Дыхательный комплекс I

Дыхательный комплекс I , EC 7.1.1.2 (также известный как НАДН:убихиноноксидоредуктаза , НАДН-дегидрогеназа I типа и митохондриальный комплекс I ) — первый крупный белковый комплекс дыхательных цепей многих организмов от бактерий до человека. Он катализирует перенос электронов от НАДН к коферменту Q10 (CoQ10) и перемещает протоны через внутреннюю митохондриальную мембрану у эукариот или плазматическую мембрану бактерий.
| Дыхательный комплекс I | |
|---|---|
| Идентификаторы | |
| Символ | Дыхательный комплекс I |
| Суперсемейство OPM | 246 |
| белок OPM | 6г72 |
| Мембраном | 255 |
| Идентификаторы | |||
|---|---|---|---|
| Номер ЕС. | 7.1.1.2 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Этот фермент необходим для нормального функционирования клеток, а мутации в его субъединицах приводят к широкому спектру наследственных нервно-мышечных и метаболических нарушений. Дефекты этого фермента ответственны за развитие ряда патологических процессов, таких как ишемия/реперфузионное повреждение ( инсульт и инфаркт ), болезнь Паркинсона и другие. [ нужна ссылка ]
Функция
[ редактировать ]

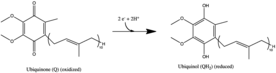
Комплекс I — первый фермент митохондриальной цепи переноса электронов . В цепи переноса электронов присутствуют три фермента-передатчика энергии — НАДН:убихиноноксидоредуктаза (комплекс I), кофермент Q — цитохром с-редуктаза (комплекс III) и цитохром с-оксидаза (комплекс IV). [1] Комплекс I — самый крупный и сложный фермент цепи переноса электронов. [2]
Реакция, катализируемая комплексом I:
- НАДН + Н + + CoQ + 4H + в → НАД + + CoQH 2 + 4H + вне
В этом процессе комплекс перемещает четыре протона через внутреннюю мембрану на молекулу окисленного НАДН . [3] [4] [5] помогает создать разность электрохимических потенциалов , используемую для производства АТФ . Комплекс I Escherichia coli (НАДН-дегидрогеназа) способен перемещать протоны в том же направлении, что и установленный Δψ , что показывает, что в тестируемых условиях ионом связи является H + . [6] Уже + наблюдался транспорт в противоположном направлении, и хотя Na + не был необходим для каталитической или протонной транспортной активности, его присутствие усиливало последнюю. ЧАС + был транслоцирован комплексом I Paracoccus denitrificans , но в данном случае H + транспорт не подвергался влиянию Na + и На + транспорта не наблюдалось. Возможно, комплекс I E. coli имеет два центра энергетического связывания (один Na + независимый и другой Na + зависит), как это наблюдается для комплекса I Rhodothermus marinus , тогда как механизм связывания фермента P. denitrificans полностью основан на Na + независимый. Также возможно, что захват Na катализирует другой переносчик. + . Трансдукция энергии комплекса I посредством протонной накачки может быть свойственна не только ферменту R. marinus . На + /ЧАС + Антипортовая активность, по-видимому, не является общим свойством комплекса I. [6] Однако существование Na + -транслокационная активность комплекса I остается под вопросом.
Реакцию можно обратить вспять – это называется аэробным НАД, поддерживаемым сукцинатом. + восстановление убихинолом – при наличии высокого мембранного потенциала, но точный каталитический механизм остается неизвестным. Движущей силой этой реакции является потенциал на мембране, который может поддерживаться либо за счет гидролиза АТФ, либо за счет комплексов III и IV при окислении сукцината. [7]
Комплекс I может играть роль в запуске апоптоза . [8] Фактически, было показано, что существует корреляция между активностью митохондрий и запрограммированной гибелью клеток (PCD) во время развития соматических эмбрионов. [9]
Комплекс I не гомологичен Na. + -транслоцирующая НАДН-дегидрогеназа (NDH) семейства ( TC# 3.D.1 ), член Na + перевозка суперсемейства Mrp .
В результате окисления двух молекул НАДН до НАД+ три молекулы АТФ могут образовываться Комплексом V ( АТФ-синтаза ) ниже по дыхательной цепи.
Механизм
[ редактировать ]Общий механизм
[ редактировать ]Все окислительно-восстановительные реакции происходят в гидрофильном домене комплекса I. НАДН первоначально связывается с комплексом I и переносит два электрона на простетическую группу флавинмононуклеотида (FMN) фермента, создавая FMNH 2 . Акцептор электронов – изоаллоксазиновое кольцо – FMN идентичен таковому FAD . Затем электроны передаются через FMN через серию железо-серных кластеров (Fe-S). [10] и, наконец, коэнзим Q10 (убихинон). Этот поток электронов меняет окислительно-восстановительное состояние белка, вызывая конформационные изменения белка, которые изменяют значения p K ионизируемой боковой цепи и вызывают выкачивание четырех ионов водорода из митохондриального матрикса. [11] Убихинон (CoQ) принимает два электрона и восстанавливается до убихинола (CoQH 2 ). [1]
Механизм переноса электронов
[ редактировать ]Предлагаемый путь транспорта электронов перед восстановлением убихинона выглядит следующим образом: НАДН – ФМН – N3 – N1b – N4 – N5 – N6a – N6b – N2 – Q, где Nx представляет собой соглашение о маркировке для кластеров железа и серы. [10] Высокий восстановительный потенциал кластера N2 и относительная близость остальных кластеров в цепи обеспечивают эффективный перенос электронов на большие расстояния в белке (со скоростью переноса от НАДН к железо-серному кластеру N2 около 100 мкс). [12] [13]
Динамика равновесия Комплекса I в первую очередь определяется окислительно-восстановительным циклом хинона. В условиях высокой движущей силы протонов (и, соответственно, пула, концентрированного убихинолом), фермент работает в обратном направлении. Убихинол окисляется до убихинона, и образующиеся в результате высвобождения протоны уменьшают движущую силу протонов. [14]
Механизм транслокации протонов
