Перо

Перья — это эпидермальные наросты, которые образуют характерный внешний покров или оперение (птиц), так и у как у птиц некоторых нептичьих динозавров и других архозавров . Это наиболее сложные покровные структуры, встречающиеся у позвоночных. [1] [2] и пример сложной эволюционной новизны. [3] Они относятся к числу характеристик, отличающих ныне живущих птиц от других ныне живущих групп. [4]
Хотя перья покрывают большую часть тела птицы, они возникают только из определенных четко выраженных участков кожи. Они помогают в полете, теплоизоляции и гидроизоляции. Кроме того, окраска помогает в общении и защите . [5] Изучение перьев называется плюмологией (или наукой о оперении ). [6] [7]
Люди используют перья во многих практических, культурных и религиозных целях. Перья мягкие и отлично удерживают тепло ; высокого класса таким образом, их иногда используют в постельных принадлежностях , особенно в подушках , одеялах и матрасах . Их также используют в качестве наполнителя для зимней одежды и уличных постельных принадлежностей, таких как стеганые пальто и спальные мешки . Гусиный и гагачий пух обладают большой высотой , способностью расширяться из сжатого, хранящегося состояния, чтобы улавливать большое количество разделенного на отсеки изолирующего воздуха. [8] Перья крупных птиц (чаще всего гусей ) использовались и используются для изготовления перьевых ручек. Исторически сложилось так, что охота на птиц ради декоративных перьев ставила под угрозу исчезновение одних видов и способствовала исчезновению других. [9] Сегодня перья, используемые в моде, а также в военных головных уборах и одежде, получают как отходы птицеводства, в том числе кур , гусей , индеек , фазанов и страусов . Эти перья красят и обрабатывают, чтобы улучшить их внешний вид, поскольку перья домашней птицы от природы часто бывают тусклыми по сравнению с перьями диких птиц.
Этимология
«Перо» происходит от древнеанглийского «feþer», имеющего германское происхождение; связано с голландским «veer» и немецким «Feder», от индоевропейского корня, общего с санскритским «patra», означающим «крыло», латинским «penna», означающим «перо», и греческим «pteron», «pterux», означающим «крыло». '.
Поскольку перья являются неотъемлемой частью перьев , которые раньше использовались для письма, само слово «перо» происходит от латинского penna , что означает перо. [10] Французское слово «плюм» может означать перо , перо или перо .
Структуры и характеристики

- У них есть
- Вал, рахис
- Барб
- Последний вал, постперо
- Перо, аир


Перья являются одними из самых сложных покровных придатков, обнаруженных у позвоночных , и образуются в крошечных фолликулах в эпидермисе или наружном слое кожи, которые производят кератиновые белки . β -кератины в перьях, клюве и когтях , а также когтях, чешуе и панцирях рептилий структуры, даже более прочные , состоят из белковых нитей, связанных водородными связями в β-складчатые листы , которые затем скручиваются и сшиваются дисульфидными мостиками в чем α -кератины млекопитающих волос , рогов и копыт . [11] [12] Точные сигналы , вызывающие рост перьев на коже, неизвестны, но было обнаружено, что фактор транскрипции cDermo-1 индуцирует рост перьев на коже и чешуи на ногах. [13]
Классификация
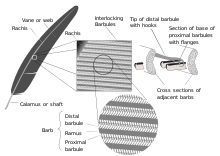
Существует два основных типа перьев: лопастные перья, покрывающие внешнюю часть тела, и пуховые перья , находящиеся под лопастными перьями. Перистые перья представляют собой лопастные перья. Перистые перья, также называемые контурными перьями, возникают из пучков и покрывают все тело. Третий, более редкий тип перьев, нитевидные перья, похожи на волосы, тесно связаны с перистыми перьями и часто полностью скрыты ими, при этом один или два нитевидных пера прикреплены и прорастают почти из той же точки кожи, что и каждое перистое перо, по крайней мере. на голове, шее и туловище птицы. [14] [15] полностью отсутствуют Филоплюмы у бескилевых . [16] У некоторых воробьинообразных нитевидные перья выходят за пределы перистых перьев на шее. [1] Ремиги, или маховые перья крыла, и прямоугольники, или маховые перья хвоста, являются наиболее важными перьями для полета. Типичное перо имеет основной стержень, называемый рахисом . К позвоночнику срослись ряд ветвей или зазубрин ; сами бородки также разветвлены и образуют бородки . У этих усиков есть крошечные крючки, называемые бородками, для перекрестного прикрепления. Пуховые перья пушистые, потому что в них отсутствуют усики, поэтому усики плавают свободно друг от друга, позволяя пуху задерживать воздух и обеспечивать отличную теплоизоляцию. У основания пера рахис расширяется, образуя полый трубчатый аир (или иголку ), который прикрепляется к фолликулу на коже . Базальная часть аира без лопастей. Эта часть встроена в кожный фолликул и имеет отверстие у основания (проксимальный отдел пупка) и небольшое отверстие сбоку (дистальный отдел пупка). [17]
У вылупившихся птиц некоторых видов есть особый вид натальных пуховых перьев (неоссоптили), которые выталкиваются при появлении нормальных перьев (телеоптилей). [1]
Маховые перья усилены, чтобы работать против воздуха при движении вниз, но поддаются в других направлениях. Обнаружено, что характер ориентации волокон β-кератина в перьях летающих птиц отличается от такового у нелетающих птиц: волокна лучше ориентированы вдоль оси стержня в направлении кончика, [18] [19] а на боковых стенках области позвоночного столба имеется структура перекрещенных волокон. [20] [21]
Функции
Перья защищают птиц от воды и низких температур. Их также можно ощипать, чтобы выстелить гнездо и обеспечить изоляцию яиц и птенцов. Отдельные перья крыльев и хвоста играют важную роль в управлении полетом. [20] У некоторых видов имеется гребень на голове из перьев. Хотя перья легкие, оперение птицы весит в два-три раза больше ее скелета, так как многие кости полые и содержат воздушные мешки. Цветовые узоры служат камуфляжем для птиц в их среде обитания и служат камуфляжем для хищников, ищущих еду. Как и в случае с рыбами, цвета верхней и нижней части могут быть разными, чтобы обеспечить маскировку во время полета. Поразительные различия в рисунке и окраске перьев являются частью полового диморфизма многих видов птиц и особенно важны при выборе пар для спаривания. В некоторых случаях существуют различия в отражательной способности перьев УФ-излучения у разных полов, хотя в видимом диапазоне различий в цвете не наблюдается. [22] Крыльевые перья самцов булавокрылых манакинов Machaeropterus deliciosus имеют особые структуры, которые используются для воспроизведения звуков путем стридуляции . [23]

У некоторых птиц есть запас пуховых перьев, которые растут непрерывно, при этом мелкие частицы регулярно откалываются от концов усиков. Эти частицы образуют порошок , который просачивается через перья на теле птицы и действует как гидроизоляционный агент и кондиционер для перьев . Пуховый пух развился независимо у нескольких таксонов и встречается как в пухе, так и в перьевых перьях. Они могут быть разбросаны по оперению, как у голубей и попугаев, или в виде локальных пятен на груди, животе или боках, как у цапель и лягушачьих пасть. Цапли используют свой клюв, чтобы разбить порошок на перья и распределить его, а какаду могут использовать голову как пуховку для нанесения порошка. [24] Водонепроницаемость может быть потеряна из-за воздействия эмульгаторов из-за загрязнения окружающей среды человеком. Перья могут намокнуть, в результате чего птица утонет. Также очень сложно чистить и спасать птиц, перья которых были загрязнены разливами нефти . Перья бакланов впитывают воду и помогают снизить плавучесть, тем самым позволяя птицам плавать под водой. [25]

Щетинки — жесткие, сужающиеся перья с большим стержнем, но с небольшим количеством зазубрин. риктальные щетинки Вокруг глаз и клюва расположены . Они могут служить той же цели, что ресницы и вибриссы млекопитающих и . Хотя четких доказательств пока нет, было высказано предположение, что риктальные щетинки выполняют сенсорные функции и могут помогать насекомоядным птицам ловить добычу. [26] В одном исследовании было обнаружено, что ивовые мухоловки ( Empidonax Traillii ) одинаково хорошо ловят насекомых до и после удаления приктальных щетинок. [27]
Поганки имеют необычную привычку заглатывать собственные перья и скармливать ими своих детенышей. Наблюдения за рыбным рационом и частотой поедания перьев позволяют предположить, что проглатывание перьев, особенно с боков, способствует образованию легко выбрасываемых гранул. [28]
Распределение

Контурные перья распределены по коже птицы неравномерно, за исключением некоторых групп, таких как пингвины , бескилевые и крикуны. [29] У большинства птиц перья растут из определенных участков кожи, называемых птерилями ; между птерилями имеются участки, свободные от перьев, называемые аптерилами (или аптериями ). Из аптерилей могут возникать нитевидные перья и пух. Расположение этих перьевых трактов, птерилез или птерилография, варьируется в зависимости от семейства птиц и в прошлом использовалось как средство определения эволюционных взаимоотношений семейств птиц. [30] [31] Виды, высиживающие собственные яйца, часто теряют перья на части живота, образуя участок для высиживания яиц . [32]
Окраска

Слева: турацин (красный) и тураковердин (зеленый, с некоторой структурной синей переливчатостью на нижнем конце) на крыле Tauraco Bannermani.
Справа: каротиноиды (красные) и меланины (темные) на брюхе/крыльях Ramphocelus bresilius.
