexillum – множественное число: vexilla . Паутинное пространство гибких бородок, более или менее соединенных между собой крючками , простирающееся бородок с каждой стороны дистальной части стержня пера, известное как рахис . Одна сторона лопасти, называемая внешней лопастью, перекрывает внутреннюю сторону лопасти рядом с ней. [8]
- черт возьми
- Общий термин для всех бородок, отходящих от ветви пера бородки . [28]
- лопастное перо
- Классификационный термин для перистых перьев (также известных как контурные перья), покрывающих тело птицы снаружи. [435] к лопастному свойству таких перьев, то есть к наличию уплощенной паутины из гибких бородок , более или менее соединенных между собой крючками бородок , Это относится идущих с каждой стороны дистальной части стержня пера, известной как рахис . Они составляют один из трех широких классов перьев, остальные представляют собой пуховые перья (которые лежат под лопастными перьями, если они есть) и волосовидные нитевидные перья . [8]
- вентиляционное отверстие
- Наружное отверстие клоаки . [436]
- вентиляционное клюнье
- Аномальное поведение птиц в неволе, проявляемое преимущественно товарными курами-несушками , характеризующееся клевыми повреждениями клоаки , окружающей кожи и подлежащих тканей. [437] Клевание клоаки часто происходит сразу после откладки яйца, когда клоака часто остается частично вывернутой, обнажая слизистую оболочку . [438]
В [ править ]
- плетень
- А еще подвес ; лапта ; палеария . Специализированный тип: риктальный плетень . Разновидность мясца , плетень представляет собой лоскут или лоскут плоти, часто яркого цвета, который свисает с головы некоторых птиц, часто с подбородка или горла. [439] [440] Если их найти в углу рта, их можно назвать риктальными сережками , например, у чибисов в масках . [441] К птицам с сережками относятся индейки , цесарки , северные казуары , чернолицые ибисы , кокакос Северного острова , плетистые скворцы (самцы в брачном оперении ) и самцы бородатых колокольчиков , причем последние имеют поразительные, похожие на бороду сережки. [442] [443]
- свистящее
- Поведение в полете, при котором птица быстро опускается зигзагообразными движениями в сторону . Во время свиста некоторые птицы меняют аэродинамическую механику своего тела, которая обычно обеспечивает подъемную силу — летая, переворачивая тело вверх тормашками, но поворачивая шею на 180 градусов, чтобы удерживать голову в нормальном полетном положении, — что приводит к быстрому падению перед быстрым падением. остановка падения путем возобновления нормальной ориентации в полете. [444] [445] [446]
- крыло бар
- Полоса на верхнем крыле образована контрастом кончиков основных и вторичных кроющих. [447] Эта особенность может исчезнуть, когда птица линяет и приобретает другое сезонное оперение . [448] Крыльевые перемычки часто встречаются парами. [449] Контраст: зеркало .
- обрезка крыльев
- А также обрезка крыльев . птицы, Спорная практика обрезки основных перьев крыльев или ремиджей чтобы она не была полностью способна к полету, пока у нее не вырастут сменные перья во время следующей линьки . [450] Если все выполнено правильно, то это безболезненная процедура. [451] Тем не менее, эта практика может привести к травмам, например, кровотечению в результате кровавых перьев . случайной обрезки [452]
- кроющие крыла
- Также определены: кроющие верхних крыльев ; подкрыльевые кроющие ; вторичные кроющие ; первичные кроющие ; большие ( первичные / вторичные ) кроющие ; срединные ( первично- / вторично- ) кроющие ; меньшие ( первичные / вторичные ) кроющие (последние также называются луковыми кроющими ).
скрытые перья птицы имеются На крыле . Существует ряд классов и подклассов. Верхние кроющие крыла делятся на две группы: те, что на внутреннем крыле , которые перекрывают вторичные маховые перья , известны как вторичные кроющие, и те, что на внешнем крыле , которые перекрывают первичные маховые перья , известны как первичные кроющие. Внутри каждой группы перья образуют несколько рядов. Перья самого дальнего, самого большого ряда называются большими (первичными/вторичными) кроющими; те, что в следующем ряду, являются срединными (первичными/вторичными) кроющими, а все оставшиеся ряды называются меньшими (первичными/вторичными) кроющими. Подкрылье . имеет соответствующие наборы кроющих (для обозначения соответствующих наборов используются названия «верхние кроющие» и «нижние кроющие») Кроме того, передний край крыла покрыт группой перьев, называемых краевыми кроющими. Внутри каждой группы кроющих крыльев ряды перьев перекрывают друг друга, как черепица (на большие кроющие перекрываются средние кроющие, которые, в свою очередь, перекрываются крайним рядом меньших кроющих и так далее). [17] См. также: подмышечные ; кроющие ушей ; подкладка крыла (краевые кроющие). - формула крыла
- Математическая классификация формы дистального конца птичьего крыла. Его можно использовать, чтобы помочь различать виды со схожим оперением , и поэтому он особенно полезен для тех, кто кольцевает (кольцевает) птиц. Чтобы определить формулу крыла птицы, открытое расстояние между кончиком самого дистального основного пера и кончиком его большого кроющего пера (самого длинного из перьев, которые покрывают и защищают стержень этого основного пера) измеряется в миллиметрах. В некоторых случаях это приводит к положительному числу (например, первичное тело выходит за пределы своего большого кроющего), а в других случаях к отрицательному числу (например, первичное полностью покрыто большим кроющим, как это происходит у некоторых видов воробьиных). ). Затем определяется самое длинное первичное перо и измеряется разница в миллиметрах между: i) длиной этого первичного пера; ii) всех остальных праймериз; и iii) самой длинной средней школы. Если на каком-либо основном пере имеется выемка или выемка , это отмечается и измеряется расстояние между кончиком пера и любой выемкой, а также глубина выемки. Все измерения расстояний производятся при закрытых крыльях птицы, чтобы сохранить относительное положение перьев. Несмотря на то, что среди представителей одного вида могут быть значительные различия, и хотя на результаты явно влияют эффекты линька и регенерация перьев — даже очень близкие виды демонстрируют явные различия в формуле крыльев. [18]
- обшивка крыла
- А также краевые кроющие . Группа очень мягких кроющих перьев выстилает передний край нижней стороны крыла. Хотя подкладка крыльев состоит из рядов этих перьев, на вид они гладкие, поэтому часто невозможно увидеть границы рядов только на глаз. [26]
- крылья
- Определены типы крыльев: эллиптические ; высокоскоростной ; высокое соотношение сторон ; парящий . птицы Передние конечности , являющиеся ключом к полету. Каждое крыло имеет центральную ось, состоящую из трёх костей конечностей: плечевой , локтевой и лучевой . Рука, или manus , которая изначально состояла из пяти пальцев, редуцируется до трех пальцев (цифры II, III и IV или I, II, III в зависимости от используемой схемы). [453] ), который служит опорой для основных маховых перьев , одной из двух групп маховых перьев, отвечающих за форму профиля крыла. Другой набор маховых перьев, расположенный за запястным суставом на локтевой кости, называется вторичными . Остальные перья на крыле известны как кроющие , их три набора. На крыле иногда имеются рудиментарные когти. У большинства видов они теряются к тому времени, когда птица становится взрослой (например, хорошо заметные, используемые для активного лазания птенцами гоацина ), но когти сохраняются и во взрослом возрасте у птицы-секретаря , крикунов , плавников , страусов , некоторых стрижей и многих других. , как локальный признак, у немногих экземпляров. Крылья птиц обычно делятся на четыре типа: i) эллиптические крылья — короткие, закругленные и имеющие низкое удлинение, они позволяют маневрировать в ограниченном пространстве, например, в густой растительности. Они распространены у лесных хищников, многих мигрирующих видов воробьиных , а также у таких видов, как фазаны и куропатки , которые используют быстрый взлет, чтобы уклониться от хищников. ii) Высокоскоростные крылья — короткие заостренные крылья, которые в сочетании с большой нагрузкой на крыло и быстрыми взмахами крыльев обеспечивают энергетически затратную высокую скорость, используемую птицей с самой высокой скоростью крыльев, сапсан . У большинства уток и чистиковых крыльев есть этот тип крыльев, но они используют эту конфигурацию для другой цели — «летать» под водой. iii) Крылья с большим удлинением - характеризуются низкой нагрузкой на крыло и будучи намного длиннее, чем ширина, они используются для более медленного полета, который может принимать форму почти зависания (как это делают пустельги , крачки и козодои ) или парения. и планирующий полет, особенно динамическое парение , используемое морскими птицами , которое использует изменение скорости ветра на разных высотах ( сдвиг ветра ) над океанскими волнами для обеспечения подъемной силы. iv) Парящие крылья с глубокими прорезями — часто встречаются у крупных видов внутренних птиц, таких как орлы , стервятники , пеликаны и аисты . Прорези на концах крыльев, между основными элементами, уменьшают индуцированное сопротивление и вихри на законцовках крыла , а более короткий размер крыльев способствует взлету (крыльям с большим удлинением требуется длинное руление, чтобы подняться в воздух). [454]
- размах крыльев
- Расстояние между кончиками полностью раскрытых крыльев . [455] Из ныне живущих птиц с самым большим размахом крыльев — странствующий альбатрос , размер которого обычно составляет от 2,5 до 3,5 м (от 8 футов 2 дюйма до 11 футов 6 дюймов). [456] Напротив, из ныне живущих видов птиц с наименьшим размахом крыльев — это колибри-пчёла (также самая маленькая птица в целом) с размахом крыльев всего 6,6 см (2,6 дюйма). [457]
- зимнее оперение
- См. основное оперение .




З [ править ]
- зигодактильный
- А еще косолапый . Также определено: гетеродактильный . Описывает тетрадактильных (четырехпалых) птиц, у которых строение стопы состоит из двух пальцев, выступающих вперед, и двух пальцев, выступающих назад, например, у попугаев , дятлов и кукушек . У большинства птиц с зигодактильными лапами выступающие вперед пальцы — это второй и третий пальцы, а выступающие назад — четвертый палец и большой палец . Однако только у трогонов третий и четвертый пальцы находятся спереди, а второй и большой палец сзади. Такое расположение иногда обозначают отдельно и называют гетеродактильным . [20] [458]
См. также [ править ]
Пояснительные сноски [ править ]
- ↑ Орнитологи из Американского музея естественной истории в статье 2016 года, опубликованной в PLOS ONE , предположили, что использование « концепции биологического вида » — классификации группы птиц как составляющих один вид по их способности к размножению — устарело и следует отбросить в пользу видовой модели, которая учитывает сходство и несходство таких характеристик, как оперение, рисунок и окраска, для индивидуализации видов птиц. Согласно такой модели предполагается, что по состоянию на 2016 год в мире насчитывается ± 18 000 отдельных видов птиц. [4]
- ^ Эму и казуары являются исключением, поскольку их послевые перья не являются ни пушистыми, ни миниатюрными по сравнению с лопастями, а сравнимы с ними по размеру; такие перья иногда называют «двойными». Перья страуса и нанду , также нелетающих птиц, напротив, имеют «мягкий и тонкий» вид и лишены каких-либо усиков , придающих жесткость, необходимую для рабочих маховых перьев . [7]
- ^ К птицам, которые подвергаются второй линьке, не связанной с размножением (третья линька в целом), относятся: куропатки ( Lagopus ) (летом); длиннохвостая утка ( Clangula hyemalis ) (зимой); ерш ( Philomachus pugnax ) (концом зимы или ранней весной); и может встречаться у других представителей семейства куликов (Scolopacidae). [31]
- ^ Что касается боков, определяемых как нижние части, в трактате «Орнитология в лаборатории и в полевых условиях » автор заявляет: «[а] хотя технически стороны тела принадлежат как к верхним , так и к нижним частям , воображаемая линия, разделяющая две поверхности расположены настолько высоко на туловище, что боковые стороны тела обычно считаются только частями». [96]
Цитаты [ править ]
- ^ Прум, Ричард О. Прум (19 декабря 2008 г.). «Кто твой папа?». Наука . 322 (5909): 1799–1800. дои : 10.1126/science.1168808 . ПМИД 19095929 . S2CID 206517571 .
- ^ Ошибка включения: {{ En }} предназначен только для использования в пространстве имен файлов. Вместо этого используйте {{ lang-en }} или {{ in lang |en}} . Гилл, Ф., Д. Донскер и П. Расмуссен (редакторы). 2024. Всемирный список птиц МОК (версия 14.1). https://www.worldbirdnames.org. Архивировано 4 октября 2020 г. в Wayback Machine.
- ^ Ньютон, Ян (2003). Видообразование и биогеография птиц . Профессиональное издательство Персидского залива. п. 93. ИСБН 978-0-12-517375-9 .
- ^ Уотерс, Ханна (15 декабря 2016 г.). «Новое исследование удваивает количество видов птиц в мире за счет нового определения понятия «вид» » . Национальное общество Одюбона. Архивировано из оригинала 21 декабря 2016 года.
