антиоксидант

Антиоксиданты — это соединения , которые ингибируют окисление (обычно происходящее в виде автоокисления ), химическую реакцию , которая может привести к образованию свободных радикалов . Автоокисление приводит к распаду органических соединений, в том числе живого. Антиоксиданты часто добавляют в промышленные продукты, такие как полимеры , топливо и смазочные материалы , чтобы продлить срок их использования. [1] Продукты питания также обрабатываются антиоксидантами, чтобы предотвратить порчу , в частности прогоркание масел , и жиров . В клетках антиоксиданты, такие как глутатион , микотиол или бациллитиол , а также ферментные системы, такие как супероксиддисмутаза , могут предотвратить повреждение от окислительного стресса . [2]
Известными пищевыми антиоксидантами являются витамины А , С и Е , но термин «антиоксидант» также применяется к многочисленным другим пищевым соединениям, которые обладают антиоксидантными свойствами только in vitro , при этом мало доказательств наличия антиоксидантных свойств in vivo . [3] пищевые добавки, продаваемые как антиоксиданты, не поддерживают здоровье и не предотвращают заболевания у людей. Доказано, что [3] [4]
История
[ редактировать ]В рамках адаптации к морской жизни наземные растения начали вырабатывать неморские антиоксиданты, такие как аскорбиновая кислота ( витамин С ), полифенолы и токоферолы . Эволюция покрытосеменных растений между 50 и 200 миллионами лет назад привела к развитию многих антиоксидантных пигментов – особенно в юрский период – в качестве химической защиты от активных форм кислорода , которые являются побочными продуктами фотосинтеза . [5] Первоначально термин «антиоксидант» конкретно относился к химическому веществу, которое предотвращало потребление кислорода. В конце 19-го и начале 20-го веков обширные исследования были сосредоточены на использовании антиоксидантов в важных промышленных процессах, таких как предотвращение коррозии металлов , вулканизация резины и полимеризация топлива при загрязнении двигателей внутреннего сгорания . [6]
Ранние исследования роли антиоксидантов в биологии были сосредоточены на их использовании для предотвращения окисления ненасыщенных жиров , которое является причиной прогоркания . [7] Антиоксидантную активность можно было бы измерить, просто поместив жир в закрытый контейнер с кислородом и измерив скорость потребления кислорода. Однако именно идентификация витаминов С и Е как антиоксидантов произвела революцию в этой области и привела к осознанию важности антиоксидантов в биохимии живых организмов . [8] [9] Возможные механизмы действия антиоксидантов были впервые изучены, когда было признано, что вещество с антиоксидантной активностью, вероятно, само по себе легко окисляется. [10] Исследования того, как витамин Е предотвращает процесс перекисного окисления липидов, привели к идентификации антиоксидантов как восстановителей, которые предотвращают окислительные реакции, часто удаляя активные формы кислорода , прежде чем они смогут повредить клетки. [11]
Использование в технологии
[ редактировать ]Пищевые консерванты
[ редактировать ]Антиоксиданты используются в качестве пищевых добавок , чтобы защитить продукты от порчи . Воздействие кислорода и солнечного света являются двумя основными факторами окисления продуктов питания, поэтому продукты консервируют, храня их в темноте и запечатывая в контейнерах или даже покрывая воском, как в случае с огурцами. Однако, поскольку кислород также важен для дыхания растений , хранение растительного сырья в анаэробных условиях приводит к появлению неприятного вкуса и непривлекательного цвета. [12] Следовательно, упаковка свежих фруктов и овощей содержит ≈8% кислородной атмосферы. Антиоксиданты представляют собой особенно важный класс консервантов, поскольку, в отличие от бактериальной или грибковой порчи, реакции окисления в замороженных или охлажденных продуктах протекают относительно быстро. [13] Эти консерванты включают природные антиоксиданты, такие как аскорбиновая кислота (АА, E300) и токоферолы (E306), а также синтетические антиоксиданты, такие как пропилгаллат (PG, E310), третичный бутилгидрохинон (TBHQ), бутилированный гидроксианизол (BHA, E320) и бутилированный гидроксианизол. гидрокситолуол (ВНТ, Е321). [14] [15]
Ненасыщенные жиры могут быть очень восприимчивы к окислению, вызывая прогоркание . [16] Окисленные липиды часто обесцвечиваются и могут придавать неприятный вкус и аромат. Таким образом, эти продукты редко сохраняются путем сушки; вместо этого их консервируют путем копчения , соления или ферментации . Даже менее жирные продукты, такие как фрукты, перед сушкой на воздухе опрыскивают сернистыми антиоксидантами. Металлы катализируют окисление. Некоторые жирные продукты, такие как оливковое масло, частично защищены от окисления благодаря натуральному содержанию антиоксидантов. Жирная пища чувствительна к фотоокислению. [17] который образует гидроперекиси путем окисления ненасыщенных жирных кислот и эфиров . [18] Воздействие ультрафиолетового (УФ) излучения может вызвать прямое фотоокисление и разложение пероксидов и молекул карбонила . Эти молекулы подвергаются свободнорадикальным цепным реакциям, но антиоксиданты ингибируют их, предотвращая процессы окисления. [18]
Косметические консерванты
[ редактировать ]Антиоксидантные стабилизаторы также добавляются в косметические средства на жировой основе, такие как губная помада и увлажняющие средства, чтобы предотвратить прогоркание. [19] Антиоксиданты в косметических продуктах предотвращают окисление активных ингредиентов и содержания липидов. Например, фенольные антиоксиданты, такие как стильбены , флавоноиды и гидроксикоричная кислота , сильно поглощают УФ-излучение из-за присутствия хромофоров . Они уменьшают окислительный стресс от воздействия солнца, поглощая ультрафиолетовый свет. [20]
Промышленное использование
[ редактировать ]
Антиоксиданты могут добавляться в промышленные продукты, такие как стабилизаторы топлива , для предотвращения окисления и полимеризации , и присадки к смазочным материалам которые приводят к образованию остатков загрязнения двигателя. [21]
| Топливная присадка | Компоненты [22] | Приложения [22] |
|---|---|---|
| АО-22 | N,N'-ди-2-бутил-1,4-фенилендиамин | Турбинные масла, трансформаторные масла , гидравлические жидкости , воски и смазки |
| АО-24 | N,N'-ди-2-бутил-1,4-фенилендиамин | Низкотемпературные масла |
| АО-29 | 2,6-ди-трет-бутил-4-метилфенол (ВНТ) | Турбинные масла, трансформаторные масла, гидравлические жидкости, воски, смазки и бензины. |
| АО-30 | 2,4-диметил-6-трет-бутилфенол | Топливо для реактивных двигателей и бензины, включая авиационные бензины |
| АО-31 | 2,4-диметил-6-трет-бутилфенол | Топливо для реактивных двигателей и бензины, в том числе авиационные бензины |
| АО-32 | 2,4-диметил-6-трет-бутилфенол и 2,6-ди-трет-бутил-4-метилфенол | Топливо для реактивных двигателей и бензины, в том числе авиационные бензины |
| АО-37 | 2,6-ди-трет-бутилфенол | Реактивное топливо и бензины, широко одобренные в качестве авиационного топлива. |
Антиоксидантные полимерные стабилизаторы широко используются для предотвращения деградации полимеров , таких как каучуки, пластмассы и клеи , что приводит к потере прочности и гибкости этих материалов. [23] Полимеры, содержащие двойные связи в своих основных цепях, такие как натуральный каучук и полибутадиен , особенно подвержены окислению и озонолизу . Их можно защитить антиозонантами . Окисление может быть ускорено УФ-излучением естественного солнечного света, вызывая фотоокисление . различные специализированные светостабилизаторы, такие как HALS Чтобы предотвратить это, в пластик можно добавлять . Ниже представлен обзор некоторых из наиболее часто применяемых антиоксидантов для полимерных материалов:
(Затрудненные) фенольные антиоксиданты : действуют путем удаления свободных радикалов, образующихся в процессе термического окисления, тем самым предотвращая цепные реакции, которые приводят к деградации полимера.
Фосфиты : действуют путем разложения пероксидов на нерадикальные продукты, тем самым предотвращая дальнейшее образование свободных радикалов и способствуя общей окислительной стабильности полимера. Фосфиты часто используются в сочетании с фенольными антиоксидантами для достижения синергетического эффекта.
Тиоэфиры : действуют путем разложения пероксидов на нерадикальные продукты. Тиоэфиры также используются в качестве состабилизаторов с первичными антиоксидантами.
Светостабилизаторы на основе затрудненных аминов (HALS) : HALS действуют путем удаления свободных радикалов, образующихся во время фотоокисления, тем самым защищая полимерный материал от УФ-излучения.
Витамины : природные антиоксиданты, такие как витамин С и витамин Е, используются для конкретных целей.
Смеси . Обычно применяются смеси различных типов антиоксидантов, поскольку они могут служить различным целям.