[ редактировать ]В настоящее время предполагается, что взаимодействие транслокации протонов и транспорта электронов в Комплексе I является непрямым (конформационные изменения на большие расстояния), а не прямым (окислительно-восстановительные промежуточные соединения в водородных насосах, как в гемовых группах Комплексов III и IV ). [10] Архитектура гидрофобной области комплекса I демонстрирует множественные переносчики протонов, которые механически связаны между собой. Считается, что три центральных компонента, вносящие вклад в это событие конформационного изменения на большие расстояния, - это связанный с pH N2-железо-серный кластер, восстановление хинона и субъединицы трансмембранной спирали плеча мембраны. Трансдукция конформационных изменений для управления трансмембранными переносчиками, связанными «шатуном» во время восстановления убихинона, может составлять два или три из четырех протонов, перекачиваемых на каждый окисленный НАДН. Оставшийся протон должен быть перекачен путем прямого связывания в месте связывания убихинона. Предполагается, что механизмы прямой и косвенной связи объясняют накачку четырех протонов. [15]
Близость кластера N2 к близлежащему остатку цистеина приводит к конформационным изменениям при восстановлении близлежащих спиралей, что приводит к небольшим, но важным изменениям в общей конформации белка. [16] Дальнейшие исследования электронного парамагнитного резонанса переноса электронов показали, что большая часть энергии, которая высвобождается во время последующего восстановления CoQ, приходится на финальную стадию образования убихинола из семихинона , что свидетельствует о «одиночном ударе» H. + механизм транслокации (т.е. все четыре протона перемещаются через мембрану одновременно). [14] [17] Альтернативные теории предполагают «двухтактный механизм», при котором каждый этап восстановления ( семихинон и убихинол ) приводит к попаданию двух протонов в межмембранное пространство. [18] [19]
Образующийся убихинол, локализованный в мембранном домене, взаимодействует с отрицательно заряженными остатками в плече мембраны, стабилизируя конформационные изменения. [10] механизм Антипортерный (Na + /ЧАС + swap) было предложено с использованием доказательств консервативности остатков Asp в мембранном ответвлении. [20] Присутствие остатков Lys, Glu и His обеспечивает гейтирование протонов (протонирование с последующим событием депротонирования через мембрану), управляемое pK a остатков. [10]
Состав и структура
[ редактировать ]НАДН:убихиноноксидоредуктаза — самый крупный из дыхательных комплексов. У млекопитающих фермент содержит 44 отдельных водорастворимых периферических мембранных белка, которые прикреплены к составным компонентам мембраны. Особое функциональное значение имеют флавина простетическая группа (ФМН) и восемь железо-серных кластеров (FeS). Из 44 субъединиц семь кодируются митохондриальным геномом . [21] [22] [23]
Структура имеет форму буквы «L» с длинным мембранным доменом (около 60 трансмембранных спиралей) и гидрофильным (или периферическим) доменом, который включает все известные окислительно-восстановительные центры и сайт связывания НАДН. [24] Все тринадцать белков E. coli , которые содержат НАДН-дегидрогеназу I, кодируются опероном nuo и гомологичны субъединицам митохондриального комплекса I. Каждая антипортер-подобная субъединица NuoL/M/N содержит по 14 консервативных трансмембранных (ТМ) спиралей. Два из них прерывисты, но субъединица NuoL содержит амфипатическую α-спираль длиной 110 Å, охватывающую всю длину домена. Субъединица NuoL связана с Na. + / ч + антипортеры TC# 2.A.63.1.1 (PhaA и PhaD).
Три из консервативных мембраносвязанных субъединиц НАДН-дегидрогеназы связаны друг с другом и с антипортерами протонов натрия Mrp. Структурный анализ двух прокариотических комплексов показал, что каждая из трех субъединиц содержит четырнадцать трансмембранных спиралей, которые перекрываются в структурном выравнивании: перемещение трех протонов может координироваться соединяющей их боковой спиралью. [25]
Комплекс I содержит карман для связывания убихинона на границе раздела субъединиц 49 кДа и PSST. Рядом с кластером железо-сера N2, предполагаемым непосредственным донором электронов для убихинона, высококонсервативный тирозин представляет собой критический элемент сайта восстановления хинона. Возможный путь хинонового обмена ведет от кластера N2 к N-концевому бета-листу субъединицы 49 кДа. [26] Секвенированы все 45 субъединиц бычьего NDHI. [27] [28] Каждый комплекс содержит нековалентно связанный ФМН, кофермент Q и несколько железо-серных центров. Бактериальные NDH имеют 8-9 железо-серных центров.
В недавнем исследовании использовались спектры электронного парамагнитного резонанса (ЭПР) и двойного электрон-электронного резонанса (DEER) для определения пути переноса электронов через комплексы железо-сера, которые расположены в гидрофильном домене. Семь из этих кластеров образуют цепь от сайтов связывания флавина к хинону; восьмой кластер расположен по другую сторону от флавина, функция его неизвестна. Результаты ЭПР и ДЭЭР предполагают переменный или «американский» профиль потенциальной энергии для переноса электронов между активными центрами и вдоль кластеров железо-сера, который может оптимизировать скорость перемещения электронов и обеспечить эффективное преобразование энергии в комплексе I. [29]
| # | Человеческая / бычья субъединица | Человеческий белок | Описание белка ( UniProt ) | Семейство Pfam с человеческим белком | |
|---|---|---|---|---|---|
| Основные субъединицы а | |||||
| 1 | НДУФС7/ПССТ/НУКМ | NDUS7_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 7, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF01058 | |
| 2 | НДУФС8 / ПУШКА / НУИМ | NDUS8_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 8, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF12838 | |
| 3 | НДУФВ2/24кД/НУХМ с | НДУВ2_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] флавопротеин 2, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF01257 | |
