Птичий мозг
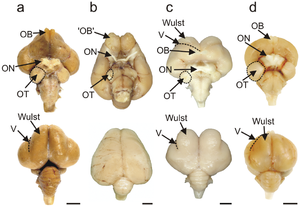
Птичий мозг — центральный орган нервной системы птиц. Птицы обладают большим и сложным мозгом , который обрабатывает , интегрирует и координирует информацию, полученную из окружающей среды, и принимает решения о том, как реагировать остальными частями тела. у всех хордовых , птичий мозг находится в костях черепа головы Как и .
Мозг птицы разделен на несколько отделов, каждый из которых выполняет свою функцию. Головной мозг или телэнцефалон разделен на два полушария и контролирует высшие функции. В конечном мозге преобладает большой мантий , который соответствует млекопитающих коре головного мозга и отвечает за когнитивные функции птиц. Паллий состоит из нескольких основных структур: гиперпаллия, дорсальной выпуклости паллия, встречающейся только у птиц, а также нидопаллия, мезопаллия и архипаллия. Ядерная структура конечного мозга птиц, в которой нейроны распределены в трехмерно расположенных кластерах, без крупномасштабного разделения белого и серого вещества , хотя существуют слоистые и столбчатые связи. Структуры мантии связаны с восприятием , обучением и познанием . Под паллием расположены два компонента субпаллия: полосатое тело и паллидум . Субпаллиум соединяет различные части конечного мозга и играет важную роль в ряде критических форм поведения. В задней части конечного мозга располагаются таламус , средний мозг и мозжечок . соединяет Задний мозг остальную часть головного мозга со спинным мозгом.
Размер и структура птичьего мозга позволяют птицам совершать заметные действия, такие как полет и вокализация . Специальные структуры и пути объединяют слуховые и зрительные чувства, сильные у большинства видов птиц, а также обычно более слабые обонятельные и тактильные чувства. Социальное поведение , широко распространенное среди птиц, зависит от организации и функций мозга. Некоторые птицы демонстрируют сильные познавательные способности, обусловленные уникальной структурой и физиологией птичьего мозга.
Структура и функции
[ редактировать ]Теленцефалон
[ редактировать ]
Теленцефалон , или передний мозг , обычно представляет собой крупную структуру у птиц. Он состоит из трех основных доменов: стриарного домена, паллидного домена и паллиального домена. [1]
Плащ
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( октябрь 2023 г. ) |
Паллиальный домен разделен на две основные части: Вульст, или гиперпаллий , на дорсальной поверхности паллиума и дорсо-желудочковый гребень, который включает нидопаллий , аркопаллий и архипаллий . Схема птичьего мозга устроена таким образом, что цепи таких процессов, как зрение и познание, проходят между структурами мантии, при этом связи внутри мантии ориентированы как горизонтально, так и вертикально. [2] В сенсорных областях мантии нейроны имеют слоистую организацию в виде столбцов с горизонтальными связями; следовательно, часть паллия, ответственная за сенсорную схему, также называется сенсорной корой . у птиц [3]
Считается, что в мантии нидопаллиум каудолатере является центром целенаправленного действия. Это часть мантии, которая обладает наиболее иннервацией чувствительной к дофамину и имеет структуру, предполагающую сильный дофаминовый контроль. Нейронная активность nidopallium caudolatere коррелирует с вознаграждениями, правилами, категориями и информацией, хранящейся в памяти . Нидопаллиум каудолатере также пропорционально намного крупнее у птиц с более сильными когнитивными способностями. Разнообразие мозга птиц и, возможно, роль nidopallium caudolatere заключается в разделении этой структуры на несколько структур у певчих птиц. [2]
Латерализация
[ редактировать ]Птичий конечный мозг, как и его человеческий аналог, состоит из двух отдельных полушарий . Такие функции, как обучение пению у певчих птиц, зрение, слуховая и обонятельная, а также магнитная сенсорная системы , являются латерализованными, а это означает, что одно полушарие мозга доминирует над определенной функцией. Левое полушарие мозга доминирует над деятельностью, требующей отделения важных стимулов от отвлекающих, тогда как правое полушарие легче отвлекается и обладает более широким вниманием. Правое полушарие также доминирует над реакциями страха и бегства . [4] У зебровых амадин ( Taeniopygia Guttata ) левое полушарие также доминирует в задачах обучения. [5] Птицы могут чувствовать ориентацию магнитных полей и использовать их для навигации. Правое полушарие мозга является доминирующим в магнетическом смысле. [4]