Опасности для окружающей среды и здоровья
[ редактировать ]Синтетические фенольные антиоксиданты (СПА) [24] и аминные антиоксиданты [25] несут потенциальную опасность для здоровья человека и окружающей среды. SPA часто встречаются в пыли внутри помещений, мелких частицах воздуха, отложениях, сточных водах, речной воде и сточных водах. [26] Они синтезируются из фенольных соединений и включают 2,6-ди-трет-бутил-4-метилфенол (ВНТ) , 2,6-ди-трет-бутил-п-бензохинон (ВНТ-Q), 2,4-ди- трет-бутилфенол (DBP) и 3- трет -бутил-4-гидроксианизол (BHA) . BHT может вызвать гепатотоксичность и повреждение эндокринной системы , а также может увеличить скорость развития опухолей из-за 1,1-диметилгидразина . [27] BHT-Q может вызвать повреждение ДНК и несоответствия [28] в процессе расщепления с образованием супероксидных радикалов. [26] DBP токсичен для морской жизни при длительном воздействии. Фенольные антиоксиданты обладают низкой биоразлагаемостью, но не обладают серьезной токсичностью по отношению к водным организмам в низких концентрациях. Другой тип антиоксиданта, дифениламин (ДФА) , обычно используется в производстве коммерческих, промышленных смазочных материалов и резиновых изделий, а также служит добавкой к автомобильным моторным маслам. [29]
Окислительная проблема в биологии
[ редактировать ]
Подавляющему большинству сложной жизни на Земле необходим кислород для метаболизма , но этот же кислород является высокореактивным элементом , который может повредить живые организмы. [2] [30] Организмы содержат химические вещества и ферменты , которые сводят к минимуму окислительное повреждение, не мешая при этом полезному эффекту кислорода. [31] [32] В целом антиоксидантные системы либо предотвращают образование этих реактивных частиц, либо удаляют их, тем самым сводя к минимуму их ущерб. [30] [31] Активные формы кислорода могут выполнять полезные клеточные функции, такие как передача окислительно-восстановительных сигналов . Таким образом, в идеале антиоксидантные системы не удаляют оксиданты полностью, а поддерживают их в некоторой оптимальной концентрации. [33]
Активные формы кислорода, вырабатываемые в клетках, включают перекись водорода (H 2 O 2 ), гипохлористую кислоту (HClO) и свободные радикалы, такие как гидроксильный радикал (·OH), и супероксид-анион (O 2 − ). [34] Гидроксильный радикал особенно нестабилен и быстро и неспецифически реагирует с большинством биологических молекул. Этот вид образуется из перекиси водорода в окислительно -восстановительных реакциях, катализируемых металлами, таких как реакция Фентона . [35] Эти окислители могут повредить клетки, запуская цепные химические реакции, такие как перекисное окисление липидов , или окисляя ДНК или белки. [31] Повреждение ДНК может вызвать мутации и, возможно, рак , если его не обратить вспять механизмами репарации ДНК . [36] [37] в то время как повреждение белков вызывает ингибирование ферментов, денатурацию и деградацию белков . [38]
Использование кислорода как часть процесса производства метаболической энергии приводит к образованию активных форм кислорода. [39] В этом процессе супероксид-анион образуется как побочный продукт нескольких стадий цепи переноса электронов . [40] Особенно важно восстановление кофермента Q в комплексе III , так как в качестве интермедиата образуется высокореактивный свободный радикал (Q · − ). Этот нестабильный промежуточный продукт может привести к «утечке» электронов, когда электроны перепрыгивают непосредственно на кислород и образуют супероксид-анион, вместо того, чтобы проходить через нормальную серию хорошо контролируемых реакций цепи переноса электронов. [41] Пероксид также образуется в результате окисления восстановленных флавопротеинов , таких как комплекс I. [42] Однако, хотя эти ферменты могут производить окислители, относительная важность цепи переноса электронов для других процессов, в которых генерируется пероксид, неясна. [43] [44] В растениях, водорослях и цианобактериях активные формы кислорода также производятся в процессе фотосинтеза . [45] особенно в условиях высокой интенсивности света . [46] Этот эффект частично компенсируется участием каротиноидов в фотоингибировании , а у водорослей и цианобактерий — большим количеством йодида и селена . [47] который включает в себя реакцию этих антиоксидантов с чрезмерно восстановленными формами фотосинтетических реакционных центров, чтобы предотвратить выработку активных форм кислорода. [48] [49]
Примеры биологически активных антиоксидантных соединений
[ редактировать ]Физиологические антиоксиданты подразделяются на два больших отдела в зависимости от того, растворимы ли они в воде ( гидрофильные ) или в липидах ( липофильные ). В целом водорастворимые антиоксиданты реагируют с окислителями в цитозоле клеток и плазме крови , тогда как жирорастворимые антиоксиданты защищают клеточные мембраны от перекисного окисления липидов . [31] Эти соединения могут синтезироваться в организме или получаться с пищей. [32] Различные антиоксиданты присутствуют в широком диапазоне концентраций в жидкостях и тканях организма, причем некоторые из них, такие как глутатион или убихинон, в основном присутствуют в клетках, тогда как другие, такие как мочевая кислота, распределяются более системно (см. таблицу ниже). Некоторые антиоксиданты обнаружены только у некоторых организмов и могут быть патогенами или факторами вирулентности . [50]
Взаимодействия между этими различными антиоксидантами могут быть синергическими и взаимозависимыми. [51] [52] Таким образом, действие одного антиоксиданта может зависеть от правильного функционирования других членов антиоксидантной системы. [32] Степень защиты, обеспечиваемая любым антиоксидантом, также будет зависеть от его концентрации, его реактивности по отношению к конкретным рассматриваемым активным формам кислорода и статуса антиоксидантов, с которыми он взаимодействует. [32]
Некоторые соединения способствуют антиоксидантной защите, хелатируя переходные металлы и не позволяя им катализировать образование свободных радикалов в клетке. Одной из таких функций является способность связывать железо для железосвязывающих белков , таких как трансферрин и ферритин . [44] Селен и цинк обычно называют антиоксидантными минералами , но эти химические элементы сами по себе не обладают антиоксидантным действием, а скорее необходимы для активности антиоксидантных ферментов, таких как глутатионредуктаза и супероксиддисмутаза . (См. также селен в биологии и цинк в биологии .)
| антиоксидант | Растворимость | Концентрация в сыворотке человека ( мкМ ) | Концентрация в ткани печени ( мкмоль/кг ) |
|---|---|---|---|
| Аскорбиновая кислота ( витамин С ) | Вода | 50–60 [53] | 260 (человек) [54] |
| Глутатион | Вода | 4 [55] | 6400 (человек) [54] |
| Липоевая кислота | Вода | 0.1–0.7 [56] | 4–5 (крыса) [57] |
| Мочевая кислота | Вода | 200–400 [58] | 1600 (человек) [54] |
| Каротины | Липид | β-каротин : 0,5–1 [59] | 5 (человек, общие каротиноиды) [61] |
| α-Токоферол (витамин Е) | Липид | 10–40 [60] | 50 (человек) [54] |
| Убихинол (коэнзим Q) | Липид | 5 [62] | 200 (человек) [63] |
Мочевая кислота
[ редактировать ]Мочевая кислота имеет самую высокую концентрацию среди всех антиоксидантов крови. [58] и обеспечивает более половины общей антиоксидантной способности сыворотки человека. [64] Антиоксидантная активность мочевой кислоты также сложна, учитывая, что она не реагирует с некоторыми окислителями, такими как супероксид , но действует против пероксинитрита . [65] пероксиды и хлорноватистая кислота . [66] Обеспокоенность по поводу повышенного вклада МК в развитие подагры следует рассматривать как один из многих факторов риска. [67] Сам по себе риск развития подагры, связанный с МК, при высоких уровнях (415–530 мкмоль/л) составляет всего 0,5% в год с увеличением до 4,5% в год при уровнях перенасыщения МК (535+ мкмоль/л). [68] Многие из этих вышеупомянутых исследований определили антиоксидантное действие УК в пределах нормального физиологического уровня. [69] [65] а некоторые обнаружили антиоксидантную активность на уровне 285 мкмоль/л. [70]
Витамин С
[ редактировать ]Аскорбиновая кислота или витамин С , окислительно-восстановительный ( окислительно-восстановительный ) катализатор, обнаруженный как в животных, так и в растениях. [71] может уменьшить и тем самым нейтрализовать активные формы кислорода, такие как перекись водорода. [71] [72] Помимо прямого антиоксидантного действия, аскорбиновая кислота также является субстратом для окислительно-восстановительного фермента аскорбатпероксидазы , функции, которая используется для устойчивости растений к стрессу. [73] Аскорбиновая кислота присутствует в больших количествах во всех частях растений и может достигать концентрации 20 миллимолярных в хлоропластах . [74]
Глутатион
[ редактировать ]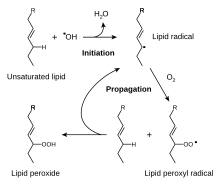
Глутатион обладает антиоксидантными свойствами, поскольку тиоловая группа в его цистеиновой части является восстановителем и может обратимо окисляться и восстанавливаться. В клетках глутатион сохраняется в восстановленной форме с помощью фермента глутатионредуктазы и, в свою очередь, восстанавливает другие метаболиты и ферментные системы, такие как аскорбат в глутатион-аскорбатном цикле , глутатионпероксидазы и глутаредоксины , а также реагируя непосредственно с окислителями. [75] Благодаря своей высокой концентрации и центральной роли в поддержании окислительно-восстановительного состояния клетки глутатион является одним из наиболее важных клеточных антиоксидантов. [76] У некоторых организмов глутатион заменен другими тиолами, например микотиолом у актиномицетов , бациллитиолом у некоторых грамположительных бактерий , [77] [78] или трипанотионом в кинетопластидах . [79] [80]
Витамин Е
[ редактировать ]Витамин Е — это собирательное название набора из восьми родственных токоферолов и токотриенолов , которые представляют собой жирорастворимые витамины с антиоксидантными свойствами. [81] [82] Из них наиболее изучен альфа-токоферол, поскольку он имеет самую высокую биодоступность , при этом организм преимущественно усваивает и метаболизирует эту форму. [83]
Утверждалось, что форма α-токоферола является наиболее важным жирорастворимым антиоксидантом и что она защищает мембраны от окисления, реагируя с липидными радикалами, образующимися в цепной реакции перекисного окисления липидов. [81] [84] Это удаляет промежуточные продукты свободных радикалов и предотвращает продолжение реакции распространения. В результате этой реакции образуются окисленные α-токофероксильные радикалы, которые можно вернуть обратно в активную восстановленную форму за счет восстановления другими антиоксидантами, такими как аскорбат, ретинол или убихинол. [85] Это согласуется с данными, показывающими, что α-токоферол, но не водорастворимые антиоксиданты, эффективно защищает клетки с дефицитом глутатионпероксидазы 4 ( GPX4 ) от гибели клеток. [86] GPx4 — единственный известный фермент, который эффективно восстанавливает гидропероксиды липидов в биологических мембранах.
