Паллий (нейроанатомия)
| Плащ | |
|---|---|
 | |
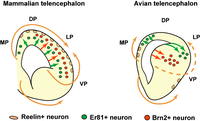 Схематическая иллюстрация различий в спецификации нейронов и моделях миграции между млекопитающих и птиц. мантией | |
| Подробности | |
| Часть | Теленцефалон |
| Идентификаторы | |
| латинский | мантия или кора головного мозга |
| НейроЛекс ID | бирнлекс_1494 |
| ТА98 | A14.1.09.003 |
| ТА2 | 5527 |
| ТО | (нейроанатомия)_by_E5.14.3.4.3.1.30 E5.14.3.4.3.1.30 |
| Анатомические термины нейроанатомии | |
В нейроанатомии паллий ( мн. паллия или паллиум ) относится к слоям серого и белого вещества, которые покрывают верхнюю поверхность головного мозга у позвоночных. Непаллиальная часть конечного мозга образует субпаллий. У базальных позвоночных паллий представляет собой относительно простую трехслойную структуру, включающую 3–4 гистогенетически различных домена, а также обонятельную луковицу .
Раньше считалось, что паллий равен коре, а субпаллий — телэнцефальным ядрам, но оказалось, согласно сравнительным данным, предоставленным молекулярными маркерами , что паллий развивает как корковые структуры (аллокортекс и изокортекс), так и паллиальные ядра ( клаустроамигдалоидный комплекс ), тогда как в субпаллиуме развиваются полосатое , паллидное, диагонально-безымянное и преоптическое ядра, а также кортикоидная структура обонятельного бугорка. [1]
У млекопитающих корковая часть мантия демонстрирует определенный эволюционный рост сложности, образуя кору головного мозга , большая часть которой состоит из постепенно расширяющейся шестислойной части изокортекса с более простыми трехслойными корковыми областями аллокортекса по краям. Аллокортекс подразделяется на аллокортекс гиппокампа медиально и обонятельный аллокортекс латерально (включая рострально обонятельную луковицу и передние обонятельные области).
Структура
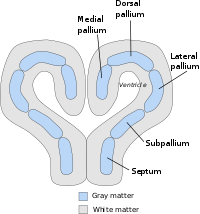
[ редактировать ]Общее расположение или план тела мантия уже четко определен у животных с относительно простым мозгом, в том числе у миног, акул и земноводных. У костистых рыб, рептилий, птиц и млекопитающих паллиальная архитектура сильно модифицирована (резко дивергентна у рыб), с дифференциальным ростом и специализацией различных секторов сохранившегося паллиального бауплана. У всех позвоночных мозг теленцефалический передний мозг состоит из двух полушарий, соединенных по средней линии областью, называемой перегородкой. Перегородка непрерывна с преоптической областью в плоскости, определяемой передней спайкой; он в основном субпаллиальный, но также содержит небольшую паллиальную часть, где образуется спайка гиппокампа, примыкающая к медиальному паллиуму. Теленцефальная часть ростральной хориоидальной тела (крышная пластинка, продолжающаяся каудально с диэнцефальной частью) прикрепляется к задней части этой спайки в том месте, где у млекопитающих виден субфорникальный околожелудочковый орган, и простирается латерально над межжелудочковым отверстием в крыловидную медиальную часть. телэнцефальная область, так называемая хориоидальная щель. Здесь хориоидальная ткань прикрепляется к бахромкам гиппокампа (также известным как кортикальная область рубца), граничащая в продольном направлении с медиальным паллием. На своих ростральном и каудальном концах медиальный паллий контактирует с вентральным паллием, который образует паллиальную часть, которая контактирует с субпаллием через субпаллиальную границу паллио, наблюдаемую на латеральной стенке конечного мозга. Внутри кольца, образованного таким образом медиальной и вентральной мантией, находится своего рода островок, содержащий дорсальную и латеральную мантии. В более старой литературе паллий подразделялся только на три зоны: медиальный паллий, дорсальный (или дорсолатеральный) паллий и латеральный паллий. Старый боковой мантий включал в себя современные боковую и вентральную части мантия. Медиальный паллий является прародителем гиппокампа млекопитающих и, как полагают, участвует в пространственном когнитивном картировании и формировании памяти у широкого круга видов. Латеральный и вентральный паллий является предшественником грушевидной коры млекопитающих и выполняет обонятельную функцию у всех видов, у которых он был изучен. Эволюционную диверсификацию и специализацию функций дорсального мантии объяснить труднее. Широко распространено мнение, что он является прародителем основной части коры головного мозга млекопитающих, хотя некоторые анатомы считают, что доказательства этого еще не убедительны. [2] У млекопитающих и птиц дорсальный паллиум увеличился в размерах и стал преобладающей областью мозга для сенсорной обработки и конечным местом сенсорного сознания, предполагаемая причина этого связана с ночным и роющим образом жизни предков млекопитающих, а также древесным и летучим образом жизни. предков птиц. [3]
Важно отметить, что латеральная и вентральная части мантии также производят глубоко в соответствующие сектора субпиальной обонятельной коры наборы паллиальных ядер, нейроны, входящие в клауструм, рострально, и в паллиальную миндалевидное тело, каудально. Понятие гипопаллия относится к этому гистогенетически единому комплексу обонятельной (грушевидной) коры и глубоких паллиальных ядер. У рептилий и птиц гипопаллий дифференциально увеличивается (наибольший у крокодилов и птиц, обонятельная кора которых тем не менее редуцируется), тогда как у млекопитающих он редуцируется до клаустроамигдалоидного комплекса и относительно увеличенной обонятельной (препириформной и грушевидной) коры.