Субпаллиальный телэнцефалон
[ редактировать ]Субпаллиальный тенецефалон птиц, вентральнее мантия, содержит нервные системы с критическими функциями. Его можно разделить на полосатый домен и паллидный домен, которые являются зонами-предшественниками, из которых развиваются структуры мозга. Субпаллиальный конечный мозг также сгруппирован в несколько основных функциональных систем. Это дорсальные соматомоторные базальные ганглии , вентральные висцеролимбические базальные ганглии, субпаллиальная расширенная миндалина, базальная кортикопетальная система и перегородка. Кроме того, преоптическая область считается частью субпаллиального конечного мозга. Функциональные системы существуют в обоих субпаллиальных доменах. [6]
Исследования 1960-х годов показали, что базальные ганглии птиц занимают только вентромедиальную часть конечного мозга , а не весь передний мозг, - исторически сложившееся мнение. Базальные ганглии включают дорсальные соматомоторные базальные ганглии (DSBG), которые обладают как паллидным, так и стриарным компонентом и состоят из медиального полосатого тела, латерального полосатого тела, бледного шара и внутриножкового ядра. У певчих птиц он также содержит область X, которая отвечает за аспекты голосовой функции певчих птиц. DSBG выполняет важные функции в произвольном управлении моторикой. Пути, включающие DSBG, позволяют птицам совершать движения, когда это необходимо, и отказываться от движения, когда оно нежелательно. К базальным ганглиям также относятся вентральные висцеролимбические базальные ганглии (ВВБГ). VVBG, как и DSBG, занимает часть как полосатого тела, так и паллидума. Он содержит вентральную часть медиального полосатого тела, прилежащее ядро , обонятельный бугорок и вентральное бледное тело. ВВБГ функционирует как « центр вознаграждений ». а также посредник в « выборе действий ». В этих ролях VVBG играет важную роль в поддержке поведения, направленного на вознаграждение , и в противодействии поведению, ведущему к негативным стимулам . [7]
Зона X, уникальная для певчих птиц, имеет решающее значение для обучения пению , которое является критически важным для размножения поведением певчих птиц. Ученые описали его как аналог полосатого тела млекопитающих из-за схожего состава с этой частью мозга. Однако он также содержит клетки головного мозга паллидума. Область X активируется наиболее сильно, когда певчие птицы изучают новые песни, и после этого ее активность снижается. [8]
Расширенная миндалина состоит из центральной расширенной миндалевидного тела и медиальной расширенной миндалевидного тела. Центральная расширенная миндалина связана с поведением, связанным с приемом пищи , а также со стрессом, тревогой и страхом. Роль и структура медиальной расширенной миндалины обсуждаются учеными. Медиальная расширенная миндалина получает сигналы от обонятельной луковицы , а также от остальной части обонятельной системы , включая органы обоняния в передней части головы птицы. Это связано с сексуальным и социальным поведением. Было показано, что некоторые структуры срединной расширенной миндалевидного тела у птиц имеют половой диморфизм . Исследования показывают, что у кур ( Gallus Gallus ) часть средней расширенной миндалевидного тела играет роль в сексуальном поведении самцов. [7]
Базальная кортикопетальная система у птиц представлена тремя ядрами : базальным магноцеллюлярным ядром , а также горизонтальным и вертикальным коленами ядер диагональной полосы. Роль базальной кортикопетальной системы у птиц плохо изучена, хотя известно, что эта система коррелирует с памятью у млекопитающих. Однако было показано, что повреждение базальной кортикопетальной системы ухудшает память птенцов ( Gallus Gallus ). [7]
На основании исследований различных воробьиных птиц перегородка птиц разделена на четыре основные части: латеральную перегородку, медиальную перегородку, септо -гиппокампальную перегородку и каудоцентральную перегородку. Перегородка выполняет функции, связанные со стрессом , а также реакцией на такие раздражители , как свет . Преоптическая область выполняет функции, связанные с сексуальным поведением. [7]
Ствол мозга и мозжечок
[ редактировать ]Мозжечок