Однако роль и значение различных форм витамина Е в настоящее время неясны. [87] [88] и даже было высказано предположение, что наиболее важной функцией α-токоферола является сигнальная молекула , причем эта молекула не играет существенной роли в антиоксидантном метаболизме. [89] [90] Функции других форм витамина Е еще менее изучены, хотя γ-токоферол является нуклеофилом , который может реагировать с электрофильными мутагенами. [83] и токотриенолы могут играть важную роль в защите нейронов от повреждений. [91]
Прооксидантная активность
[ редактировать ]Антиоксиданты, являющиеся восстановителями, также могут действовать как прооксиданты. Например, витамин С обладает антиоксидантной активностью, когда восстанавливает окисляющие вещества, такие как перекись водорода; [92] однако он также снижает содержание ионов металлов, таких как железо и медь. [93] которые генерируют свободные радикалы посредством реакции Фентона . [35] [94] Хотя аскорбиновая кислота является эффективным антиоксидантом, она также может окислительно изменять вкус и цвет пищи. При наличии переходных металлов низкие концентрации аскорбиновой кислоты могут действовать как поглотитель радикалов в реакции Фентона. [93]
2 Фе 3+ + Аскорбат → 2 Fe 2+ + Дегидроаскорбат
2 Фе 2+ + 2 H 2 O 2 → 2 Fe 3+ + 2 ОН · + 2 ОН −
Относительная важность антиоксидантной и прооксидантной активности антиоксидантов является областью текущих исследований, но витамин С, который оказывает свое действие как витамин путем окисления полипептидов, по-видимому, оказывает в организме человека преимущественно антиоксидантное действие. [94]
Ферментные системы
[ редактировать ]![{\displaystyle {\ce {{\underset {Кислород}{O2}}->{\underset {Супероксид}{*O2^{-}}}->[{\ce {Супероксид \на вершине дисмутазы}}]{\ underset {Водород \поверх перекиси}{H2O2}}->[{\ce {Пероксидазы \поверх каталазы}}]{\underset {Water}{H2O}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8feecd26377be56b431d182ea9a398ab6b5d3b7f)
Как и в случае с химическими антиоксидантами, клетки защищены от окислительного стресса взаимодействующей сетью антиоксидантных ферментов. [30] [31] Здесь супероксид, выделяемый в результате таких процессов, как окислительное фосфорилирование, сначала преобразуется в перекись водорода, а затем восстанавливается с образованием воды. Этот путь детоксикации является результатом работы множества ферментов: первый этап катализирует супероксиддисмутаза, а затем каталазы и различные пероксидазы, удаляющие перекись водорода. Как и в случае с антиоксидантными метаболитами, вклад этих ферментов в антиоксидантную защиту может быть трудно отделить друг от друга, но создание трансгенных мышей, лишенных хотя бы одного антиоксидантного фермента, может быть информативным. [95]
Супероксиддисмутаза, каталаза и пероксиредоксины
[ редактировать ]Супероксиддисмутазы (СОД) представляют собой класс близкородственных ферментов, которые катализируют расщепление супероксид-аниона на кислород и перекись водорода. [96] [97] Ферменты СОД присутствуют почти во всех аэробных клетках и во внеклеточных жидкостях. [98] Ферменты супероксиддисмутазы содержат кофакторы ионов металлов, которыми в зависимости от изофермента могут быть медь, цинк, марганец или железо. У человека СОД меди/цинка присутствует в цитозоле , а СОД марганца присутствует в митохондриях . [97] также существует третья форма СОД Во внеклеточных жидкостях , которая содержит в своих активных центрах медь и цинк. [99] Митохондриальный изофермент, по-видимому, является наиболее биологически важным из этих трех, поскольку мыши, лишенные этого фермента, умирают вскоре после рождения. [100] Напротив, мыши, лишенные СОД меди/цинка (Sod1), жизнеспособны, но имеют многочисленные патологии и уменьшенную продолжительность жизни (см. статью о супероксиде ), в то время как мыши без внеклеточной СОД имеют минимальные дефекты (чувствительны к гипероксии ). [95] [101] У растений изоферменты СОД присутствуют в цитозоле и митохондриях, при этом СОД железа обнаруживается в хлоропластах и отсутствует у позвоночных и дрожжей . [102]
Каталазы — это ферменты, которые катализируют превращение перекиси водорода в воду и кислород с использованием кофактора железа или марганца. [103] [104] Этот белок локализован в пероксисомах большинства эукариотических клеток. [105] Каталаза является необычным ферментом, поскольку, хотя перекись водорода является ее единственным субстратом, она действует по механизму пинг-понга . Здесь его кофактор окисляется одной молекулой перекиси водорода, а затем регенерируется путем переноса связанного кислорода на вторую молекулу субстрата. [106] Несмотря на очевидную важность удаления перекиси водорода, люди с генетическим дефицитом каталазы — « акаталаземия » — или мыши, генетически модифицированные для полного отсутствия каталазы, испытывают мало побочных эффектов. [107] [108]

Пероксиредоксины — пероксидазы, катализирующие восстановление перекиси водорода, органических гидроперекисей , а также пероксинитрита . [110] Их разделяют на три класса: типичные 2-цистеиновые пероксиредоксины; атипичные 2-цистеиновые пероксиредоксины; и 1-цистеинпероксиредоксины. [111] Эти ферменты имеют один и тот же основной каталитический механизм, при котором окислительно-восстановительный цистеин (пероксидатический цистеин) в активном центре окисляется до сульфеновой кислоты пероксидным субстратом. [112] Чрезмерное окисление этого остатка цистеина в пероксиредоксинах инактивирует эти ферменты, но это можно обратить вспять действием сульфиредоксина . [113] Пероксиредоксины, по-видимому, играют важную роль в антиоксидантном метаболизме, поскольку мыши, лишенные пероксиредоксина 1 или 2, сокращают продолжительность жизни и развивают гемолитическую анемию , в то время как растения используют пероксиредоксины для удаления перекиси водорода, образующейся в хлоропластах. [114] [115] [116]
Тиоредоксиновая и глутатионовая системы
[ редактировать ]Тиоредоксиновая система содержит белок тиоредоксин массой 12 кДа и его компаньон тиоредоксинредуктазу . [117] Белки, родственные тиоредоксину, присутствуют во всех секвенированных организмах. Растения, такие как Arabidopsis thaliana , имеют особенно большое разнообразие изоформ. [118] Активный центр тиоредоксина состоит из двух соседних цистеинов, как часть высококонсервативного мотива CXXC , который может циклически переключаться между активной формой дитиола (восстановленной) и окисленной дисульфидной формой. В активном состоянии тиоредоксин действует как эффективный восстановитель, удаляя активные формы кислорода и поддерживая другие белки в восстановленном состоянии. [119] После окисления активный тиоредоксин регенерируется под действием тиоредоксинредуктазы с использованием НАДФН в качестве донора электронов . [120]
Глутатионовая , система включает глутатион глутатионредуктазу , глутатионпероксидазы и глутатионS - трансферазы . [76] Эта система встречается у животных, растений и микроорганизмов. [76] [121] Глутатионпероксидаза — фермент, содержащий четыре селен - кофактора , катализирующий расщепление перекиси водорода и органических гидроперекисей. глутатионпероксидазы . существует по крайней мере четыре различных изофермента У животных [122] Глутатионпероксидаза 1 является наиболее распространенной и является очень эффективным поглотителем перекиси водорода, тогда как глутатионпероксидаза 4 наиболее активна в отношении гидропероксидов липидов. Удивительно, но глутатионпероксидаза 1 необязательна, поскольку мыши, лишенные этого фермента, имеют нормальную продолжительность жизни. [123] но они сверхчувствительны к индуцированному окислительному стрессу. [124] Кроме того, глутатион S -трансферазы проявляют высокую активность в отношении пероксидов липидов. [125] Эти ферменты находятся на особенно высоком уровне в печени и также участвуют в детоксикационном метаболизме. [126]
Исследования в области здравоохранения
[ редактировать ]Отношение к диете
[ редактировать ]Диетические антиоксидантные витамины А, С и Е необходимы и необходимы в определенных ежедневных количествах для предотвращения заболеваний. [3] [127] [128] Полифенолы , которые обладают антиоксидантными свойствами in vitro благодаря своим свободным гидроксильным группам . [129] интенсивно метаболизируются катехол-О-метилтрансферазой , которая метилирует свободные гидроксильные группы и тем самым предотвращает их действие в качестве антиоксидантов in vivo. [130] [131]
Взаимодействия
[ редактировать ]Обычные фармацевтические препараты (и добавки) с антиоксидантными свойствами могут влиять на эффективность некоторых противораковых препаратов и лучевой терапии . [132] Фармацевтические препараты и добавки, обладающие антиоксидантными свойствами, подавляют образование свободных радикалов, подавляя процессы окисления. Лучевая терапия вызывает окислительный стресс, который повреждает важные компоненты раковых клеток, такие как белки, нуклеиновые кислоты и липиды, составляющие клеточные мембраны. [133]
Побочные эффекты
[ редактировать ]
Относительно сильные восстанавливающие кислоты могут оказывать антипитательное действие, связываясь с пищевыми минералами, такими как железо и цинк, в желудочно-кишечном тракте и предотвращая их всасывание. [134] Примерами являются щавелевая кислота , дубильные вещества и фитиновая кислота , высокое содержание которых содержится в растительных диетах. [135] Дефицит кальция и железа не является редкостью в рационе питания в развивающихся странах , где едят меньше мяса и высок уровень потребления фитиновой кислоты из бобов и пресного цельнозернового хлеба. Однако проращивание, замачивание или микробная ферментация — все это методы домашнего хозяйства, позволяющие снизить содержание фитатов и полифенолов в нерафинированных злаках. Сообщалось об увеличении абсорбции Fe, Zn и Ca у взрослых, которых кормили дефитинизированными зерновыми, по сравнению с зерновыми, содержащими их нативный фитат. [136]
| Продукты питания | Уменьшение присутствия кислоты |
|---|---|
| Какао-бобы и шоколад, шпинат , репа и ревень [137] | Щавелевая кислота |
| Цельнозерновые , кукуруза, бобовые [138] | Фитиновая кислота |
| Чай, фасоль , капуста [137] [139] | Танины |
Высокие дозы некоторых антиоксидантов могут иметь вредные долгосрочные последствия. Исследование бета-каротина и ретинола эффективности (CARET) среди пациентов с раком легких показало, что у курильщиков, получавших добавки, содержащие бета-каротин и витамин А, наблюдался повышенный уровень заболеваемости раком легких. [140] Последующие исследования подтвердили эти побочные эффекты. [141] Эти вредные эффекты могут также наблюдаться у некурящих, поскольку один метаанализ , включающий данные примерно 230 000 пациентов, показал, что прием добавок β-каротина, витамина А или витамина Е связан с увеличением смертности, но не выявил значимого эффекта от витамина С. [142] Никакого риска для здоровья не наблюдалось, когда все рандомизированные контролируемые исследования рассматривались вместе, но увеличение смертности было обнаружено, когда отдельно рассматривались только исследования высокого качества и с низким уровнем систематической ошибки. [143] Поскольку большинство этих исследований с низкой предвзятостью касались либо пожилых людей , либо людей с заболеваниями, эти результаты могут быть неприменимы к населению в целом. [144] Этот метаанализ позже был повторен и расширен теми же авторами, подтвердив предыдущие результаты. [143] Эти две публикации согласуются с некоторыми предыдущими метаанализами, которые также предположили, что добавление витамина Е увеличивает смертность. [145] и что антиоксидантные добавки повышают риск рака толстой кишки . [146] Бета-каротин также может способствовать развитию рака легких . [146] [147] В целом, большое количество клинических испытаний, проведенных с антиоксидантными добавками, позволяет предположить, что либо эти продукты не оказывают никакого влияния на здоровье, либо вызывают небольшое увеличение смертности среди пожилых или уязвимых групп населения. [127] [148] [142]
Физические упражнения и боль в мышцах
[ редактировать ]Обзор 2017 года показал, что прием антиоксидантных пищевых добавок до или после тренировки вряд ли приведет к заметному уменьшению болезненности мышц после тренировки. [149]
Уровни в еде
[ редактировать ]
Витамины-антиоксиданты содержатся в овощах, фруктах, яйцах, бобовых и орехах. Витамины А, С и Е могут разрушаться при длительном хранении или длительной кулинарной обработке. [150] Последствия приготовления пищи и обработки пищевых продуктов сложны, поскольку эти процессы также могут увеличить биодоступность антиоксидантов, таких как некоторые каротиноиды в овощах. [151] Обработанные продукты содержат меньше витаминов-антиоксидантов, чем свежие и сырые продукты, поскольку при приготовлении они подвергаются воздействию тепла и кислорода. [152]
| Витамины-антиоксиданты | Продукты, содержащие высокий уровень витаминов-антиоксидантов [139] [153] [154] |
|---|---|
| Витамин С (аскорбиновая кислота) | Свежие или замороженные фрукты и овощи |
| Витамин Е (токоферолы, токотриенолы) | Растительные масла , орехи и семена |
| Каротиноиды ( каротины как провитамин А ) | Фрукты, овощи и яйца |
Другие антиоксиданты не поступают с пищей, а вырабатываются в организме. Например, убихинол (коэнзим Q) плохо всасывается в кишечнике и вырабатывается мевалонатным путем . [63] Другой пример — глутатион , который состоит из аминокислот. Поскольку любой глутатион в кишечнике расщепляется до свободного цистеина, глицина и глутаминовой кислоты перед всасыванием, даже большой пероральный прием мало влияет на концентрацию глутатиона в организме. [155] [156] Хотя большое количество серосодержащих аминокислот, таких как ацетилцистеин, может повысить уровень глутатиона, [157] Нет никаких доказательств того, что употребление высоких доз этих предшественников глутатиона полезно для здоровых взрослых. [158]
Измерение и аннулирование ORAC
[ редактировать ]Измерение содержания полифенолов и каротиноидов в пище — непростой процесс, поскольку антиоксиданты в совокупности представляют собой разнообразную группу соединений с различной реакционной способностью по отношению к различным активным формам кислорода. В пищевых исследованиях in vitro способность поглощать кислородные радикалы (ORAC) когда-то была отраслевым стандартом для оценки антиоксидантной силы цельных продуктов, соков и пищевых добавок, главным образом, на основе присутствия полифенолов . [159] [160] Более ранние измерения и рейтинги Министерства сельского хозяйства США были отозваны в 2012 году как биологически не имеющие отношения к здоровью человека, ссылаясь на отсутствие физиологических доказательств того, что полифенолы обладают антиоксидантными свойствами in vivo . [161] Следовательно, метод ORAC, полученный только в результате экспериментов in vitro больше не считается актуальным для человеческого питания или биологии . , с 2010 года [161]
Альтернативные измерения содержания антиоксидантов в пищевых продуктах in vitro, также основанные на присутствии полифенолов, включают реагент Фолина-Чиокальтеу и анализ эквивалентной антиоксидантной способности Тролокса . [162]
Ссылки
[ редактировать ]- ^ Клемчук, Петр П. (2000). «Антиоксиданты». Энциклопедия промышленной химии Ульмана . дои : 10.1002/14356007.a03_091 . ISBN 3527306730 .