- ^ Энциклопедия perthensis, или Универсальный словарь искусств, наук, литературы и т. д . Эдинбург: Джон Браун. 1816. с. 166.
- ↑ Перейти обратно: Перейти обратно: а б с Уивер 1981 , с. 12
- ^ Корни 2006 , с. 16
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Петтингилл 1985 , с. 32
- ^ Мандал, Фатик Баран (2012). Учебник поведения животных . Нью-Дели: PHI Learning Pvt. ООО с. 188. ИСБН 978-81-203-4519-5 .
- ↑ Перейти обратно: Перейти обратно: а б с Хамфри, Филип С.; Паркс, Кеннет К. (январь 1959 г.). «Подход к изучению линьки и оперения». Аук . Том. 76, нет. 1. С. 1–31. дои : 10.2307/4081839 . JSTOR 4081839 .
- ↑ Перейти обратно: Перейти обратно: а б с Саммерс, Кайл; Креспи, Бернар (2013). Социальная эволюция человека: фундаментальные работы Ричарда Д. Александра . Издательство Оксфордского университета. п. 192. ИСБН 978-0-19-933963-1 .
- ^ Хауэлл, Стив Н.Г. (2014). Справочник Петерсона по линьке североамериканских птиц . Хоутон Миффлин Харкорт. п. 8. ISBN 978-0-547-48769-4 .
- ↑ Перейти обратно: Перейти обратно: а б Джонстон, Ричард (2013). Современная орнитология . Springer Science & Business Media. п. 34. ISBN 978-1-4615-6781-3 .
- ↑ Перейти обратно: Перейти обратно: а б Урфи, Эй Джей (2011). Раскрашенный аист: экология и охрана . Springer Science & Business Media. п. 88. ИСБН 978-1-4419-8468-5 .
- ^ Скотт, Линнетт (2008). Реабилитация дикой природы . Национальная ассоциация реабилитаторов дикой природы. п. 50. ISBN 978-1-931439-23-7 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Петтингилл 2013 , с. 371
- ↑ Перейти обратно: Перейти обратно: а б Петтингилл, 1985 , стр. 16–21.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Кэмпбелл и Лэк, 1985 , с. 656
- ^ Петтингилл 1985 , с. 187
- ↑ Перейти обратно: Перейти обратно: а б Гилл 1995 , стр. 59–61.
- ^ Осборн, Софи А.Х. (сентябрь 1998 г.). «Муравьи американской медведицы ( Cinclus mexicanus )» (PDF) . Бюллетень Уилсона . Том. 110, нет. 3. С. 423–425. Архивировано (PDF) из оригинала 14 ноября 2019 г. Проверено 22 января 2017 г.
- ^ Альдерфер, Джонатан К.; Данн, Джон (январь 2007 г.). Основы National Geographic Birding: все инструменты, методы и советы, которые вам нужны, чтобы начать и стать лучшим орнитологом . Нэшнл Географик. п. 216 . ISBN 978-1-4262-0135-6 .
- ↑ Перейти обратно: Перейти обратно: а б Проктор и Линч 1998 , с. 98
- ^ Улмер, Мартин Джон; Хаупт, Роберт Э.; Хикс, Эллис А. (1962). Сравнительная анатомия хордовых: Лабораторное пособие . Харпер и Роу. п. 169.
- ^ Ньютон и др. 1899 , с. 25
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Лэк, 1985 , с. 60
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Виникомб 2014 , с. 15
- ↑ Перейти обратно: Перейти обратно: а б Чендлер 1916 , с. 252
- ↑ Перейти обратно: Перейти обратно: а б с Петтингилл, 1985 , стр. 32–33.
- ^ Аллен, Эльза Гердрум (1951). История американской орнитологии до Одюбона . Рассел и Рассел. п. 458.
- ↑ Перейти обратно: Перейти обратно: а б с Петтингилл, 1985 , стр. 189–192.
- ↑ Перейти обратно: Перейти обратно: а б Куэ 1890 , с. 198
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Проктор и Линч 1998 , с. 66
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Гилл 1995 , с. 148
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Кэмпбелл и Лэк, 1985 , с. 47
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Лэк, 1985 , с. 598
- ↑ Перейти обратно: Перейти обратно: а б с Кэмпбелл и Лэк, 1985 , с. 375
- ^ Партингтон, Чарльз Фредерик (1835). Британская циклопедия естественной истории: объединение научной классификации животных, растений и минералов . Орр и Смит. п. 417 .
- ^ Ральф, Чарльз Л. (май 1969 г.). «Контроль цвета у птиц». Американский зоолог . Том. 9, нет. 2. С. 521–530. дои : 10.1093/icb/9.2.521 . JSTOR 3881820 . ПМИД 5362278 .
- ↑ Перейти обратно: Перейти обратно: а б с Хауэлл 2007 , с. 23
- ↑ Перейти обратно: Перейти обратно: а б Грандин, Темпл (2010). Улучшение благополучия животных: практический подход . Оксфордшир, Великобритания: CABI. п. 110. ИСБН 978-1-84593-541-2 .
- ^ «Птичьи клювы: анатомия, уход и болезни» . Департамент ветеринарных и водных услуг, д-р. Фостер и Смит. Архивировано из оригинала 4 июня 2012 года . Проверено 29 января 2017 г.
- ^ Эш, Лидия. «Справиться со своим Раптором» . Современный ученик. Архивировано из оригинала 6 апреля 2005 года . Проверено 29 января 2017 г.
- ^ Бэрд, Спенсер Фуллертон; Брюэр, Томас Мэйо; Риджуэй, Роберт (1874). История птиц Северной Америки . Маленький, Браун. п. 535.
- ^ Бирма, Натан (12 августа 2004 г.). «Добавьте это в список жизни: «Птицы» вдохновили на множество слов» . Чикаго Трибьюн . Архивировано из оригинала 12 августа 2012 г. Проверено 20 января 2017 г.
- ^ Ленды 1980 , с. 232
- ^ Шрайбер, Элизабет Энн; Бургер, Джоанна, ред. (2002). Биология морских птиц . Бока-Ратон, Флорида: CRC Press. п. 325. ИСБН 978-0-8493-9882-7 .
- ^ Каннингем, Сьюзен Дж.; Элли, Морис Р.; Кастро, Изабель; Поттер, Мюррей А.; Каннингем, Малькольм; Пайн, Майкл Дж. (2010). «Морфология клюва ибисов предполагает наличие дистанционно-тактильной сенсорной системы для обнаружения добычи». Аук . Том. 127, нет. 2. С. 308–316. дои : 10.1525/аук.2009.09117 . S2CID 85254980 .
- ^ Буссе, Пшемыслав; Мейснер, Влодзимеж (2015). Инструкция по устройству кольцевания птиц . Варшава: Де Грютер. ISBN 978-83-7656-053-3 .
- ^ Уивер 1981 , с. 23
- ↑ Перейти обратно: Перейти обратно: а б с Сибли и др. 2001 , с. 17
- ^ Уивер 1981 , с. 24
- ^ Вийемье, Франсуа, изд. (2011). Американский музей естественной истории птиц восточного региона Северной Америки . Издательство ДК. п. 473. ИСБН 978-0-7566-7387-1 .
- ^ Ранкини, Хуан Омедес; Гарсия, Франсиско Аро (1958). Зоогенетика . сохранено
- ^ «Определение Мерриама-Вебстера» . Архивировано из оригинала 26 марта 2017 года . Проверено 12 марта 2017 г.
- ^ Тернер, Дж. Скотт (1997). «О теплоемкости птичьего яйца, согреваемого выводком». Физиологическая зоология . Том. 70, нет. 4. С. 470–80. дои : 10.1086/515854 . ПМИД 9237308 .
- ^ Арагон, С.; Мёллер, АП; Солер, Джей-Джей; Солер, М. (1999). «Молекулярная филогения кукушек подтверждает полифилетическое происхождение выводкового паразитизма». Журнал эволюционной биологии . Том. 12, нет. 3. С. 495–506. дои : 10.1046/j.1420-9101.1999.00052.x . S2CID 16923328 .
- ^ Соренсон, доктор медицины; Пейн, РБ (2001). «Единое древнее происхождение выводкового паразитизма у африканских вьюрков: последствия коэволюции хозяина и паразита» (PDF) . Эволюция . Том. 55, нет. 12. С. 2550–2567. doi : 10.1554/0014-3820(2001)055[2550:asaoob]2.0.co;2 . hdl : 2027.42/72018 . ПМИД 11831669 .
- ^ Соренсон, доктор медицины; Пейн, РБ (2002). «Молекулярно-генетические взгляды на паразитизм птичьего выводка». Интегративная и сравнительная биология . Том. 42, нет. 2. С. 388–400. дои : 10.1093/icb/42.2.388 . ПМИД 21708732 .
- ↑ Перейти обратно: Перейти обратно: а б Ритчисон, Гэри. «Орнитология (Био 554/754): Эволюция перьев» . Департамент биологических наук Университета Восточного Кентукки. Архивировано из оригинала 25 февраля 2017 года . Проверено 16 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Чендлер 1916 , с. 250
- ^ «Латинский словарь» . Архивировано из оригинала 16 октября 2021 года . Проверено 1 мая 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Эрлих, Пол Р.; Добкин, Дэвид С.; Эй, Дэррил. « Голоса птиц» и «Вокальное развитие» из эссе «Птицы Стэнфорда» . Архивировано из оригинала 25 июля 2018 года . Проверено 9 сентября 2008 г.
- ^ Ленгань, Т.; Лауга, Дж.; Обен, Т. (2001). «Внутрислоговые акустические сигналы, используемые королевским пингвином для распознавания родителей и птенцов: экспериментальный подход» (PDF) . Журнал экспериментальной биологии . Том. 204, нет. Пт. 4. стр. 663–672. ПМИД 11171348 . Архивировано (PDF) из оригинала 30 сентября 2007 г. Проверено 1 апреля 2017 г.
- ^ Маккей, Барри Кент (январь 2001 г.). Звуки птиц: как и почему птицы поют, кричат, болтают и визжат . Книги Стэкпола. п. 28. ISBN 978-0-8117-2787-7 .
- ^ Алкок, Джон (1998). Поведение животных: эволюционный подход (6-е изд.). Сандерленд: Sinauer Associates. ISBN 978-0-87893-009-8 .
- ^ Браун, CH (1982). «Чревовещание и локализуемые вокализации у птиц». Журнал психологии животных . Том 59, № 4. С. 338–350. дои : 10.1111/j.1439-0310.1982.tb00346.x .
- ↑ Перейти обратно: Перейти обратно: а б Уивер 1981 , с. 34
- ^ Сибли и др. 2001 , стр. 68–69.
- ^ Ровински, Кейт (2011). Радость наблюдения за птицами: руководство для начинающих . Skyhorse Publishing Inc., стр. 22–. ISBN 978-1-61608-122-5 .
- ^ Торп, штат Вашингтон (1963). «Антифонное пение птиц как свидетельство времени слуховой реакции птиц». Природа . Том. 197, нет. 4869. стр. 774–776. Бибкод : 1963Natur.197..774T . дои : 10.1038/197774a0 . S2CID 30542781 .
- ^ Стоукс, AW; Уильямс, HW (1968). «Антифонный крик перепела» (PDF) . Аук . Том. 85, нет. 1. С. 83–89. дои : 10.2307/4083626 . JSTOR 4083626 . Архивировано (PDF) из оригинала 29 апреля 2014 г. Проверено 1 апреля 2017 г.
- ^ Харрис, Тони; Франклин, Ким (2000). Сорокопуты и кустарниковые сорокопуты . Издательство Принстонского университета. стр. 257–260. ISBN 978-0-691-07036-0 .
- ^ Осмастон, Б.Б. (1941). «Дуэтинг» у птиц». Ибис . Том. 5, нет. 2. С. 310–311. дои : 10.1111/j.1474-919X.1941.tb00620.x .
- ^ Власть, ДМ (1966). «Антифонный дуэт и данные о времени слуховой реакции у попугая с оранжевым подбородком». Аук . Том. 83, нет. 2. С. 314–319. дои : 10.2307/4083033 . JSTOR 4083033 .
- ^ Хэнкок, Джеймс; Кушлан, Джеймс (2010). Справочник по цапли . Лондон: A&C Black. стр. 20, 101–102. ISBN 978-1-4081-3496-2 .
- ^ Флойд 2008 , с. 495
- ^ Шарп, Ричард Боудлер (1888). Каталог птиц Британского музея . Зоологический отдел Британского музея естественной истории.
- ^ Одюбон, Джон Джеймс; Амадон, Дин; Булл, Джон Л. (1967). Птицы Америки . Дуврские публикации. OCLC 555150 .
- ^ Берк, Дон (2005). Полный задний двор Берка: Лучшая книга информационных бюллетеней . п. 702. ИСБН 9781740457392 .