[ редактировать ]Мозжечок типов — относительно консервативная часть мозга, схема которой, как правило, одинакова у разных позвоночных . Как и у млекопитающих, мозжечок птиц представляет собой сильно складчатую структуру. Птичий мозжечок обычно делится на десять различных групп складок, называемых дольками. У некоторых групп птиц мозжечок расширен для удовлетворения различных потребностей. Например, у птиц, совершающих много манипуляций с клювом (вороны, дятлы, попугаи), наблюдается расширение зрительной части мозжечка. Напротив, часть мозжечка, связанная с хвостом, расширена. [9]
Существуют значительные различия в слоении (складчатости) мозжечка птиц, хотя у всех птиц имеется хотя бы некоторая складчатость мозжечка. Складчатость мозжечка также не связана строго со структурой и размером остального мозга: пингвины, морские птицы, попугаи и вороны демонстрируют одинаковую степень складчатости, несмотря на очень разные характеристики мозга. Аналогичным образом, совы, курообразные и голуби имеют схожие модели складывания. [10] В целом структура складок мозжечка у птиц отражает различия в поведении, а также различия в форме черепа, ограничивающие развитие мозжечка, а также сенсорные и сенсомоторные потребности животных, ведущих разный образ жизни. [11]
Физиология
[ редактировать ]Метаболизм
[ редактировать ]Нейроны , как правило, представляют собой энергоемкие клетки, содержание которых требует высоких затрат, однако у птиц высокая плотность и абсолютное количество нейронов в мозгу. Из энергии, потребляемой мозгом, около 70-80% используется нейронами. Большое количество нейронов в птичьем мозге обеспечивается относительно низкими удельными энергетическими потребностями. Эксперименты с голубями показали, что потребность нейронов птиц в глюкозе более чем в 3 раза меньше, чем у млекопитающих, у которых энергетические затраты нейронов не меняются между таксонами. Пока неизвестно, почему птичьему мозгу требуется гораздо меньше глюкозы, но два способствующих этому фактора: размер нейронов и температуру тела исследователи установили . Хотя по состоянию на 2023 год окончательных исследований по этой теме не проводилось, предполагается, что нейроны птиц меньше, чем у млекопитающих, поскольку мозг птиц имеет гораздо более высокую плотность нейронов на единицу объема по сравнению с мозгом млекопитающих аналогичного размера. Нейроны меньшего размера могут потреблять меньше энергии, поскольку их меньшая площадь поверхности и объем содержат меньше рецепторы , ионные каналы и митохондрии мембраны , обладая при этом более низкой емкостью , а это означает, что для изменения мембранного потенциала требуется меньше энергии. Меньший нейрон также требует меньше энергии для поддержания. Кроме того, более высокая температура мозга птиц, достигающая 42 °C (108 °F) у голубей , также способствует снижению потребления энергии . Более высокие температуры сокращают время, необходимое для активации и деактивации ионных каналов, что снижает потребности в энергии. [12]
Эволюция
[ редактировать ]Мозг птиц сегодня заметно отличается от мозга млекопитающих, от которых в перми отделилась линия птиц , и от всех других рептилий, от которых птицы разошлись в триасе .
Нидопаллиум каудолатере птичьего мозга, отвечающий за целенаправленное действие, был обнаружен у крокодилов , ближайших ныне живущих родственников птиц , эволюционно разделенных 245 миллионами лет. Хотя это может представлять собой пример конвергентной эволюции крокодилов и птиц, учёные полагают, что более вероятно, что последний общий предок птиц и крокодилов обладал nidopallium caudolatere. [3]
В юрском и меловом периоде
[ редактировать ]Птицы произошли от нептичьих динозавров , а линия динозавров, ведущая к птицам, демонстрирует значительное увеличение и изменение формы мозга. Эндокасты динозавров черепных коробок позволяют ученым изучить взаимосвязь между размером мозга и размером тела у вымерших видов. В частности, размеры мозга претерпели несколько значительных изменений от первых динозавров- теропод до современных птиц. Размеры и форма мозга большинства динозавров аналогичны таковым у современных рептилий. [13] Однако у целурозаврии , группы динозавров, содержащей (среди других таксонов ) тираннозавроидов и птиц, мозг как минимум в два раза больше, чем у других динозавров аналогичного размера тела. [14] Группа целурозавров, Maniraptoriformes , в которую входят птицы, а также орнитомимозавры , обладает мозгом, который снова в два раза больше, чем у других целурозавров. [14] Размер мозга снова увеличивается у Maniraptora , группы, состоящей из птиц и овирапторозавров . [14] Кроме того, форма мозга, приобретенная у Maniraptora, с большим мозжечком и зрительной долей, расположенной латерально от конечного