- ^ Перейти обратно: а б Хелберг, Джулиан; Пратт, Дерек А. (2021). «Аутоокисление против антиоксидантов – борьба навсегда». Обзоры химического общества . 50 (13): 7343–7358. дои : 10.1039/D1CS00265A . ПМИД 34037013 . S2CID 235200305 .
- ^ Перейти обратно: а б с «Антиоксиданты: подробно» . Национальный центр дополнительного и интегративного здравоохранения, Национальные институты здравоохранения США. 1 ноября 2013 года . Проверено 17 марта 2023 г.
- ^ Фанг, Юн-Чжун; Ян, Шэн; Ву, Гояо (2002). «Свободные радикалы, антиоксиданты и питание». Питание . 18 (10): 872–879. дои : 10.1016/s0899-9007(02)00916-4 . ПМИД 12361782 .
- ^ Бензи ИФ (сентябрь 2003 г.). «Эволюция пищевых антиоксидантов». Сравнительная биохимия и физиология А. 136 (1): 113–26. дои : 10.1016/S1095-6433(02)00368-9 . hdl : 10397/34754 . ПМИД 14527634 .
- ^ Мэттилл Х.А. (1947). «Антиоксиданты». Ежегодный обзор биохимии . 16 : 177–92. дои : 10.1146/annurev.bi.16.070147.001141 . ПМИД 20259061 .
- ^ Немецкий JB (1999). «Пищевая промышленность и окисление липидов». Влияние переработки на безопасность пищевых продуктов . Достижения экспериментальной медицины и биологии. Том. 459. стр. 23–50. дои : 10.1007/978-1-4615-4853-9_3 . ISBN 978-0-306-46051-7 . ПМИД 10335367 .
- ^ Джейкоб Р.А. (1996). «Введение: три эпохи открытия витамина С». Три эпохи открытия витамина С. Субклеточная биохимия. Том. 25. стр. 1–16. дои : 10.1007/978-1-4613-0325-1_1 . ISBN 978-1-4613-7998-0 . ПМИД 8821966 .
- ^ Найт Дж.А. (1998). «Свободные радикалы: их история и современный статус в вопросах старения и болезней». Анналы клинической и лабораторной науки . 28 (6): 331–46. ПМИД 9846200 .
- ^ Мурё С., Дюфрэс С. (1922). «Об автоокислении: Антикислороны». Отчеты заседаний и мемуары Общества биологии (на французском языке). 86 : 321–322.
- ^ Вольф Г. (март 2005 г.). «Открытие антиоксидантной функции витамина Е: вклад Генри А. Мэттилла» . Журнал питания . 135 (3): 363–6. дои : 10.1093/jn/135.3.363 . ПМИД 15735064 .
- ^ Кадер А.А., Загори Д., Кербель Э.Л. (1989). «Упаковка фруктов и овощей в модифицированной атмосфере». Критические обзоры в области пищевой науки и питания . 28 (1): 1–30. дои : 10.1080/10408398909527490 . ПМИД 2647417 .
- ^ Заллен Э.М., Хичкок М.Дж., Герц Г.Е. (декабрь 1975 г.). «Охлаждаемые пищевые системы. Влияние хранения в охлажденном виде на качество говяжьих буханок». Журнал Американской диетической ассоциации . 67 (6): 552–7. дои : 10.1016/S0002-8223(21)14836-9 . ПМИД 1184900 .
- ^ Айверсон Ф (июнь 1995 г.). «Фенольные антиоксиданты: исследования Отделения здравоохранения по бутилированному гидроксианизолу». Письма о раке . 93 (1): 49–54. дои : 10.1016/0304-3835(95)03787-W . ПМИД 7600543 .
- ^ «Индекс номера E» . Британский кулинарный гид. Архивировано из оригинала 4 марта 2007 года . Проверено 5 марта 2007 г.
- ^ Робардс К., Керр А.Ф., Патсалидес Э. (февраль 1988 г.). «Прогорклость и ее измерение в пищевых маслах и закусках. Обзор». Аналитик . 113 (2): 213–24. Бибкод : 1988Ана...113..213R . дои : 10.1039/an9881300213 . ПМИД 3288002 .
- ^ Дель Карло М., Саккетти Г., Ди Маттиа К., Компаньоне Д., Мастрокола Д., Либераторе Л., Чичелли А. (июнь 2004 г.). «Вклад фенольной фракции в антиоксидантную активность и окислительную стабильность оливкового масла». Журнал сельскохозяйственной и пищевой химии . 52 (13): 4072–9. дои : 10.1021/jf049806z . ПМИД 15212450 .
- ^ Перейти обратно: а б Франкель, Эдвин Н. (1 января 2012 г.), Франкель, Эдвин Н. (ред.), «Глава 3 - Фотоокисление ненасыщенных жиров» , Окисление липидов (второе издание) , Серия библиотек липидов Oily Press, Woodhead Publishing, стр. 51 –66, ISBN 978-0-9531949-8-8 , получено 15 апреля 2023 г.
- ^ «Окончательный отчет об измененной оценке безопасности пропилгаллата». Международный журнал токсикологии . 26 (3_приложение): 89–118. 2007. дои : 10.1080/10915810701663176 . ПМИД 18080874 . S2CID 39562131 .
Пропилгаллат является общепризнанным безопасным (GRAS) антиоксидантом, защищающим жиры, масла и жиросодержащие продукты от прогоркания, возникающего в результате образования пероксидов.
- ^ Дебора, Жаклин; Клейд, Вивиан; Лусиана, Оливейра; Розмейр, Апаресида (7 августа 2019 г.). «Полифенолы как природные антиоксиданты в косметике». Журнал косметической дерматологии . 19 (1): 33–37. дои : 10.1111/jocd.13093 . ПМИД 31389656 . S2CID 201156301 .
- ^ Бузер CE, Хаммонд Г.С., Гамильтон CE, Сен JN (1955). «Окисление углеводородов воздухом.1II. Стехиометрия и судьба ингибиторов в бензоле и хлорбензоле». Журнал Американского химического общества . 77 (12): 3233–7. дои : 10.1021/ja01617a026 .
- ^ Перейти обратно: а б «Топливные антиоксиданты» . Инноспек Кемикалс. Архивировано из оригинала 15 октября 2006 года . Проверено 27 февраля 2007 г.
- ^ «Зачем использовать антиоксиданты?» . Специальные химические клеи. Архивировано из оригинала 11 февраля 2007 года . Проверено 27 февраля 2007 г.
- ^ Лю, Рунцэн; Мэбери, Скотт А. (6 октября 2020 г.). «Синтетические фенольные антиоксиданты: обзор возникновения, судьбы, воздействия на человека и токсичности в окружающей среде». Экологические науки и технологии . 54 (19): 11706–11719. Бибкод : 2020EnST...5411706L . doi : 10.1021/acs.est.0c05077 . ПМИД 32915564 . S2CID 221637214 .
- ^ Сюй, Цзин; Хао, Янфэнь; Ян, Жируо; Ли, Вэньцзюань; Се, Вэньцзин; Хуанг, Яни; Ван, Делян; Он, Юцин; Лян, Юн; Мацико, Юлиус; Ван, Пу (7 ноября 2022 г.). «Резиновые антиоксиданты и продукты их трансформации: появление в окружающей среде и потенциальное воздействие» . Международный журнал экологических исследований и общественного здравоохранения . 19 (21): 14595. doi : 10.3390/ijerph192114595 . ПМЦ 9657274 . ПМИД 36361475 .
- ^ Перейти обратно: а б Ли, Чао; Цуй, Синьи; Чен, Йи; Ляо, Чуньян; Ма, Лена К. (февраль 2019 г.). «Синтетические фенольные антиоксиданты и их основные метаболиты в ногте человека» . Экологические исследования . 169 : 308–314. Бибкод : 2019ER....169..308L . дои : 10.1016/j.envres.2018.11.020 . ПМИД 30500685 . S2CID 56486425 .
- ^ Лю, Рунцэн; Мэбери, Скотт А. (11 сентября 2020 г.). «Синтетические фенольные антиоксиданты: обзор возникновения, судьбы, воздействия на человека и токсичности в окружающей среде» . Окружающая среда. наук. Технол . 54 (19): 11706–11719. Бибкод : 2020EnST...5411706L . doi : 10.1021/acs.est.0c05077 . ПМИД 32915564 . S2CID 221637214 .
- ^ Ван, Ваньи; Сюн, Пин; Чжан, Хэ; Чжу, Цинцин; Ляо, Чуньян; Цзян, Гуйбинь (1 октября 2021 г.). «Анализ, возникновение, токсичность и риски для здоровья окружающей среды синтетических фенольных антиоксидантов: обзор» . Экологические исследования . 201 : 111531. Бибкод : 2021ER....201k1531W . дои : 10.1016/j.envres.2021.111531 . ISSN 0013-9351 . ПМИД 34146526 .
- ^ Чжан, Цзы-Фэн; Чжан, Сюэ; Сверко, Эд; Марвин, Кристофер Х.; Йобст, Карл Дж.; Смит, Ширли Энн; Ли, И-Фан (11 февраля 2020 г.). «Определение антиоксидантов дифениламина в сточных водах/твердых биологических веществах и отложениях» . Письма об экологической науке и технологиях . 7 (2): 102–110. Бибкод : 2020EnSTL...7..102Z . doi : 10.1021/acs.estlett.9b00796 . ISSN 2328-8930 . S2CID 213719260 .
- ^ Перейти обратно: а б с Дэвис К.Дж. (1995). «Окислительный стресс: парадокс аэробной жизни». Симпозиум Биохимического общества . 61 : 1–31. дои : 10.1042/bss0610001 . ПМИД 8660387 .
- ^ Перейти обратно: а б с д и Сис Х (март 1997 г.). «Окислительный стресс: оксиданты и антиоксиданты» . Экспериментальная физиология . 82 (2): 291–5. doi : 10.1113/expphysicalol.1997.sp004024 . ПМИД 9129943 . S2CID 20240552 .
- ^ Перейти обратно: а б с д Вертуани С., Ангусти А., Манфредини С. (2004). «Сеть антиоксидантов и проантиоксидантов: обзор». Текущий фармацевтический дизайн . 10 (14): 1677–94. дои : 10.2174/1381612043384655 . ПМИД 15134565 .
- ^ Ри С.Г. (июнь 2006 г.). «Клеточная сигнализация. H2O2, необходимое зло для клеточной сигнализации». Наука . 312 (5782): 1882–3. дои : 10.1126/science.1130481 . ПМИД 16809515 . S2CID 83598498 .
- ^ Валко М., Лейбфриц Д., Монкол Дж., Кронин М.Т., Мазур М., Тельсер Дж. (2007). «Свободные радикалы и антиоксиданты в нормальных физиологических функциях и заболеваниях человека». Международный журнал биохимии и клеточной биологии . 39 (1): 44–84. doi : 10.1016/j.biocel.2006.07.001 . ПМИД 16978905 .
- ^ Перейти обратно: а б Стохс С.Дж., Багчи Д. (февраль 1995 г.). «Окислительные механизмы токсичности ионов металлов» (PDF) . Свободная радикальная биология и медицина (представлена рукопись). 18 (2): 321–36. CiteSeerX 10.1.1.461.6417 . дои : 10.1016/0891-5849(94)00159-H . ПМИД 7744317 .