- ^ Баратти, Мариэлла; Амманнати, Мартина; Магнелли, Клаудия; Массоло, Алессандро; Десси-Фульгери, Франческо (2010). «Связаны ли большие сережки с определенными генотипами MHC у фазанов-самцов?». Генетика . Том 138, нет. 6. стр. 657–665. дои : 10.1007/s10709-010-9440-5 . ISSN 0016-6707 . ПМИД 20145977 . S2CID 35053439 .
- ^ Бухгольц, Ричард (1996). «Терморегуляционная роль неоперенной головы и шеи у самцов диких индеек» (PDF) . Аук . Том. 113, нет. 2. С. 310–318. дои : 10.2307/4088897 . JSTOR 4088897 . Архивировано (PDF) из оригинала 18 мая 2014 г. Проверено 11 февраля 2017 г.
- ^ Блейк, Эммет Рид (1977). Руководство неотропических птиц . Издательство Чикагского университета. п. 428. ИСБН 978-0-226-05641-8 .
- ^ Пратт, Тейн К.; Билер, Брюс М. (2014). Птицы Новой Гвинеи (2-е изд.). Оксфорд: Издательство Принстонского университета. п. 262. ИСБН 978-1-4008-6511-6 .
- ^ Рейли, Эдгар М.; Национальное общество Одюбона (1968). Иллюстрированный справочник Audubon по американским птицам . МакГроу-Хилл. п. 142 .
- ^ Стейнегер, Леонард Гесс (1885). Итоги орнитологических исследований на Командорских островах и на Камчатке . Типография правительства США. стр. 52–53 .
- ^ Национальное географическое общество (2009). Харрис, Тим (ред.). Полное издание «Птицы мира» . Нэшнл Географик. п. 226. ИСБН 978-1-4262-0403-6 .
- ^ Мобли и др. 2008 , с. 339
- ^ Карнаби 2008 , с. 124
- ^ Мобли и др. 2008 , с. 441
- ^ Юпитер, Тони; Парр, Майк (2010). Попугаи: Путеводитель по попугаям мира . А&С Черный. п. 17. ISBN 978-1-4081-3575-4 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Штеттенхайм, Питер Р. (2000). «Покровная морфология современных птиц — обзор» . Интегративная и сравнительная биология . Том. 40, нет. 4. С. 461–477. дои : 10.1093/icb/40.4.461 . Архивировано из оригинала (PDF) 20 апреля 2012 г.
- ^ Мужо, Франсуа; Арройо, Беатрис Э. (22 июня 2006 г.). «Отражение ультрафиолета в мозговиках хищников». Письма по биологии . Том. 2, нет. 2. С. 173–176. дои : 10.1098/rsbl.2005.0434 . ПМЦ 1618910 . ПМИД 17148356 .
- ^ Леопольд, Альдо Старкер (1972). Дикая природа Мексики: дичь, птицы и млекопитающие . Беркли, Калифорния: Издательство Калифорнийского университета. п. 202 . ISBN 978-0-520-00724-6 .
- ^ Олдертон, Дэвид (1996). Руководство для птицеводов по волнистым попугайчикам . Тетра Пресс. п. 12.
- ↑ Перейти обратно: Перейти обратно: а б с д Петтингилл 1985 , с. 9
- ↑ Перейти обратно: Перейти обратно: а б с д и Виникомб 2014 , с. 14
- ^ Эрлих, Пол Р.; Добкин, Дэвид С.; Сыворотка, Дэррил (1988). «Питьевой» . Птицы Стэнфорда . Стэнфордский университет. Архивировано из оригинала 2 мая 2006 года . Проверено 29 января 2017 г.
- ^ Царь, Элла; Мартинес Дель Рио, К.; Ижаки И.; Арад, З. (2005). «Могут ли птицы быть аммонотелическими? Баланс и выделение азота у двух плодоядных». Журнал экспериментальной биологии . Том. 208, нет. 6. С. 1, 025–34. дои : 10.1242/jeb.01495 . ПМИД 15767304 .
- ^ Скадхауге, Э.; Эрльвангер, К.Х.; Рузива, SD; Данцер, В.; Эльбронд, В.С.; Чамунорва, JP (2003). «Обладает ли страусиный копродеум ( Struthio Camelus ) электрофизиологическими свойствами и микроструктурой других птиц?». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 134 (4): 749–755. дои : 10.1016/S1095-6433(03)00006-0 . ПМИД 12814783 .
- ^ Йонг, Эд (6 июня 2013 г.). «Феномены: не совсем ракетостроение. Как куры потеряли свои пенисы (а утки сохранили свои)» . Феномены.nationalgeographic.com. Архивировано из оригинала 9 июня 2013 года . Проверено 29 января 2017 г.
- ^ «Орнитология, 3-е издание - Водоплавающие птицы: отряд Anseriformes» . Архивировано из оригинала 22 июня 2015 года . Проверено 29 января 2017 г.
- ^ Петтингилл 1985 , с. 376
- ^ Максвелл, Кеннет Э. (2013). Секс-императив: эволюционная история сексуального выживания . Спрингер. п. 99. ИСБН 978-1-4899-5988-1 .
- ^ Донли, Боб (2016). Птичья медицина и хирургия на практике: птицы-компаньоны и вольеры, второе издание . ЦРК Пресс. п. 12. ISBN 978-1-4822-6019-9 .
- ^ Лак, Дэвид (1947) [1887]. «Значение размера кладки (часть I-II)» . Ибис . Том. 89, нет. 2. С. 302–352. дои : 10.1111/j.1474-919X.1947.tb04155.x .
- ^ Петтингилл 1985 , с. 310
- ^ Маккейб, Роберт А. (1943). Венгерская куропатка (perdix perdix linn.) изучается в Висконсине . Университет Висконсина — Мэдисон. п. 28.
- ^ Андерсон, Дэйв (февраль – март 2008 г.). «Куриный гребешок». Журнал о птицеводстве на заднем дворе .
- ^ Гилл 1995 , стр. 138–141.
- ^ Данчин, Э.; Вагнер, Р.Х. (1997). «Эволюция колониальности: появление новых перспектив» (PDF) . Тенденции экологии и эволюции . Том. 12, нет. 2. С. 342–347. дои : 10.1016/S0169-5347(97)01124-5 . ПМИД 21238100 . Архивировано из оригинала (PDF) 3 октября 2011 г. Проверено 1 апреля 2017 г.
- ^ Роллан, Сесиль; Данчин, Этьен; де Фрайпон, Мишель (1998). «Эволюция колониальности птиц в зависимости от еды, среды обитания, хищничества и особенностей жизненного цикла: сравнительный анализ» (PDF) . Американский натуралист . Том. 151, нет. 6. С. 514–529. дои : 10.1086/286137 . ПМИД 18811373 . Архивировано из оригинала (PDF) 20 июня 2010 г.
- ^ Куэ 1890 , с. 155
- ^ Кэмпбелл и Лак 1985 , с. 105
- ^ Кляйн, Эдвард (1785). Элементы гистологии . Филадельфия: Леа. п. 124 .
- ^ Pierce, R.M. (1911). Dictionary of aviation . Рипол Классик. pp. 69–70. ISBN 978-5-87745-565-8 .
- ↑ Перейти обратно: Перейти обратно: а б Мобли и др. 2008 , с. 295
- ^ Элфик 2016 , стр. 22–23
- ^ Карнаби 2008 , с. 10
- ^ Тай, Хилари (декабрь 1985 г.). «Эректильный гребень и другие оперения на голове рода Picathartes» . Бюллетень Клуба британских орнитологов . Том. 106, нет. 3. п. 91. Архивировано из оригинала 07 августа 2018 г. Проверено 14 января 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б Проктор и Линч 1998 , с. 64
- ↑ Перейти обратно: Перейти обратно: а б Хайфилл, Кэрол. «Эти великолепные гребни какаду» . Какадуовый рай. Архивировано из оригинала 1 марта 2017 года . Проверено 23 марта 2017 г.
- ^ Гилл 1995 , стр. 318–319.
- ^ Мустаки, Никки (2005). Попугаи для чайников (1-е изд.). Индианаполис, Индиана: Уайли. ISBN 978-0764583537 .
- ↑ Перейти обратно: Перейти обратно: а б с д Петтингилл 1985 , с. 21
- ^ Бернс, Джим (2008). Джим Бернс «Птицы Аризоны: от заднего двора до глуши» . Издательство Университета Аризоны. п. 166. ИСБН 978-0-8165-2644-4 .
- ^ Рамель, Гордон (29 сентября 2008 г.). «Пищеварительный канал птиц» . Архивировано из оригинала 1 февраля 2017 года . Проверено 16 января 2017 г.
- ^ Леви, Венделл (1977). Голубь . Levi Publishing Co, Inc. Самтер, Южная Каролина: ISBN 978-0-85390-013-9 .
- ↑ Перейти обратно: Перейти обратно: а б Сильвер, Рэй (1984). «Пролактин и воспитание детей в семье голубей» (PDF) . Журнал экспериментальной зоологии . Том. 232, нет. 3. С. 617–625. дои : 10.1002/jez.1402320330 . ПМИД 6394702 . Архивировано из оригинала 13 сентября 2016 года.
{{cite magazine}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Эрауд, К.; Дори, А.; Жаке, А.; Фавр, Б. (2008). «Урожайное молоко: потенциальный новый путь родительского воздействия, опосредованного каротиноидами». Журнал птичьей биологии . Том. 39, нет. 2. С. 247–251. дои : 10.1111/j.0908-8857.2008.04053.x .
- ^ Эрлих, Пол Р.; Добкин, Дэвид С. и Уэй, Дэррил (1988). Птичье молоко . Архивировано 15 октября 2012 г. в Wayback Machine . Стэнфорд.edu
- ^ Филлипс, Кэмпбелл (22 сентября 2011 г.). «Разгаданы тайны голубиного молока» . Австралийское географическое издание. Архивировано из оригинала 14 августа 2013 года . Проверено 17 августа 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Петтингилл 1985 , с. 10
- ^ Гилл 1995 , с. 106
- ^ Райзер, Фред А. (1985). Птицы Большого Бассейна: Естественная история . Университет Невады Пресс. п. 22. ISBN 978-0-87417-080-1 . [ постоянная мертвая ссылка ]
- ^ Кэмпбелл и Лак 1985 , с. 127
- ^ Куэ 1890 , с. 152
- ^ Пайл, Питер; Хауэлл, Стив Н.Г.; Юник, Роберт П.; ДеСанте, Дэвид Ф. (1987). Справочник по идентификации воробьинообразных Северной Америки . Болинас, Калифорния: Slate Creek Press. стр. 6–7. ISBN 978-0-9618940-0-9 .
- ^ Боррас, А.; Паскаль, Дж.; Сенар, JC (осень 2000 г.). «Что измеряют различные размеры клюва и какой метод лучше всего использовать у зерноядных птиц?» (PDF) . Журнал полевой орнитологии . Том. 71, нет. 4. С. 606–611. дои : 10.1648/0273-8570-71.4.606 . JSTOR 4514529 . Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 17 января 2017 г.
- ^ Малларни, Киллиан; Свенссон, Ларс; Зеттерстрем, Дэн; Грант, Питер Дж. (1999). Путеводитель Коллинза по птицам: наиболее полный путеводитель по птицам Британии и Европы . Лондон: Харпер Коллинз. п. 357. ИСБН 978-0-00-711332-3 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Карнаби 2008 , с. 334
- ^ Моллес, Мануэль С. младший (1999). Экология: концепции и приложения (международное изд.). Дубьюк: МакГроу-Хилл. п. 510 . ISBN 978-0-07-042716-7 .
- ^ Хоузи, Джефф; Мелфи, Вики; Шейла Панкхерст (2013). Животные зоопарка: поведение, управление и благополучие . ОУП Оксфорд. п. 419. ИСБН 978-0-19-969352-8 .
- ^ Сканес, Колин Г. (2014). Птичья физиология Стерки . Эльзевир Наука. п. 61. ИСБН 978-0-12-407243-5 .
- ^ Адамс, Рик А.; Педерсен, Скотт К. (2000). Онтогенез, функциональная экология и эволюция летучих мышей . Издательство Кембриджского университета. п. 124. ИСБН 978-1-139-42949-8 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж Ирвин, Марк Д.; Стоунер, Джон Б.; Кобо, Аарон М. (2013). Зоопарк: введение в науку и технологии . Издательство Чикагского университета. п. 166. ИСБН 978-0-226-92532-5 .
- ↑ Перейти обратно: Перейти обратно: а б Мур, Питер Д. (1 января 2009 г.). Тропические леса . Издательство информационной базы. п. 164. ИСБН 978-1-4381-1874-1 .
- ↑ Перейти обратно: Перейти обратно: а б Класинг, Кирк К. (21 мая 1998 г.). Сравнительное питание птиц . Такси Интернешнл. п. 3. ISBN 978-0-85199-219-8 .
- ^ Свон, Уильям Дрейпер (1848). Поучительный читатель, или Курс чтения по естествознанию, науке и литературе: предназначен для школ . Томас, Каупертвейт. п. 71.