Паллиальная миндалина содержит в основном так называемую базолатеральную миндалину, охватывающую латеральные, базолатеральные (базальные) и базомедиальные (добавочные базальные) ядра, а также переднюю, миндалино-грушевидную и заднелатеральную кортикоидные области на ее поверхности. Медиальный паллий также может вносить вклад в паллиальную миндалевидное тело, образуя миндалино-гиппокампальное ядро и заднемедиальную кортикоидную область. Было высказано предположение, что нейроны, образующие ядро латерального обонятельного тракта, происходят из дорсального мантии и мигрируют по касательной в его конечное положение каудальнее обонятельного бугорка. Вентральнее мантия в основном плане переднего мозга позвоночных (хотя и представляет собой топологически ростральное поле на картах судеб нервной пластинки) находится еще одна область телэнцефального серого вещества, известная как субпаллий , которая является предшественником области базальных ганглиев , набора структур. которые играют решающую роль в исполнительном контроле поведения. Субпаллиальная область имеет отчетливые полосатую, паллидную, диагональную и преоптическую субобласти, которые наклонно вытянуты между средней линией перегородки и миндалевидным телом на заднем полюсе конечного мозга. По крайней мере, стриатум, паллидум и диагональные домены простираются в миндалевидное тело, представляя там субпаллиальную миндалину, образующую ее центральное и медиальное ядра, а также миндалевидный конец комплекса стриа терминальных ядер ложа.
Таким образом, миндалевидное тело включает в себя гетерогенную группу субпаллиальных ядер и гипопаллиальных обонятельных и миндалино-гиппокампальных корково-ядерных клеточных масс, которые в целом активно участвуют в эмоциях и мотивации. Паллиальные части образуют аналитический или перцептивный конец этого комплекса, тогда как субпаллиальные части представляют собой соответствующий выходной или эфферентный функциональный полюс.Обонятельная луковица представляет собой своеобразный паллиальный отросток (возможно, индуцированный афферентными к нему первичными обонятельными волокнами, идущими от сенсорных нейронов, развитых в обонятельной плакоде), проекционные нейроны которого (митральные и тафтинговые нейроны) имеют паллиальное происхождение и соответственно являются возбуждающими. Напротив, поверхностные перигломерулярные нейроны, различные промежуточные нейроны и глубокие гранулярные клетки имеют субпаллиальное происхождение и мигрируют тангенциально из стриарной части субпаллия (вероятно, из дорсального субсектора этого домена) через так называемый ростральный мигрирующий поток. в обонятельную луковицу. Все эти чрезвычайно многочисленные субпаллиальные клетки являются тормозными. Таким образом, обонятельная луковица образована меньшинством автохтонных паллиальных нейронов и большинством иммигрировавших тормозных субпаллиальных клеток (тем не менее, она классифицируется как часть вентрального паллиума). У основания основной имеется также видоизмененная добавочная обонятельная луковица, которая связана именно с входящими афферентами от органа Якобсона, расположенного в перегородке носа. Дополнительный обонятельный путь максимально развит у некоторых рептилий (например, у змей) и утрачен у птиц.