| 4 | НДУФС3/30кД/НУГМ | NDUS3_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 3, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF00329 | |
| 5 | НДУФС2/49кД/НУЦМ | NDUS2_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 2, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF00346 | |
| 6 | НДУФВ1/51кД/НУБМ | НДУВ1_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] флавопротеин 1, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF01512 | |
| 7 | НДУФС1/75кД/НУАМ | NDUS1_HUMAN | Субъединица НАДН-убихиноноксидоредуктаза 75 кДа, митохондриальная EC 1.6.5.3 EC 1.6.99.3 | Пфам PF00384 | |
| 8 | НД1/НУ1М | NU1M_HUMAN | НАДН-убихинон оксидоредуктаза цепь 1 EC 1.6.5.3 | Пфам PF00146 | |
| 9 | НД2/НУ2М | NU2M_HUMAN | НАДН-убихиноноксидоредуктаза цепь 2 EC 1.6.5.3 | Пфам ПФ00361 , Пфам ПФ06444 | |
| 10 | НД3/НУ3М | NU3M_HUMAN | НАДН-убихинон оксидоредуктаза цепь 3 EC 1.6.5.3 | Пфам PF00507 | |
| 11 | НД4 / НУ4М | NU4M_HUMAN | НАДН-убихинон оксидоредуктаза цепь 4 EC 1.6.5.3 | Пфам ПФ01059 , Пфам ПФ00361 | |
| 12 | ND4L / НУЛМ | NU4LM_HUMAN | НАДН-убихиноноксидоредуктазная цепь 4L EC 1.6.5.3 | Пфам PF00420 | |
| 13 | НД5/НУ5М | NU5M_HUMAN | НАДН-убихиноноксидоредуктаза цепь 5 EC 1.6.5.3 | Пфам ПФ00361 , Пфам ПФ06455 , Пфам ПФ00662 | |
| 14 | НД6 / НУ6М | NU6M_HUMAN | НАДН-убихиноноксидоредуктаза цепь 6 EC 1.6.5.3 | Пфам PF00499 | |
| Основные вспомогательные субъединицы б | |||||
| 15 | НДУФС6/13А | NDUS6_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 6, митохондриальный | Пфам PF10276 | |
| 16 | НДУФА12/Б17.2 | НДУАК_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 12 субкомплекса | Пфам PF05071 | |
| 17 | НДУФС4/АКДК | NDUS4_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 4, митохондриальный | Пфам PF04800 | |
| 18 | НДУФА9/39кДа | NDUA9_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-подкомплекс, субъединица 9, митохондриальная | Пфам PF01370 | |
| 19 | НДУФАБ1/АКПМ | ACPM_HUMAN | Ацильный белок-переносчик, митохондриальный | Пфам PF00550 | |
| 20 | НДУФА2/В8 | NDUA2_HUMAN | НАДН-дегидрогеназа [убихинон] 1 субъединица альфа-субъединицы 2 | Пфам PF05047 | |
| 21 | НДУФА1 / МФВЕ | NDUA1_HUMAN | НАДН-дегидрогеназа [убихинон] 1 субъединица альфа-субъединицы 1 | Пфам PF15879 | |
| 22 | НДУФБ3/Б12 | NDUB3_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица 3 субкомплекса | Пфам PF08122 | |
| 23 | НДУФА5/АБ13 | NDUA5_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица субкомплекса 5 | Пфам PF04716 | |
| 24 | НДУФА6/В14 | NDUA6_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 6 субкомплекса | Пфам PF05347 | |
| 25 | НДУФА11/Б14.7 | НДУАБ_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 11 субкомплекса | Пфам PF02466 | |
| 26 | НДУФБ11/ЭССС | НДУББ_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 11, митохондриальная | Пфам PF10183 | |
| 27 | НДУФС5/ПФФД | NDUS5_HUMAN | НАДН-дегидрогеназа [убихинон] железо-серный белок 5 | Пфам PF10200 | |
| 28 | НДУФБ4/Б15 | NDUB4_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 4 | Пфам PF07225 | |
| 29 | НДУФА13/А13 | НДУАД_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 13 субкомплекса | Пфам PF06212 | |
| 30 | НДУФБ7/Б18 | NDUB7_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 7 | Пфам PF05676 | |
| 31 | НДУФА8 / ПГИВ | NDUA8_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 8 субкомплекса | Пфам PF06747 | |
| 32 | НДУФБ9/Б22 | NDUB9_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 9 | Пфам PF05347 | |
| 33 | НДУФБ10/ПДСВ | НДУБА_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 10 | Пфам PF10249 | |
| 34 | НДУФБ8 / АШИ | NDUB8_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 8, митохондриальная | Пфам PF05821 | |
| 35 | НДУФК2/Б14.5Б | NDUC2_HUMAN | НАДН-дегидрогеназа [убихинон] 1 субъединица C2 | Пфам PF06374 | |
| 36 | НДУФБ2/АГГГ | NDUB2_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 2, митохондриальная | Пфам PF14813 | |
| 37 | НДУФА7/Б14.5А | NDUA7_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 7 субкомплекса | Пфам PF07347 | |
| 38 | НДУФА3/В9 | NDUA3_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 3 субкомплекса | Пфам PF14987 | |
| 39 | NDUFA4 / MLRQ в, г | NDUA4_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 4 субкомплекса | Пфам PF06522 | |
| 40 | НДУФБ5/СГДХ | NDUB5_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 5, митохондриальная | Пфам PF09781 | |
| 41 | НДУФБ1/МНЛЛ | NDUB1_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 1 | Пфам PF08040 | |
| 42 | NDUFC1 / KFYI | NDUC1_HUMAN | НАДН-дегидрогеназа [убихинон] 1 субъединица C1, митохондриальная | Пфам PF15088 | |
| 43 | НДУФА10/42кД | НДУАА_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 альфа-подкомплекс, субъединица 10, митохондриальная | Пфам PF01712 | |
| 44 | НДУФА4L2 | NUA4L_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субъединица 4-подобного субкомплекса 2 | Пфам PF15880 | |
| 45 | НДУФВ3 | НДУВ3_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] флавопротеин 3, 10 кДа | - | |
| 46 | НДУФБ6 | NDUB6_HUMAN | НАДН-дегидрогеназа [убихинон] 1 бета-субъединица субъединицы 6 | Пфам PF09782 | |
| Белки фактора сборки [31] | |||||
| 47 | НДУФАФ1 с | ЦРУ30_ЧЕЛОВЕК | НАДН-дегидрогеназа [убихинон] 1 альфа-субкомплекс, фактор сборки 1 | Пфам PF08547 | |
| 48 | НДУФАФ2 | MIMIT_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субкомплекс, фактор сборки 2 | Пфам PF05071 | |
| 49 | НДУФАФ3 | NDUF3_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субкомплекс, фактор сборки 3 | Пфам PF05071 | |
| 50 | НДУФАФ4 | NDUF4_HUMAN | НАДН-дегидрогеназа [убихинон] 1 альфа-субкомплекс, фактор сборки 4 | Пфам PF06784 | |
Примечания:
- а Встречается у всех видов, кроме грибов.
- б Может присутствовать или отсутствовать у любого вида
- с Встречается в таких видах грибов, как Schizosaccharomyces pombe.