- ^ Ньютон, Ян (1998). Ограничение численности птиц . Академическая пресса. п. 341 . ISBN 978-0-08-087923-9 .
- ^ Селби, Майкл Джон (1969). Поверхность земли . Касселл. п. 417 . ISBN 978-0-304-92858-3 .
- ^ Элфик 2016 , стр. 110–111
- ^ Спирман, Ричард Ян Кэмпбелл (1973). Покровы: Учебник биологии кожи . Издательство Кембриджского университета. п. 97 . ISBN 978-0-521-20048-6 .
- ↑ Перейти обратно: Перейти обратно: а б с дель Хойо, Эллиотт и Саргатал 1992 , с. 39
- ^ Скотт 2010 , с. 31
- ^ Элфик 2016 , стр. 75–77
- ^ Горман, Джерард (2014). Дятлы мира: Фотопутеводитель . Книги Светлячка. п. 28. ISBN 978-1-77085-309-6 .
- ^ «Бекас Лэтэма — ты знал?» . Птицы во дворах . Жизнь птиц Австралии. Архивировано из оригинала 25 января 2017 года . Проверено 25 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Джаспер, Теодор (1878). Популярная орнитология Штудера… . Колумбус, Огайо: JH Studer & Company. п. 2.
- ↑ Перейти обратно: Перейти обратно: а б Фарнер, Дональд С.; Кинг, Джеймс Р. (2013). Птичья биология . Эльзевир. п. 13. ISBN 978-1-4832-6942-9 .
- ^ Пикрафт, WP (1908). Scientific American: Приложение: Любопытные явления пигментации яичной скорлупы . Манн и компания. п. 376.
- ^ Хилл, Джеффри Эдвард; МакГроу, Кевин Дж. (2006). Окраска птиц: механизмы и измерения . Издательство Гарвардского университета. п. 368. ИСБН 978-0-674-01893-8 .
- ^ Ариас, Дж.Л.; Фернандес, MS (2001). «Роль молекул внеклеточного матрикса в формировании и структуре оболочки». Мировой научный журнал по птицеводству . Том. 57, нет. 4. С. 349–357. дои : 10.1079/WPS20010024 . S2CID 86152776 .
- ↑ Перейти обратно: Перейти обратно: а б Хантон, П. (2005). «Исследование структуры и качества яичной скорлупы: исторический обзор». Revista Brasileira de Ciencia Avícola . Том. 7, нет. 2. С. 67–71. дои : 10.1590/S1516-635X2005000200001 .
- ^ Нюс, Ив; Гаутрон, Жоэль; Гарсия-Руис, Хуан М.; Хинкке, Максвелл Т. (2004). «Минерализация яичной скорлупы птиц: биохимическая и функциональная характеристика белков матрикса». Comptes Рендус Палевол . Том. 3, нет. 6–7. стр. 549–562. дои : 10.1016/j.crpv.2004.08.002 .
- ^ Романофф, Алексис Л.; Романофф, Анастасия Дж. (1949). Птичье яйцо . Нью-Йорк: Уайли. OCLC 191965542 .
- ^ Берли, RW; Вадехра, Дхарам В. (1989). Птичье яйцо: химия и биология . Нью-Йорк: Уайли. ISBN 978-0-471-84995-7 .
- ^ Лавелен, И., Н.; Мейри, М. Пайнс (2000). «Новый взгляд на формирование яичной скорлупы». Птицеводство . Том. 79, нет. 7. С. 1014–1017. дои : 10.1093/ps/79.7.1014 . ПМИД 10901204 .
{{cite magazine}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Олдертон, Дэвид (1998). Полное руководство по уходу за птицами . Книжный дом Хауэлла. ISBN 978-0-87605-038-5 .
- ^ Харрисон, Джон (2011). Садовые куры и другая домашняя птица . Литтл, Группа Брауновой книги. п. 125. ИСБН 978-0-7160-2276-3 .
- ^ Дамероу, Гейл (2013). Высиживание и выращивание собственных цыплят: цыплят, индеек, уток, гусей, цесарок . Стори Паблишинг. п. 142. ИСБН 978-1-60342-878-1 .
- ^ Паттен, Брэдли М. (2008). Ранняя эмбриология цыпленка . ООО «Вайлдсайд Пресс». п. 17. ISBN 978-1-4344-7385-1 .
- ^ «Дракон Сумо». экзотическим животным доктора К. Скорая помощь Сезон 03 E01. 21 мая 2016 г. Прошло 26 минут. Nat Geo Wild .
- ^ Стаут, Джейн Д. (май 2016 г.). «Общие чрезвычайные ситуации у домашних птиц». Ветеринарные клиники Северной Америки. Практика с экзотическими животными . Том. 19, нет. 2. С. 513–41. дои : 10.1016/j.cvex.2016.01.002 . ПМИД 26948267 .
- ^ Бэрроуз, Эдвард М. (2000). Справочник по поведению животных: Словарь по поведению, экологии и эволюции животных, второе издание . ЦРК Пресс. п. 79 . ISBN 978-0-8493-2005-7 .
- ^ Гилл 1995 , с. 459
- ^ Уорхэм, Дж (1990). Буревестники: их экология и системы размножения . Лондон: Академическая пресса. ISBN 978-0-12-735420-0 .
- ^ Кэмпбелл и Лак 1985 , с. 178
- ↑ Перейти обратно: Перейти обратно: а б с Джел, Джозеф Р. младший (сентябрь 1968 г.). «Яичный зуб некоторых ржанкообразных птиц» (PDF) . Бюллетень Уилсона . Том. 80, нет. 3. С. 328–9. Архивировано (PDF) из оригинала 22 октября 2020 г. Проверено 19 октября 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Перринс, Кристофер М.; Аттенборо, Дэвид; Арлотт, Норман (1987). Путеводитель нового поколения по птицам Британии и Европы . Остин, Техас: Издательство Техасского университета. п. 205. ИСБН 978-0-292-75532-1 .
- ^ Кларк, Джордж А. младший (сентябрь 1961 г.). «Появление и время появления яичных зубов у птиц» (PDF) . Бюллетень Уилсона . Том. 73, нет. 3. С. 268–278. Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 20 января 2017 г.
- ^ Гилл 1995 , с. 427
- ^ Гилл 1995 , с. 428
- ↑ Перейти обратно: Перейти обратно: а б Трейл 2001 , с. 6
- ↑ Перейти обратно: Перейти обратно: а б с Прум, Ричард О .; Кисть, АХ (2002). «Эволюционное происхождение и разнообразие перьев» (PDF) . Ежеквартальный обзор биологии . Том. 77, нет. 3. С. 261–295. дои : 10.1086/341993 . ПМИД 12365352 . Архивировано (PDF) из оригинала 29 июня 2011 г. Проверено 17 января 2017 г.
- ^ Прум, Ричард О.; Кисть, AH (март 2003 г.). «Что было первым: перо или птица?» (PDF) . Научный американец . Том. 288, нет. 3. С. 84–93. Бибкод : 2003SciAm.288c..84P . doi : 10.1038/scientificamerican0303-84 . ПМИД 12616863 . Архивировано (PDF) из оригинала 29 июня 2011 г. Проверено 17 января 2017 г.
- ^ Прум, Ричард О. (1999). «Развитие и эволюционное происхождение перьев» (PDF) . Журнал экспериментальной зоологии . Том. 285, нет. 4. С. 291–306. doi : 10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9 . ПМИД 10578107 . Архивировано (PDF) из оригинала 02 мая 2018 г. Проверено 12 апреля 2017 г.
- ^ Ли, Цюаньго (9 марта 2012 г.). «Реконструкция микрораптора и эволюция радужного оперения». Наука . Том. 335, нет. 6073. стр. 1215–1219. Бибкод : 2012Sci...335.1215L . дои : 10.1126/science.1213780 . ПМИД 22403389 . S2CID 206537426 .
- ↑ Перейти обратно: Перейти обратно: а б с Петтингилл 1985 , с. 29
- ↑ Перейти обратно: Перейти обратно: а б Чендлер 1916 , с. 285
- ^ Маклелланд, Дж. (1991). Цветной атлас анатомии птиц . WB Saunders Co. ISBN 978-0-7216-3536-1 .
- ^ Хубер-Эйхер, Б.; Себо, Ф. (2001). «Распространенность расклевывания перьев и их развитие в товарных стадах кур-несушек». Прикладная наука о поведении животных . Том. 74, нет. 3. С. 223–231. дои : 10.1016/S0168-1591(01)00173-3 .
- ^ Шервин, CM; Ричардс, Дж.Дж.; Никол, CJ (2010). «Сравнение благосостояния кур-несушек в четырех системах содержания в Великобритании». Британская наука о птицеводстве . Том. 51, нет. 4. С. 488–499. дои : 10.1080/00071668.2010.502518 . ПМИД 20924842 . S2CID 8968010 .
- ^ Батлер, округ Колумбия; Дэвис, К. (2010). «Влияние пластиковых насадок на состояние и поведение фазанов, выращенных в неволе». Ветеринарный учет . Том. 166, нет. 13. С. 398–401. дои : 10.1136/vr.b4804 . ПМИД 20348469 . S2CID 7549266 .
- ^ Шервин, CM (2010). «Индюки: поведение, управление и благополучие». В пруду, Уилсон Г.; Белл, Алан В. (ред.). Энциклопедия зоотехники . Марсель Деккер. стр. 847–849.
- ^ Густафсон, Лесли А.; Ченг, Хэн-Вэй; Гарнер, Джозеф П.; Пайорк, Эдмонд А.; Менч, Джой А. (2007). «Влияние обрезки клюва мускусных уток на поведение, прибавку массы тела и морфопатологию клюва». Прикладная наука о поведении животных . Том. 103, нет. 1–2. стр. 59–74. дои : 10.1016/j.applanim.2006.04.003 .
- ^ Райшль, Э.; Самбраус, HH (2003). «Клев перьев африканских страусов в Израиле». Ветеринарное обследование . Том 58, № 7. С. 364–369.
- ^ МакЭди, ТМ; Килинг, ЖЖ (2002). «Социальная передача клевания перьев у кур-несушек: влияние окружающей среды и возраста». Прикладная наука о поведении животных . Том. 75, нет. 2. С. 147–159. дои : 10.1016/S0168-1591(01)00182-4 .
- ^ Люмей, Йоханнес Т.; Хоммерс, Кэролайн Дж. (2008). «Обогащение собирательства как лечение птеротилломании» (PDF) . Прикладная наука о поведении животных . Том. 111, нет. 1–2. стр. 85–94. дои : 10.1016/j.applanim.2007.05.015 . Архивировано из оригинала (PDF) 23 ноября 2015 г. Проверено 20 декабря 2020 г.
- ^ «Ощипывание перьев у домашних птиц» . Красота птиц . 2011. Архивировано из оригинала 2 ноября 2019 года . Проверено 21 декабря 2020 г.
- ^ Стивс, Сьюзен А. (21 декабря 2005 г.). «Поведение попугаев отражает психические расстройства человека» . Университет Пердью. Архивировано из оригинала 10 апреля 2021 года . Проверено 21 декабря 2020 г.
- ^ ван Зеланд, Ивонн Р.А.; Спрут, Берри М.; Роденбург, Т. Басс; и др. (ноябрь 2009 г.). «Поведение попугаев, повреждающее перья: обзор с учетом сравнительных аспектов» . Прикладная наука о поведении животных . Том. 121, нет. 2. С. 75–95. дои : 10.1016/j.applanim.2009.09.006 . Архивировано из оригинала 28 мая 2023 г. Проверено 21 декабря 2020 г.
- ^ Шуаньер, Ж.; Моубрей Голдинг, К.; Везо, Т. (2005). Что это за птица? . Стори Паблишинг . п. 33 . ISBN 978-1-58017-554-8 .
- ^ Сибли и др. 2001 , с. 78
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Лэк, 1985 , с. 433
- ↑ Перейти обратно: Перейти обратно: а б О'Коннор, Р.Дж. (1984). Рост и развитие птиц . Уайли Интерсайенс . п. 174. ИСБН 978-0-471-90345-1 .
- ^ Риклефс, RE (1996). «Поведение молодых кактусовых крапивников и кривоклювых молотилок» (PDF) . Бюллетень Уилсона . Том. 78, нет. 1. С. 47–56. Архивировано из оригинала (PDF) 10 октября 2008 г.
- ^ Ницш, Кристиан Людвиг (1867). Птерилография Нича . Рэйское общество. п. 14.
- ^ Чендлер 1916 , с. 261
- ^ Проктор и Линч 1998 , с. 96
- ^ Чендлер 1916 , с. 253
- ^ Истман, Джон (2000). Птицы поля и берега: луга и береговые птицы восточной части Северной Америки . Книги Стэкпола. п. 13. ISBN 978-0-8117-2699-3 .
- ^ Стителер, Шэрон (2013). 1001 секрет, который должен знать каждый птицелов: советы и мелочи для заднего двора и за его пределами . Лондон: Бегущая пресса. п. 389. ИСБН 978-0-7624-4832-6 .