Эволюция
[ редактировать ]Эволюция дорсального мантия еще до конца не изучена. Некоторые авторы считают, что он в значительной степени способствует развитию аллокортикальных и парагиппокампальных мезокортикальных (переходных) областей гиппокампа млекопитающих. Другие постулируют, что он непосредственно трансформируется в шестислойный изокортекс (неокортекс), характерный для млекопитающих, а третьи полагают, что медиальная и латеральная части дорсального мантия вносят вклад (возможно, с некоторым вкладом латерального мантия) в альтернативные аллокортикальные и изокортикальные судьбы. [4] [5]
У людей
[ редактировать ]Человеческий паллиум ( плащ латинский из-за обширного поверхностного расширения ) покрывает большую часть конечного мозга изокортекса. Теленцефальный паллий классически описывался как состоящий из трех частей: архипаллий , палеопаллий и неопаллий , но эти концепции в настоящее время считаются устаревшими, поскольку были заменены концепцией медиального паллия, дорсального паллия, латерального паллия и вентрального паллия, упомянутых выше в разделе паллиальный Бауплан. Исторически сложилось так, что в учебниках анатомии утверждалось, что паллий эквивалентен коре головного мозга, а субпаллий — ядрам телэнцефала. Однако исследования с использованием молекулярных маркеров показывают, что в паллиуме развиваются как корковые структуры (аллокортекс и изокортекс), так и мантийные ядра (клаустроамигдалоидный комплекс), тогда как в субпаллиуме развиваются полосатое, паллидное, диагонально-безымянное и преоптическое ядра, а также кортикоидная структура обонятельного отдела. туберкулум.
У амфибий и других анамниот
[ редактировать ]У амфибий в конечном мозге отчетливо видны медиальная, дорсальная, латеральная и вентральная части мантия, а также полосатая, паллидная, диагональная и преоптическая части базальных ядер . Однако мантийные части не имеют видимой слоистости. У них уже есть смесь глутаматергических (возбуждающих) и ГАМКергических (тормозных) нейронов, тогда как субпаллий в основном заселен тормозными нейронами. Эта структура очень похожа на структуру, обычно встречающуюся у анамниот, хотя у хрящевых рыб действительно наблюдается послойное расположение паллиальных нейронов.
У рептилий и птиц
[ редактировать ]У рептилий развилась отчетливая трехслойная структура медиальной и дорсальной частей мантии - морфологическая схема, называемая концепцией аллокортекса. Напротив, у рептилий латеральный и вентральный секторы мантия имеют гипопаллиальное строение (поверхностная обонятельная кора, прикрывающая глубокие паллиальные ядра). Гипопаллиальная область также известна как дорсальный желудочковый гребень и имеет переднюю и заднюю (амигдалоидную) области. [4]
Птицы по существу демонстрируют значительно повышенную клеточность, сохраняя морфологическую схему рептилий, что приводит к кажущемуся исчезновению наслоений в медиальном и дорсальном паллиальном секторах. Обонятельная кора сильно редуцирована, тогда как ядра гипопаллиального или дорсального гребня желудочков значительно увеличиваются в размерах и относительной дифференцировке. От передней к задней части части называются гиперпаллием , мезопаллием , нидопаллием и аркопаллием . [5]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Фишер, Робин; Се, Юань-Юнь (4 октября 2010 г.). «Дефекты роста дорсального мантия после генетически направленной абляции основных препластинных нейронов и нейробластов: морфометрический анализ» . АСН Нейро . 2 (5): AN20100022. дои : 10.1042/AN20100022 . ПМК 2949088 . ПМИД 20957077 .
- ^ Ханс Дж. тен Донкелаар; Мартин Ламменс; Акира Хори (7 сентября 2006 г.). Клиническая нейроэмбриология: развитие и нарушения развития центральной нервной системы человека . Спрингер. п. 372. ИСБН 978-3-540-34659-3 .
- ^ Фейнберг, Т.Э.; Маллатт, Дж. М. (2016). Древние истоки сознания: как мозг создавал опыт . Кембридж, Массачусетс: MIT Press. стр. 118, 122–125. ISBN 9780262034333 .
- ^ Перейти обратно: а б Батлер, Энн Б.; Райнер, Антон; Картен, Харви Дж. (апрель 2011 г.). «Эволюция мантии амниота и происхождение неокортекса млекопитающих» . Анналы Нью-Йоркской академии наук . 1225 (1): 14–27. дои : 10.1111/j.1749-6632.2011.06006.x . ПМК 3384708 . ПМИД 21534989 .
- ^ Перейти обратно: а б Джарвис, Эрик Дж. (2009). «Эволюция паллия у птиц и рептилий». Энциклопедия нейробиологии . Берлин: Шпрингер. ISBN 978-3-540-29678-2 .