- ^ Накабеппу Ю, Сакуми К, Сакамото К, Цучимото Д, Цузуки Т, Накацу Ю (апрель 2006 г.). «Мутагенез и канцерогенез, вызванные окислением нуклеиновых кислот». Биологическая химия . 387 (4): 373–9. дои : 10.1515/BC.2006.050 . ПМИД 16606334 . S2CID 20217256 .
- ^ Валко М., Изакович М., Мазур М., Роудс С.Дж., Тельсер Дж. (ноябрь 2004 г.). «Роль кислородных радикалов в повреждении ДНК и заболеваемости раком». Молекулярная и клеточная биохимия . 266 (1–2): 37–56. дои : 10.1023/B:MCBI.0000049134.69131.89 . ПМИД 15646026 . S2CID 207547763 .
- ^ Штадтман Э.Р. (август 1992 г.). «Окисление белков и старение» . Наука . 257 (5074): 1220–4. Бибкод : 1992Sci...257.1220S . дои : 10.1126/science.1355616 . ПМИД 1355616 .
- ^ Раха С., Робинсон Б.Х. (октябрь 2000 г.). «Митохондрии, свободные радикалы кислорода, болезни и старение». Тенденции биохимических наук . 25 (10): 502–8. дои : 10.1016/S0968-0004(00)01674-1 . ПМИД 11050436 .
- ^ Леназ Г (2001). «Митохондриальное производство активных форм кислорода: механизмы и значение в патологии человека» . ИУБМБ Жизнь . 52 (3–5): 159–64. дои : 10.1080/15216540152845957 . ПМИД 11798028 . S2CID 45366190 .
- ^ Финкель Т., Холбрук, штат Нью-Джерси (ноябрь 2000 г.). «Окислители, окислительный стресс и биология старения». Природа . 408 (6809): 239–47. Бибкод : 2000Natur.408..239F . дои : 10.1038/35041687 . ПМИД 11089981 . S2CID 2502238 .
- ^ Херст Дж., Кинг М.С., Прайд К.Р. (октябрь 2008 г.). «Производство активных форм кислорода комплексом I». Труды Биохимического общества . 36 (Часть 5): 976–80. дои : 10.1042/BST0360976 . ПМИД 18793173 .
- ^ Сивер LC, Имлей Дж. А. (ноябрь 2004 г.). «Являются ли дыхательные ферменты основным источником внутриклеточной перекиси водорода?» . Журнал биологической химии . 279 (47): 48742–50. дои : 10.1074/jbc.M408754200 . ПМИД 15361522 .
- ^ Перейти обратно: а б Имлай Дж.А. (2003). «Пути окислительного повреждения». Ежегодный обзор микробиологии . 57 : 395–418. дои : 10.1146/annurev.micro.57.030502.090938 . ПМИД 14527285 .
- ^ Деммиг-Адамс Б., Адамс В.В. (декабрь 2002 г.). «Антиоксиданты в фотосинтезе и питании человека». Наука . 298 (5601): 2149–53. Бибкод : 2002Sci...298.2149D . дои : 10.1126/science.1078002 . ПМИД 12481128 . S2CID 27486669 .
- ^ Кригер-Лишкай А. (январь 2005 г.). «Производство синглетного кислорода при фотосинтезе». Журнал экспериментальной ботаники . 56 (411): 337–46. CiteSeerX 10.1.1.327.9651 . дои : 10.1093/jxb/erh237 . ПМИД 15310815 .
- ^ Куппер ФК, Карпентер Л.Дж., МакФигганс ГБ, Палмер С.Дж., Уэйт Т.Дж., Бонеберг Э.М., Войтш С., Вейллер М., Абела Р., Гролимунд Д., Потин П., Батлер А., Лютер Г.В., Кронек ПМХ, Мейер-Клауке В., Фейтерс М.С. ( 2008). «Накопление йодида обеспечивает водоросли неорганическим антиоксидантом, влияющим на химический состав атмосферы» . Труды Национальной академии наук . 105 (19): 6954–6958. Бибкод : 2008PNAS..105.6954K . дои : 10.1073/pnas.0709959105 . ISSN 0027-8424 . ПМК 2383960 . ПМИД 18458346 .
- ^ Сабо I, Бергантино Э, Джакометти ГМ (июль 2005 г.). «Световой и кислородный фотосинтез: рассеивание энергии как механизм защиты от фотоокисления» . Отчеты ЭМБО . 6 (7): 629–34. дои : 10.1038/sj.embor.7400460 . ПМЦ 1369118 . ПМИД 15995679 .
- ^ Керфельд, Калифорния (октябрь 2004 г.). «Водорастворимые каротиноиды цианобактерий» (PDF) . Архив биохимии и биофизики (Представлена рукопись). 430 (1): 2–9. дои : 10.1016/j.abb.2004.03.018 . ПМИД 15325905 . S2CID 25306222 .
- ^ Миллер Р.А., Бритиган Б.Е. (январь 1997 г.). «Роль оксидантов в микробной патофизиологии» . Обзоры клинической микробиологии . 10 (1): 1–18. дои : 10.1128/CMR.10.1.1 . ПМК 172912 . ПМИД 8993856 .
- ^ Шодьер Ж, Феррари-Илиу Р (1999). «Внутриклеточные антиоксиданты: от химических к биохимическим механизмам». Пищевая и химическая токсикология . 37 (9–10): 949–62. дои : 10.1016/S0278-6915(99)00090-3 . ПМИД 10541450 .
- ^ Сис Х (июль 1993 г.). «Стратегии антиоксидантной защиты» . Европейский журнал биохимии . 215 (2): 213–9. дои : 10.1111/j.1432-1033.1993.tb18025.x . PMID 7688300 .
- ^ Хоу К.Т., Вудхаус П. (июнь 1995 г.). «Взаимосвязь витамина С, инфекции, гемостатических факторов и сердечно-сосудистых заболеваний» . БМЖ . 310 (6994): 1559–63. дои : 10.1136/bmj.310.6994.1559 . ПМК 2549940 . ПМИД 7787643 .
- ^ Перейти обратно: а б с д Эвелсон П., Травасио М., Репетто М., Эскобар Дж., Ллесуй С., Лисси Э.А. (апрель 2001 г.). «Оценка общего реактивного антиоксидантного потенциала (TRAP) гомогенатов тканей и их цитозолей». Архив биохимии и биофизики . 388 (2): 261–6. дои : 10.1006/abbi.2001.2292 . ПМИД 11368163 .
- ^ Моррисон Дж.А., Якобсен Д.В., Спречер Д.Л., Робинсон К., Хури П., Дэниелс С.Р. (ноябрь 1999 г.). «Сывороточный глутатион у мужчин-подростков предсказывает ишемическую болезнь сердца у родителей» . Тираж . 100 (22): 2244–7. дои : 10.1161/01.CIR.100.22.2244 . ПМИД 10577998 .
- ^ Тейхерт Дж., Прейсс Р. (ноябрь 1992 г.). «ВЭЖХ-методы определения липоевой кислоты и ее восстановленной формы в плазме человека». Международный журнал клинической фармакологии, терапии и токсикологии . 30 (11): 511–2. ПМИД 1490813 .
- ^ Акиба С., Мацуго С., Пакер Л., Кониси Т. (май 1998 г.). «Анализ липоевой кислоты, связанной с белками, в тканях новым ферментативным методом». Аналитическая биохимия . 258 (2): 299–304. дои : 10.1006/abio.1998.2615 . ПМИД 9570844 .
- ^ Перейти обратно: а б Гланцунис Г.К., Цимояннис Е.К., Каппас А.М., Галарис Д.А. (2005). «Мочевая кислота и окислительный стресс». Текущий фармацевтический дизайн . 11 (32): 4145–51. дои : 10.2174/138161205774913255 . ПМИД 16375736 .
- ^ Эль-Сохеми А., Байлин А., Кабагамбе Э., Ашерио А., Шпигельман Д., Кампос Х. (июль 2002 г.). «Индивидуальные концентрации каротиноидов в жировой ткани и плазме как биомаркеры поступления с пищей» . Американский журнал клинического питания . 76 (1): 172–9. дои : 10.1093/ajcn/76.1.172 . ПМИД 12081831 .
- ^ Перейти обратно: а б Соуэлл А.Л., Хафф Д.Л., Йегер П.Р., Каудилл С.П., Гюнтер Э.В. (март 1994 г.). «Ретинол, альфа-токоферол, лютеин/зеаксантин, бета-криптоксантин, ликопин, альфа-каротин, транс-бета-каротин и четыре эфира ретинила в сыворотке, определяемые одновременно с помощью обращенно-фазовой ВЭЖХ с многоволновым обнаружением» . Клиническая химия . 40 (3): 411–6. дои : 10.1093/клинчем/40.3.411 . ПМИД 8131277 .
- ^ Шталь В., Шварц В., Сундквист А.Р., Сис Х. (апрель 1992 г.). «Цис-транс-изомеры ликопина и бета-каротина в сыворотке и тканях человека». Архив биохимии и биофизики . 294 (1): 173–7. дои : 10.1016/0003-9861(92)90153-N . ПМИД 1550343 .
- ^ Зита С., Овервад К., Мортенсен С.А., Синдберг К.Д., Моэсгаард С., Хантер Д.А. (2003). «Концентрация коэнзима Q10 в сыворотке крови у здоровых мужчин, принимавших 30 или 100 мг коэнзима Q10 в течение двух месяцев в рандомизированном контролируемом исследовании». Биофакторы . 18 (1–4): 185–93. дои : 10.1002/biof.5520180221 . ПМИД 14695934 . S2CID 19895215 .
- ^ Перейти обратно: а б Турунен М., Олссон Дж., Даллнер Г. (январь 2004 г.). «Метаболизм и функции коэнзима Q» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1660 (1–2): 171–99. дои : 10.1016/j.bbamem.2003.11.012 . ПМИД 14757233 .
- ^ Беккер Б.Ф. (июнь 1993 г.). «К физиологической функции мочевой кислоты». Свободно-радикальная биология и медицина . 14 (6): 615–31. дои : 10.1016/0891-5849(93)90143-I . ПМИД 8325534 .
- ^ Перейти обратно: а б Саутин Ю.Ю., Джонсон Р.Дж. (июнь 2008 г.). «Мочевая кислота: окислительно-антиоксидантный парадокс» . Нуклеозиды, нуклеотиды и нуклеиновые кислоты . 27 (6): 608–19. дои : 10.1080/15257770802138558 . ПМЦ 2895915 . ПМИД 18600514 .
- ^ Эномото А., Эндо Х. (сентябрь 2005 г.). «Роль переносчиков органических анионов (ОАТ) и переносчика уратов (URAT1) в патофизиологии заболеваний человека». Клиническая и экспериментальная нефрология . 9 (3): 195–205. дои : 10.1007/s10157-005-0368-5 . ПМИД 16189627 . S2CID 6145651 .
- ^ Эггебин AT (сентябрь 2007 г.). «Подагра: обновление» . Американский семейный врач . 76 (6): 801–8. ПМИД 17910294 .
- ^ Кэмпион Э.В., Глинн Р.Дж., ДеЛабри Л.О. (март 1987 г.). «Бессимптомная гиперурикемия. Риски и последствия в исследовании нормативного старения». Американский медицинский журнал . 82 (3): 421–6. дои : 10.1016/0002-9343(87)90441-4 . ПМИД 3826098 .
- ^ Бэйли Дж.К., Бейтс М.Г., Томпсон А.А., Уоринг В.С., Партридж Р.В., Шнопп М.Ф., Симпсон А., Гулливер-Слоан Ф., Максвелл С.Р., Уэбб DJ (май 2007 г.). «Эндогенное производство уратов увеличивает антиоксидантную способность плазмы у здоровых жителей равнин, находящихся на большой высоте». Грудь . 131 (5): 1473–8. дои : 10.1378/сундук.06-2235 . ПМИД 17494796 .