- ^ Гилл 1995 , с. 136 и далее.
- ^ Корни 2006 , с. различный
- ^ Макнаб, Брайан К. (октябрь 1994 г.). «Энергосбережение и эволюция нелетаемости птиц». Американский натуралист . Том. 144, нет. 4. С. 628–42. дои : 10.1086/285697 . JSTOR 2462941 .
- ^ Ковач, Кристофер Э.; Мейерс, Р.А. (2000). «Анатомия и гистохимия летательных мышц крылатой ныряющей птицы атлантического тупика Fratercula arctica ». Журнал морфологии . Том. 244, нет. 2. С. 109–25. doi : 10.1002/(SICI)1097-4687(200005)244:2<109::AID-JMOR2>3.0.CO;2-0 . ПМИД 10761049 .
- ^ Баумель, Джулиан Дж. (1993). Справочник по анатомии птиц: Nomina Anatomica Avium . Орнитологический клуб Наттолла.
- ^ Уитни, Уильям Дуайт (1906). Словарь и циклопедия Century . Запись "Перо". Компания Century Co., с. 2 , 164. OCLC 825679 .
- ^ Бакли, Пенсильвания (январь – декабрь 1966 г.). «Гребля ногами четырех американских чаек с комментариями о его возможной функции и стимуляции». Этология . Том. 23, нет. 4. С. 395–402. ПМИД 5992179 .
- ^ Уоллс, Гордон Л. (1942). «Адаптация к дневной активности» . Бюллетень: Глаз позвоночных и его адаптивное излучение . Научный институт Крэнбрука. п. 188.
- ^ Холден, Питер (2016). Птицы RSPB: их скрытый мир . Лондон: Издательство Блумсбери. п. 245. ИСБН 978-1-4729-3224-2 .
- ^ Элфик 2016 , с. 61
- ^ Гилл 1995 , с. 84
- ^ Дэвисон, GWH (ноябрь 1982 г.). «Заметки о вымершем Argusianus Bipuntatus» . Бюллетень Клуба британских орнитологов . Том. 103, нет. 3. п. 87. Архивировано из оригинала 5 августа 2017 г. Проверено 14 января 2018 г.
- ^ Виделер, Джон Дж. (2005). Птичий полет . ОУП Оксфорд. стр. 58–61. ISBN 978-0-19-929992-8 .
- ^ Кэмпбелл и Лак 1985 , с. 533
- ^ Гроу, Катрина Ван (2013). Неоперённая птица . Издательство Принстонского университета. п. 58. ИСБН 978-0-691-15134-2 .
- ^ Альвареза Ф., Санчес К.С., Ангуло С. (2005). «Лобный щит камышницы: половые различия и связь с состоянием тела». Этология, экология и эволюция . Том. 17, нет. 2. С. 135–148. CiteSeerX 10.1.1.158.6095 . дои : 10.1080/08927014.2005.9522603 . S2CID 85892241 .
- ^ Галлион, Гордон В. (1951). «Лобный щит американской лысухи». Бюллетень Уилсона . Том. 63, нет. 3. С. 157–166.
- ^ Гилл 1995 , стр. 134–136.
- ^ «Фуркула» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ↑ Перейти обратно: Перейти обратно: а б Ньюман, Кеннет Б. (2000). Птицы Ньюмана по цветам . Струйк. п. 14. ISBN 978-1-86872-448-2 .
- ^ Уилрайт, Северная Каролина (1985). «Размер плода, ширина зева и рацион птиц, питающихся фруктами» (PDF) . Экология . Том. 66, нет. 3. С. 808–818. дои : 10.2307/1940542 . JSTOR 1940542 . Архивировано из оригинала (PDF) 8 апреля 2016 г.
- ^ Зикефуз, Джули (25 июля 2014 г.). «Таинственные птицы двора» . Дайджест орнитолога. Архивировано из оригинала 13 декабря 2010 года . Проверено 29 января 2017 г.
- ^ Элфик 2016 , с. 71
- ^ Петтингилл 1985 , с. 77
- ^ Ньякупфука, Эндрю (3 апреля 2013 г.). Глобальные деликатесы: разнообразие, экзотика, странность, странность, релятивизм . Бальбоа Пресс. п. 120. ИСБН 978-1-4525-6791-4 .
- ^ Петерсон, Роджер ; Маунтфорт, Гай; Холлом, PAD (1954). Полевой справочник по птицам Британии и Европы . Лондон: Коллинз. п. 251.
- ^ Данн, Джон; Гарретт, Кимбалл (1997). Славки . Нью-Йорк: Полевые гиды Петерсона . стр. 18, 377, 437, 470, 551. ISBN. 978-0-395-78321-4 .
- ^ Сибли и др. 2001 , стр. 360–361, 377, 390, 404, 427, 435–436, 441, 451, 545.
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Лэк, 1985 , с. 254
- ^ Рассел, Питер Дж.; Вулф, Стивен Л.; Герц, Пол Э.; Старр, Сеси (2008). Биология: динамическая наука . Том. 2. Белмонт, Калифорния: Томсон Брукс/Коул. п. 1255. ИСБН 978-0-495-01033-3 .
- ^ Хауэлл, Стив Н.Г. (2002). Колибри Северной Америки: Фотопутеводитель . Академическая пресса. п. 1. ISBN 978-0-12-356955-4 .
- ^ Риджели, Роберт С.; Тюдор, Гай; Браун, Уильям Л. (1994). Птицы Южной Америки: воробьинообразные . Остин, Техас: Издательство Техасского университета. п. 771. ИСБН 978-0-292-77063-8 .
- ^ Кристенсен, Глен К. (1970). Чукарская куропатка: ее введение, история жизни и содержание . Рино, Невада: Департамент рыболовства и охоты штата Невада. п. 33.
- ^ Сандсторм, Боб (8 апреля 2015 г.). «Сияющие доспехи колибри» . BirdNote (Подкаст). Рассказывает Мэри Макканн. Примечание птицы. Архивировано из оригинала 27 декабря 2017 года . Проверено 26 декабря 2017 г.
- ^ Моррис, Кристофер Г.; Академическое издательство (1992). Академический словарь прессы по науке и технологиям . Профессиональное издательство Персидского залива. п. 971. ИСБН 978-0-12-200400-1 .
- ^ Дэй, Лесли; Рипе, Дон (2015). Полевой путеводитель по окрестным птицам Нью-Йорка . Джу Пресс. п. 36. ISBN 978-1-4214-1617-5 .
- ^ Ледерер, Роджер (2016). Клювы, кости и песни птиц: как борьба за выживание сформировала птиц и их поведение . Лесной Пресс. п. 157. ИСБН 978-1-60469-752-0 .
- ↑ Перейти обратно: Перейти обратно: а б Ловетт и Фитцпатрик, 2016 , с. 181.
- ^ Федучча 1999 , с. 13
- ^ Берд 2004 , с. 4
- ^ Одум, Юджин П.; Куэнцлер, Эдвард Дж. (1955). «Измерение территории и размеров ареала у птиц». Аук . Том. 72, нет. 2. С. 128–137. дои : 10.2307/4081419 . ISSN 0004-8038 . JSTOR 4081419 .
- ^ Элфик 2016 , стр. 135–136
- ^ Харестад, AS; Баннел, Флорида (1979). «Диапазон дома и масса тела — переоценка». Экология . Том. 60, нет. 2. С. 389–402. дои : 10.2307/1937667 . ISSN 0012-9658 . JSTOR 1937667 .
- ^ Петтингилл 1985 , стр. 98, 190.
- ↑ Перейти обратно: Перейти обратно: а б Элфик 2016 , стр. 25–26
- ^ Юнг, Джэ-Ён; Нэйлуэй, Стивен Э.; Яраги, Николас А.; Эррера, Стивен; Шерман, Винсент Р.; Бушонг, Эрик А.; Эллисман, Марк Х.; Кисаилус, Давид; МакКиттрик, Джоанна (2016). «Структурный анализ языка и подъязычного аппарата дятла». Акта Биоматериалы . Том. 37. стр. 1–13. doi : 10.1016/j.actbio.2016.03.030 . ISSN 1742-7061 . ПМК 5063634 . ПМИД 27000554 .
- ^ Пайкрафт, Уильям Плейн (1911). . В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 10 (11-е изд.). Издательство Кембриджского университета. п. 227.
- ^ Дюрден, JE (июль 1922 г.). «Происхождение перьев от чешуи рептилий» . Журнал Министерства сельского хозяйства . Том. 5, нет. 1. п. 67. Архивировано из оригинала 18 апреля 2017 г. Проверено 17 апреля 2017 г.
- ^ Скотт 2010 , с. 43
- ^ Циммер, Кевин Дж. (2000). Наблюдение за птицами на американском Западе: Справочник . Издательство Корнельского университета. п. 47. ИСБН 978-0-8014-8328-8 .
- ^ Макдональд, Дэвид (март 1996 г.). «Этимология слова Jizz» . Заметки о Канберрских птицах . Том. 21, нет. 1. стр. 2–11. Архивировано из оригинала 18 апреля 2016 г. Проверено 9 сентября 2017 г.
- ^ «Джизз» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Дули, Шон (2005). Большой Твич . Аллен и Анвин. п. 78. ИСБН 978-1-74114-528-1 .
- ^ Дэйв Мэдден (2 августа 2011 г.). Настоящее животное: внутри странного и захватывающего мира таксидермии . Пресса Святого Мартина. стр. 127–. ISBN 978-1-4299-8762-2 .
- ^ Хью Харроп (2004). Отчет о млекопитающих Шетландских островов, 2003 год . Группа млекопитающих Шетландского моря.
- ^ Эндрю Тайзак (30 июня 2013 г.). Рисование и раскрашивание насекомых . Кровуд Пресс, Лимитед. стр. 147–. ISBN 978-1-84797-625-3 .
- ^ Камминс, Джим (1 апреля 1996 г.). «Анатомия полета» . Архивировано из оригинала 11 февраля 2005 года . Проверено 14 марта 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Рамель, Гордон. «Анатомия птиц» . Веб-продукция Earth-Life. Архивировано из оригинала 7 февраля 2005 года . Проверено 14 марта 2017 г.
- ^ Элфик 2016 , с. 129
- ^ Фиске, Педер; Ринтамяки, Пекка Т.; Карвонен, Эви (январь 1998 г.). «Успех спаривания у токирующих самцов: метаанализ». Поведенческая экология . Том. 9, нет. 4. С. 328–338. дои : 10.1093/beheco/9.4.328 .
- ↑ Перейти обратно: Перейти обратно: а б Элфик 2016 , с. 151
- ^ Майр, Джеральд (2005). «Новая эоценовая мышевидная птица, похожая на Chascacocolius (Aves: Coliiformes), с замечательной зияющей адаптацией» (PDF) . Организмы, разнообразие и эволюция . Том. 5, нет. 3. С. 167–171. дои : 10.1016/j.ode.2004.10.013 . Архивировано (PDF) из оригинала 11 апреля 2016 г. Проверено 19 января 2017 г.
- ^ Кайзер, Гэри В. (2007). Внутренняя птица: анатомия и эволюция . Ванкувер, Британская Колумбия: UBC Press. п. 19. ISBN 978-0-7748-1343-3 .
- ^ Стоукс, Дональд В. и Стоукс, Лилиан К. (2003). Книга Стоукса о птицах на заднем дворе . Эммаус, Пенсильвания: Родейл. п. 141. ИСБН 1-57954-864-4 .
- ^ Кэмпбелл и Лак 1985 , с. 101.
- ^ Кэмпбелл и Лак 1985 , стр. 101–105.
- ^ Сибли и др. 2001 , стр. 55–59.
- ^ Кричер, Джон (2020). Справочное руководство Петерсона по поведению птиц . Хоутон Миффлин Харкорт. п. 20. ISBN 978-1-328-78736-1 .
- ^ Петтингилл 1985 , стр. 232–234.
- ^ Бертольд, Питер; Бауэр, Ганс-Гюнтер; Вестхед, Валери (2001). Миграция птиц: общий обзор . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-850787-1 .
- ^ Секерджиоглу, Швейцария (2007). «Экология сохранения: территория превосходит мобильность в результате исчезновения фрагментов птиц». Современная биология . Том. 17, нет. 8. стр. 283–286. дои : 10.1016/j.cub.2007.04.045 . ПМИД 1743770 .
- ^ Роллан, Джонатан; Жиге, Фредерик; Йонссон, Кнуд Андреас; Кондамин, Фабьен Л.; Морлон, Элен (2014). «Расселение сезонных мигрантов способствует диверсификации птиц». Труды Королевского общества Б. Том. 281, нет. 1784. с. 20140473. дои : 10.1098/rspb.2014.0473 . ПМК 4043101 . ПМИД 24759866 .
- ^ Ньютон, Ян (2008). Миграционная экология птиц . Эльзевир. ISBN 978-0-12-517367-4 .