- д Недавние исследования показали, что NDUFA4 является субъединицей комплекса IV , а не комплекса I. [34]
Ингибиторы
[ редактировать ]Буллатацин ( ацетогенин, обнаруженный в плодах Asimina triloba ) является наиболее мощным известным ингибитором НАДН-дегидрогеназы (убихинон) ( IC 50 = 1,2 нМ, сильнее, чем ротенон). [35] Самый известный ингибитор комплекса I — ротенон (обычно используемый в качестве органического пестицида). Ротенон и ротеноиды представляют собой изофлавоноиды, встречающиеся в нескольких родах тропических растений, таких как Antonia ( Loganiaceae ), Derris и Lonchocarpus ( Faboideae , Fabaceae ). Сообщения о том, что коренные жители Французской Гвианы использовали ротенонсодержащие растения для ловли рыбы из-за ихтиотоксического эффекта, появились еще в 17 веке. [36] Ротенон связывается с сайтом связывания убихинона комплекса I, а также с пиерицидином А , другим мощным ингибитором с близким структурным гомологом убихинону.
Ацетогенины Annonaceae . являются еще более мощными ингибиторами комплекса I. Они перекрестно сшиваются с субъединицей ND2, что позволяет предположить, что ND2 необходим для связывания хинонов [37] Роллиниастатин-2, ацетогенин, является первым обнаруженным ингибитором комплекса I, который не имеет того же сайта связывания, что и ротенон. [38]
Несмотря на более чем 50-летние исследования комплекса I, ингибиторов, блокирующих поток электронов внутри фермента, обнаружено не было. Гидрофобные ингибиторы, такие как ротенон или пиерицидин, скорее всего, нарушают перенос электронов между концевым кластером N2 FeS и убихиноном. Было показано, что длительное системное ингибирование комплекса I ротеноном может вызывать селективную дегенерацию дофаминергических нейронов. [39]
Комплекс I также блокируется аденозиндифосфатрибозой – обратимым конкурентным ингибитором окисления НАДН – путем связывания с ферментом в месте связывания нуклеотидов. [40] И гидрофильные НАДН, и гидрофобные аналоги убихинона действуют в начале и в конце пути внутреннего транспорта электронов соответственно.
Было показано, что противодиабетический препарат метформин вызывает легкое и временное ингибирование комплекса I дыхательной цепи митохондрий, и это ингибирование, по-видимому, играет ключевую роль в механизме его действия. [41]
Ингибирование комплекса I связано с гепатотоксичностью , связанной с различными лекарственными средствами, например флутамидом и нефазодоном . [42] Кроме того, было показано, что ингибирование комплекса I запускает НАД. + -независимый катаболизм глюкозы . [43]
Активный/неактивный переход
[ редактировать ]Каталитические свойства эукариотического комплекса I непросты. В любом препарате фермента существуют две каталитически и структурно различные формы: одна — полностью компетентная, так называемая «активная» А-форма, а другая — каталитически молчащая, «спящая», «неактивная» D-форма. После воздействия бездействующего фермента на повышенные, но физиологические температуры (>30°С) в отсутствие субстрата фермент переходит в D-форму. Эта форма каталитически некомпетентна, но может активироваться в результате медленной реакции (k~4 мин. −1 ) окисления НАДН с последующим восстановлением убихинона. После одного или нескольких оборотов фермент становится активным и может катализировать физиологическую реакцию НАДН:убихинон с гораздо более высокой скоростью (k ~ 10 4 мин −1 ). В присутствии двухвалентных катионов (Mg 2+ , Как 2+ ), или при щелочном pH активация занимает гораздо больше времени.
Высокая энергия активации (270 кДж/моль) процесса дезактивации указывает на возникновение серьезных конформационных изменений в организации комплекса I. Однако до сих пор единственным конформационным различием, наблюдаемым между этими двумя формами, является количество цистеина. подвергающихся воздействию остатков на поверхности фермента. Обработка D-формы комплекса I сульфгидрильными реагентами N-этилмалеимидом или ДТНБ необратимо блокирует критические остатки цистеина, отменяя способность фермента реагировать на активацию и тем самым необратимо инактивируя его. А-форма комплекса I нечувствительна к сульфгидрильным реагентам. [44] [45]
Было обнаружено, что эти конформационные изменения могут иметь очень важное физиологическое значение. Неактивная, но не активная форма комплекса I поддавалась ингибированию нитрозотиолами и пероксинитритом . [46] Вероятно, переход от активной формы комплекса I к неактивной происходит при патологических состояниях, когда оборот фермента ограничен при физиологических температурах, например при гипоксии , ишемии. [47] [48] или когда соотношение оксид азота :кислород в тканях увеличивается (т.е. метаболическая гипоксия). [49]
Производство супероксида
[ редактировать ]Недавние исследования показывают, что комплекс I является мощным источником активных форм кислорода . [50] Комплекс I может производить супероксид (а также перекись водорода ) как минимум двумя разными путями. Во время прямого переноса электронов образуется лишь очень небольшое количество супероксида (вероятно, менее 0,1% от общего потока электронов). [50] [51] [52]
Во время обратного переноса электронов комплекс I может быть наиболее важным местом производства супероксида в митохондриях, при этом около 3-4% электронов направляются на образование супероксида. [53] Обратный перенос электронов - процесс, посредством которого электроны из восстановленного пула убихинола (поставляемого сукцинатдегидрогеназой , глицерин-3-фосфатдегидрогеназой , электронопереносящим флавопротеином или дигидрооротатдегидрогеназой в митохондриях млекопитающих) проходят через комплекс I для восстановления НАД. + НАДН, управляемый электрическим потенциалом внутренней мембраны митохондрий. Хотя точно не известно, при каких патологических условиях обратный перенос электронов будет происходить in vivo, эксперименты in vitro показывают, что этот процесс может быть очень мощным источником супероксида, когда концентрации сукцината высоки, а концентрации оксалоацетата или малата низки. [54] Это может произойти при ишемии тканей, когда блокируется доставка кислорода. [55]
Супероксид — это активная форма кислорода, которая способствует клеточному окислительному стрессу и связана с нервно-мышечными заболеваниями и старением. [56] НАДН-дегидрогеназа производит супероксид путем переноса одного электрона от FMNH 2 (или полувосстановленного флавина) к кислороду (O 2 ). Оставшийся радикал-флавин нестабилен и переносит оставшийся электрон к железо-серным центрам. Это соотношение НАДН к НАД. + что определяет скорость образования супероксида. [57] [58]
Патология
[ редактировать ]Мутации в субъединицах комплекса I могут вызывать митохондриальные заболевания , в том числе синдром Ли . Точечные мутации в различных субъединицах комплекса I, происходящих из митохондриальной ДНК ( мтДНК ), также могут приводить к наследственной оптической нейропатии Лебера . Имеются некоторые свидетельства того, что дефекты комплекса I могут играть роль в этиологии болезни Паркинсона , возможно, из-за активных форм кислорода (комплекс Я могу, как и комплекс III , отдавать электроны кислороду, образуя высокотоксичный супероксид ).