Цвет перьев создается пигментами, микроскопическими структурами, которые могут преломлять , отражать или рассеивать определенные длины волн света или комбинировать то и другое.
Большинство пигментов перьев представляют собой меланины (коричневые и бежевые феомеланин , черный и серый эумеланин ) и каротиноиды (красный, желтый, оранжевый); другие пигменты встречаются только в определенных таксонах - от желтого до красного пситтакофульвина. [33] (обнаружен у некоторых попугаев ), а также красный турацин и зеленый тураковердин ( порфириновые пигменты, обнаруженные только у турако ).
Структурная окраска [5] [34] [35] участвует в создании синих цветов, радужности , большей части отражения ультрафиолета и в усилении пигментных цветов. Сообщалось о структурной радужности [36] в ископаемых перьях возрастом 40 миллионов лет. Белые перья лишены пигмента и рассеивают свет рассеянно; альбинизм у птиц вызван дефектной выработкой пигмента, хотя структурная окраска не изменяется (как это можно видеть, например, у сине-белых волнистых попугаев ).

Синий и ярко-зеленый цвет у многих попугаев возникает в результате конструктивной интерференции света, отражающегося от разных слоев структур перьев. В случае зеленого оперения, помимо желтого, некоторые называют специфическую структуру перьев текстурой Дайка. [37] [38] Меланин часто участвует в поглощении света; в сочетании с желтым пигментом дает тусклый оливково-зеленый цвет.

У некоторых птиц цвет перьев может создаваться или изменяться за счет выделений уропигиальной железы , также называемой прихорашивающейся железой. Желтый цвет клюва многих птиц-носорогов образуется в результате таких выделений. Было высказано предположение, что существуют и другие цветовые различия, которые можно увидеть только в ультрафиолетовой области. [24] но исследования не смогли найти доказательств. [39] Масляный секрет уропигиальной железы также может оказывать ингибирующее действие на перьевые бактерии. [40]
Красный, оранжевый и желтый цвет многих перьев обусловлен различными каротиноидами. Пигменты на основе каротиноидов могут быть честным сигналом о физической форме, поскольку они получаются в результате специальной диеты и, следовательно, их может быть трудно получить. [41] [42] и/или потому, что каротиноиды необходимы для иммунной функции и, следовательно, сексуальные проявления наносят ущерб здоровью. [43]
Перья птиц изнашиваются и периодически заменяются в течение жизни птицы в результате линьки . Новые перья, известные при развитии как кровяные, или штыревые перья , в зависимости от стадии роста образуются из тех же фолликулов, из которых оперились старые. Наличие меланина в перьях повышает их устойчивость к истиранию. [44] В одном исследовании отмечается, что перья на основе меланина разлагаются быстрее под действием бактерий даже по сравнению с непигментированными перьями того же вида, чем непигментированные перья или перья с каротиноидными пигментами. [45] Однако в другом исследовании, проведенном в том же году, сравнили действие бактерий на пигментацию двух видов певчих воробьев и обнаружили, что перья с более темной пигментацией были более устойчивыми; авторы процитировали другое исследование, также опубликованное в 2004 году, в котором утверждалось, что повышенное содержание меланина обеспечивает большую устойчивость. Они заметили, что большее сопротивление более темных птиц подтверждает правило Глогера . [46]
Хотя половой отбор играет важную роль в развитии перьев, в частности, о цвете перьев, это не единственный доступный вывод. Новые исследования показывают, что уникальные перья птиц также оказывают большое влияние на многие важные аспекты поведения птиц, например, на высоту, на которой разные виды строят свои гнезда. Поскольку самки являются основными опекунами, эволюция помогла выбрать самок, которые имеют более тусклые цвета внизу, чтобы они могли сливаться с окружающей средой гнездования. Положение гнезда и вероятность того, что оно подвергнется нападению хищников, оказывают ограничения на оперение самок птиц. [47] Видам птиц, которые гнездятся на земле, а не на кронах деревьев, необходимо иметь более тусклые цвета, чтобы не привлекать внимание к гнезду. Исследование высоты показало, что птицы, гнездящиеся в кронах деревьев, часто подвергаются гораздо большему нападению хищников из-за более яркого цвета перьев, которые демонстрирует самка. [47] Еще одно влияние эволюции, которое могло сыграть роль в том, почему перья птиц такие красочные и имеют так много узоров, может быть связано с тем, что птицы развили свои яркие цвета под влиянием растительности и цветов, которые растут вокруг них. Птицы развивают свои яркие цвета, живя рядом с определенными цветами. Большинство видов птиц часто сливаются с окружающей средой из-за некоторой степени маскировки, поэтому, если среда обитания вида полна цветов и узоров, виды в конечном итоге эволюционируют, чтобы сливаться с окружающей средой, чтобы их не съели. Перья птиц имеют широкий спектр цветов, даже превосходящий разнообразие цветов многих растений, листьев и цветов. [48]
Паразиты
Поверхность перьев является домом для некоторых эктопаразитов, особенно перьевых вшей ( Phthiraptera ) и перьевых клещей. Перьевые вши обычно живут на одном хозяине и могут перемещаться только от родителей к птенцам, между спаривающимися птицами и, иногда, путем форезии . Такая история жизни привела к тому, что большинство видов паразитов специфичны для хозяина и развиваются совместно с хозяином, что делает их интересными для филогенетических исследований. [49]
Перьевые отверстия представляют собой следы жевания вшей (вероятнее всего, вшей Brueelia spp.) на перьях крыльев и хвоста. Они были описаны на ласточках , и из-за простоты подсчета многие публикации по эволюции, экологии и поведению используют их для количественной оценки интенсивности заражения.
Паразитические кукушки, которые растут в гнездах других видов, также имеют перьевых вшей, специфичных для хозяина, и они, по-видимому, передаются только после того, как молодые кукушки покидают гнездо-хозяин. [50]
Птицы поддерживают состояние перьев, прихорашиваясь и купаясь в воде или пыли . Было высказано предположение, что своеобразное поведение птиц, муравьевание , при котором муравьи внедряются в оперение, способствует уменьшению паразитов, но подтверждающих доказательств не обнаружено. [51]
Использование человеком
Утилитарный


Перья птиц издавна использовались для оперения стрел . Разноцветные перья, например, у фазанов, использовались для украшения рыболовных приманок .
Перья также помогают идентифицировать виды в судебно-медицинских исследованиях, особенно при столкновении птиц с самолетами. Соотношение изотопов водорода в перьях помогает определить географическое происхождение птиц. [52] Перья также могут быть полезны при неразрушающем отборе проб загрязняющих веществ. [53]
Птицеводство производит большое количество перьев в качестве отходов, которые, как и другие формы кератина, медленно разлагаются. Отходы перьев использовались в ряде промышленных применений в качестве среды для культивирования микробов. [54] биоразлагаемые полимеры, [55] и производство ферментов. [56] Белки перьев были опробованы в качестве клея для древесных плит. [57]
Некоторые группы коренных жителей Аляски использовали перья куропатки в качестве закалки (непластичных добавок) в гончарном производстве с первого тысячелетия до нашей эры, чтобы повысить устойчивость к термическому удару и прочность. [58]
В религии и культуре

Орлиные перья имеют большую культурную и духовную ценность для коренных американцев в Соединенных Штатах и коренных народов Канады как религиозные объекты. В Соединенных Штатах религиозное использование орлиных и ястребиных перьев регулируется Законом об орлиных перьях — федеральным законом, ограничивающим владение орлиными перьями сертифицированным и зарегистрированным членам признанных на федеральном уровне индейских племен.
В Южной Америке отвары из перьев кондоров используются в традиционных лекарствах. [59] В Индии перья индийского павлина используются в традиционной медицине при укусах змей, бесплодии и кашле. [60] [61]
Известно, что члены шотландского клана Кэмпбелл носят перья на своих шляпах, чтобы обозначить власть внутри клана. Вожди кланов носят три, вожди - два, а оруженосец - один. Любой член клана, который не соответствует критериям, не имеет права носить перья как часть традиционной одежды, и это считается самонадеянным. [62]
В XVIII, XIX и начале XX веков наблюдался бурный рост международной торговли перьями для экстравагантных женских шляп и других головных уборов (в том числе викторианской моды ). Фрэнк Чепмен отметил в 1886 году, что перья 40 видов птиц использовались примерно в трех четвертях из 700 женских шляпок, которые он наблюдал в Нью-Йорке. [63] Например, перья южноамериканских колибри в прошлом использовались для украшения некоторых миниатюрных птиц, изображенных в ящиках для певчих птиц . Эта торговля нанесла серьезный ущерб популяциям птиц (например, цапель и американских журавлей ). Защитники природы возглавили крупную кампанию против использования перьев в шляпах. Это способствовало принятию Закона Лейси в 1900 году и изменениям в моде. Рынок декоративных перьев тогда практически рухнул. [64] [65]
В последнее время петушиное оперение стало популярной тенденцией в качестве аксессуара для прически : перья, которые раньше использовались в качестве рыболовных приманок, теперь используются для придания волосам цвета и стиля. [66]
Производство изделий из перьев в Европе сократилось за последние 60 лет, главным образом из-за конкуренции со стороны Азии.Перья украшали шляпы на многих престижных мероприятиях, таких как свадьбы и женский день на ипподромах (Royal Ascot).