- ^ Назаревич Р.Р., Циолковски В., Ваккаро П.С., Гафурифар П. (декабрь 2007 г.). «Влияние краткосрочной кетогенной диеты на окислительно-восстановительный статус крови человека». Исследования омоложения . 10 (4): 435–40. дои : 10.1089/rej.2007.0540 . ПМИД 17663642 .
- ^ Перейти обратно: а б «Витамин С» . Информационный центр по микроэлементам, Институт Лайнуса Полинга, Университет штата Орегон, Корваллис, Орегон. 1 июля 2018 года . Проверено 19 июня 2019 г.
- ^ Падаятти С.Дж., Кац А., Ван Й., Эк П., Квон О., Ли Дж.Х., Чен С., Корпе С., Датта А., Датта С.К., Левин М. (февраль 2003 г.). «Витамин С как антиоксидант: оценка его роли в профилактике заболеваний» . Журнал Американского колледжа питания . 22 (1): 18–35. дои : 10.1080/07315724.2003.10719272 . ПМИД 12569111 . S2CID 21196776 .
- ^ Сигеока С., Исикава Т., Тамои М., Миягава Ю., Такеда Т., Ябута Ю., Ёсимура К. (май 2002 г.). «Регуляция и функция изоферментов аскорбатпероксидазы» . Журнал экспериментальной ботаники . 53 (372): 1305–19. дои : 10.1093/jexbot/53.372.1305 . ПМИД 11997377 .
- ^ Смирнофф Н., Уиллер Г.Л. (2000). «Аскорбиновая кислота в растениях: биосинтез и функции». Критические обзоры по биохимии и молекулярной биологии . 35 (4): 291–314. дои : 10.1080/10409230008984166 . ПМИД 11005203 . S2CID 85060539 .
- ^ Мейстер А. (апрель 1994 г.). «Антиоксидантная система глутатион-аскорбиновая кислота у животных» . Журнал биологической химии . 269 (13): 9397–400. дои : 10.1016/S0021-9258(17)36891-6 . ПМИД 8144521 .
- ^ Перейти обратно: а б с Мейстер А., Андерсон М.Е. (1983). «Глутатион». Ежегодный обзор биохимии . 52 : 711–60. дои : 10.1146/annurev.bi.52.070183.003431 . ПМИД 6137189 .
- ^ Габалла А., Ньютон Г.Л., Антельманн Х., Парсонейдж Д., Аптон Х., Рават М., Клэйборн А., Фэйи Р.К., Хелманн Дж.Д. (апрель 2010 г.). «Биосинтез и функции бациллитиола, основного низкомолекулярного тиола в бациллах» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (14): 6482–6. Бибкод : 2010PNAS..107.6482G . дои : 10.1073/pnas.1000928107 . ПМК 2851989 . ПМИД 20308541 .
- ^ Ньютон Г.Л., Рават М., Ла Клер Дж.Дж., Джотивасан В.К., Будиарто Т., Гамильтон С.Дж., Клэйборн А., Хелманн Дж.Д., Фэйи Р.С. (сентябрь 2009 г.). «Бациллитиол — это антиоксидантный тиол, вырабатываемый бациллами» . Химическая биология природы . 5 (9): 625–627. дои : 10.1038/nchembio.189 . ПМЦ 3510479 . ПМИД 19578333 .
- ^ Фэйи RC (2001). «Новые тиолы прокариот». Ежегодный обзор микробиологии . 55 : 333–56. дои : 10.1146/annurev.micro.55.1.333 . ПМИД 11544359 .
- ^ Фэрламб А.Х., Керами А. (1992). «Метаболизм и функции трипанотиона в кинетопластидах». Ежегодный обзор микробиологии . 46 : 695–729. дои : 10.1146/annurev.mi.46.100192.003403 . ПМИД 1444271 .
- ^ Перейти обратно: а б Эррера Э., Барбас С. (март 2001 г.). «Витамин Е: действие, метаболизм и перспективы». Журнал физиологии и биохимии . 57 (2): 43–56. дои : 10.1007/BF03179812 . HDL : 10637/720 . ПМИД 11579997 . S2CID 7272312 .
- ^ Пакер Л., Вебер С.Ю., Римбах Г. (февраль 2001 г.). «Молекулярные аспекты антиоксидантного действия альфа-токотриенола и передачи сигналов клеткам» . Журнал питания . 131 (2): 369С–73С. дои : 10.1093/jn/131.2.369S . ПМИД 11160563 .
- ^ Перейти обратно: а б Бригелиус-Флоэ Р., Трабер М.Г. (июль 1999 г.). «Витамин Е: функция и обмен веществ» . Журнал ФАСЭБ . 13 (10): 1145–55. CiteSeerX 10.1.1.337.5276 . дои : 10.1096/fasebj.13.10.1145 . ПМИД 10385606 . S2CID 7031925 .
- ^ Трабер М.Г., Аткинсон Дж. (июль 2007 г.). «Витамин Е, антиоксидант и ничего больше» . Свободно-радикальная биология и медицина . 43 (1): 4–15. doi : 10.1016/j.freeradbiomed.2007.03.024 . ПМК 2040110 . ПМИД 17561088 .
- ^ Ван X, Куинн Пи Джей (июль 1999 г.). «Витамин Е и его функция в мембранах». Прогресс в исследованиях липидов . 38 (4): 309–36. дои : 10.1016/S0163-7827(99)00008-9 . ПМИД 10793887 .
- ^ Зайлер А., Шнайдер М., Фёрстер Х., Рот С., Вирт Е.К., Калмзее С., Плеснила Н., Креммер Е., Родмарк О., Вурст В., Борнкамм Г.В., Швейцер У., Конрад М. (сентябрь 2008 г.). «Глутатионпероксидаза 4 воспринимает и преобразует окислительный стресс в 12/15-липоксигеназно-зависимую и AIF-опосредованную гибель клеток» . Клеточный метаболизм . 8 (3): 237–48. дои : 10.1016/j.cmet.2008.07.005 . ПМИД 18762024 .
- ^ Бригелиус-Флоэ Р., Дэвис К.Дж. (июль 2007 г.). «Является ли витамин Е антиоксидантом, регулятором передачи сигналов и экспрессии генов, или «нездоровой» пищей? Комментарии к двум сопроводительным статьям: «Молекулярный механизм действия альфа-токоферола» А. Аззи и «Витамин Е, антиоксидант и ничего больше» М. Трабера и Дж. Аткинсона». Свободно-радикальная биология и медицина . 43 (1): 2–3. doi : 10.1016/j.freeradbiomed.2007.05.016 . ПМИД 17561087 .
- ^ Аткинсон Дж., Эпанд РФ, Эпанд Р.М. (март 2008 г.). «Токоферолы и токотриенолы в мембранах: критический обзор». Свободно-радикальная биология и медицина . 44 (5): 739–64. doi : 10.1016/j.freeradbiomed.2007.11.010 . ПМИД 18160049 .
- ^ Аззи А (июль 2007 г.). «Молекулярный механизм действия альфа-токоферола». Свободно-радикальная биология и медицина . 43 (1): 16–21. doi : 10.1016/j.freeradbiomed.2007.03.013 . ПМИД 17561089 .
- ^ Зингг Дж. М., Аззи А. (май 2004 г.). «Неантиоксидантная активность витамина Е» . Современная медицинская химия . 11 (9): 1113–33. дои : 10.2174/0929867043365332 . ПМИД 15134510 . Архивировано из оригинала 6 октября 2011 года.
- ^ Сен К.К., Ханна С., Рой С. (март 2006 г.). «Токотриенолы: витамин Е за пределами токоферолов» . Науки о жизни . 78 (18): 2088–98. дои : 10.1016/j.lfs.2005.12.001 . ПМК 1790869 . ПМИД 16458936 .
- ^ Дуарте Т.Л., Лунец Дж. (июль 2005 г.). «Обзор: Когда антиоксидант не является антиоксидантом? Обзор новых действий и реакций витамина С». Свободные радикальные исследования . 39 (7): 671–86. дои : 10.1080/10715760500104025 . ПМИД 16036346 . S2CID 39962659 .
- ^ Перейти обратно: а б Шен, Цзяци; Гриффитс, Пол Т.; Кэмпбелл, Стивен Дж.; Утингер, Баттист; Калберер, Маркус; Полсон, Сюзанна Э. (1 апреля 2021 г.). «Окисление аскорбата железом, медью и активными формами кислорода: обзор, разработка модели и вывод ключевых констант скорости» . Научные отчеты . 11 (1): 7417. Бибкод : 2021NatSR..11.7417S . дои : 10.1038/s41598-021-86477-8 . ISSN 2045-2322 . ПМК 8016884 . ПМИД 33795736 .
- ^ Перейти обратно: а б Карр А., Фрей Б. (июнь 1999 г.). «Действует ли витамин С как прооксидант в физиологических условиях?» . Журнал ФАСЭБ . 13 (9): 1007–24. дои : 10.1096/fasebj.13.9.1007 . ПМИД 10336883 . S2CID 15426564 .
- ^ Перейти обратно: а б Хо Ю.С., Магненат Дж.Л., Гаргано М., Цао Дж. (октябрь 1998 г.). «Природа механизмов антиоксидантной защиты: урок трансгенных исследований» . Перспективы гигиены окружающей среды . 106 (Приложение 5): 1219–28. дои : 10.2307/3433989 . JSTOR 3433989 . ПМЦ 1533365 . ПМИД 9788901 .
- ^ Зелько И.Н., Мариани Т.Дж., Фольц Р.Дж. (август 2002 г.). «Мультигенное семейство супероксиддисмутазы: сравнение структур, эволюции и экспрессии генов CuZn-SOD (SOD1), Mn-SOD (SOD2) и EC-SOD (SOD3)». Свободно-радикальная биология и медицина . 33 (3): 337–49. дои : 10.1016/S0891-5849(02)00905-X . ПМИД 12126755 .
- ^ Перейти обратно: а б Баннистер СП, Баннистер WH, Ротилио Г (1987). «Аспекты структуры, функции и применения супероксиддисмутазы». Критические обзоры CRC по биохимии . 22 (2): 111–80. дои : 10.3109/10409238709083738 . ПМИД 3315461 .
- ^ Джонсон Ф., Джуливи С. (2005). «Супероксиддисмутазы и их влияние на здоровье человека». Молекулярные аспекты медицины . 26 (4–5): 340–52. дои : 10.1016/j.mam.2005.07.006 . ПМИД 16099495 .
- ^ Нозик-Грейк Э., Сулиман Х.Б., Пиантадоси Калифорния (декабрь 2005 г.). «Внеклеточная супероксиддисмутаза». Международный журнал биохимии и клеточной биологии . 37 (12): 2466–71. doi : 10.1016/j.biocel.2005.06.012 . ПМИД 16087389 .
- ^ Мелов С., Шнайдер Дж.А., Дэй Б.Дж., Хинерфельд Д., Коскун П., Мирра С.С., Крапо Дж.Д., Уоллес Д.К. (февраль 1998 г.). «Новый неврологический фенотип у мышей, лишенных митохондриальной супероксиддисмутазы марганца». Природная генетика . 18 (2): 159–63. дои : 10.1038/ng0298-159 . ПМИД 9462746 . S2CID 20843002 .