Хотя точная этиология болезни Паркинсона неясна, вполне вероятно, что большую роль может играть митохондриальная дисфункция, наряду с ингибированием протеасом и токсинами окружающей среды. Фактически было показано, что ингибирование комплекса I вызывает выработку пероксидов и снижение активности протеасом , что может привести к болезни Паркинсона. [59] Кроме того, Эстевес и др. (2010) обнаружили, что клеточные линии с болезнью Паркинсона демонстрируют повышенную утечку протонов в комплексе I, что приводит к снижению максимальной дыхательной способности. [60]
Ишемия/реперфузионное повреждение головного мозга опосредовано нарушением комплекса I. [61] Недавно было обнаружено, что кислородное голодание приводит к состояниям, при которых митохондриальный комплекс I теряет свой природный кофактор — флавинмононуклеотид (ФМН) и становится неактивным. [62] [63] В присутствии кислорода фермент катализирует физиологическую реакцию окисления НАДН убихиноном, доставляя электроны ниже дыхательной цепи (комплексы III и IV). Ишемия приводит к резкому повышению уровня сукцината . В присутствии сукцината митохондрии катализируют обратный перенос электронов , так что часть электронов сукцината направляется вверх по ходу к FMN комплекса I. Обратный перенос электронов приводит к восстановлению комплекса I FMN, [53] повышенное образование АФК с последующей потерей восстановленного кофактора (FMNH 2 ) и нарушением производства энергии митохондриями. Утрату FMN в результате повреждения комплекса I и I/R можно облегчить введением предшественника FMN, рибофлавина. [63]
Недавние исследования изучили другие роли активности комплекса I в мозге. Андреацца и др. (2010) обнаружили, что уровень активности комплекса I значительно снижается у пациентов с биполярным расстройством, но не у пациентов с депрессией или шизофренией. Они обнаружили, что у пациентов с биполярным расстройством наблюдается повышенное окисление и нитрование белков в префронтальной коре. Эти результаты позволяют предположить, что будущие исследования должны быть нацелены на комплекс I для потенциальных терапевтических исследований биполярного расстройства. [64] Аналогичным образом, Моран и др. (2010) обнаружили, что у пациентов с тяжелым дефицитом комплекса I наблюдается снижение скорости потребления кислорода и замедление темпов роста. Однако они обнаружили, что мутации в разных генах комплекса I приводят к разным фенотипам, тем самым объясняя вариации патофизиологических проявлений дефицита комплекса I. [65]
Воздействие пестицидов также может ингибировать комплекс I и вызывать симптомы заболевания. Например, было показано, что хроническое воздействие низких концентраций дихлофоса, фосфорорганического соединения, используемого в качестве пестицида, вызывает дисфункцию печени. Это происходит потому, что дихлофос изменяет уровни активности комплексов I и II, что приводит к снижению активности переноса электронов в митохондриях и снижению синтеза АТФ. [66]
В хлоропластах
[ редактировать ]Комплекс НАДН-дегидрогеназы, перекачивающий протоны и использующий убихинон, гомологичный комплексу I, обнаружен в геномах хлоропластов большинства наземных растений под названием ndh . Этот комплекс унаследован от первоначального симбиоза цианобактерий, но утрачен у большинства эукариотических водорослей, некоторых голосеменных ( Pinus и гнетофитов ) и некоторых очень молодых линий покрытосеменных . Назначение этого комплекса изначально загадочно, поскольку хлоропласты не участвуют в дыхании, но теперь известно, что ndh служит для поддержания фотосинтеза в стрессовых ситуациях. Это делает его, по крайней мере, частично необязательным в благоприятных условиях. Очевидно, что линии покрытосеменных без ndh не живут долго с раннего возраста, но выживают на суше без ndh . неизвестно, как голосеменные так долго [67]
Гены
[ редактировать ]Ниже приводится список генов человека, кодирующих компоненты комплекса I:
- НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс
- NDUFA1 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 1, 7,5 кДа
- NDUFA2 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 2, 8 кДа
- NDUFA3 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 3, 9 кДа
- NDUFA4 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 4, 9 кДа - недавно описано как часть комплекса IV. [34]
- NDUFA4L - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 4-подобный
- NDUFA4L2 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 4-подобный 2
- NDUFA5 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 5, 13 кДа
- NDUFA6 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 6, 14 кДа
- NDUFA7 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 7, 14,5 кДа
- NDUFA8 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 8, 19 кДа
- NDUFA9 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 9, 39 кДа
- NDUFA10 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, 10, 42 кДа
- NDUFA11 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 11, 14,7 кДа
- NDUFA12 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 12
- NDUFA13 - НАДН-дегидрогеназа (убихинон) 1 альфа-субкомплекс, 13
- NDUFAB1 - НАДН-дегидрогеназа (убихинон) 1, альфа/бета-субкомплекс, 1, 8 кДа
- NDUFAF1 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, фактор сборки 1
- NDUFAF2 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, фактор сборки 2
- NDUFAF3 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, фактор сборки 3
- NDUFAF4 - субкомплекс НАДН-дегидрогеназы (убихинон) 1 альфа, фактор сборки 4
- НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс
- NDUFB1 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 1,7 кДа
- NDUFB2 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 2, 8 кДа
- NDUFB3 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 3, 12 кДа
- NDUFB4 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 4, 15 кДа
- NDUFB5 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 5, 16 кДа
- NDUFB6 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 6, 17 кДа
- NDUFB7 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 7, 18 кДа
- NDUFB8 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 8, 19 кДа
- NDUFB9 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 9, 22 кДа
- NDUFB10 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 10, 22 кДа
- NDUFB11 - НАДН-дегидрогеназа (убихинон) 1 бета-субкомплекс, 11, 17,3 кДа
- НАДН-дегидрогеназа (убихинон) 1, субкомплекс неизвестен
- НАДН-дегидрогеназа (убихинон) белок Fe-S
- NDUFS1 - НАДН-дегидрогеназа (убихинон), белок Fe-S 1, 75 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS2 - НАДН-дегидрогеназа (убихинон), белок Fe-S 2, 49 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS3 - НАДН-дегидрогеназа (убихинон), белок Fe-S 3, 30 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS4 - НАДН-дегидрогеназа (убихинон), белок Fe-S 4, 18 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS5 - НАДН-дегидрогеназа (убихинон), белок Fe-S 5, 15 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS6 - НАДН-дегидрогеназа (убихинон), белок Fe-S 6, 13 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS7 - НАДН-дегидрогеназа (убихинон), белок Fe-S 7, 20 кДа (НАДН-коэнзим Q-редуктаза)
- NDUFS8 - НАДН-дегидрогеназа (убихинон), белок Fe-S 8, 23 кДа (НАДН-коэнзим Q-редуктаза)
- НАДН-дегидрогеназа (убихинон) флавопротеин 1
- митохондриально кодируемая субъединица НАДН-дегидрогеназы
- MT-ND1 - субъединица 1 НАДН-дегидрогеназы, кодируемая митохондриями.