Эволюция


Функциональные соображения
Функциональный взгляд на эволюцию перьев традиционно фокусировался на изоляции, полете и демонстрации. Открытия нелетающих пернатых динозавров позднего мела в Китае. [67] однако предполагают, что полет не мог быть первоначальной основной функцией, поскольку перья просто не были способны обеспечивать какую-либо подъемную силу. [68] [69] Были предположения, что перья, возможно, выполняли свою первоначальную функцию терморегуляции, гидроизоляции или даже в качестве поглотителя метаболических отходов, таких как сера. [70] Утверждается, что недавние открытия подтверждают функцию терморегуляции, по крайней мере, у более мелких динозавров. [71] [72] Некоторые исследователи даже утверждают, что терморегуляция возникла из-за щетинок на лице, которые использовались в качестве тактильных датчиков. [73] Хотя предполагалось, что перья произошли от чешуи рептилий , против этой идеи существует множество возражений, а более поздние объяснения возникли на основе парадигмы эволюционной биологии развития . [2] Теории происхождения перьев, основанные на чешуе, предполагают, что плоская структура чешуи была модифицирована для развития перьев путем расщепления с образованием перепонки; однако этот процесс развития включает трубчатую структуру, возникающую из фолликула, и трубку, разделяющуюся в продольном направлении, образующую перепонку. [1] [2] Количество перьев на единицу площади кожи у мелких птиц выше, чем у крупных, и эта тенденция указывает на их важную роль в теплоизоляции, поскольку более мелкие птицы теряют больше тепла из-за относительно большей площади поверхности по отношению к массе тела. . [5] Миниатюризация птиц также сыграла роль в развитии механизированного полета. [74] Считается, что окраска перьев развилась главным образом в результате полового отбора . У ископаемых экземпляров паравиана Anchiornis huxleyi и птерозавра Tupandactylus imperator особенности сохранились настолько хорошо, что меланосом можно наблюдать структуру цвет и рисунок перьев Anchiornis и Tupandactylus . (пигментных клеток). Сравнивая форму ископаемых меланосом с меланосомами современных птиц, можно было определить [75] [76] Было обнаружено, что у Anchiornis на передних и задних конечностях перья с черно-белым рисунком и красновато-коричневым гребнем. Этот рисунок похож на окраску многих современных видов птиц, которые используют окраску оперения для демонстрации и общения, включая половой отбор и маскировку. Вполне вероятно, что нептичьи виды динозавров использовали модели оперения для выполнения тех же функций, что и современные птицы, до того, как они начали летать. Во многих случаях о физиологическом состоянии птиц (особенно самцов) свидетельствует качество их перьев, и это используется (самками) при выборе партнера . [77] [78] Кроме того, при сравнении разных экземпляров Ornithomimus edmontonicus у пожилых особей было обнаружено пеннибрахий (крылообразная структура, состоящая из удлиненных перьев), а у более молодых - нет. Это говорит о том, что пеннибрахий был вторичным половым признаком и, вероятно, выполнял сексуальную функцию. [79]
Молекулярная эволюция
несколько генов Было обнаружено , определяющих развитие перьев. Они будут ключом к пониманию эволюции перьев. Например, некоторые гены преобразуют чешуйки в перья или пероподобные структуры, когда они экспрессируются или индуцируются в лапах птиц, например конвертеры чешуйчатых перьев Sox2 , Zic1 , Grem1 , Spry2 и Sox18 . [80]
Перья и чешуя состоят из двух различных форм кератина , и долгое время считалось, что каждый тип кератина свойственен только каждой структуре кожи (перьям и чешуйкам). Однако кератин перьев также присутствует на ранних стадиях развития чешуи американского аллигатора . Этот тип кератина, который ранее считался специфичным для перьев, подавляется во время эмбрионального развития аллигатора и поэтому не присутствует в чешуе взрослых аллигаторов. Присутствие этого гомологичного кератина как у птиц, так и у крокодилов указывает на то, что он был унаследован от общего предка. [81]
Это может свидетельствовать о том, что чешуя крокодила, перья птиц и динозавров, а также пикноволокна птерозавров являются проявлениями развития одних и тех же примитивных структур кожи архозавров; предполагая, что перья и пикноволокна могут быть гомологичными. [82] Методы молекулярного датирования, проведенные в 2011 году, показывают, что подсемейство β-кератинов перьев, обнаруженное у современных птиц, начало расходиться 143 миллиона лет назад, что позволяет предположить, что перистые перья Anchiornis не состояли из β-кератинов перьев, присутствующих у современных птиц. [83] Однако исследование ископаемых перьев динозавра Sinosauropteryx и других окаменелостей с использованием инфракрасной спектроскопии и серно-рентгеновской спектроскопии выявило следы белков бета-листа. Было показано, что присутствие обильных альфа-белков в некоторых ископаемых перьях является артефактом процесса окаменения, поскольку структуры бета-белков легко превращаются в альфа-спирали во время термической деградации. [84] В 2019 году ученые обнаружили, что гены производства перьев развились в основании архозаврий, что подтверждает, что перья присутствовали у ранних орнитодиран и согласуются с летописью окаменелостей. [85]
Пернатые динозавры

У некоторых нептичьих динозавров на конечностях были перья, которые не позволяли летать. [67] [2] Одна теория предполагает, что перья изначально появились у динозавров из-за их изоляционных свойств; затем мелкие виды динозавров, у которых были более длинные перья, возможно, нашли их полезными для планирования, что привело к эволюции протоптиц, таких как археоптерикс и микрораптор zhaoianus . Другая теория утверждает, что первоначальным адаптивным преимуществом ранних перьев была их пигментация или переливчатость, способствующая сексуальным предпочтениям при выборе партнера. [86] Динозавры, у которых были перья или протоперья, включают Pedopenna daohugouensis. [87] и Dilong paradoxus , тираннозавроид , который на 60-70 миллионов лет старше тираннозавра рекса . [88]
Большинство динозавров, которые, как известно, имели перья или протоперья, являются тероподами , однако перьеподобные «нитевидные покровные структуры» также известны у птицетазовых динозавров Tianyulong и Psittacosaurus . [89] Точная природа этих структур все еще изучается. Однако считается, что перья стадии 1 (см. раздел «Этапы эволюции» ниже), подобные тем, которые наблюдаются у этих двух птицетазовых, вероятно, функционировали демонстративно. [90] птицетазовый кулиндадромеус имеет структуры, напоминающие перья третьей стадии. В 2014 году сообщалось, что [91] Вероятность развития чешуи у ранних предков динозавров высока. Однако это было сделано на основе предположения, что примитивные птерозавры были чешуйчатыми. [92] [93] Исследование 2016 года проанализировало морфологию пульпы хвостовых щетинок Psittacosaurus и обнаружило, что они похожи на перья, но отмечает, что они также похожи на щетинки на голове конголезского павлина , бороду индейки и позвоночник на голове. рогатого крикуна . [94] Переоценка максимальной вероятности, проведенная палеонтологом Томасом Хольцем, показывает, что нити, скорее всего, были предковым состоянием динозавров. [95]
В 2010 году у кархародонтозаврид по имени Concavenator corcovatus были обнаружены ремиги на локтевой кости, что позволяет предположить, что у него могли быть игольчатые структуры на амортизирующих мышцах. [96] Однако Фот и др. 2014 г. не согласны с публикацией, в которой указывают, что шишки на локтевой кости конкавенатора находятся на передне-латеральной стороне , что в отличие от ремижей, находящихся на задне-латеральной части локтевой кости некоторых птиц, они считают более вероятным, что это крепления межкостных связок. [97] Это опровергли Куэста Фидальго и ее коллеги, они указали, что эти бугорки на локтевой кости являются заднелатеральными, что не похоже на таковые у межкостных связок. [98]
С 1990-х годов десятки пернатых динозавров были обнаружены в кладе Maniraptora , которая включает кладу Avialae и недавних общих предков птиц — Oviraptorosauria и Deinonychosauria . В 1998 году открытие пернатого овирапторозавра Caudipteryx zoui бросило вызов представлению о перьях как структуре, исключительной для Avialae. [99] Похороненный в формации Исянь в провинции Ляонин, Китай, C. zoui жил в раннемеловом периоде. Присутствует на передних конечностях и хвосте, их покровная структура принята. [ кем? ] в виде перистых лопастных перьев, основанных на ости и елочном узоре бородок. В кладе Deinonychosauria продолжающееся расхождение перьев наблюдается также у семейств Troodontidae и Dromaeosauridae . Разветвленные перья с гребнями, зазубринами и бородками были обнаружены у многих представителей, включая Sinornithosaurus millenii , дромеозаврида, обнаруженного в формации Исянь (124,6 млн лет назад). [100]
Раньше в эволюции перьев существовал временной парадокс — тероподы с высокоразвитыми птицеподобными характеристиками возникли позже, чем археоптерикс, — что позволяет предположить, что потомки птиц возникли раньше предка. Однако открытие Anchiornis huxleyi в позднеюрской формации Тяоцзишань (160 млн лет назад) на западе Ляонина в 2009 г. [101] [102] разрешил этот парадокс. Предшествуя Археоптериксу , Анхиорнис доказывает существование современного пернатого предка теропода, что дает представление о переходе от динозавра к птице. На экземпляре видно распределение крупных перистых перьев на передних конечностях и хвосте, что означает, что перистые перья распространились на остальную часть тела на более ранней стадии эволюции теропод. [103] Развитие перистых перьев не заменило более ранние нитевидные перья. Нитчатые перья сохранились наряду с современными маховыми перьями, в том числе с модификациями, обнаруженными в перьях современных ныряющих птиц, в янтаре возрастом 80 миллионов лет из Альберты. [104]
Два маленьких крыла, заключённые в янтаре, датируемые 100 млн лет назад, свидетельствуют о том, что оперение существовало у некоторых предшественников птиц. Крылья, скорее всего, принадлежали энантиорнитам , разнообразной группе птичьих динозавров. [105] [106]
Большой филогенетический анализ ранних динозавров, проведенный Мэтью Бароном, Дэвидом Б. Норманом и Полом Барреттом (2017), показал, что Theropoda на самом деле более тесно связана с Ornithischia , с которой она образовала сестринскую группу в кладе Ornithoscelida . Исследование также показало, что если перьевые структуры теропод и птицетазовых имеют общее эволюционное происхождение, то вполне возможно, что перья были ограничены Ornithoscelida. Если это так, то происхождение перьев, вероятно, произошло еще в среднем триасе . [107] хотя с этим не согласились. [108] [109] Отсутствие перьев у крупных зауроподов и анкилозавров могло быть связано с тем, что перья были подавлены геномными регуляторами. [110]
Эволюционные этапы

Несколько исследований развития перьев у эмбрионов современных птиц в сочетании с распределением типов перьев среди различных предшественников доисторических птиц позволили ученым попытаться восстановить последовательность, в которой перья впервые эволюционировали и превратились в типы, обнаруженные у современных птиц.