- ^ Реоме А.Г., Эллиотт Дж.Л., Хоффман Э.К., Ковалл Н.В., Ферранте Р.Дж., Сивек Д.Ф., Уилкокс Х.М., Флуд Д.Г., Бил М.Ф., Браун Р.Х., Скотт Р.В., Снайдер В.Д. (май 1996 г.). «Мотонейроны у мышей с дефицитом супероксиддисмутазы Cu/Zn развиваются нормально, но демонстрируют повышенную гибель клеток после повреждения аксонов». Природная генетика . 13 (1): 43–7. дои : 10.1038/ng0596-43 . ПМИД 8673102 . S2CID 13070253 .
- ^ Ван Кэмп В., Инзе Д., Ван Монтегю М. (1997). «Регуляция и функция супероксиддисмутазы табака» . Свободно-радикальная биология и медицина . 23 (3): 515–20. дои : 10.1016/S0891-5849(97)00112-3 . ПМИД 9214590 .
- ^ Челикани П., Фита И., Левен ПК (январь 2004 г.). «Разнообразие структур и свойств каталаз» (PDF) . Клеточные и молекулярные науки о жизни (представленная рукопись). 61 (2): 192–208. дои : 10.1007/s00018-003-3206-5 . hdl : 10261/111097 . ПМИД 14745498 . S2CID 4411482 .
- ^ Замоцкий М, Коллер Ф (1999). «Понимание структуры и функции каталаз: подсказки молекулярной эволюции и мутагенеза in vitro» . Прогресс биофизики и молекулярной биологии . 72 (1): 19–66. дои : 10.1016/S0079-6107(98)00058-3 . ПМИД 10446501 .
- ^ дель Рио Л.А., Сандалио Л.М., Пальма Х.М., Буэно П., Корпус Ф.Д. (ноябрь 1992 г.). «Метаболизм кислородных радикалов в пероксисомах и клеточные последствия». Свободно-радикальная биология и медицина . 13 (5): 557–80. дои : 10.1016/0891-5849(92)90150-F . ПМИД 1334030 .
- ^ Хинер А.Н., Рэйвен Э.Л., Торнели Р.Н., Гарсиа-Кановас Ф., Родригес-Лопес Х.Н. (июль 2002 г.). «Механизмы образования соединения I в гемопероксидазах». Журнал неорганической биохимии . 91 (1): 27–34. дои : 10.1016/S0162-0134(02)00390-2 . ПМИД 12121759 .
- ^ Мюллер С., Ридель Х.Д., Штреммель В. (декабрь 1997 г.). «Прямое доказательство того, что каталаза является преобладающим ферментом, удаляющим H2O2, в эритроцитах человека» . Кровь . 90 (12): 4973–8. дои : 10.1182/blood.V90.12.4973 . ПМИД 9389716 .
- ^ Огата М. (февраль 1991 г.). «Акаталаземия». Генетика человека . 86 (4): 331–40. дои : 10.1007/BF00201829 . ПМИД 1999334 . S2CID 264033871 .
- ^ Пасторский Д., Янгблад Д., Сарма Г., Вуд З., Карплюс П., Пул Л. (2005). «Анализ связи между ферментативной активностью и олигомерным состоянием AhpC, бактериального пероксиредоксина» . Биохимия . 44 (31): 10583–92. дои : 10.1021/bi050448i . ПМЦ 3832347 . ПМИД 16060667 . ПДБ 1YEX
- ^ Ри С.Г., Че Х.З., Ким К. (июнь 2005 г.). «Пероксиредоксины: исторический обзор и умозрительный обзор новых механизмов и новых концепций клеточной сигнализации». Свободно-радикальная биология и медицина . 38 (12): 1543–52. doi : 10.1016/j.freeradbiomed.2005.02.026 . ПМИД 15917183 .
- ^ Вуд З.А., Шредер Э., Робин Харрис Дж., Пул Л.Б. (январь 2003 г.). «Структура, механизм и регуляция пероксиредоксинов». Тенденции биохимических наук . 28 (1): 32–40. дои : 10.1016/S0968-0004(02)00003-8 . ПМИД 12517450 .
- ^ Клэйборн А., Йе Дж.И., Маллет Т.С., Люба Дж., Крейн Э.Дж., Чарриер В., Пасторский Д. (ноябрь 1999 г.). «Белко-сульфеновые кислоты: разнообразные роли маловероятного игрока в ферментативном катализе и окислительно-восстановительной регуляции». Биохимия . 38 (47): 15407–16. дои : 10.1021/bi992025k . ПМИД 10569923 . S2CID 29055779 .
- ^ Йонссон Т.Дж., Лоутер В.Т. (2007). «Белки репарации пероксиредоксина». Пероксиредоксиновые системы . Субклеточная биохимия. Том. 44. С. 115–41. дои : 10.1007/978-1-4020-6051-9_6 . ISBN 978-1-4020-6050-2 . ПМК 2391273 . ПМИД 18084892 .
- ^ Нойманн К.А., Краузе Д.С., Карман К.В., Дас С., Дубей Д.П., Абрахам Дж.Л., Бронсон Р.Т., Фудзивара Ю., Оркин Ш.Х., Ван Эттен Р.А. (июль 2003 г.). «Основная роль пероксиредоксина Prdx1 в антиоксидантной защите эритроцитов и подавлении опухолей» (PDF) . Природа . 424 (6948): 561–5. Бибкод : 2003Natur.424..561N . дои : 10.1038/nature01819 . ПМИД 12891360 . S2CID 3570549 .
- ^ Ли Т.Х., Ким СУ, Ю СЛ, Ким Ш., Пак Д.С., Мун Х.Б., До Ш., Квон К.С., Квон Х.Дж., Хан Ю.Х., Чон С., Кан С.В., Шин Х.С., Ли К.К., Ри С.Г., Ю Д.И. (июнь 2003 г.) ). «Пероксиредоксин II необходим для поддержания продолжительности жизни эритроцитов у мышей» . Кровь . 101 (12): 5033–8. дои : 10.1182/кровь-2002-08-2548 . ПМИД 12586629 .
- ^ Дитц К.Дж., Джейкоб С., Оэльце М.Л., Лакса М., Тогнетти В., де Миранда С.М., Байер М., Финкемайер I (2006). «Функция пероксиредоксинов в окислительно-восстановительном метаболизме органелл растений» . Журнал экспериментальной ботаники . 57 (8): 1697–709. дои : 10.1093/jxb/erj160 . ПМИД 16606633 .
- ^ Нордберг Дж., Арнер Э.С. (декабрь 2001 г.). «Активные формы кислорода, антиоксиданты и тиоредоксиновая система млекопитающих». Свободно-радикальная биология и медицина . 31 (11): 1287–312. дои : 10.1016/S0891-5849(01)00724-9 . ПМИД 11728801 .
- ^ Виейра Дос Сантос К., Рей П. (июль 2006 г.). «Растительные тиоредоксины являются ключевыми участниками реакции на окислительный стресс». Тенденции в науке о растениях . 11 (7): 329–34. doi : 10.1016/j.tplants.2006.05.005 . ПМИД 16782394 .
- ^ Арнер Э.С., Холмгрен А. (октябрь 2000 г.). «Физиологические функции тиоредоксина и тиоредоксинредуктазы» . Европейский журнал биохимии . 267 (20): 6102–9. дои : 10.1046/j.1432-1327.2000.01701.x . ПМИД 11012661 .
- ^ Мустачич Д., Повис Дж. (февраль 2000 г.). «Тиоредоксинредуктаза» . Биохимический журнал . 346 (1): 1–8. дои : 10.1042/0264-6021:3460001 . ПМК 1220815 . ПМИД 10657232 .
- ^ Крейссен Г., Бродбент П., Стивенс Р., Веллберн А.Р., Муллино П. (май 1996 г.). «Манипулирование метаболизмом глутатиона в трансгенных растениях». Труды Биохимического общества . 24 (2): 465–9. дои : 10.1042/bst0240465 . ПМИД 8736785 .
- ^ Бригелиус-Флоэ Р. (ноябрь 1999 г.). «Тканеспецифические функции отдельных глутатионпероксидаз». Свободно-радикальная биология и медицина . 27 (9–10): 951–65. дои : 10.1016/S0891-5849(99)00173-2 . ПМИД 10569628 .
- ^ Хо Ю.С., Магненат Дж.Л., Бронсон РТ, Као Дж., Гаргано М., Сугавара М., Funk CD (июнь 1997 г.). «Мыши с дефицитом клеточной глутатионпероксидазы развиваются нормально и не проявляют повышенной чувствительности к гипероксии» . Журнал биологической химии . 272 (26): 16644–51. дои : 10.1074/jbc.272.26.16644 . ПМИД 9195979 .
- ^ де Хаан Дж.Б., Бладье С., Гриффитс П., Келнер М., О'Ши Р.Д., Ченг Н.С., Бронсон Р.Т., Сильвестро М.Дж., Уайлд С., Чжэн СС, Берт П.М., Герцог П.Дж., Кола I (август 1998 г.). «Мыши с гомозиготной нулевой мутацией наиболее распространенной глутатионпероксидазы, Gpx1, демонстрируют повышенную восприимчивость к параквату и перекиси водорода, вызывающим окислительный стресс» . Журнал биологической химии . 273 (35): 22528–36. дои : 10.1074/jbc.273.35.22528 . hdl : 10536/DRO/DU:30101410 . ПМИД 9712879 .
- ^ Шарма Р., Ян Й., Шарма А., Авасти С., Авасти Ю.К. (апрель 2004 г.). «Антиоксидантная роль глутатион-S-трансфераз: защита от окислительной токсичности и регуляция стресс-опосредованного апоптоза». Антиоксиданты и окислительно-восстановительная сигнализация . 6 (2): 289–300. дои : 10.1089/152308604322899350 . ПМИД 15025930 .
- ^ Хейс Дж. Д., Фланаган Дж. Ю., Джоуси И. Р. (2005). «Глутатионтрансферазы». Ежегодный обзор фармакологии и токсикологии . 45 : 51–88. doi : 10.1146/annurev.pharmtox.45.120403.095857 . ПМИД 15822171 .
- ^ Перейти обратно: а б Стэннер С.А., Хьюз Дж., Келли С.Н., Баттрисс Дж. (май 2004 г.). «Обзор эпидемиологических данных в пользу «антиоксидантной гипотезы» » . Питание общественного здравоохранения . 7 (3): 407–22. дои : 10.1079/PHN2003543 . ПМИД 15153272 .
- ^ Еда, питание, физическая активность и профилактика рака: глобальная перспектива. Архивировано 23 сентября 2015 года в Wayback Machine . Всемирный фонд исследования рака (2007). ISBN 978-0-9722522-2-5 .
- ^ Райс-Эванс, Кэтрин А.; Миллер, Николас Дж.; Паганга, Джордж (1996). «Взаимосвязь структуры и антиоксидантной активности флавоноидов и фенольных кислот» . Свободнорадикальная биология и медицина . 20 (7): 933–956. дои : 10.1016/0891-5849(95)02227-9 . ПМИД 8743980 .
- ^ Дель Рио Д., Родригес-Матеос А., Спенсер Дж.П., Тоньолини М., Борхес Г., Крозье А. (май 2013 г.). «Диетические (поли)фенолы для здоровья человека: структура, биодоступность и доказательства защитного действия против хронических заболеваний» . Антиоксиданты и окислительно-восстановительная сигнализация . 18 (14): 1818–1892. дои : 10.1089/ars.2012.4581 . ПМК 3619154 . ПМИД 22794138 .
- ^ «Флавоноиды» . Институт Лайнуса Полинга, Университет штата Орегон, Корваллис. 2016 . Проверено 24 июля 2016 г.
- ^ Леммо В. (сентябрь 2014 г.). «Потенциальное взаимодействие рецептурных и безрецептурных препаратов, обладающих антиоксидантными свойствами, с лучевой и химиотерапией» . Международный журнал рака . 137 (11): 2525–33. дои : 10.1002/ijc.29208 . ПМИД 25220632 . S2CID 205951215 .