- MT-ND2 - субъединица 2 НАДН-дегидрогеназы, кодируемая митохондриями.
- MT-ND3 - субъединица 3 НАДН-дегидрогеназы, кодируемая митохондриями.
- MT-ND4 - митохондриально кодируемая субъединица 4 НАДН-дегидрогеназы
- MT-ND4L - митохондриально кодируемая субъединица 4L НАДН-дегидрогеназы
- MT-ND5 - субъединица 5 НАДН-дегидрогеназы, кодируемая митохондриями.
- MT-ND6 - субъединица 6 НАДН-дегидрогеназы, кодируемая митохондриями.
Ссылки
[ редактировать ]- ^ Jump up to: а б Берг Дж., Тимочко Дж., Страйер Л. (2006). Биохимия (6-е изд.). Нью-Йорк: WH Freeman & Company. стр. 509–513.
- ^ Брандт У (2006). «Энергопреобразование НАДН:хиноноксидоредуктаза (комплекс I)». Ежегодный обзор биохимии . 75 : 69–92. doi : 10.1146/annurev.biochem.75.103004.142539 . ПМИД 16756485 .
- ^ Викстрем М. (апрель 1984 г.). «Два протона выкачиваются из митохондриального матрикса на один электрон, передаваемый между НАДН и убихиноном» . Письма ФЭБС . 169 (2): 300–4. дои : 10.1016/0014-5793(84)80338-5 . ПМИД 6325245 .
- ^ Галкин А., Дрёзе С., Брандт У. (декабрь 2006 г.). «Стехиометрия протонной накачки очищенного митохондриального комплекса, который я восстановил в протеолипосомы» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1757 (12): 1575–81. дои : 10.1016/j.bbabio.2006.10.001 . ПМИД 17094937 .
- ^ Галкин А.С., Гривенникова В.Г., Виноградов А.Д. (май 1999 г.). «-->H+/2e-стехиометрия в реакциях НАДН-хинонредуктазы, катализируемых субмитохондриальными частицами бычьего сердца» . Письма ФЭБС . 451 (2): 157–61. дои : 10.1016/s0014-5793(99)00575-x . ПМИД 10371157 . S2CID 2337382 .
- ^ Jump up to: а б Батиста А.П., Перейра М.М. (март 2011 г.). «Влияние натрия на трансдукцию энергии комплексами I из Escherichia coli и Paracoccus denitrificans» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1807 (3): 286–92. дои : 10.1016/j.bbabio.2010.12.008 . ПМИД 21172303 .
- ^ Гривенникова В.Г., Котляр А.Б., Карлинер Дж.С., Чеккини Г., Виноградов А.Д. (сентябрь 2007 г.). «Редокс-зависимое изменение сродства нуклеотидов к активному центру комплекса I млекопитающих» . Биохимия . 46 (38): 10971–8. дои : 10.1021/bi7009822 . ПМЦ 2258335 . ПМИД 17760425 .
- ^ Чомова М., Ракай П. (март 2010 г.). «Митохондриальный комплекс I в сети известных и неизвестных фактов» . Общая физиология и биофизика . 29 (1): 3–11. дои : 10.4149/gpb_2010_01_3 . ПМИД 20371875 .
- ^ Петрусса Э., Бертолини А., Касоло В., Крайнакова Дж., Макри Ф., Вианелло А. (декабрь 2009 г.). «Митохондриальная биоэнергетика связана с проявлением запрограммированной гибели клеток во время соматического эмбриогенеза Abies alba». Планта . 231 (1): 93–107. дои : 10.1007/s00425-009-1028-x . ПМИД 19834734 . S2CID 25828432 .
- ^ Jump up to: а б с д и Сазанов Л.А. (июнь 2015). «Гигантский молекулярный протонный насос: структура и механизм дыхательного комплекса I». Обзоры природы. Молекулярно-клеточная биология . 16 (6): 375–88. дои : 10.1038/nrm3997 . ПМИД 25991374 . S2CID 31633494 .
- ^ Voet DJ, Voet GJ, Пратт CW (2008). «Глава 18, Синтез митохондриального АТФ». Принципы биохимии, 3-е издание . Уайли. п. 608. ИСБН 978-0-470-23396-2 .
- ^ Ониши Т. (май 1998 г.). «Железосерные кластеры/семихиноны в комплексе I» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1364 (2): 186–206. дои : 10.1016/s0005-2728(98)00027-9 . ПМИД 9593887 .
- ^ Бриджес HR, Билл Э., Херст Дж. (январь 2012 г.). «Мессбауэровская спектроскопия на дыхательном комплексе I: кластерный ансамбль железо-сера в НАДН-восстановленном ферменте частично окислен» . Биохимия . 51 (1): 149–58. дои : 10.1021/bi201644x . ПМК 3254188 . ПМИД 22122402 .
- ^ Jump up to: а б Ефремов Р.Г., Сазанов Л.А. (октябрь 2012 г.). «Механизм сцепления дыхательного комплекса I - структурный и эволюционный взгляд» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1817 (10): 1785–95. дои : 10.1016/j.bbabio.2012.02.015 . ПМИД 22386882 .