мозга, [15] обнаружен у Ихтиорниса , птицы, жившей 70 миллионов лет назад и тесно связанной с современными птицами. [16] Понимание эволюции размера мозга современных птиц осложняется ограниченной выборкой эндокастов мозга стволовых птиц. Только четыре мезозойских авиалана имеют эндокасты головного мозга: археоптерикс , который не всегда классифицируется как авиалан (ближе к современным птицам, чем к дромеозавру ), [14] Cerebavis , неполный экземпляр, у которого отсутствует гиперпаллий и классификация которого неясна, Ichthyornis и MPM-334-1, базикраний , принадлежавший энантиорнитиновой птице возрастом восемьдесят миллионов лет, сохранивший задний мозг . [16] [17] Ни один конечный мозг не известен из окончательных примеров Enanitornithes, самой большой и самой разнообразной группы птиц за пределами радиации современных птиц. [16]

Археоптерикс традиционно считается первой птицей, и иногда его называют Урвогелем (первоначальная птица). [18] Несмотря на то, что мозг археоптерикса почти наверняка был приспособлен к полету, конечный мозг археоптерикса не был особенно большим по сравнению с родственными ему динозаврами. Действительно, несколько овирапторозавров, а также троодонт Занабазар и джинфенгоптеригин троодонтид. [19] IGM100/1126 обладают мозгом, превышающим размер тела, чем у археоптерикса. Другие элементы черепной анатомии археоптерикса также аналогичны таковым у других динозавров-манирапторов, что позволяет предположить, что эти животные также могли иметь необходимые неврологические возможности для полета . [20]
Вульст, физически выступающий гиперпаллий, представляет интерес для эволюции мозга птиц, поскольку он отсутствует ни у каких других ныне живущих рептилий, кроме птиц. Более того, считается, что он аналогичен неокортексу млекопитающих и играет важную роль в высшем познании. Ни один нептичий динозавр не обладает Вульстом, как и Археоптерикс , примитивная птица. Однако птицевидный Ichthyornis, несмотря на то, что форма его мозга напоминает примитивных манирапторанов , обладает Wulst, показывая, что эта структура, вероятно, возникла раньше в эволюции птиц и существует за пределами современных птиц. [16]
В целом соотношение мозга и тела динозавров удвоилось от базальных теропод до целурозаврии и снова удвоилось от целурозаврии до манирапториформ . От Maniraptoriformes общая форма мозга приняла форму, которая сохранится по крайней мере у ихтиорнисов, близких к современным птицам. [14] произошли три основных изменения в соотношении мозга и тела Ученые предполагают, что в ходе эволюции птиц от базального Paraves до основания кроны Neoaves . [21]
В кайнозое
[ редактировать ]У современных птиц существует существенное разнообразие размеров и расположения мозга. В палеогене средний размер мозга птиц по отношению к телу имел тенденцию к увеличению; во многих линиях, возникших в результате палеогеновой радиации птиц, размер мозга имел тенденцию увеличиваться быстрее, чем размер тела. [22] [21] Ученые предполагают, что такой рост размера мозга произошел в результате адаптивной радиации, которую птицы испытали при переходе K-Pg , и что более крупный мозг коронных птиц позволил им адаптироваться к нестабильной окружающей среде. [22]
Огромное разнообразие современных птиц приводит к разнообразию моделей мозга. В кладе Neoaves , в которую входят все птицы, за исключением птиц и палеогнатов , соотношение мозга и тела увеличивается, но это обусловлено, прежде всего, уменьшением среднего размера тела. [22] [21] Такая закономерность наблюдается у стрижей , колибри , куликов , перепелов , а также у линии, ведущей к Теллуравсу , «высшим наземным птицам». Однако многие группы водоплавающих птиц , известные под общим названием Aequornithes , не следуют этой тенденции, а скорее имеют тенденцию увеличивать размер тела и размер мозга с одинаковой скоростью. Самые базовые теллюравы — это две ветви крупных хищных птиц. Один, в Afroaves , состоит из сов , у которых большой мозг развился для остроты зрения, и Accipitriformes , включая ястребов и орлов . Другой, в Австралавесе , включает соколов , серием и вымерших ужасных птиц . Внутри афроавов наблюдается последовательный сдвиг в сторону более высокого соотношения мозга и тела. и У мышевидных птиц катунов Coraciimorphae соотношение мозга к телу больше, чем у афроавов, а у дятлов, гнездящихся в Coraciimorphae, соотношение еще больше. [21]