- ^ Чан, Кен (январь 2001 г.). «Частичная миграция австралийских наземных птиц: обзор». Эму . Том. 101, нет. 4. С. 281–292. дои : 10.1071/MU00034 .
- ^ Хаммель, Д.; Бойкенберг, М. (1989). «Аэродинамические интерференционные эффекты в групповом полете птиц». Дж. Орнитол . Том 130. С. 15–24. дои : 10.1007/BF01647158 .
- ^ Каттс, CJ; Спикмен, младший (1994). «Энергосбережение при групповом полете розовоногих гусей» (PDF) . Дж. Эксп. Биол . Том. 189, нет. 1. С. 251–261. ПМИД 9317742 . Архивировано (PDF) из оригинала 31 октября 2008 г. Проверено 1 апреля 2017 г.
- ^ Лаборатория орнитологии Корнельского университета (март 1974 г.). «Запланированные поездки по наблюдению за птицами по штату, присоединяйтесь к Ньюэллу» . Ножницы-хвост . Том. 24. с. 32. ISSN 0582-2637 .
- ^ Биди, Тед; Пандольфино, Эд (2013). Птицы Сьерра-Невады: их естественная история, статус и распространение . Издательство Калифорнийского университета. п. 47. ИСБН 978-0-520-95447-2 .
- ^ Террес 1980 , стр. 616–617
- ^ Флойд 2008 , с. 497
- ^ Кинг и Маклелланд 1985 , с. 376
- ^ Эллиот, Даниэль Жиро (1898). Дикая птица Соединенных Штатов и британских владений . Нью-Йорк, штат Нью-Йорк: Ф. П. Харпер. п. XVIII. LCCN 98001121 .
- ^ Перринс, Кристофер М. (1974). Птицы . Лондон, Великобритания: Коллинз. п. 24. ISBN 978-0-00-212173-6 .
- ^ Петри, Чак (2006). Почему утки это делают: 40 характерных особенностей поведения уток, объясненных и сфотографированных . Миноква, Висконсин: Willow Creek Press. п. 31. ISBN 978-1-59543-050-2 .
- ^ Гудман, Дональд Чарльз; Фишер, Харви И. (1962). Функциональная анатомия пищевого аппарата водоплавающих птиц (Aves: Anatidae) . Карбондейл, Иллинойс: Издательство Университета Южного Иллинойса. п. 179. OCLC 646859135 .
- ^ Кинг и Маклелланд 1985 , с. 421
- ^ Кэмпбелл и Лак 1985 , с. 470
- ↑ Перейти обратно: Перейти обратно: а б Гилл 1995 , с. 108
- ^ Хоскер, Энн (1936). «Исследования эпидермальных структур птиц». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . Том. 226, нет. 533. стр. 143–188. Бибкод : 1936RSPTB.226..143H . дои : 10.1098/rstb.1936.0006 . ISSN 0080-4622 . JSTOR 92253 .
- ^ Берд 2004 , с. 281
- ^ Берд 2004 , с. 277
- ^ Кэмпбелл и Лак 1985 , с. 386
- ^ Кэмпбелл и Лак 1985 , с. 345
- ^ Скуч, Александр Ф (1960). «Гнездо как общежитие». Ибис . Том. 103, нет. 1. С. 50–70. дои : 10.1111/j.1474-919X.1961.tb02420.x .
- ^ Гиббон, Джонни (20 апреля 2015 г.). «Птичьи гнезда: разнообразие — ключ к успеху мировых авиационных архитекторов» . Смитсоновский институт. Архивировано из оригинала 3 января 2017 года . Проверено 7 февраля 2017 г.
- ^ Кэмпбелл и Лак 1985 , с. 387
- ^ Уильямс, Дэвид Л.; Флах, Э. (март 2003 г.). «Симблефарон с аберрантным выпячиванием мигательной перепонки у полярной совы ( Nyctea scandiaca )». Ветеринарная офтальмология . Том. 6, нет. 1. С. 11–13. дои : 10.1046/j.1463-5224.2003.00250.x . ПМИД 12641836 .
- ^ Гилл 1995 , с. 185
- ^ Петтингилл 1985 , с. 11
- ^ Хаубер, Марк Э. (1 августа 2014 г.). Книга яиц: путеводитель по яйцам шестисот видов птиц мира в натуральную величину . Чикаго: Издательство Чикагского университета. п. 31. ISBN 978-0-226-05781-1 . Архивировано из оригинала 24 февраля 2024 года . Проверено 14 июня 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б Гилл 1995 , с. 117
- ^ Уитни, Уильям Дуайт; Смит, Бенджамин Эли (1911). Словарь и циклопедия Century, Vol. 6 . Нью-Йорк: Компания Century. п. 4123. LCCN 11031934 .
- ^ Бок, Уолтер Дж. (1989). «Организмы как функциональные машины: объяснение связности». Американский зоолог . Том. 29, нет. 3. С. 1119–1132. дои : 10.1093/icb/29.3.1119 . JSTOR 3883510 .
- ^ Уивер 1981 , с. 83
- ^ Майр, Эрнст (1946). «Число видов птиц» (PDF) . Аук . Том. 63, нет. 1. С. 64–69. дои : 10.2307/4079907 . JSTOR 4079907 . Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 21 января 2017 г.
- ^ Кеннеди, Адам Скотт (2014). Птицы заповедника Серенгети и Нгоронгоро . Издательство Принстонского университета. п. 198. ИСБН 978-0-691-15910-2 . Архивировано из оригинала 28 мая 2023 г. Проверено 19 марта 2023 г.
- ^ Элфик 2016 , с. 77
- ^ Буш, Сара Э.; Вилла, Скотт М.; Бовес, Тан Дж.; Брюэр, Даллас; Белтхофф, Джеймс Р. (1 апреля 2012 г.). «Влияние морфологии клюва и стоп на эктопаразитов сипух» . Журнал паразитологии . Том. 98, нет. 2. С. 256–261. дои : 10.1645/GE-2888.1 . ПМИД 22010835 .
- ↑ Перейти обратно: Перейти обратно: а б Чеке, Манн и Аллен 2001 , с. 15.
- ^ Чеке, Манн и Аллен 2001 , с. 23.
- ^ Эванс, Мэтью Р. и Хэтчвелл, Би Джей (1992). «Экспериментальное исследование мужских украшений малахитовой нектарницы с алыми хохолками: I. Роль нагрудных пучков в территориальной защите» . Поведенческая экология и социобиология . 29 (6): 413–419. JSTOR 4600642 .
- ^ «GEOL 204 Летопись окаменелостей: Пернатые драконы: Происхождение птиц и птичьего полета» . Департамент геологии Университета Мэриленда. Архивировано из оригинала 17 марта 2017 года . Проверено 29 января 2017 г.
- ^ Элфик 2016 , с. 136
- ^ Проктор и Линч 1998 , с. 86
- ^ Атан, Мэтти Сью (2008). Путеводитель по попугаю-квакеру . Образовательная серия Бэррона. п. 57. ИСБН 978-0-7641-3668-9 .
- ^ Петтингилл 1985 , стр. 41–42.
- ↑ Перейти обратно: Перейти обратно: а б Ян Дж. Х. Дункан; Пенни Хокинс (2010). Благополучие домашней птицы и других птиц, содержащихся в неволе . Спрингер. стр. 84–85. ISBN 978-90-481-3649-0 .
- ^ Хадсон, Вашингтон (1910). Естественная история Хармсворта: полный обзор животного мира . Лондон: Дом кармелитов. п. 926.
- ^ «Оперение» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Петтингилл 1985 , с. 194
- ^ Розенберг, Дэниел К.; Гейст, Деннис; Харпп, Карен. «Плюмология Галапагосских островов» (PDF) . darwinfoundation.org . База данных коллекций Чарльза Дарвина Фонда Чарльза Дарвина. ISSN 1390-2830 . Архивировано из оригинала 17 марта 2016 года . Проверено 29 января 2017 г.
- ^ Эйххорн, изд. Манфред (2005). Специальный биологический словарь Лангеншайдта на английском языке: английский – немецкий, немецкий – английский (1-е изд.). Берлин [среди прочих]: Лангеншайдт. п. 537. ИСБН 978-3-86117-228-4 . [ постоянная мертвая ссылка ]
- ^ Элфик 2016 , с. 33
- ^ Подулка, Сэнди; Рорбо, Рональд В.; Бонни, Рик, ред. (2003). Курс домашнего обучения по биологии птиц, второе издание . Корнеллская лаборатория орнитологии. п. 55 (Глоссарий).
- ^ Кэмпбелл и Лак 1985 , с. 208
- ^ Можжевельник, Тони; Парр, Майк (2003). Попугаи: Путеводитель по попугаям мира . Лондон: Кристофер Хелм. п. 17. ISBN 978-0-7136-6933-6 .
- ^ Калстон, Ширли (2006). Аллергия на домашних животных?: Революционное руководство по жизни с любимыми животными . Нью-Йорк, штат Нью-Йорк: Бантам Делл. стр. 34–35 . ISBN 978-0-553-38367-6 .
- ↑ Перейти обратно: Перейти обратно: а б Ловетт и Фитцпатрик, 2016 , с. 126
- ^ Хауэлл, Стив Н.Г.; Льюингтон, Ян; Рассел, Уилл (16 февраля 2014 г.). Редкие птицы Северной Америки . Издательство Принстонского университета. п. 34. ISBN 978-1-4008-4807-2 .
- ^ Ханна, ДР (2005). Биология птиц . Издательство Дискавери. п. 109. ИСБН 978-81-7141-933-3 .
- ^ Вальтер, Бруно А. (2005). «Уход за сложными украшениями обходится дорого: свидетельство того, что уход за ними требует больших затрат». Поведенческая экология . Том. 16, нет. 1. С. 89–95. дои : 10.1093/beheco/arh135 .
- ^ Шоки, Мэтью Д.; Пиллаи, Шрикумар Р.; Хилл, Джеффри Э. (2003). «Химическая война? Влияние уропигиального масла на бактерии, разлагающие перья». Журнал птичьей биологии . Том. 34, нет. 4. С. 345–49. дои : 10.1111/j.0908-8857.2003.03193.x .
- ^ Эрлих, Пол Р. (1986). «Адаптативное значение Anting» (PDF) . Аук . Том. 103, нет. 4. с. 835. Архивировано из оригинала (PDF) 5 марта 2016 года.
- ^ Эрлих и др. 1994 , с. 219
- ^ Эрлих и др. 1994 , с. 79
- ^ Мюллер, Вернер; Патоне, Джаннино (1998). «Пропускаемость перьев по воздуху» (PDF) . Журнал экспериментальной биологии . Том. 201, нет. 18. стр. 2591–2599. ПМИД 9716511 .
- ↑ Перейти обратно: Перейти обратно: а б Дженни, Лукас; Винклер, Рафаэль (1994). Линька и старение европейских воробьинообразных . Лондон: Академическая пресса. п. 7. ISBN 978-0-12-384150-6 .
- ↑ Перейти обратно: Перейти обратно: а б с дель Хойо, Эллиотт и Саргатал 1992 , с. 37
- ↑ Перейти обратно: Перейти обратно: а б Кауфман, Кенн (1990). Продвинутое наблюдение за птицами . Бостон: Хоутон Миффлин. стр. 186 . ISBN 978-0-395-53376-5 .
- ^ Свенссон, Ларс; Грант, Питер Дж. (1999). Путеводитель Коллинза по птицам: наиболее полный путеводитель по птицам Британии и Европы . Лондон: ХарперКоллинз. п. 231. ИСБН 978-0-00-219728-1 .
- ^ Кристи, Томас Алерстам; перевод Дэвида А. (1993). Миграция птиц . Кембридж [Англия]: Издательство Кембриджского университета . п. 253. ИСБН 978-0-521-44822-2 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Элфик 2016 , с. 49
- ^ Холл, К.; Сюзанна, С. (2005). «У воробьинообразных с девятью первичными перьями девять или десять первичных перьев? Эволюция концепции». Журнал орнитологии . Том. 146, нет. 2. С. 121–126. дои : 10.1007/s10336-004-0070-5 . S2CID 36055848 .
- ^ Пикрафт, WP (1895). «К птерилографии гоацина ( Opisthocomus cristatus )». Ибис . Том. 37, нет. 3. С. 345–373. дои : 10.1111/j.1474-919X.1895.tb06744.x .
- ^ Бэрд, Спенсер Фуллертон; Брюэр, Томас Мэйо; Риджуэй, Роберт (1874). История птиц Северной Америки . Маленький, Браун. п. 554.
- ^ Чаттерджи, Санкар (2015). Возникновение птиц: 225 миллионов лет эволюции . Джу Пресс. стр. 114–115. ISBN 978-1-4214-1614-4 .
- ^ Ловетт и Фицпатрик 2016 , с. 177
- ^ Элфик 2016 , с. 117
- ↑ Перейти обратно: Перейти обратно: а б Петтингилл, 1985 , стр. 29–32.