- ^ Треберг-младший, Куинлан С.Л., Брэнд, доктор медицинских наук (август 2011 г.). «Доказательства наличия двух мест производства супероксида митохондриальной НАДН-убихинон оксидоредуктазой (комплекс I)» . Журнал биологической химии . 286 (31): 27103–10. дои : 10.1074/jbc.M111.252502 . ПМК 3149303 . ПМИД 21659507 .
- ^ Беррисфорд Дж.М., Сазанов Л.А. (октябрь 2009 г.). «Структурные основы механизма I дыхательного комплекса» . Журнал биологической химии . 284 (43): 29773–83. дои : 10.1074/jbc.m109.032144 . ПМЦ 2785608 . ПМИД 19635800 .
- ^ Баранова Е.А., Морган Д.Д., Сазанов Л.А. (август 2007 г.). «Анализ одиночных частиц подтверждает дистальное расположение субъединиц NuoL и NuoM в комплексе I Escherichia coli». Журнал структурной биологии . 159 (2): 238–42. дои : 10.1016/j.jsb.2007.01.009 . ПМИД 17360196 .
- ^ Брандт У (октябрь 2011 г.). «Механизм стабилизации-изменения с двумя состояниями для комплекса протонной накачки I» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1807 (10): 1364–9. дои : 10.1016/j.bbabio.2011.04.006 . ПМИД 21565159 .
- ^ Цикерманн В., Вирт К., Насири Х., Зигмунд К., Швальбе Х., Ханте К., Брандт Ю. (январь 2015 г.). «Структурная биология. Механистическое понимание кристаллической структуры митохондриального комплекса I» (PDF) . Наука . 347 (6217): 44–9. дои : 10.1126/science.1259859 . ПМИД 25554780 . S2CID 23582849 .
- ^ Хант С., Скрапанти Э., Вентури М., Римон А., Падан Э., Мишель Х. (июнь 2005 г.). «Структура антипортера Na +/H + и понимание механизма действия и регуляции pH». Природа . 435 (7046): 1197–202. Бибкод : 2005Natur.435.1197H . дои : 10.1038/nature03692 . ПМИД 15988517 . S2CID 4372674 .
- ^ Фут JG, Фут D (2004). Биохимия (3-е изд.). Нью-Йорк: Дж. Уайли и сыновья. стр. 813 –826. ISBN 0-471-19350-Х .
- ^ Кэрролл Дж., Фернли И.М., Скехел Дж.М., Шеннон Р.Дж., Херст Дж., Уокер Дж.Э. (октябрь 2006 г.). «Бычий комплекс I представляет собой комплекс из 45 различных субъединиц» . Журнал биологической химии . 281 (43): 32724–7. дои : 10.1074/jbc.M607135200 . ПМИД 16950771 .
- ^ Бальса Э., Марко Р., Пералес-Клементе Э., Шклярчик Р., Кальво Э., Ландазури М.О., Энрикес Х.А. (сентябрь 2012 г.). «NDUFA4 представляет собой субъединицу комплекса IV цепи переноса электронов млекопитающих» . Клеточный метаболизм . 16 (3): 378–86. дои : 10.1016/j.cmet.2012.07.015 . ПМИД 22902835 .
- ^ Сазанов Л.А. , Хинчлифф П. (март 2006 г.). «Структура гидрофильного домена дыхательного комплекса I Thermus thermophilus» . Наука . 311 (5766): 1430–6. Бибкод : 2006Sci...311.1430S . дои : 10.1126/science.1123809 . ПМИД 16469879 . S2CID 1892332 .
- ^ Ефремов Р.Г., Барадаран Р., Сазанов Л.А. (май 2010 г.). «Архитектура дыхательного комплекса I». Природа . 465 (7297): 441–5. Бибкод : 2010Natur.465..441E . дои : 10.1038/nature09066 . ПМИД 20505720 . S2CID 4372778 .
- ^ Точилеску М.А., Цикерманн В., Цвикер К., Брандт У. (декабрь 2010 г.). «Связывание и восстановление хинонов дыхательным комплексом I» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1797 (12): 1883–90. дои : 10.1016/j.bbabio.2010.05.009 . ПМИД 20493164 .
- ^ Кардол П., Ванробэйс Ф., Девриз Б., Ван Беумен Дж., Матань Р.Ф., Ремакл С. (октябрь 2004 г.). «Высший растительный субъединичный состав митохондриального комплекса I Chlamydomonas Reinhardtii: 31 консервативный компонент среди эукариот» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1658 (3): 212–24. дои : 10.1016/j.bbabio.2004.06.001 . ПМИД 15450959 .
- ^ Габальдон Т., Рейни Д., Хуйнен М.А. (май 2005 г.). «Отслеживание эволюции большого белкового комплекса у эукариот НАДН:убихиноноксидоредуктаза (Комплекс I)». Журнал молекулярной биологии . 348 (4): 857–70. дои : 10.1016/j.jmb.2005.02.067 . ПМИД 15843018 .
- ^ Росслер М.М., Кинг М.С., Робинсон А.Дж., Армстронг Ф.А., Хармер Дж., Херст Дж. (февраль 2010 г.). «Прямое отнесение спектров ЭПР к структурно определенным железо-серным кластерам в комплексе I методом двойного электрон-электронного резонанса» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (5): 1930–5. Бибкод : 2010ПНАС..107.1930Р . дои : 10.1073/pnas.0908050107 . ПМК 2808219 . ПМИД 20133838 .
- ^ Кардол П. (ноябрь 2011 г.). «Митохондриальный НАДН:убихиноноксидоредуктаза (комплекс I) у эукариот: высококонсервативный состав субъединиц, выявленный при анализе баз данных белков» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1807 (11): 1390–7. дои : 10.1016/j.bbabio.2011.06.015 . ПМИД 21749854 .
- ^ Огилви И., Кеннауэй Н.Г., Шубридж Э.А. (октябрь 2005 г.). «Молекулярный шаперон сборки митохондриального комплекса I мутирует при прогрессирующей энцефалопатии» . Журнал клинических исследований . 115 (10): 2784–92. дои : 10.1172/JCI26020 . ПМК 1236688 . ПМИД 16200211 .
- ^ Даннинг СиДжей, Маккензи М., Сугиана С., Лазару М., Силке Дж., Коннелли А., Флетчер Дж.М., Кирби Д.М., Торберн Д.Р., Райан М.Т. (июль 2007 г.). «Человеческий CIA30 участвует в ранней сборке митохондриального комплекса I, а мутации в его гене вызывают заболевания» . Журнал ЭМБО . 26 (13): 3227–37. дои : 10.1038/sj.emboj.7601748 . ПМК 1914096 . ПМИД 17557076 .