Некоторые из самых больших соотношений мозга и тела у птиц, особенно конечного мозга, той части мозга, которая отвечает за познание, обнаружены у Psittaciformes (попугаи) и Corvidae (вороны, вороны, сойки, сороки и их союзники). оба члены австралавов. [23] Действительно, попугаи и врановые уникальны среди птиц своим большим размером мозга. Более того, ученые полагают, что увеличение соотношения мозга и тела у птиц развивалось быстрее всех других изменений соотношения мозга и тела. [21]
В отличие от того, что в настоящее время считается относительно небольшим изменением соотношения мозга и тела птиц в мезозое, исследователи обнаружили, что девять таких сдвигов произошли в палеоцене , возможно, в ответ на вымирание K-Pg. событие . Более того, они обнаружили, что наибольшие размеры мозга у птиц развились лишь недавно, с неогеновой радиацией у коронованных врановых и коронованных попугаев. Более того, в этих линиях плотность нейронов в мозге также увеличивается, что способствует значительной когнитивной сложности. [21]
Сравнение с мозгом млекопитающих
[ редактировать ]Мозг птиц часто сравнивают с мозгом млекопитающих. Первые нейробиологи, активно изучавшие птиц, такие как Людвиг Эдингер , были поражены различиями, которые они наблюдали между мозгом птиц и млекопитающих. Сегодня взаимоотношения между гомологичными структурами в мозге птиц и млекопитающих более известны; кроме того, наблюдались многие конвергентные особенности мозга птиц и млекопитающих. Некоторые из сходств между мозгом птиц и млекопитающих включают обработку специализированной сенсорной информации, участие в более высоких когнитивных функциях, высокую плотность нейронов и структуру волокон мозга. Более того, птичий мозг демонстрирует наличие сенсорного сознания . [24] Многие из структур, которые, как считается, способствуют интеллекту млекопитающих, такие как шестислойный неокортекс , отсутствуют у птиц. Несмотря на это, такие птицы, как врановые и попугаи, демонстрируют интеллектуальное поведение, сравнимое с поведением высокоинтеллектуальных млекопитающих, таких как человекообразные обезьяны . Ученые считают, что это пример конвергентной эволюции , когда радикально разные грубые структуры эволюционировали в сторону связующего сходства, что привело к сопоставимым результатам. [25]
Исчерченность паллия
[ редактировать ]У млекопитающих неокортекс, тесно связанный с высшим мышлением и познанием, многослойный и состоит из 6 слоев. У птиц подобная слоистость отсутствует на большей части мантия, соответствующего коре головного мозга. [24] Однако гиперпаллий дорсального конечного мозга действительно имеет некоторую слоистость: либо три, либо четыре слоя, в зависимости от размера гиперпаллия рассматриваемой птицы. И в неокортексе млекопитающих, и в гиперпаллии птиц слои имеют разные типы связей и нейрохимический состав. [25]
Аналоги префронтальной коры
[ редактировать ]Префронтальная кора млекопитающих, активно вовлеченная в процессы, поддерживающие сложное обучение, может иметь аналог у птиц в виде nidopallium caudolatere. Исследователи обнаружили, что существует сходство между тем, как нейроны каудолатерального нидопаллия активируются в ответ на определенные тесты, разработанные для префронтальной коры, аналогично тому, как ведут себя сами нейроны префронтальной коры. Повреждение каудолатерального нидопаллия также вызывает недостатки в выполнении этого класса задач. Однако роль каудолатерального нидопаллия до сих пор не ясна, и он был протестирован только на голубях, обладающих относительно небольшим мозгом. Таким образом , роль каудолатерального нидопаллия в более сложных целенаправленных действиях неясна. [25] [26]
История
[ редактировать ]Научное изучение мозга птиц ускорилось в 19 веке, когда новые методы подготовки мозга позвоночных стали более совершенными. На рубеже 20-го века немецкий невролог Людвиг Эдингер разработал теорию эволюции головного мозга позвоночных , которая вскоре стала доминирующей научной точкой зрения. Затем голландский невролог К.У. Ариенс-Капперс усовершенствовал модель, адаптировав ее к птичьему мозгу. Следуя линейной модели эволюции, Ариенс-Капперс выдвинул идею о том, что структуры мозга возникали последовательно в ходе эволюционных линий , называемую аккреционной теорией. Согласно его теории, птицы и млекопитающие унаследовали от рыб и амфибий примитивные базальные ганглии, известные как палеостриатум . Они также унаследовали от рептилий новые базальные ганглии, известные как неостриатум . Предполагалось, что рептилии развили неостриатум в две разные структуры, которые затем были переданы птицам. Ариенс-Капперс также предположил, что самый верхний слой птичьего мозга образован клетками полосатого мозга, содержащими массу, которую он назвал гиперстриатумом. Поэтому в течение многих лет считалось, что конечный мозг птиц почти полностью состоит из крупных базальных ганглиев. [27] Первый стереотаксический атлас птичьего мозга, вышедший в 1967 году, последовал за взглядом Ариена-Каппера на птичий мозг и еще больше закрепил неправильную концепцию «примитивного» птичьего мозга. [28]