В 2009 году Сюй и Го разбили эволюцию перьев на следующие этапы: [90]
- Одиночная нить
- Несколько нитей соединены у основания.
- Несколько нитей, соединенных у основания с центральной нитью.
- Множественные нити по длине центральной нити.
- Множественные нити, отходящие от края мембранозной структуры.
- Перистое перо с лопастями бородок и бородок и центральным стержнем.
- Перистое перо с асимметричным стержнем.
- Недифференцированная лопасть с центральным рахисом
Однако Фот (2011) показал, что некоторые из этих предполагаемых стадий (в частности, стадии 2 и 5), вероятно, являются просто артефактами консервации, вызванными тем, как ископаемые перья измельчаются и сохраняются остатки или отпечатки перьев. Фот переинтерпретировал перья стадии 2 как раздавленные или ошибочно идентифицированные перья как минимум стадии 3, а перья стадии 5 - как раздавленные перья стадии 6. [111]
Следующая упрощенная диаграмма взаимоотношений динозавров следует за этими результатами и показывает вероятное распределение перистых (пуховых) и перистых (лопастных) перьев среди динозавров и доисторических птиц. Схема соответствует схеме, представленной Сюй и Го (2009). [90] модифицировано с учетом результатов Фота (2011) [111] Цифры, сопровождающие каждое имя, указывают на наличие определенных стадий перьев. Обратите внимание, что буква «s» указывает на известное наличие чешуи на теле.
У птерозавров
Давно известно, что птерозавры имеют нитевидные, похожие на мех структуры, покрывающие их тело, известные как пикноволокна , которые обычно считались отличными от «настоящих перьев» птиц и их родственников динозавров. Однако исследование, проведенное в 2018 году двух небольших, хорошо сохранившихся окаменелостей птерозавров из юрского периода Внутренней Монголии ( , Китай) показало, что птерозавры были покрыты множеством пикноволокон разной структуры (а не просто нитевидными), причем некоторые из этих структур демонстрируют диагностические признаки. особенности перьев, такие как сгруппированные нити без жилок и двусторонне разветвленные нити, которые первоначально считались эксклюзивными для птиц и других динозавров-манирапторов. Учитывая эти данные, вполне возможно, что перья имеют глубокое эволюционное происхождение у предков архозавров , хотя существует также вероятность того, что эти структуры независимо эволюционировали и стали напоминать птичьи перья в результате конвергентной эволюции . [112] Майк Бентон, старший автор исследования, подтвердил прежнюю теорию, заявив: «Мы не смогли найти никаких анатомических доказательств того, что четыре типа пикноволокон каким-либо образом отличаются от перьев птиц и динозавров. Следовательно, поскольку они одинаковы , они должны иметь общее эволюционное происхождение, и это было около 250 миллионов лет назад, задолго до возникновения птиц». [113] [114] [115] [116] покровные структуры экземпляров анурогнатид Но , как отметила Лилиана Д'Альба, по-прежнему основаны на грубой морфологии. Пикноволокна двух экземпляров анурогнатид, возможно, не гомологичны нитевидным придаткам динозавров. [117] Пол М. Барретт подозревает, что в ходе покровной эволюции птерозавров птерозавры примитивно потеряли чешую и начали появляться пикноволокна. [93]
Каскокауда был почти полностью покрыт обширным слоем пикноволокон, которые, по-видимому, были двух типов. Первые представляют собой простые изогнутые нити длиной от 3,5 до 12,8 мм. Эти нити покрывают большую часть животного, включая голову, шею, тело, конечности и хвост. Второй тип состоит из пучков нитей, соединенных у основания, похожих на ветвящиеся вниз перья птиц и других целурозавровых динозавров, длиной около 2,5–8,0 мм и покрывают только перепонки крыльев. Исследования образцов пикноволокон выявили наличие внутри нитей микротельц, напоминающих пигменты меланосом, идентифицированные в других ископаемых покровах, в частности в феомеланосомах. Кроме того, инфракрасный спектральный анализ этих пикноволокон показывает спектры поглощения, аналогичные рыжим человеческим волосам . Эти пикноволокна, вероятно, обеспечивали и изоляцию, и, возможно, помогали обтекать тело и крылья во время полета. [118] Идентичность этих ветвящихся структур как пикноволокон или перьев была оспорена Анвином и Мартиллом (2020), которые интерпретировали их как сгруппированные и деградированные актинофибриллы – волокна, придающие жесткость, обнаруженные в мембране крыльев птерозавров – и приписали меланосомы и кератин коже, а не коже. чем нити. [119] Эти утверждения были опровергнуты Янгом и его коллегами, которые утверждают, что интерпретации Анвина и Мартилла несовместимы с сохранностью образца. А именно, они утверждают, что последовательная структура, регулярное расположение и расширение нитей за пределы мембраны крыла подтверждают их идентификацию как пикноволокон. Кроме того, они утверждают, что ограничение количества меланосом и кератина в волокнах, как это происходит в перьях ископаемых динозавров, подтверждает тот факт, что они являются нитями, и не согласуется с загрязнением сохранившейся кожи. [120] Протоперья, вероятно, развились у ранних архозавров вскоре после вымирания PT, когда скорость метаболизма ранних архозавров и синапсид увеличивалась, позы становились выпрямленными и сохраняли устойчивую активность. [85]
См. также
Ссылки
- ^ Jump up to: а б с д Прум, Ричард О .; А. Х. Кисть (2002). «Эволюционное происхождение и разнообразие перьев» (PDF) . Ежеквартальный обзор биологии . 77 (3): 261–295. дои : 10.1086/341993 . ПМИД 12365352 . S2CID 6344830 . Архивировано (PDF) из оригинала 29 июня 2011 года . Проверено 7 июля 2010 г.
- ^ Jump up to: а б с д Прум, Р.О. и Браш, АХ (март 2003 г.). «Что было первым: перо или птица?» (PDF) . Научный американец . 288 (3): 84–93. Бибкод : 2003SciAm.288c..84P . doi : 10.1038/scientificamerican0303-84 (неактивен 30 июня 2024 г.). ПМИД 12616863 . Архивировано (PDF) из оригинала 29 июня 2011 года . Проверено 7 июля 2010 г.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июнь 2024 г. ( ссылка ) - ^ Прум, Ричард О (1999). «Развитие и эволюционное происхождение перьев» (PDF) . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 285 (4): 291–306. Бибкод : 1999JEZ...285..291P . doi : 10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9 . ПМИД 10578107 . Архивировано из оригинала (PDF) 9 апреля 2011 года . Проверено 7 июля 2010 г.
- ^ Ли, Цюаньго (9 марта 2012 г.). «Реконструкция микрораптора и эволюция радужного оперения». Наука . 335 (6073): 1215–1219. Бибкод : 2012Sci...335.1215L . дои : 10.1126/science.1213780 . ПМИД 22403389 . S2CID 206537426 .
- ^ Jump up to: а б с Петтингилл, О.С. младший (1970). Орнитология в лаборатории и полевых условиях. Четвертое издание . Издательская компания Берджесс. стр. 29–58 . ISBN 0-8087-1609-3 .
- ^ «Плюмология Галапагосских островов» (PDF) . darwinfoundation.org . База данных коллекций Чарльза Дарвина Фонда Чарльза Дарвина. Архивировано из оригинала 17 марта 2016 года . Проверено 24 апреля 2015 г.
- ^ Эйххорн, изд. Манфред (2005). Специальный словарь биологии Лангеншайдта на английском языке: английский – немецкий, немецкий – английский (1-е изд.). Берлин [среди прочих]: Лангеншайдт. п. 537. ИСБН 3-86117-228-3 . Архивировано из оригинала 17 декабря 2017 года . Проверено 24 апреля 2015 г.
- ^ Бонсер, RHC; Доусон, К. (1999). «Структурно-механические свойства пуховых перьев и биоимитирующих натуральные изоляционные материалы». Журнал материаловедческих писем . 18 (21): 1769–1770. дои : 10.1023/A:1006631328233 . S2CID 135061282 .
- ^ Джонстон, Николь и Парсонс, Джин (20 сентября 2018 г.). «Перья: под угрозой исчезновения – фауна и мода» . Университета Миссури . Коллекция исторических костюмов и текстиля
- ^ "ручка(3)" . Интернет-словарь Мерриам-Вебстера . Merriam-Webster, Inc. Архивировано из оригинала 19 сентября 2011 года . Проверено 16 октября 2010 г.