- ↑ Перейти обратно: Перейти обратно: а б Гилл 1995 , с. 80
- ^ Лилли, Фрэнк Р. (ноябрь 1941 г.). «О развитии перьев». Биологические обзоры . Том. 17, нет. 3. п. 247. дои : 10.1111/j.1469-185X.1942.tb00439.x . S2CID 84592534 .
- ^ дель Хойо, Эллиотт и Саргатал 1992 , стр. 84–85, 91, 104
- ^ дель Ойо, Эллиот и Саргатал 1992 , стр. 141
- ^ Мэдж, Стив; Макгоуэн, Фил (2002). Фазаны, куропатки и тетерева . Лондон: Кристофер Хелм. п. 375. ИСБН 978-0-7136-3966-7 .
- ^ дель Ойо, Эллиот и Саргатал 1992 , стр. 105
- ^ Подулка, Сэнди; Рорбо, Рональд В.; Бонни, Рик, ред. (2003). Курс домашнего обучения по биологии птиц, второе издание . Итака, Нью-Йорк: Корнельская лаборатория орнитологии. п. 1.11.
- ^ След 2001 , с. 8
- ^ Моллер, Андерс Папе; Хоглунд, Джейкоб (1991). «Модели флуктуирующей асимметрии в орнаментах из птичьих перьев: значение для моделей полового отбора». Труды: Биологические науки . Том. 245, нет. 1312. стр. 1–5. Бибкод : 1991РСПСБ.245....1П . дои : 10.1098/rspb.1991.0080 . S2CID 84991514 .
- ^ Штреземанн, Эрвин (ноябрь – декабрь 1963 г.). «Вариации количества праймериз» (PDF) . Кондор . Том. 65, нет. 6. С. 449–459. дои : 10.2307/1365506 . JSTOR 1365506 . Архивировано (PDF) из оригинала 19 октября 2021 г. Проверено 5 февраля 2017 г.
- ^ Ловетт и Фицпатрик 2016 , с. 458
- ^ Террилл, Скотт Б.; Абель, Кеннет П. (январь 1988 г.). «Терминология миграции птиц» (PDF) . Аук . Том. 105. с. 206. дои : 10.1093/аук/105.1.205 . Архивировано (PDF) из оригинала 19 октября 2021 г. Проверено 19 марта 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Ледерер, Роджер Дж. «Роль птичьих риктальных щетинок» (PDF) . Бюллетень Уилсона . Том. 84, нет. 2. стр. 193–197. Архивировано (PDF) из оригинала 4 февраля 2014 г. Проверено 20 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Коновер, Майкл Р.; Миллер, Дон Э. (ноябрь 1980 г.). «Риктальная щетинка у ивовой мухоловки» (PDF) . Кондор . Том. 82, нет. 4. С. 469–471. дои : 10.2307/1367580 . JSTOR 1367580 . Архивировано (PDF) из оригинала 12 августа 2017 г. Проверено 20 января 2017 г.
- ^ Ледерер, Роджер Дж. (1972). «Роль птичьих щетинок» (PDF) . Бюллетень Уилсона . Том. 84. стр. 193–97. Архивировано (PDF) из оригинала 4 февраля 2014 г. Проверено 20 января 2017 г.
- ^ Харрис, Майк П. (2014). «Старение атлантических тупиков Fratercula arctica летом и зимой» (PDF) . Морская птица . Том. 27. С. 22–40. Архивировано из оригинала (PDF) 11 июня 2016 г.
- ^ «Информационный бюллетень о тупиках с острова Скомер» (PDF) . welshwildlife.org . Фонд дикой природы Южного и Западного Уэльса. Май 2011 г. Архивировано из оригинала (PDF) 19 октября 2020 г. Проверено 20 января 2017 г.
- ^ Гирлинг, Саймон (2003). Ветеринарный уход за экзотическими домашними животными . Оксфорд, Великобритания: Blackwell Publishing. п. 4. ISBN 978-1-4051-0747-1 .
- ^ Самур, Хайме, изд. (2000). Птичья медицина . Лондон, Великобритания: Мосби. п. 296. ИСБН 978-0-7234-2960-9 .
- ^ Бонс, Р.Х.; Виттер, Марк С. (1993). «Твердость кератина клюва европейского скворца» (PDF) . Кондор . Том. 95, нет. 3. С. 736–738. дои : 10.2307/1369622 . JSTOR 1369622 . Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 19 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Петтингилл 1985 , с. 8
- ^ Дамероу, Гейл (2012). Куриная энциклопедия: иллюстрированный справочник . Стори Паблишинг, ООО. п. 278. ИСБН 978-1-60342-561-2 .
- ^ Лукас, Альфред М. (1972). Анатомия птиц – покровы . Ист-Лансинг, Мичиган: Проект анатомии птиц Министерства сельского хозяйства США, Университет штата Мичиган. стр. 67, 344, 394–601.
- ^ Сойер, Р.Х., Кнапп, Л.В. 2003. Развитие кожи птиц и эволюционное происхождение перьев. J.Exp.Zool. (Мол.Дев.Эвол) 298Б:57–72.
- ↑ Перейти обратно: Перейти обратно: а б с Дуайи, Д. (апрель 2009 г.). «Новый сценарий эволюционного происхождения волос, перьев и птичьей чешуи». Журнал анатомии . Том. 214, нет. 4. С. 587–606. дои : 10.1111/j.1469-7580.2008.01041.x . ПМК 2736124 . ПМИД 19422430 .
- ↑ Перейти обратно: Перейти обратно: а б с Лукас, Альфред Мартин; Штеттенхайм, Питер Рич (1972). Птичья анатомия Покровы . Типография правительства США. п. 597.
- ^ Чжэн, X.; Чжоу, З.; Ван, X.; Чжан, Ф.; Чжан, X.; Ван, Ю.; Сюй, X. (2013). «Задние крылья базальных птиц и эволюция перьев ног». Наука . Том. 339, нет. 6125. стр. 1309–1312. Бибкод : 2013Sci...339.1309Z . CiteSeerX 10.1.1.1031.5732 . дои : 10.1126/science.1228753 . ПМИД 23493711 . S2CID 206544531 .
- ^ Кэмпбелл и Лак 1985 , с. 52
- ^ Скотт, С. Дэвид; Макфарланд, Кейси (2010). Перья птиц: Путеводитель по видам Северной Америки . Книги Стэкпола. п. 15. ISBN 978-0-8117-4217-7 .
- ^ Петтингилл 1985 , с. 39
- ^ Андерссон, Мальте Б. (1994). Половой отбор . Издательство Принстонского университета . п. 269. ИСБН 978-0-691-00057-2 . Архивировано из оригинала 19 января 2023 г. Проверено 15 марта 2017 г.
- ^ Бернс, Челси М.; Адамс, Дин К. (11 ноября 2012 г.). «Становиться разными, но оставаться похожими: закономерности диморфизма полового размера и формы клювов колибри». Эволюционная биология . Том. 40, нет. 2. С. 246–260. дои : 10.1007/s11692-012-9206-3 . ISSN 0071-3260 .
- ^ МакГроу, К.Дж.; Хилл, GE; Стради, Р.; Паркер, Р.С. (2002). «Влияние пищевых каротиноидов на половой дихроматизм и пигментный состав оперения американского щегла» (PDF) . Сравнительная биохимия и физиология. Часть B-Биохимия и молекулярная биология . Том. 131, нет. 2. С. 261–269. дои : 10.1016/S1096-4959(01)00500-0 . ПМИД 11818247 . Архивировано из оригинала (PDF) 28 августа 2005 г.
- ^ Оуэнс, IPF; Хартли, ИК (1998). «Половой диморфизм у птиц: почему так много разных форм диморфизма?». Труды Королевского общества Б. Том. 265, нет. 1394. стр. 397–407. дои : 10.1098/rspb.1998.0308 . JSTOR 50849 . ПМК 1688905 .
- ^ Фридрих II (император Священной Римской империи) (1943). Вуд, Кейси Альберт; Файф, Флоренс Марджори (ред.). De Arte Venardi Cum Avibus: Быть De Arte Venandi Cum Avibus Фридриха II Гогенштауфена [ Искусство соколиной охоты ]. Издательство Стэнфордского университета. п. 72. ИСБН 978-0-8047-0374-1 .
- ^ Купер, Дж. Г. (1870). Бэрд, Сан-Франциско (ред.). Орнитология . Геологическая служба Калифорнии, Дж. Д. Уитни, государственный геолог. п. 570. ИСБН 978-5-87984-211-1 .
- ^ Риджели, Роберт С.; Гвинн, Джон А. младший (1989). Птицы Панамы с Коста-Рикой, Никарагуа и Гондурасом . Издательство Принстонского университета. ISBN 978-0-691-08529-6 .
- ^ Рэнд, Алабама (июнь 1954 г.). «На шпорах на птичьих крыльях» (PDF) . Бюллетень Уилсона . Том. 66, нет. 2. С. 127–134. Архивировано (PDF) из оригинала 3 марта 2016 г. Проверено 24 апреля 2017 г.
- ^ Петтингилл 1985 , с. 56
- ^ Данн, Джон Л.; Альдерфер, Джонатан, ред. (2006). Полевой справочник по птицам Северной Америки . Вашингтон, округ Колумбия: Национальное географическое общество. п. 10. ISBN 978-0-7922-5314-3 .
- ^ Ленды 1980 , с. 995
- ^ Ларсен, Онтарио; Голлер, Франц (2002). «Прямое наблюдение за функцией сирингеальных мышц у певчих птиц и попугая» . Журнал экспериментальной биологии . Том. 205, нет. Ч. 1. стр. 25–35. ПМИД 11818409 . Архивировано из оригинала 22 декабря 2015 г. Проверено 22 января 2017 г.
- ^ Сазерс, РА (октябрь 1990 г.). «Вклад в пение птиц с левой и правой стороны неповрежденного сиринкса». Природа . Том. 347, нет. 6292. стр. 473–477. Бибкод : 1990Natur.347..473S . дои : 10.1038/347473a0 .
- ^ Кейд, Томас Джозеф (1982). Соколы мира . Итака, Нью-Йорк: Комсток/Издательство Корнельского университета. п. 13. ISBN 978-0-8014-1454-1 .
- ^ Мёллер, АП; Барбоза, А.; Куэрво, Джей Джей; Лопе, Ф.Д.; Мерино, С.; Сайно, Н. (1998). «Половой отбор и хвостовые стримеры у ласточки». Труды Королевского общества B: Биологические науки . Том. 265, нет. 1394. стр. 409–414. дои : 10.1098/rspb.1998.0309 . ISSN 0962-8452 . ПМЦ 1688894 .
- ^ Матиясяк, Петр; Яблонский, Петр Г. (2001). «Гипотетические механизмы начальной эволюции сексуально диморфных хвостовых стримеров у Hirundinidae». Эволюция . Том. 55, нет. 2. п. 446. doi : 10.1554/0014-3820(2001)055[0446:HMOTIE]2.0.CO;2 . ISSN 0014-3820 . ПМИД 11308101 .
- ^ Бьюкенен, КЛ (2000). «Влияние длины хвостовой ленты на аэродинамические характеристики ласточки». Поведенческая экология . Том. 11, нет. 2. С. 228–238. дои : 10.1093/beheco/11.2.228 . ISSN 1465-7279 .
- ^ Фаулер, Д.В.; Фридман, Э.А.; Сканнелла, Дж. Б. (2009). Пиццари, Том (ред.). «Хищная функциональная морфология хищников: межпальцевые вариации размера когтей связаны с техникой удержания добычи и иммобилизации». ПЛОС ОДИН . Том. 4, нет. 11. стр. e7999. Бибкод : 2009PLoSO...4.7999F . дои : 10.1371/journal.pone.0007999 . ПМЦ 2776979 . ПМИД 19946365 .
- ^ Фейн, Мэтью Г.; Уд, Питер (2004). «Параллельные излучения в первичных кладах птиц» (PDF) . Эволюция . Том. 58, нет. 11. С. 2558–2573. дои : 10.1554/04-235 . ПМИД 15612298 . Архивировано из оригинала (PDF) 9 июля 2017 года.
- ^ Паркер, В.К. (1891). «О морфологии рептилии-птицы Opisthocomus hoazin » . Труды Лондонского зоологического общества . Том. 13, нет. 2. стр. 43–89. дои : 10.1111/j.1096-3642.1891.tb00045.x . Архивировано из оригинала 01 июля 2017 г. Проверено 21 февраля 2018 г.
- ^ Сэр Уолтер Лоури Буллер (1888): История птиц Новой Зеландии. из Лондона. Отрывок Архивировано 27 июля 2011 г. в коллекции Wayback Machine из коллекции Электронного текстового центра Зеландии.
- ^ Ньютон и др. 1899 , с. 948
- ^ Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс. стр. 205–208. ISBN 978-0-03-910284-5 .
- ^ Петтингилл 1985 , с. 33
- ^ Проктор и Линч 1998 , с. 54
- ^ Фергюсон-Лис, Джеймс; Кристи, Дэвид А. (2001). Хищники мира . Лондон: Кристофер Хелм. п. 27. ISBN 978-0-7136-8026-3 .