В течение многих лет ученые предполагали, что птицы не способны к продвинутому мышлению, поскольку считалось, что их мозг лишен сложных мантийных структур. Более того, у них отсутствовали поперечно-полосатые структуры, подобные тем, которые обнаружены в коре головного мозга млекопитающих, которые, как считалось, отвечают за сложное познание . На самом деле неврологи считали, что птицы — это существа, которые действуют скорее инстинктивно , чем на основании каких-либо мыслей, и что они крайне неразумны. Из этой концепции возник разговорный термин « птичий мозг» , используемый для унижения людей как неразумных. [27]
Исследования познания, поведения и анатомии птиц, а также мозга, в частности, показали, что традиционный аккреционный взгляд на эволюцию телэнцефалов позвоночных неверен. Стало также ясно, что то, что тогда называли полосатыми частями мозга, на самом деле имело паллиальное происхождение. В конце 1990-х и начале 2000-х годов началось движение, достигшее кульминации в 2002 году, в результате которого была создана новая номенклатура птичьего мозга, которая с тех пор стала стандартной. [28] Новое исследование нейронных сетей птиц привело к лучшему пониманию структуры и организации птичьего мозга. [29]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Джарвис, Эрих Д.; Гюнтюркюн, Онур; Брюс, Лаура; Чиллаг, Андраш; Картен, Харви; Куэнзель, Уэйн; Медина, Лорета; Паксинос, Джордж; Перкель, Дэвид Дж.; Симидзу, Тору; Стридтер, Георг; Уайлд, Дж. Мартин; Болл, Грегори Ф.; Дугас-Форд, Дженнифер; Дюран, Сара Э. (1 февраля 2005 г.). «Птичий мозг и новое понимание эволюции мозга позвоночных» . Обзоры природы Неврология . 6 (2): 151–159. дои : 10.1038/nrn1606 . ISSN 1471-0048 . ПМК 2507884 . ПМИД 15685220 .
- ^ Jump up to: а б Роуз, Джонас (24 октября 2022 г.). «Птичий мозг» (PDF) . Современные учебники по биологии . 32 (20): Р1076–Р1079. дои : 10.1016/j.cub.2022.07.072 . ПМИД 36283368 . S2CID 253105801 . Архивировано (PDF) из оригинала 9 октября 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б Гюнтюркюн, Онур; фон Ойген, Кая; Пакхайзер, Джулиан; Пуш, Роланд (01 декабря 2021 г.). «Птичьи мантии и познание: сравнение с млекопитающими» . Современное мнение в нейробиологии . Эволюция мозга и вычислений. 71 : 29–36. дои : 10.1016/j.conb.2021.08.007 . ISSN 0959-4388 . ПМИД 34562800 . S2CID 237603077 . Архивировано из оригинала 9 октября 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б Роджерс, Лесли Дж. (15 июня 2008 г.). «Развитие и функция латерализации в мозгу птиц» . Бюллетень исследований мозга . Специальный выпуск: Механизмы мозга, познание и поведение птиц. 76 (3): 235–244. дои : 10.1016/j.brainresbull.2008.02.001 . ISSN 0361-9230 . ПМИД 18498936 . S2CID 26031891 .
- ^ Мурман, Санне; Гобес, Шэрон М.Х.; ван де Камп, Фердинанд К.; Зандберген, Маттейс А.; Болхуис, Йохан Дж. (12 марта 2015 г.). «Доминирование полушарий мозга, связанное с обучением, у спящих певчих птиц» . Научные отчеты . 5 (1): 9041. Бибкод : 2015NatSR...5E9041M . дои : 10.1038/srep09041 . ISSN 2045-2322 . ПМК 4356971 . ПМИД 25761654 .
- ^ Куэнзель, Уэйн Дж.; Медина, Лорета; Чсиллаг, Андрас; Перкель, Дэвид Дж.; Райнер, Антон (18 ноября 2011 г.). «Птичий субпаллий: новое понимание структурных и функциональных подразделений, занимающих латеральную субпаллиальную стенку, и их эмбриологического происхождения» . Исследования мозга . 1424 : 67–101. дои : 10.1016/j.brainres.2011.09.037 . ISSN 0006-8993 . ПМЦ 3378669 . ПМИД 22015350 .
- ^ Jump up to: а б с д Сканес, Колин Г.; Дриди, Сами (2021). Птичья физиология Стерки (7-е изд.). Академическая пресса. стр. 257–274. ISBN 9780323853514 .
- ^ Чжан, Ютао; Чжоу, Лифан; Цзо, Цзячунь; Ван, Сунгари; Мэн, Вэй (2023). «Аналогии человеческой речи и пения птиц: от вокального обучения до его нейронной основы» . Границы в психологии . 14 . дои : 10.3389/fpsyg.2023.1100969 . ISSN 1664-1078 . ПМЦ 9992734 . ПМИД 36910811 .