- ^ Шор, Р.; Кримм, С. (1961). «Исследования структуры кератина перьев: II. Модель β-спирали для структуры кератина перьев» . Биофиз. Дж . 1 (6): 489–515. Бибкод : 1961BpJ.....1..489S . дои : 10.1016/S0006-3495(61)86904-X . ПМЦ 1366335 . ПМИД 19431311 .
- ^ Полинг, Лайнус; Кори, Роберт Б. (1951). «Структура кератина перьевого гребня» . Труды Национальной академии наук Соединенных Штатов Америки . 37 (5): 256–261. Бибкод : 1951ПНАС...37..256П . дои : 10.1073/pnas.37.5.256 . ПМЦ 1063351 . ПМИД 14834148 .
- ^ Хорник, К.; Кришан, К.; Юсуф, Ф.; Скааль, М.; Бранд-Сабери, Б. (2005). «Неправильная экспрессия cDermo-1 приводит к образованию плотной дермы, перьев и чешуек» . Биология развития . 277 (1): 42–50. дои : 10.1016/j.ydbio.2004.08.050 . ПМИД 15572138 .
- ^ Ницш, Кристиан Людвиг (1867). Птерилография Нича . Рэйское общество. п. 14 .
- ^ Чендлер 1916 , с. 261
- ^ Чендлер, Аса К. (1916). «Изучение строения перьев с учетом их таксономического значения» . Публикации Калифорнийского университета по зоологии . 13 (11): 243–446 [284].
- ^ Маклелланд, Дж. (1991). Цветной атлас анатомии птиц . WB Saunders Co. ISBN 0-7216-3536-9 .
- ^ Кэмерон, Г.; Весс, Т.; Бонсер, Р. (2003). «Модуль Юнга варьируется в зависимости от дифференциальной ориентации кератина в перьях». Журнал структурной биологии . 143 (2): 118–23. дои : 10.1016/S1047-8477(03)00142-4 . ПМИД 12972348 .
- ^ Бонсер, Р.; Сакер, Л.; Джеронимидис, Г. (2004). «Анизотропия прочности кератина перьев». Журнал материаловедения . 39 (8): 2895–2896. Бибкод : 2004JMatS..39.2895B . doi : 10.1023/B:JMSC.0000021474.75864.ff . S2CID 135873731 .
- ^ Jump up to: а б Ван, Бин (2016). «Легкий, как перышко: волокнистый натуральный композит, форма которого меняется от круглой до квадратной» . Передовая наука . 4 (3): 1600360. doi : 10.1002/advs.201600360 . ПМК 5357985 . ПМИД 28331789 .
- ^ Лингхэм-Солиар, Теагартен (2013). «Новая спиральная структура b-кератина с перекрещенными волокнами в маховых перьях и ее биомеханические последствия» . ПЛОС ОДИН . 8 (6): e65849. Бибкод : 2013PLoSO...865849L . дои : 10.1371/journal.pone.0065849 . ПМЦ 3677936 . ПМИД 23762440 .
- ^ Итон, Мьюир Д.; Ланьон, Скотт М. (2003). «Повсеместное отражение ультрафиолетового оперения птиц» . Труды: Биологические науки . 270 (1525): 1721–1726. дои : 10.1098/rspb.2003.2431 . ПМЦ 1691429 . ПМИД 12965000 .
- ^ Боствик, Кимберли С.; Ричард О., Прум (2005). «Ухаживающие птицы поют стригущимися перьями крыльев» (PDF) . Наука . 309 (5735): 736. doi : 10.1126/science.1111701 . ПМИД 16051789 . S2CID 22278735 . Архивировано (PDF) из оригинала 7 июля 2010 года . Проверено 19 июля 2010 г.
- ^ Jump up to: а б Дели, К; Питерс, А.; Кемпенаерс, Б. (2007). «Косметическая окраска птиц: возникновение, функции и эволюция» (PDF) . Являюсь. Нат . 169 : С145–158. дои : 10.1086/510095 . ПМИД 19426089 . S2CID 29592388 . Архивировано из оригинала (PDF) 3 декабря 2007 года.
- ^ Рыбак, Г.; Вейс, Д.; Арад, З. (2005). «Задержка воды в оперении ныряющих больших бакланов Phalacrocorax carbo sinensis ». Дж. Авиан Биол . 36 (2): 89–95. дои : 10.1111/j.0908-8857.2005.03499.x .
- ^ Ледерер, Роджер Дж. (1972). «Роль птичьих щетинок» (PDF) . Бюллетень Уилсона . 84 : 193–97. Архивировано (PDF) из оригинала 4 февраля 2014 г.
- ^ Коновер, MR; Миллер, Делавэр (1980). «Риктальная щетинка у ивовой мухоловки» . Кондор . 82 (4): 469–471. дои : 10.2307/1367580 . JSTOR 1367580 . Архивировано из оригинала 22 февраля 2014 года.
- ^ Пирсма, Т; ван Эрден, MR (1989). «Поедание перьев у больших хохлатых поганок Podiceps cristatus : уникальное решение проблем мусора и желудочных паразитов у рыбоядных птиц» (PDF) . Ибис . 131 (4): 477–486. дои : 10.1111/j.1474-919X.1989.tb04784.x .
- ^ Демэй, Ида С. (1940). «Исследование птерилеза и пневматичности крикуна» . Кондор . 42 (2): 112–118. дои : 10.2307/1364475 . JSTOR 1364475 . Архивировано из оригинала 21 февраля 2014 года.
- ^ Холл, К.; Сюзанна С. (2005). «У воробьинообразных с девятью первичными перьями девять или десять первичных перьев? Эволюция концепции». Журнал орнитологии . 146 (2): 121–126. дои : 10.1007/s10336-004-0070-5 . S2CID 36055848 .
- ^ Пикрафт, WP (1895). «К птерилографии гоацина ( Opisthocomus cristatus )» . Ибис . 37 (3): 345–373. дои : 10.1111/j.1474-919X.1895.tb06744.x .
- ^ Тернер, Дж. Скотт (1997). «О теплоемкости птичьего яйца, согретого расплодом» (PDF) . Физиологическая зоология . 70 (4): 470–80. дои : 10.1086/515854 . ПМИД 9237308 . S2CID 26584982 . Архивировано из оригинала (PDF) 20 октября 2022 года . Проверено 29 июля 2020 г. - через EBSCO.
- ^ МакГроу, К.Х.; Ногаре, MC (2005). «Распространение уникальных красных пигментов перьев у попугаев» . Письма по биологии . 1 (1): 38–43. дои : 10.1098/rsbl.2004.0269 . ПМК 1629064 . ПМИД 17148123 .
- ^ Хаусманн, Ф.; Арнольд, Кентукки; Маршалл, Нью-Джерси; Оуэнс, IPF (2003). «Ультрафиолетовые сигналы у птиц особенные» . Труды Королевского общества Б. 270 (1510): 61–67. дои : 10.1098/rspb.2002.2200 . ПМК 1691211 . ПМИД 12590772 .
- ^ Шоки, Мэтью Д.; Хилл, Джеффри Э (2005). «Каротиноидам нужны структурные цвета, чтобы сиять» (PDF) . Биол. Летт . 1 (2): 121–124. дои : 10.1098/rsbl.2004.0289 . ПМЦ 1626226 . ПМИД 17148144 . Архивировано из оригинала (PDF) 26 марта 2009 года.
- ^ Винтер, Якоб; Бриггс, Дерек Э.Г.; Кларк, Джулия; Майр, Джеральд; Прум, Ричард О. (2009). «Структурная окраска ископаемого пера» (PDF) . Письма по биологии . 6 (1): 128–31. дои : 10.1098/rsbl.2009.0524 . ПМЦ 2817243 . ПМИД 19710052 . Архивировано из оригинала (PDF) 21 июня 2010 года . Проверено 19 июля 2010 г.
- ^ Дайк, Дж. (1971). «Структура и спектральная отражательная способность зеленых и синих перьев неразлучника ( Agapornis roseicollis )». Биол. СКР . 18 : 1–67.
- ^ Шоки, доктор медицины; Хилл, GE (2005). «Перья в мелком масштабе» . Аук . 121 (3): 652–655. doi : 10.1642/0004-8038(2004)121[0652:FAAFS]2.0.CO;2 .
- ^ Дели, К.; Питерс, А.; Бидерманн, PHW; Кемпенаерс, Б. (2008). «Оптические свойства секрета уропигиальной железы: нет доказательств использования УФ-косметики у птиц» . Naturwissenschaften . 95 (10): 939–46. Бибкод : 2008NW.....95..939D . дои : 10.1007/s00114-008-0406-8 . hdl : 11858/00-001M-0000-0010-509C-A . ПМИД 18560743 .
- ^ Шоки, доктор медицины; Пиллаи, СР; Хилл, GE (2003). «Химическая война? Влияние уропигиального масла на бактерии, разлагающие перья» (PDF) . Журнал птичьей биологии . 34 (4): 345–349. дои : 10.1111/j.0908-8857.2003.03193.x . Архивировано из оригинала (PDF) 10 сентября 2008 года.
- ^ Эндлер, Дж. А. (1980). «Естественный отбор по цветовым узорам Poeci-lia reticulata». Эволюция . 34 (1): 76–91. дои : 10.2307/2408316 . JSTOR 2408316 . ПМИД 28563214 .