- ^ Хикман, Скотт (2008). «Проблема с терциалами». Аук . Том. 125, нет. 2. п. 493. дои : 10.1525/аук.2008.2408 .
- ^ Бергер, Эй Джей; Ланк, Вашингтон (1954). «Птерилез птенца Coua ruficeps» (PDF) . Бюллетень Уилсона . Том. 66, нет. 2. С. 119–126. Архивировано (PDF) из оригинала 8 августа 2022 г. Проверено 16 января 2017 г.
- ^ Карнаби 2008 , с. 11
- ^ Шодде, Р; Мейсон, Эй-Джей (1999). Справочник австралийских птиц: Воробьиные: Воробьиные . Издательство Csiro. п. 1122. ИСБН 978-0-643-10293-4 .
- ^ Класинг, Кирк К. (1999). «Анатомия и физиология желудочно-кишечного тракта птиц». Семинары по медицине птиц и экзотических домашних животных . Том. 8, нет. 2. С. 42–50. дои : 10.1016/S1055-937X(99)80036-X .
- ^ Госнер, Кеннет Л. (июнь 1993 г.). «Скопат Томия: приспособление для обращения с добычей с твердым панцирем?» (PDF) . Бюллетень Уилсона . Том. 105, нет. 2. С. 316–324. Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 17 января 2017 г.
- ^ Орнелас, Хуан Франциско. «Зубчатая Томия: адаптация к краже нектара у колибри?» (PDF) . Аук . Том. 111, нет. 3. С. 703–710. Архивировано (PDF) из оригинала 5 марта 2016 г. Проверено 17 января 2017 г.
- ^ Мэдж, Стив; Берн, Хилари (1988). Дикая птица . Лондон: Кристофер Хелм. стр. 143–144. ISBN 978-0-7470-2201-5 .
- ^ Шлейхер, Эльке (2004). «Оцепенение птиц: систематика, энергетика и экология». Физиологическая и биохимическая зоология . Том. 77, нет. 6. С. 942–949. дои : 10.1086/423744 . ПМИД 15674768 .
- ^ Бригам, Р. Марк (1992). «Ежедневное оцепенение у козососа, обыкновенного бедняги (Phalaenoptilus nuttallii)». Физиологическая зоология . Том. 65, нет. 2. С. 457–472. дои : 10.1086/physzool.65.2.30158263 . JSTOR 30158263 .
- ^ «Подкрылок» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ «Размах крыльев». Информационный бюллетень РАОУ . Том. 21, нет. 4. 2011. с. 17. ОСЛК 23961486 .
- ↑ Перейти обратно: Перейти обратно: а б Уилок, Ирен Гросвенор (1904). Птицы Калифорнии: знакомство с более чем 300 обычными птицами штата и прилегающих островов . Чикаго: AC McClurg. п. 15. ОСЛК 2064174 .
- ^ «Словарь птичьих терминов» . www.nebraskabirdlibrary.org . Архивировано из оригинала 27 марта 2017 года . Проверено 26 марта 2017 г.
- ^ Гилл 1995 , с. 102
- ^ «бродяга» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Альдерфер, Джонатан К. (2012). Библия орнитолога: полная сокровищница: наука, ноу-хау, красота, знания . Нэшнл Географик. п. 32. ISBN 978-1-4262-0964-2 .
- ^ Кан, Синтия М. (2007). Руководство Merck/Merial по здоровью домашних животных: полный справочник по здоровью вашей собаки, кошки, лошади или других домашних животных – изложенный повседневным языком . Саймон и Шустер. п. 849. ИСБН 978-0-911910-99-5 .
- ^ Шервин, CM, (2010). Благосостояние и этическая оценка помещений для производства яиц. В книге «Благосостояние домашней птицы и других содержащихся в неволе птиц» , И. Дж. Х. Дункан и П. Хокинс (редакторы), Springer, стр. 237–258.
- ^ Поцш, CJ; Льюис, К.; Никол, CJ; Грин, Л.Е. (2001). «Поперечное исследование распространенности клева у кур-несушек в альтернативных системах и его связи с клеванием перьев, лечением и болезнями». Прикладная наука о поведении животных . Том. 74, нет. 4. С. 259–272. дои : 10.1016/S0168-1591(01)00167-8 .
- ^ «Плетень» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Круикшанк, Аллан Д.; Круикшанк, Хелен Гир (1976). Ответы на 1001 вопрос о птицах . Курьерская корпорация. п. 97. ИСБН 978-0-486-23315-4 .
- ^ Кочан, Джек Б. (сентябрь 1995 г.). Счета и рты . Книги Стэкпола. п. 12. ISBN 978-0-8117-3057-0 .
- ^ Старк, Артур Коуэлл (1900). Птицы Южной Африки Том. 1 . Том. 1. Лондон: Р. Х. Портер. п. 23. Архивировано из оригинала 28 сентября 2018 г. Проверено 27 сентября 2018 г.
- ^ Кричер, Джон (18 февраля 2015 г.). Неотропический спутник: введение в животных, растения и экосистемы тропиков Нового Света. Иллюстрировано Андреа С. ЛеЖён . Издательство Принстонского университета. п. 273. ИСБН 978-1-4008-6691-5 .
- ^ Огилви, Массачусетс; Уоллес, DIM (1975). «Полевая идентификация серых гусей». Британские птицы . Том. 68. стр. 57–67.
- ^ Серстемон, Сандрин (25 января 2012 г.). «Гусь, летящий вверх ногами, снят в замедленном фильме» . Новый учёный ТВ. Архивировано из оригинала 17 августа 2017 года . Проверено 19 апреля 2017 г.
- ^ Аттенборо, Дэвид (24 сентября 1998 г.). Жизнь птиц . Би-би-си. п. 49. ИСБН 978-0-563-38792-3 .
- ^ Данн, Джон Ллойд; Альдерфер, Джонатан К. (2006). Полевой справочник National Geographic по птицам Северной Америки . Вашингтон, округ Колумбия: National Geographic Books. п. 12. ISBN 978-0-7922-5314-3 .
- ^ Брукс, МЫ; Этава, CE (1872 г.). Почетные секретари (ред.). Труды Азиатского общества Бенгалии: 1872 г. - Брукс об орлах-могильниках Индии . Калькутта: CB Lewis, Baptist Mission Press. п. 65.
- ^ Флойд 2008 , с. 498
- ^ Джонс, Алан К. «Подрезка крыльев у домашних птиц - исследование и сравнение методов» . Общество попугаев Великобритании. Архивировано из оригинала 16 января 2021 года . Проверено 21 декабря 2020 г.
- ^ Гесс, Лори, DVM (27 февраля 2017 г.). «Как подрезать птичке крылья» . ПетМД . Архивировано из оригинала 29 октября 2020 года . Проверено 22 декабря 2020 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Спир, Брайан (2015). Современная терапия в птичьей медицине и хирургии . Elsevier Науки о здоровье. п. 700. ИСБН 978-0-323-24367-4 .
- ^ Баумель, Дж. Дж. (1993) Справочник по анатомии птиц: Nomina Anatomica Avium . 2-е изд. Орнитологический клуб Наттолла. Кембридж, Массачусетс
- ^ Федучча 1999 , стр. 16–18.
- ^ «Словарь терминов по птицам от А до Я» . 9 сентября 2016. Архивировано из оригинала 20 марта 2017 года . Проверено 19 марта 2017 г.
- ^ Элфик, Джонатан (2007). Атлас миграции птиц: прослеживание великих путешествий птиц мира . Струйк. стр. 154–155. ISBN 978-1-77007-499-6 . [ постоянная мертвая ссылка ]
- ^ Уильямс, Эрнест Х. Уильямс младший (2005). Справочник природы: Руководство по наблюдению за природой . Издательство Оксфордского университета. п. 94. ИСБН 978-0-19-972075-0 .
- ^ Куэ 1890 , с. 187
Библиография [ править ]
- Берд, Дэвид Майкл (2004). Птичий альманах: Путеводитель по основным фактам и цифрам о птицах мира . Книги Светлячка. ISBN 978-1-55297-925-9 .
- Кэмпбелл, Брюс; Лэк, Элизабет, ред. (1985). Словарь птиц . Карлтон, Англия: Т. и А.Д. Пойзер. ISBN 978-0-85661-039-4 .
- Карнаби, Тревор (2008). Битва вокруг Буша: Птицы . Йоханнесбург: Джакана Медиа. ISBN 978-1-77009-241-9 .
- Чендлер, Аса К. (1916). Изучение строения перьев с учетом их таксономического значения . Беркли, Калифорния: Издательство Калифорнийского университета.
- Чик, Роберт А.; Манн, Клайв Ф. и Аллен, Ричард (2001). Солнечные птицы: Путеводитель по несолнечным птицам, цветогрызам, охотникам за пауками и сахарным птицам мира . Лондон: Кристофер Хелм. ISBN 978-1-873403-80-8 .
- Куэс, Эллиотт (1890). Справочник по полевой и общей орнитологии . Лондон: Macmillan and Co. OCLC 263166207 .
- дель Ойо, Джозеф; Эллиотт, Эндрю; Саргатал, Джорди, ред. (1992). Справочник птиц мира, Том 1: от страуса до уток . Барселона: Lynx Edicions. ISBN 978-84-87334-10-8 .
- Эрлих, Пол Р .; Добкин, Дэррил А.; Сыворотка, Дэррил; Пимм, Стюарт Л. (1994). Справочник орнитолога . Издательство Оксфордского университета. ISBN 978-0-19-858407-0 .
- Элфик, Джонатан (2016). Птицы: полное руководство по их биологии и поведению . Буффало, Нью-Йорк: Firefly Books. ISBN 978-1-77085-762-9 .
- Федучча, Алан (1999). Происхождение и эволюция птиц . Издательство Йельского университета. ISBN 978-0-300-07861-9 .
- Флойд, Тед (2008). Смитсоновский полевой справочник по птицам Северной Америки . ХарперКоллинз. ISBN 978-0-06-112040-4 .
- Гилл, Фрэнк Б. (1995). Орнитология (2-е изд.). Нью-Йорк, штат Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-2415-5 .
- Хауэлл, Стив Н.Г. (2007). Чайки Америки . Нью-Йорк: Компания Houghton Mifflin. ISBN 978-0-618-72641-7 .
- Кинг, Энтони Стюарт; Маклелланд, Джон, ред. (1985). Форма и функции у птиц, том 3 . Лондон, Великобритания: Академическая пресса. ISBN 978-0-12-407503-0 .
- Ловетт, Ирби Дж.; Фитцпатрик, Джон В. (2016). Справочник по биологии птиц . Уайли. ISBN 978-1-118-29104-7 .
- Ньютон, Альфред; Гадов, Ганс Фридрих; Лидеккер, Ричард; Рой, Чарльз Смарт; Шуфельдт, Роберт Уилсон (1899). Словарь птиц . Лондон: А. и К. Блэк. OCLC 623393918 .
- Мобли, Джейсон А.; Брюэр, Дункан; Элфик, Джонатан; Хоар, Бен; Анвин, Майк; Вудворд, Джон; и др. (2008). Птицы мира . Тэрритаун, Нью-Йорк: Маршалл Кавендиш. ISBN 978-0-7614-7775-4 .
- Петтингилл, Олин Сьюэлл младший (1985). Орнитология в лабораторных и полевых условиях . Орландо, Флорида: Academic Press. ISBN 978-0-12-552455-1 .
- Петтингилл, Олин Сьюэлл младший (2013). Орнитология в лабораторных и полевых условиях . Эльзевир. ISBN 978-1-4832-6311-3 .
- Проктор, Ноубл С.; Линч, Патрик Дж. (1998). Руководство по орнитологии: строение и функции птиц . Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 978-0-300-07619-6 .
- Корни, Клайв (2006). Нелетающие птицы . Вестпорт, Коннектикут: Издательская группа Greenwood. ISBN 978-0-313-08394-5 .
- Скотт, Грэм (2010). Основная орнитология . Издательство Оксфордского университета. ISBN 978-0-19-856997-8 .
- Сибли, Дэвид ; и др. (2001). Путеводитель Сибли по жизни и поведению птиц . Лондон: Кристофер Хелм. ISBN 978-0-7136-6250-4 .
- Террес, Дж. К. (1980). Энциклопедия птиц Северной Америки Общества Одюбона . Нью-Йорк, штат Нью-Йорк: Кнопф. ISBN 978-0-394-46651-4 .
- След, Пеппер (2001). «Перья крыльев» (PDF) . Служба охраны рыбы и дикой природы США. Архивировано из оригинала (PDF) 30 сентября 2017 года . Проверено 5 августа 2017 г.
- Виникомб, Кейт (2014). Руководство Helm по идентификации птиц . Лондон: Издательство Блумсбери. ISBN 978-1-4729-0554-3 .
- Уивер, Питер (1981). Словарь орнитолога . Лондон: Издательство Блумсбери. ISBN 978-1-4081-3852-6 .