- ^ Султан, Фахад (6 сентября 2005 г.). «Почему у некоторых птиц мозг больше, чем у других» . Современная биология . 15 (17): 649–650 р. дои : 10.1016/j.cub.2005.08.043 . ISSN 0960-9822 . ПМИД 16139191 . S2CID 6559127 . Архивировано из оригинала 24 февраля 2024 г. Проверено 7 ноября 2023 г.
- ^ Иванюк, Андрей Н.; Херд, Питер Л.; Уайли, Дуглас Р.В. (14 июня 2006 г.). «Сравнительная морфология птичьего мозжечка: I. Степень слоения» . Мозг, поведение и эволюция . 68 (1): 45–62. дои : 10.1159/000093530 . ISSN 0006-8977 . ПМИД 16717442 . S2CID 1633943 . Архивировано из оригинала 24 февраля 2024 г. Проверено 7 ноября 2023 г.
- ^ Иванюк, Андрей Н.; Херд, Питер Л.; Уайли, Дуглас Р.В. (13 ноября 2006 г.). «Сравнительная морфология птичьего мозжечка: II. Размер листков» . Мозг, поведение и эволюция . 69 (3): 196–219. дои : 10.1159/000096987 . ПМИД 17108672 . S2CID 2747025 . Архивировано из оригинала 24 февраля 2024 г. Проверено 7 ноября 2023 г.
- ^ фон Ойген, Кая; Эндеполс, Хайке; Джезга, Александр; Ноймайер, Бернд; Гюнтюркюн, Онур; Бэкес, Хейко; Стрёкенс, Феликс (10 октября 2022 г.). «Нейроны птиц потребляют в три раза меньше глюкозы, чем нейроны млекопитающих» . Современная биология . 32 (19): 4306–4313.e4. дои : 10.1016/j.cub.2022.07.070 . ISSN 0960-9822 . ПМИД 36084646 . S2CID 252124960 . Архивировано из оригинала 24 февраля 2024 г. Проверено 8 октября 2023 г.
- ^ Ларссон, Ханс CE; Серено, Пол С.; Уилсон, Джеффри А. (25 сентября 2000 г.). «Увеличение переднего мозга у нептичьих динозавров-теропод» . Журнал палеонтологии позвоночных . 20 (3): 615–618. doi : 10.1671/0272-4634(2000)020[0615:FEANTD]2.0.CO;2 . ISSN 0272-4634 . S2CID 29413008 . Архивировано из оригинала 9 октября 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б с д и Хольц, Томас Р. (2012). Полный динозавр (2-е изд.). Блумингтон: Издательство Университета Индианы. стр. 358–372. ISBN 9780253008497 .
- ^ Алонсо, Патрисио Домингес; Милнер, Анджела С.; Кетчем, Ричард А.; Куксон, М. Джон; Роу, Тимоти Б. (5 августа 2004 г.). «Птичья природа мозга и внутреннего уха археоптерикса» . Природа . 430 (7000): 666–669. Бибкод : 2004Natur.430..666A . дои : 10.1038/nature02706 . ISSN 1476-4687 . ПМИД 15295597 . S2CID 4391019 . Архивировано из оригинала 5 декабря 2022 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б с д Торрес, Кристофер Р.; Норелл, Марк А.; Кларк, Джулия А. (30 июля 2021 г.). «Эволюция нейрокраниала и массы тела птиц в период массового вымирания в конце мелового периода: форма птичьего мозга оставила позади других динозавров» . Достижения науки . 7 (31). Бибкод : 2021SciA....7.7099T . дои : 10.1126/sciadv.abg7099 . ISSN 2375-2548 . ПМК 8324052 . ПМИД 34330706 .
- ^ Кьяппе, Луис М.; Навалон, Гильермо; Мартинелли, Агустин Г.; Нава, Уильям; Филд, Дэниел Дж. (28 сентября 2022 г.). «Ископаемый базикраний проясняет происхождение центральной нервной системы и внутреннего уха птиц» . Труды Королевского общества B: Биологические науки . 289 (1983). дои : 10.1098/rspb.2022.1398 . ISSN 0962-8452 . ПМЦ 9515635 . ПМИД 36168759 .
- ^ «Старая, новая одежда Урфогеля» . Наука . 02 июля 2014 г. Архивировано из оригинала 16 мая 2021 года . Проверено 8 октября 2023 г.
- ^ Хартман, Скотт; Мортимер, Микки; Уол, Уильям Р.; Ломакс, Дин Р.; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ISSN 2167-8359 . ПМК 6626525 . ПМИД 31333906 .