- ^ Бадяев А.В.; Хилл, GE (2000). «Эволюция полового дихроматизма: вклад каротиноидов в окраску по сравнению с окраской на основе меланина» . Биологический журнал Линнеевского общества . 69 (2): 153–172. дои : 10.1111/j.1095-8312.2000.tb01196.x . S2CID 201965078 .
- ^ Лозано, Джорджия (1994). «Каротиноиды, паразиты и половой отбор». Ойкос . 70 (2): 309–311. Бибкод : 1994Oikos..70..309L . дои : 10.2307/3545643 . JSTOR 3545643 . S2CID 86971117 .
- ^ Бонсер, RHC (1995). «Меланин и устойчивость перьев к истиранию» . Кондор . 97 (2): 590–591. дои : 10.2307/1369048 . JSTOR 1369048 . Архивировано из оригинала 23 февраля 2014 года.
- ^ Гранде, Дж. М.; Негр, Джей-Джей; Торрес, MJ (2004). «Эволюция окраски оперения птиц: роль бактерий, разлагающих перья?» (PDF) . Ардеола . 51 (2): 375–383. Архивировано (PDF) из оригинала 10 сентября 2008 г.
- ^ Бертт, Эдвард Х. младший; Ичида, Янн М. (2004). «Правило Глогера, бактерии, разлагающие перья, и изменение окраски певчих воробьев» (PDF) . Кондор . 106 (3): 681–686. дои : 10.1650/7383 . S2CID 5857742 . Архивировано (PDF) из оригинала 20 ноября 2012 года.
- ^ Jump up to: а б Мартин, TE; Бадяев, А.В. (1996). «Половая дихроматика у птиц; важность хищничества и расположения гнезд для самок по сравнению с самцами». Эволюция . 50 (6): 2454–2460. дои : 10.2307/2410712 . JSTOR 2410712 . ПМИД 28565684 .
- ^ Касвелл Стоддард, Мэри; Прум, Ричард О. (2011). «Насколько красочны птицы? Эволюция цветовой гаммы оперения птиц» . Поведенческая экология . 22 (5): 1042–1052. дои : 10.1093/beheco/arr088 . hdl : 10.1093/beheco/arr088 .
- ^ Тун А. и Хьюз Дж. (2008). «Являются ли вши хорошими индикаторами истории хозяина? Сравнительный анализ австралийской сороки Gymnorhina tibicen и двух видов перьевых вошей» . Наследственность . 101 (2): 127–135. дои : 10.1038/hdy.2008.37 . ПМИД 18461081 .
- ^ Брук, М. де Л.; Хироши Накамура (1998). «Приобретение перьевых вшей, специфичных для хозяина, обыкновенными кукушками ( Cuculus canorus )». Журнал зоологии . 244 (2): 167–173. дои : 10.1017/S0952836998002027 .
- ^ Ревис, Ханна К.; Дебора А. Уоллер (2004). «Бактерицидное и фунгицидное действие муравьиных химикатов на перьевых паразитов: оценка муравьиного поведения как метода самолечения у певчих птиц» . Аук . 121 (4): 1262–1268. doi : 10.1642/0004-8038(2004)121[1262:BAFAOA]2.0.CO;2 . S2CID 85677766 .
- ^ Боуэн, Габриэль Дж; Вассенаар, Леонард I; Хобсон, Кейт А. (2005). «Глобальное применение стабильных изотопов водорода и кислорода в криминалистике дикой природы». Экология . 143 (3): 337–348. Бибкод : 2005Oecol.143..337B . дои : 10.1007/s00442-004-1813-y . ПМИД 15726429 . S2CID 1762342 .
- ^ Ясперс, В.; Вурспоелс, С.; Ковачи, А.; Лепойнт, Г. и Иенс, М. (2007). «Оценка полезности птичьих перьев как инструмента неразрушающего биомониторинга органических загрязнителей: сравнительный и метааналитический подход» . Интернационал окружающей среды . 33 (3): 328–337. Бибкод : 2007EnInt..33..328J . дои : 10.1016/j.envint.2006.11.011 . hdl : 2268/1067 . ПМИД 17198730 .
- ^ Пупати, С.; Абидха, С. (2007). «Использование питательных сред на основе перьев для производства комарицидных бактерий». Биологический контроль . 43 (1): 49–55. Бибкод : 2007БиолС..43...49П . doi : 10.1016/j.biocontrol.2007.04.019 .
- ^ Шмидт, ВФ; Бароне, младший (2004). «Новое применение кератинового волокна куриных перьев». Материалы симпозиума по управлению отходами птицеводства . стр. 99–101.
- ^ Казарин, Франчиани; Бранделли, Флоренсия Кладера-Оливера Адриано; Бранделли, Адриано (2008). «Использование побочных продуктов птицеводства для производства кератинолитических ферментов». Пищевые и биотехнологические технологии . 1 (3): 301–305. дои : 10.1007/s11947-008-0091-9 . S2CID 96154497 .
- ^ Цзян, З.; Цинь, Д.; Хсе, К.; Куо, М.; Луо, З.; Ван, Г.; и др. (2008). «Предварительное исследование клеев для древесины на основе белка куриного пера» . Журнал химии и технологии древесины . 28 (3): 240–246. дои : 10.1080/02773810802347073 . S2CID 3656808 . Архивировано из оригинала 19 февраля 2014 года.
- ^ Неузиус, Сара В. и Г. Тимоти Гросс2007 В поисках нашего прошлого: введение в археологию Северной Америки. Издательство Оксфордского университета, Нью-Йорк.
- ^ Фромминг, Стив (2006). «Традиционное использование андского мерцания (Colaptes rupicola) в качестве галактагога в Перуанских Андах» . Журнал этнобиологии и этномедицины . 2:23 . дои : 10.1186/1746-4269-2-23 . ПМЦ 1484469 . ПМИД 16677398 .
- ^ Мурари, СК; Фрей, Ф.Дж.; Фрей, Б.М.; Гауда, ТВ; Вишванат, Б.С. (2005). «Использование экстракта перьев Pavo cristatus для лучшего лечения змеиных укусов: нейтрализация воспалительных реакций». Журнал этнофармакологии . 99 (2): 229–237. дои : 10.1016/j.jep.2005.02.027 . ПМИД 15894132 .
- ^ Махавар, ММ; Джароли, ДП (2007). «Традиционные знания о зоотерапевтическом использовании племени Сахария в Раджастане, Индия» . Журнал этнобиологии и этномедицины . 3:25 . дои : 10.1186/1746-4269-3-25 . ЧВК 1892771 . ПМИД 17547781 .
- ^ «Платье шотландских горцев: клан Кэмпбелл» . Общество клана Кэмпбелл . Проверено 10 ноября 2020 г.
- ^ Даути, Робин В. Мода на перья и охрана птиц, Исследование по охране природы . Издательство Калифорнийского университета. Страница 197.
- ^ Эрлих, Пол Р.; Добкин, Дэвид С.; Сыворотка, Дэррил (1988). «Плюм Трейд» . Стэнфордский университет. Архивировано из оригинала 30 сентября 2008 года.
- ↑ Торговля перьями . Архивировано 23 июня 2008 года в Wayback Machine , Смитсоновский институт.
- ^ Боннер, Джесси Л. (6 июня 2011 г.). «Высокая мода или приманка? Завязки теперь наращивают волосы» . Сиэтл Таймс . Архивировано из оригинала 10 июня 2011 года.
- ^ Jump up to: а б Сен-Флер, Николас (8 декабря 2016 г.). «Эта штука с перьями, застрявшими в янтаре? Это был хвост динозавра» . Нью-Йорк Таймс . Архивировано из оригинала 8 декабря 2016 года . Проверено 8 декабря 2016 г.
- ^ Сумида, СС; CA Брошу (2000). «Филогенетический контекст происхождения перьев» . Американский зоолог . 40 (4): 486–503. дои : 10.1093/icb/40.4.486 .
- ^ Даймонд, CC; Каюта Р.Дж.; Дж. С. Брукс (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального отображения адаптивной функции перьев у нептичьих теропод». БИОС . 82 (3): 58–63. дои : 10.1893/011.082.0302 . S2CID 98221211 .
- ^ Бок, WJ (2000). «Толковая история происхождения перьев» . Являюсь. Зоол . 40 (4): 478–485. дои : 10.1093/icb/40.4.478 .
- ^ Уитфилд, Джон (4 апреля 2012 г.). «Самый крупный пернатый динозавр, обнаруженный в Китае» . Блог новостей природы . Архивировано из оригинала 6 апреля 2012 года . Проверено 4 апреля 2012 г.
- ^ Сюй С.; Ван К.; Чжан К.; Ма К.; Син Л.; Салливан К.; Ху Д.; Ченг С.; Ван С.; и др. (2012). «Гигантский пернатый динозавр из нижнего мела Китая» (PDF) . Природа . 484 (7392): 92–95. Бибкод : 2012Natur.484...92X . дои : 10.1038/nature10906 . ПМИД 22481363 . S2CID 29689629 . Архивировано из оригинала (PDF) 17 апреля 2012 года.
- ^ Персоны, Уолтер С.; Карри, Филип Дж. (2015). «Щетинка перед пухом: новый взгляд на функциональное происхождение перьев» . Эволюция . 69 (4): 857–862. дои : 10.1111/evo.12634 . ISSN 1558-5646 . ПМИД 25756292 . S2CID 24319963 .
- ^ Де Риклс; Эй Джей; К. Падиан; Дж. Р. Хорнер; Э.Т. Ламм; Н. Мирволд (2003). «Остеогистология confuciusornis Sanctus (theropoda: Aves)». Журнал палеонтологии позвоночных . 23 (2): 373–386. doi : 10.1671/0272-4634(2003)023[0373:oocsta]2.0.co;2 . S2CID 84936431 .