- ^ Форман, Генри Джей; Чжан, Хунцяо (30 июня 2021 г.). «Нацеливание на окислительный стресс при заболеваниях: перспективы и ограничения антиоксидантной терапии» . Nature Reviews Открытие лекарств . 20 (9): 689–709. дои : 10.1038/s41573-021-00233-1 . ISSN 1474-1784 . ПМЦ 8243062 . ПМИД 34194012 .
- ^ Харрелл РФ (сентябрь 2003 г.). «Влияние источников растительного белка на биодоступность микроэлементов и минералов» . Журнал питания . 133 (9): 2973С–7С. дои : 10.1093/jn/133.9.2973S . ПМИД 12949395 .
- ^ Хант-младший (сентябрь 2003 г.). «Биодоступность железа, цинка и других микроэлементов в вегетарианской диете» . Американский журнал клинического питания . 78 (3 доп.): 633S–639S. дои : 10.1093/ajcn/78.3.633S . ПМИД 12936958 .
- ^ Гибсон Р.С., Перлас Л., Хотц С. (май 2006 г.). «Улучшение биодоступности питательных веществ растительных продуктов на бытовом уровне» . Труды Общества питания . 65 (2): 160–8. дои : 10.1079/PNS2006489 . ПМИД 16672077 .
- ^ Перейти обратно: а б Моша Т.С., Гага Х.Э., Пейс Р.Д., Ласвай Х.С., Мтебе К. (июнь 1995 г.). «Влияние бланширования на содержание антипитательных факторов в отдельных овощах». Растительные продукты для питания человека . 47 (4): 361–7. дои : 10.1007/BF01088275 . ПМИД 8577655 . S2CID 1118651 .
- ^ Сандберг А.С. (декабрь 2002 г.). «Биодоступность минералов в бобовых» . Британский журнал питания . 88 (Приложение 3): S281–5. дои : 10.1079/BJN/2002718 . ПМИД 12498628 .
- ^ Перейти обратно: а б Бичер Г.Р. (октябрь 2003 г.). «Обзор пищевых флавоноидов: номенклатура, распространение и потребление» . Журнал питания . 133 (10): 3248С–3254С. дои : 10.1093/jn/133.10.3248S . ПМИД 14519822 .
- ^ Оменн Г.С., Гудман Г.Е., Торнквист М.Д., Балмс Дж., Каллен М.Р., Гласс А., Кио Дж.П., Мейскенс Ф.Л., Валанис Б., Уильямс Дж.Х., Барнхарт С., Черняк М.Г., Бродкин К.А., Хаммар С. (ноябрь 1996 г.). «Факторы риска рака легких и эффекты вмешательства в CARET, исследовании эффективности бета-каротина и ретинола» (PDF) . Журнал Национального института рака . 88 (21): 1550–9. дои : 10.1093/jnci/88.21.1550 . ПМИД 8901853 .
- ^ Альбанес Д. (июнь 1999 г.). «Бета-каротин и рак легких: пример» . Американский журнал клинического питания . 69 (6): 1345С–50С. дои : 10.1093/ajcn/69.6.1345S . ПМИД 10359235 .
- ^ Перейти обратно: а б Белакович Г., Николова Д., Глууд Л.Л., Симонетти Р.Г., Глууд С. (февраль 2007 г.). «Смертность в рандомизированных исследованиях антиоксидантных добавок для первичной и вторичной профилактики: систематический обзор и метаанализ». ДЖАМА . 297 (8): 842–57. дои : 10.1001/jama.297.8.842 . ПМИД 17327526 .
- ^ Перейти обратно: а б Белакович Г, Николова Д, Глууд ЛЛ, Симонетти Р.Г., Глууд С (14 марта 2012 г.). «Антиоксидантные добавки для профилактики смертности здоровых участников и пациентов с различными заболеваниями» . Кокрановская база данных систематических обзоров . 2012 (3): CD007176. дои : 10.1002/14651858.CD007176.pub2 . hdl : 10138/136201 . ПМЦ 8407395 . ПМИД 22419320 .
- ^ Эксперты утверждают, что исследование, в котором упоминаются риски антиоксидантных витаминов, основано на ошибочной методологии. Пресс-релиз Университета штата Орегон опубликован на ScienceDaily. Проверено 19 апреля 2007 г.
- ^ Миллер Э.Р., Пастор-Барриузо Р., Далал Д., Римерсма Р.А., Аппель Л.Дж., Гуаллар Э. (январь 2005 г.). «Метаанализ: высокие дозы витамина Е могут увеличить смертность от всех причин» . Анналы внутренней медицины . 142 (1): 37–46. дои : 10.7326/0003-4819-142-1-200501040-00110 . ПМИД 15537682 .
- ^ Перейти обратно: а б Белакович Г., Нагорни А., Николова Д., Симонетти Р.Г., Белакович М., Глууд С. (июль 2006 г.). «Метаанализ: антиоксидантные добавки для первичной и вторичной профилактики колоректальной аденомы» . Алиментарная фармакология и терапия . 24 (2): 281–91. дои : 10.1111/j.1365-2036.2006.02970.x . ПМИД 16842454 . S2CID 20452618 .
- ^ Кортес-Жофре М., Руэда Х.Р., Асенхо-Лобос К., Мадрид Е., Бонфилл Косп X (март 2020 г.). «Препараты для профилактики рака легких у здоровых людей» . Кокрановская база данных систематических обзоров . 2020 (3): CD002141. дои : 10.1002/14651858.CD002141.pub3 . ПМК 7059884 . ПМИД 32130738 .
- ^ Шенкин А (февраль 2006 г.). «Ключевая роль микроэлементов». Клиническое питание . 25 (1): 1–13. дои : 10.1016/j.clnu.2005.11.006 . ПМИД 16376462 .
- ^ Ранчордас, Маюр К.; Роджерсон, Дэвид; Солтани, Хора; Костелло, Джозеф Т. (14 декабря 2017 г.). «Антиоксиданты для предотвращения и уменьшения мышечных болей после тренировки» . Кокрановская база данных систематических обзоров . 2017 (12): CD009789. дои : 10.1002/14651858.CD009789.pub2 . ISSN 1469-493X . ПМК 6486214 . ПМИД 29238948 .
- ^ Родригес-Амая Д.Б. (2003). «Пищевые каротиноиды: анализ, состав и изменения при хранении и обработке пищевых продуктов». Форум питания . 56 : 35–7. PMID 15806788 .
- ^ Майани Дж., Кастон М.Дж., Катаста Г., Тоти Э., Камбродон И.Г., Бистед А., Гранадо-Лоренсио Ф., Ольмедилья-Алонсо Б., Кнутсен П., Валоти М., Бём В., Майер-Мибах Е., Беснилиан Д., Шлеммер Ю. (сентябрь 2009 г.) ). «Каротиноиды: актуальные знания об источниках пищи, потреблении, стабильности и биодоступности, а также их защитной роли в организме человека» . Молекулярное питание и пищевые исследования . 53 (Приложение 2): S194–218. дои : 10.1002/mnfr.200800053 . hdl : 10261/77697 . ПМИД 19035552 . Архивировано из оригинала 27 сентября 2018 года . Проверено 18 апреля 2017 г. .
- ^ Генри С.Дж., Хеппелл Н. (февраль 2002 г.). «Пищевые потери и выгоды при переработке: будущие проблемы и проблемы» . Труды Общества питания . 61 (1): 145–8. дои : 10.1079/PNS2001142 . ПМИД 12002789 .
- ^ «Антиоксиданты и профилактика рака: информационный бюллетень» . Национальный институт рака. Архивировано из оригинала 4 марта 2007 года . Проверено 27 февраля 2007 г.
- ^ Ортега Р. (декабрь 2006 г.). «Важность функциональных продуктов в средиземноморской диете» . Питание общественного здравоохранения . 9 (8А): 1136–40. дои : 10.1017/S1368980007668530 . ПМИД 17378953 .
- ^ Витчи А., Редди С., Стофер Б., Лаутербург Б.Х. (1992). «Системная доступность перорального глутатиона». Европейский журнал клинической фармакологии . 43 (6): 667–9. дои : 10.1007/BF02284971 . ПМИД 1362956 . S2CID 27606314 .
- ^ Флэгг Э.В., Коутс Р.Дж., Эли Дж.В., Джонс Д.П., Гюнтер Э.В., Байерс Т.Е., Блок Г.С., Гринберг Р.С. (1994). «Потребление глутатиона с пищей у людей и взаимосвязь между потреблением и общим уровнем глутатиона в плазме». Питание и рак . 21 (1): 33–46. дои : 10.1080/01635589409514302 . ПМИД 8183721 .
- ^ Додд С., Дин О., Кополов Д.Л., Малхи Г.С., Берк М. (декабрь 2008 г.). «N-ацетилцистеин для антиоксидантной терапии: фармакология и клиническое применение». Экспертное мнение о биологической терапии . 8 (12): 1955–62. дои : 10.1517/14728220802517901 . ПМИД 18990082 . S2CID 74736842 .
- ^ ван де Полл MC, Деджонг CH, Сотерс ПБ (июнь 2006 г.). «Адекватный диапазон серосодержащих аминокислот и биомаркеры их избытка: уроки энтерального и парентерального питания» . Журнал питания . 136 (6 Доп.): 1694–1700С. дои : 10.1093/jn/136.6.1694S . ПМИД 16702341 .
- ^ Цао Дж., Алессио Х.М., Катлер Р.Г. (март 1993 г.). «Определение способности антиоксидантов поглощать кислородные радикалы» . Свободно-радикальная биология и медицина . 14 (3): 303–11. дои : 10.1016/0891-5849(93)90027-R . ПМИД 8458588 .
- ^ Оу Б., Хэмпш-Вудилл М., Приор Р.Л. (октябрь 2001 г.). «Разработка и проверка улучшенного анализа способности поглощать кислородные радикалы с использованием флуоресцеина в качестве флуоресцентного зонда». Журнал сельскохозяйственной и пищевой химии . 49 (10): 4619–26. дои : 10.1021/jf010586o . ПМИД 11599998 .
- ^ Перейти обратно: а б «Снято: способность некоторых продуктов поглощать радикалы кислорода (ORAC), выпуск 2 (2010 г.)» . Министерство сельского хозяйства США, Служба сельскохозяйственных исследований. 16 мая 2012 года . Проверено 13 июня 2012 г.
- ^ Приор Р.Л., Ву X, Шайх К. (май 2005 г.). «Стандартизированные методы определения антиоксидантной способности и фенольных соединений в пищевых продуктах и пищевых добавках» (PDF) . Журнал сельскохозяйственной и пищевой химии . 53 (10): 4290–302. дои : 10.1021/jf0502698 . ПМИД 15884874 . Архивировано из оригинала (PDF) 29 декабря 2016 года . Проверено 24 октября 2017 г.
Дальнейшее чтение
[ редактировать ]- Холливелл Б., Гаттеридж Дж. М. (2015). Свободные радикалы в биологии и медицине (5-е изд.). Издательство Оксфордского университета. ISBN 978-0-19-856869-8 .
- Лейн Н (2003). Кислород: молекула, создавшая мир . Издательство Оксфордского университета. ISBN 978-0-19-860783-0 .
- Покорный Дж., Янишлиева Н., Гордон М.Х. (2001). Антиоксиданты в продуктах питания: практическое применение . ЦРК Пресс. ISBN 978-0-8493-1222-9 .
Внешние ссылки
[ редактировать ] СМИ, связанные с антиоксидантами, на Викискладе?
СМИ, связанные с антиоксидантами, на Викискладе?