- ^ Баланофф, Эми М.; Бевер, Гейб С.; Роу, Тимоти Б.; Норелл, Марк А. (31 июля 2013 г.). «Эволюционное происхождение птичьего мозга» . Природа . 501 (7465): 93–96. Бибкод : 2013Natur.501...93B . дои : 10.1038/nature12424 . ISSN 1476-4687 . ПМИД 23903660 . S2CID 4451895 . Архивировано из оригинала 31 октября 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б с д и ж Ксепка, Дэниел Т.; Баланофф, Эми М.; Смит, Н. Адам; Бевер, Габриэль С.; Бхуллар, Бхарт-Анджан С.; Бурдон, Эстель; Браун, Эдвард Л.; Берли, Дж. Гордон; Кларк, Джулия А.; Кольбер, Мэтью В.; Корфилд, Джереми Р.; Дегранж, Федерико Дж.; Де Пьетри, Ванеса Л.; Рано, Кэтрин М.; Филд, Дэниел Дж. (08 июня 2020 г.). «Темп и закономерности эволюции размера птичьего мозга» . Современная биология . 30 (11): 2026–2036.e3. дои : 10.1016/j.cub.2020.03.060 . hdl : 11336/141993 . ISSN 0960-9822 . ПМИД 32330422 . S2CID 216095924 . Архивировано (PDF) из оригинала 16 июля 2020 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б с «Эволюция птичьего мозга» . Американский учёный . 2021-09-23. Архивировано из оригинала 7 июля 2022 г. Проверено 8 октября 2023 г.
- ^ Олькович, Северин; Кокоурек, Мартин; Лучан, Радек К.; Портеш, Михал; Фитч, В. Текумсе; Эркулано-Хузель, Сюзана; Немец, Павел (28 июня 2016 г.). «Птицы имеют такое же количество нейронов в переднем мозге, как и приматы» . Труды Национальной академии наук . 113 (26): 7255–7260. Бибкод : 2016PNAS..113.7255O . дои : 10.1073/pnas.1517131113 . ISSN 0027-8424 . ПМЦ 4932926 . ПМИД 27298365 .
- ^ Jump up to: а б Болл, Грегори Ф.; Бальтазар, Жак (12 июля 2021 г.). «Эволюционная нейробиология: действительно ли мозг птиц и млекопитающих настолько различен?» . Современная биология . 31 (13): Р840–Р842. дои : 10.1016/j.cub.2021.05.004 . ISSN 0960-9822 . ПМИД 34256914 . S2CID 235802619 . Архивировано из оригинала 9 октября 2023 г. Проверено 8 октября 2023 г.
- ^ Jump up to: а б с Эмери, Натан Дж.; Клейтон, Никола С. (2005). «Эволюция птичьего мозга и интеллекта» (PDF) . Современные учебники по биологии . 15 (23): 946–950 р. дои : 10.1016/j.cub.2005.11.029 . ПМИД 16332520 . S2CID 8969952 . Архивировано (PDF) из оригинала 26 июля 2022 г. Проверено 8 октября 2023 г.
- ^ Лю, Синьюй; Ван, Хун; Ли, Шан; Шан, Чжиган; Ши, Ли (30 мая 2017 г.). «Роль нидопаллия хвостолатерального в целенаправленном поведении голубей» . Поведенческие исследования мозга . 326 : 112–120. дои : 10.1016/j.bbr.2017.02.042 . ISSN 0166-4328 . ПМИД 28288807 . S2CID 4282299 .
- ^ Jump up to: а б Эмери, Натан Дж (07 декабря 2005 г.). «Когнитивная орнитология: эволюция птичьего интеллекта» . Философские труды Королевского общества B: Биологические науки . 361 (1465): 23–43. дои : 10.1098/rstb.2005.1736 . ISSN 0962-8436 . ПМК 1626540 . ПМИД 16553307 .
- ^ Jump up to: а б Райнер, Антон; Перкель, Дэвид Дж.; Мелло, Клаудио В.; Джарвис, Эрих Д. (16 июня 2004 г.). «Певчие птицы и пересмотренная номенклатура птичьего мозга» . Анналы Нью-Йоркской академии наук . 1016 (1): 77–108. Бибкод : 2004NYASA1016...77R . дои : 10.1196/анналы.1298.013 . ISSN 0077-8923 . ПМЦ 2481519 . ПМИД 15313771 .
- ^ Хардинг, Шерил Ф. (1 мая 2004 г.). «Изучение мозга птиц: как изучение мозга певчих птиц произвело революцию в нейробиологии» . Лабораторное животное . 33 (5): 28–33. дои : 10.1038/laban0504-28 . ISSN 0093-7355 . ПМИД 15141244 . S2CID 751995 . Архивировано из оригинала 9 октября 2023 г. Проверено 9 октября 2023 г.