- ^ Ли, Цюаньго; Гао, Кэ-Цинь; Винтер, Якоб; Шоки, Мэтью; Кларк, Джулия; Д'Альба, Лилиана; Мэн, Цинцзинь; Бриггс, Дерек; Прум, Ричард (12 марта 2010 г.). «Цвет оперения вымершего динозавра» (PDF) . Наука . 327 (5971): 1369–1372. Бибкод : 2010Sci...327.1369L . дои : 10.1126/science.1186290 . ПМИД 20133521 . S2CID 206525132 .
- ^ Чинкотта А., Николаи М., Кампос, HBN и др. Меланосомы птерозавров поддерживают сигнальные функции ранних перьев. Природа 604, 684–688 (2022). два : 10.1038/s41586-022-04622-3 .
- ^ Саино, Никола; Риккардо Стради (1999). «Концентрация каротиноидов в плазме, иммунный профиль и украшение оперения самцов деревенских ласточек». Американский натуралист . 154 (4): 441–448. дои : 10.1086/303246 . ПМИД 10523490 . S2CID 4400888 .
- ^ Эндлер, Джон А.; Дэвид А. Уэсткотт; Джоа Р. Мэдден; Тим Робсон и Патрик Филлипс (2005). «Зрительные системы животных и эволюция цветовых паттернов: сенсорная обработка проливает свет на эволюцию сигналов» . Эволюция . 59 (8): 1795–1818. дои : 10.1111/j.0014-3820.2005.tb01827.x . ПМИД 16329248 . S2CID 25683790 .
- ^ Зеленицкий, ДК; Терриен, Ф.; Эриксон, генеральный директор; ДеБур, CL; Кобаяши, Ю.; Эберт, Д.А.; Хэдфилд, Ф. (26 октября 2012 г.). «Пернатые нептичьи динозавры из Северной Америки дают представление о происхождении крыльев». Наука . 338 (6106): 510–514. Бибкод : 2012Sci...338..510Z . дои : 10.1126/science.1225376 . ISSN 0036-8075 . ПМИД 23112330 . S2CID 2057698 .
- ^ Ву, Пин; Ян, Цзе; Лай, Юнг-Чи; Нг, Чен Сян; Ли, Анг; Цзян, Сюэюань; Элси, Рут М.; Виделиц, Рэндалл; Баджпай, Ручи; Ли, Вэнь-Сюн; Чуонг, Ченг-Мин (1 февраля 2018 г.). «Для преобразования масштаба в перо требуется несколько регуляторных модулей» . Молекулярная биология и эволюция . 35 (2): 417–430. дои : 10.1093/molbev/msx295 . ISSN 1537-1719 . ПМЦ 5850302 . ПМИД 29177513 .
- ^ Алибарди, Л.; Кнапп, ЛВ; Сойер, Р.Х. (июнь 2006 г.). «Локализация бета-кератина в развивающейся чешуе и перьях аллигатора в связи с развитием и эволюцией перьев» . Журнал субмикроскопической цитологии и патологии . 38 (2–3): 175–192. ISSN 1122-9497 . ПМИД 17784647 .
- ^ Алибарди, Л; Кнапп, ЛВ; Сойер, Р.Х. (2006). «Локализация бета-кератина в развивающейся чешуе и перьях аллигатора в связи с развитием и эволюцией перьев» . Журнал субмикроскопической цитологии и патологии . 38 (2–3): 175–92. ПМИД 17784647 .
- ^ Гринволд, Мэтью Дж.; Сойер, Роджер Х. (15 декабря 2011 г.). «Связь молекулярной эволюции птичьего бета (β) кератина с эволюцией перьев» (PDF) . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 316Б (8): 609–616. Бибкод : 2011JEZB..316..609G . дои : 10.1002/jez.b.21436 . ПМИД 21898788 . Архивировано из оригинала (PDF) 15 октября 2022 года . Проверено 15 октября 2022 г.
- ^ Слейтер, Тиффани С; Эдвардс, Николас П; Уэбб, Сэмюэл М; Чжан, Фучэн; Макнамара, Мария Э (2023). «Сохранение роговых β-белков в мезозойских перьях» . Экология и эволюция природы . 7 (10): 1706–1713. Бибкод : 2023NatEE...7.1706S . дои : 10.1038/s41559-023-02177-8 . ПМИД 37735563 . S2CID 262125827 .
- ^ Jump up to: а б Бентон, Майкл Дж.; Дуайи, Даниэль; Цзян, Баоюй; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» (PDF) . Тенденции в экологии и эволюции . 34 (9): 856–869. Бибкод : 2019TEcoE..34..856B . дои : 10.1016/j.tree.2019.04.018 . hdl : 10468/8068 . ISSN 0169-5347 . ПМИД 31164250 . S2CID 174811556 .
- ^ Даймонд, CC; Каюта Р.Дж.; Дж. С. Брукс (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального отображения адаптивной функции перьев у нептичьих теропод». БИОС . 82 (3): 58–63. дои : 10.1893/011.082.0302 . S2CID 98221211 .
- ^ Сюй, Син; Фучэн Чжан (2005). «Новый динозавр-манираптор из Китая с длинными перьями на плюсне». Naturwissenschaften . 92 (4): 173–177. Бибкод : 2005NW.....92..173X . дои : 10.1007/s00114-004-0604-y . ПМИД 15685441 . S2CID 789908 .
- ^ Сюй, Син (2006). «Пернатые динозавры из Китая и эволюция основных птичьих персонажей» . Интегративная зоология . 1 (1): 4–11. дои : 10.1111/j.1749-4877.2006.00004.x . ПМИД 21395983 . S2CID 1516713 .
- ^ Чжэн, ХТ; HL Вы; С. Сюй и З.М. Донг (2009). «Раннемеловой гетеродонтозавридный динозавр с нитевидными покровными структурами». Природа . 458 (7236): 333–336. Бибкод : 2009Natur.458..333Z . дои : 10.1038/nature07856 . ПМИД 19295609 . S2CID 4423110 .
- ^ Jump up to: а б с Сюй, Х.; Го, Ю. (2009). «Происхождение и ранняя эволюция перьев: выводы из последних палеонтологических и неонтологических данных». Позвоночные Палазиатские . 47 (4): 311–329.
- ^ Годфруа, Паскаль; Синица, София М.; Дуайи, Даниэль; Болоцкий Юрий Л.; Сизов, Александр В.; Макнамара, Мария Э.; Бентон, Майкл Дж.; Спагна, Пол (2014). «Юрский птицетазовый динозавр из Сибири с перьями и чешуей» . Наука . 345 (6195): 451–455. Бибкод : 2014Sci...345..451G . дои : 10.1126/science.1253351 . hdl : 1983/a7ae6dfb-55bf-4ca4-bd8b-a5ea5f323103 . ПМИД 25061209 . S2CID 206556907 .
- ^ Барретт, Пол М.; Эванс, Дэвид С.; Кампионе, Николас Э. (30 июня 2015 г.). «Эволюция эпидермальных структур динозавров» . Письма по биологии . 11 (6): 20150229. doi : 10.1098/rsbl.2015.0229 . ПМЦ 4528472 . ПМИД 26041865 .
- ^ Jump up to: а б Эти перья не прилипнут: моделирование максимального правдоподобия поддерживает примитивность чешуи динозавров (66-й симпозиум по палеонтологии и сравнительной анатомии позвоночных) , 9 ноября 2018 г., заархивировано из оригинала 11 декабря 2021 г. , получено 3 октября 2021 г.
- ^ Майр, Джеральд; Питтман, Майкл; Сайтта, Эван; Кэй, Томас Г.; Винтер, Якоб (30 августа 2016 г.). Бенсон, Роджер (ред.). «Строение и гомология щетинок хвоста пситтакозавра» . Палеонтология . 59 (6): 793–802. Бибкод : 2016Palgy..59..793M . дои : 10.1111/пала.12257 . hdl : 1983/029c668f-08b9-45f6-a0c5-30ce9256e593 . S2CID 89156313 .
- ^ Хольц, Томас (19 октября 2018 г.). « Покровный статус: это сложно»: филогенетические, осадочные и биологические препятствия на пути разрешения предкового покрова мезозойской динозаврии» (PDF) . Общество палеонтологии позвоночных . Проверено 16 июля 2022 г.
- ^ Ортега, Франциско; Эскасо, Фернандо; Санс, Хосе Л. (9 сентября 2010 г.). «Причудливая горбатая Carcharodontosauria (Theropoda) из нижнего мела Испании» . Природа . 467 (7312): 203–206. Бибкод : 2010Natur.467..203O . дои : 10.1038/nature09181 . ISSN 1476-4687 . ПМИД 20829793 . S2CID 4395795 .
- ^ Фот, Кристиан; Тишлингер, Гельмут; Раухут, Оливер В.М. (2 июня 2014 г.). «Новый экземпляр археоптерикса дает представление об эволюции перистых перьев» . Природа . 511 (7507): 79–82. Бибкод : 2014Natur.511...79F . дои : 10.1038/nature13467 . ПМИД 24990749 . S2CID 4464659 .
- ^ Куэста, Елена; Ортега, Франциско; Санс, Хосе Луис (4 июля 2018 г.). «Аппендикулярная остеология Concavenator corcovatus (Theropoda: Carcharodontosauridae) из нижнего мела Испании» . Журнал палеонтологии позвоночных . 38 (4): (1)–(24). дои : 10.1080/02724634.2018.1485153 . ISSN 0272-4634 . S2CID 91976402 .
- ^ Джи, К.; Пи Джей Карри; М. А. Норелл; СА Джи (1998). «Два пернатых динозавра из северо-восточного Китая» (PDF) . Природа . 393 (6687): 753–761. Бибкод : 1998Natur.393..753Q . дои : 10.1038/31635 . S2CID 205001388 .
- ^ Сюй, Х.; Его Святейшество Чжоу и Ро Прум (2001). «Разветвленные покровные структуры у синорнитозавра и происхождение перьев». Природа . 410 (6825): 200–204. Бибкод : 2001Natur.410..200X . дои : 10.1038/35065589 . ПМИД 11242078 . S2CID 4426803 .
- ^ Ху, ДЯ; Л. Х. Хоу; Эл Джей Чжан; С. Сюй (2009). «Троодонтидный теропод до археоптерикса из Китая с длинными перьями на плюсне». Природа . 461 (7264): 640–643. Бибкод : 2009Natur.461..640H . дои : 10.1038/nature08322 . ПМИД 19794491 . S2CID 205218015 .
- ^ Сюй, Х.; Ц. Чжао; М. Норелл; К. Салливан; Д. Хоун; Г. Эриксон; XL Ван; и др. (2009). «Новая окаменелость пернатого динозавра-манираптора, которая заполняет морфологический пробел в птичьем происхождении» . Китайский научный бюллетень . 54 (3): 430–435. Бибкод : 2009SciBu..54..430X . дои : 10.1007/s11434-009-0009-6 .
- ^ Витмер, LM (2009). «Пернатые динозавры в клубке» . Природа . 461 (7264): 601–602. Бибкод : 2009Natur.461..601W . дои : 10.1038/461601a . ПМИД 19794481 . S2CID 205049989 .
- ^ «Перья динозавра найдены в янтаре Альберты» . Новости ЦБК . 15 сентября 2011 г. Архивировано из оригинала 15 сентября 2011 г.
- ^ «Редкие птичьи крылья эпохи динозавров найдены в янтаре» . 28 июня 2016 года. Архивировано из оригинала 28 июня 2016 года . Проверено 28 июня 2016 г.
- ^ Син, Лида; МакКеллар, Райан С.; Ван, Мин; Бай, Мин; О'Коннор, Цзинмай К.; Бентон, Майкл Дж.; Чжан, Цзяньпин; Ван, Ян; Ценг, Куовэй (28 июня 2016 г.). «Мумифицированные ранние птичьи крылья в бирманском янтаре среднего мела» . Природные коммуникации . 7 : 12089. Бибкод : 2016NatCo...712089X . дои : 10.1038/ncomms12089 . ПМЦ 4931330 . ПМИД 27352215 .
- ^ Барон, МГ; Норман, Д.Б.; Барретт, премьер-министр (2017). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров». Природа . 543 (7646): 501–506. Бибкод : 2017Natur.543..501B . дои : 10.1038/nature21700 . ПМИД 28332513 . S2CID 205254710 .
- ^ Ян, Цзысяо; Цзян, Баоюй; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питтман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (17 декабря 2018 г.). «Покровные структуры птерозавра со сложным перьевым ветвлением» (PDF) . Экология и эволюция природы . 3 (1): 24–30. Бибкод : 2018NatEE...3...24Y . дои : 10.1038/s41559-018-0728-7 . hdl : 1983/1f7893a1-924d-4cb3-a4bf-c4b1592356e9 . ISSN 2397-334X . ПМИД 30568282 . S2CID 56480710 .
- ^ Лангер, Макс К.; Эскурра, Мартин Д.; Раухут, Оливер В.М.; Бентон, Майкл Дж.; Нолл, Фабьен; Макфи, Блэр В.; Новас, Фернандо Э.; Пол, Диего; Брусатте, Стивен Л. (2 ноября 2017 г.). «Распутывание генеалогического древа динозавров» (PDF) . Природа . 551 (7678): Е1–Е3. Бибкод : 2017Natur.551E...1L . дои : 10.1038/nature24011 . hdl : 1983/d088dae2-c7fa-4d41-9fa2-aeebbfcd2fa3 . ISSN 1476-4687 . ПМИД 29094688 . S2CID 205260354 .
- ^ Бентон, Майкл Дж.; Дуайи, Даниэль; Цзян, Баоюй; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» (PDF) . Тенденции в экологии и эволюции . 34 (9): 856–869. Бибкод : 2019TEcoE..34..856B . дои : 10.1016/j.tree.2019.04.018 . hdl : 10468/8068 . ISSN 0169-5347 . ПМИД 31164250 . S2CID 174811556 .
- ^ Jump up to: а б Фот, К. (2011). «Об идентификации структур перьев у представителей стволовой линии птиц: данные окаменелостей и актуопалеонтологии». Палеонтологическая газета . 86 : 91–102. дои : 10.1007/s12542-011-0111-3 . S2CID 86362907 .
- ^ Анвин, Дэвид М.; Мартилл, Дэвид М. (декабрь 2020 г.). «У птерозавров нет протоперьев» . Экология и эволюция природы . 4 (12): 1590–1591. Бибкод : 2020NatEE...4.1590U . дои : 10.1038/s41559-020-01308-9 . ISSN 2397-334X . ПМИД 32989266 . S2CID 222168569 .
- ^ Ян, Цзысяо; Цзян, Баоюй; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питтман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (январь 2019 г.). «Покровные структуры птерозавра со сложным перьевым ветвлением» (PDF) . Экология и эволюция природы . 3 (1): 24–30. Бибкод : 2018NatEE...3...24Y . дои : 10.1038/s41559-018-0728-7 . hdl : 1983/1f7893a1-924d-4cb3-a4bf-c4b1592356e9 . ISSN 2397-334X . ПМИД 30568282 . S2CID 56480710 .
- ^ «Новое исследование показывает, что у птерозавров было четыре типа перьев | Палеонтология | Sci-News.com» . Последние научные новости | Sci-News.com . 18 декабря 2018 года . Проверено 19 декабря 2018 г.
- ^ Сен-Флер, Николас (17 декабря 2018 г.). «Перья и мех летают над находкой окаменелостей птерозавров. Анализ двух окаменелостей отодвинет происхождение перьев примерно на 70 миллионов лет назад, но для подтверждения может потребоваться больше образцов» . Нью-Йорк Таймс . Проверено 19 декабря 2018 г.
- ^ Бриггс, Хелен (17 декабря 2018 г.). «Мех летает над новыми окаменелостями птерозавров» . Новости Би-би-си . Проверено 19 декабря 2018 г.
- ^ Д'Альба, Лилиана (2019). «Оперение птерозавра» . Экология и эволюция природы . 3 (1): 12–13. дои : 10.1038/s41559-018-0767-0 . ISSN 2397-334X . ПМИД 30568284 . S2CID 56480834 .
- ^ Цзысяо Ян; Баоюй Цзян; Мария Э. Макнамара; Стюарт Л. Кернс; Майкл Питтман; Томас Дж. Кэй; Патрик Дж. Орр; Син Сюй; Майкл Дж. Бентон (2019). «Покровные структуры птерозавра со сложным перистым ветвлением» . Экология и эволюция природы . 3 (1): 24–30. Бибкод : 2018NatEE...3...24Y . дои : 10.1038/s41559-018-0728-7 . hdl : 1983/1f7893a1-924d-4cb3-a4bf-c4b1592356e9 . ПМИД 30568282 . S2CID 56480710 .
- ^ Анвин, DM; Мартилль, DM (2020). «У птерозавров нет протоперьев». Экология и эволюция природы . 4 (12): 1590–1591. Бибкод : 2020NatEE...4.1590U . дои : 10.1038/s41559-020-01308-9 . ПМИД 32989266 . S2CID 222168569 .
- ^ Ян, З.; Цзян, Б.; Макнамара, Мэн; Кернс, СЛ; Питтман, М.; Кэй, Т.Г.; Орр, ПиДжей; Сюй, Х.; Бентон, MJ (2020). «Ответ на: У птерозавров нет протоперьев». Экология и эволюция природы . 4 (12): 1592–1593. Бибкод : 2020NatEE...4.1592Y . дои : 10.1038/s41559-020-01309-8 . hdl : 10468/11874 . ПМИД 32989267 . S2CID 222163211 .
Дальнейшее чтение
- Хэнсон, Тор (2011). Перья: эволюция природного чуда . Нью-Йорк: Основные книги. ISBN 978-0-465-02013-3 .
- Лукас, Альфред М.; Штеттенхайм, Питер Р. (1972). «Строение перьев» . Птичья анатомия Покровы . Том. Часть 1. Вашингтон, округ Колумбия: Министерство сельского хозяйства США. стр. 235–276.
Внешние ссылки
- МакГроу, К.Дж. 2005. Полли хочет пигмент? Расшифровка химического кода красной окраски попугаев. Австралийский журнал Birdkeeper 18: 608–611.
- ДеМео, Антония М. Доступ к орлам и частям орлов: защита окружающей среды против свободного отправления религии коренными американцами (1995)
- Электронный свод федеральных правил (e-CFR), раздел 50: Дикая природа и рыболовство, ЧАСТЬ 22 — РАЗРЕШЕНИЯ EAGLE
- США против Тридцати восьми Золотых орлов (1986)
- Механическое строение перьев
- Документальный фильм об эволюции перьев
- Конспект лекций по покровам птиц
- Атлас перьев Национальной лаборатории криминалистики рыб и дикой природы США
- Federn.org