- ^ Саада А., Фогель Р.О., Хофс С.Дж., ван де Бранд М.А., Весселс Х.Дж., Виллемс П.Х., Венселаар Х., Шааг А., Баргути Ф., Рейш О., Шохат М., Хюйнен М.А., Смейтинк Дж.А., ван де Хеувел Л.П., Нейтманс Л.Г. (июнь) 2009). «Мутации в NDUFAF3 (C3ORF60), кодирующем белок сборки комплекса I, взаимодействующий с NDUFAF4 (C6ORF66), вызывают фатальные неонатальные митохондриальные заболевания» . Американский журнал генетики человека . 84 (6): 718–27. дои : 10.1016/j.ajhg.2009.04.020 . ПМК 2694978 . ПМИД 19463981 .
- ^ Jump up to: а б Бальса, Эдуардо; Марко, Рикардо; Пералес-Клементе, Эстер; Шклярчик, Радек; Кальво, Энрике; Ландазури, Мануэль О.; Энрикес, Хосе Антонио (5 сентября 2012 г.). «NDUFA4 представляет собой субъединицу комплекса IV цепи переноса электронов млекопитающих» . Клеточный метаболизм . 16 (3): 378–386. дои : 10.1016/j.cmet.2012.07.015 . ISSN 1932-7420 . ПМИД 22902835 .
- ^ Миёси Х., Осима М., Симада Х., Акаги Т., Ивамура Х., Маклафлин Дж.Л. (июль 1998 г.). «Основные структурные факторы аннонических ацетогенинов как мощных ингибиторов митохондриального комплекса I» . Биохимия и биофизика Acta (BBA) - Биоэнергетика 1365 (3): 443–52. дои : 10.1016/s0005-2728(98) 00097-8 ПМИД 9711297 .
- ^ Моретти С., Гренан П. (сентябрь 1982 г.). «[Нивре, или ихтиотоксичные растения Французской Гвианы]». Журнал этнофармакологии (на французском языке). 6 (2): 139–60. дои : 10.1016/0378-8741(82)90002-2 . ПМИД 7132401 .
- ^ Накамару-Огисо Э., Хан Х., Мацуно-Яги А., Кейнан Э., Синха С.К., Яги Т., Ониши Т. (март 2010 г.). «Субъединица ND2 метится фотоаффинным аналогом азимицина, мощного ингибитора комплекса I» . Письма ФЭБС . 584 (5): 883–8. дои : 10.1016/j.febslet.2010.01.004 . ПМЦ 2836797 . ПМИД 20074573 .
- ^ Дельи Эспости М., Гелли А., Ратта М., Кортес Д., Эсторнелл Э. (июль 1994 г.). «Природные вещества (ацетогенины) семейства Annonaceae являются мощными ингибиторами митохондриальной НАДН-дегидрогеназы (Комплекс I)» . Биохимический журнал . 301 (Часть 1) (Часть 1): 161–7. дои : 10.1042/bj3010161 . ПМЦ 1137156 . ПМИД 8037664 .
- ^ Ватабе М., Накаки Т. (октябрь 2008 г.). «Ингибитор митохондриального комплекса I ротенон ингибирует и перераспределяет везикулярный переносчик моноаминов 2 посредством нитрования в дофаминергических клетках человека SH-SY5Y». Молекулярная фармакология . 74 (4): 933–40. дои : 10.1124/моль.108.048546 . ПМИД 18599602 . S2CID 1844073 .
- ^ Жарова Т.В., Виноградов А.Д. (июль 1997 г.). «Конкурентное ингибирование митохондриальной НАДН-убихинон-оксидоредуктазы (комплекса I) АДФ-рибозой» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1320 (3): 256–64. дои : 10.1016/S0005-2728(97)00029-7 . ПМИД 9230920 .
- ^ Виолле Б., Гигас Б., Санс Гарсия Н., Леклерк Дж., Форец М., Андреелли Ф. (март 2012 г.). «Клеточные и молекулярные механизмы действия метформина: обзор» . Клиническая наука . 122 (6): 253–70. дои : 10.1042/CS20110386 . ПМК 3398862 . ПМИД 22117616 .
- ^ Наданачива С., Уилл Ю (2011). «Новые данные о митохондриальной токсичности, вызванной лекарствами». Текущий фармацевтический дизайн . 17 (20): 2100–12. дои : 10.2174/138161211796904795 . ПМИД 21718246 .
- ^ Абросимов Роман; Баекен, Мариус В.; Хауф, Сэмюэл; Виттиг, Илька; Хаджиева, Парвана; Перроне, Кармен Э.; Моосманн, Бернд (25 января 2024 г.). «Ингибирование митохондриального комплекса I запускает НАД+-независимое окисление глюкозы посредством последовательного образования НАДФН, «бесполезного» цикла жирных кислот и окисления ФАДН2» . Геронаука . дои : 10.1007/s11357-023-01059-y . ISSN 2509-2723 . ПМЦ 11226580 .
- ^ Бабот М., Берч А., Лабарбута П., Галкин А. (июль 2014 г.). «Характеристика активного/деактивного перехода митохондриального комплекса I» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1837 (7): 1083–92. дои : 10.1016/j.bbabio.2014.02.018 . ПМК 4331042 . ПМИД 24569053 .
- ^ Дрёзе С., Степанова А., Галкин А. (июль 2016 г.). «Ишемический A/D-переход митохондриального комплекса I и его роль в генерации АФК» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1857 (7): 946–57. дои : 10.1016/j.bbabio.2015.12.013 . ПМЦ 4893024 . ПМИД 26777588 .
- ^ Галкин А., Монкада С. (декабрь 2007 г.). «S-нитрозирование митохондриального комплекса I зависит от его структурной конформации» . Журнал биологической химии . 282 (52): 37448–53. дои : 10.1074/jbc.M707543200 . ПМИД 17956863 .
- ^ Ким М., Степанова А., Ниацецкая З., Сосунов С., Арндт С., Мерфи М.П. и др. (август 2018 г.). «Ослабление окислительного повреждения путем воздействия на митохондриальный комплекс I при неонатальном гипоксически-ишемическом повреждении головного мозга» . Свободно-радикальная биология и медицина . 124 : 517–524. doi : 10.1016/j.freeradbiomed.2018.06.040 . ПМК 6389362 . ПМИД 30037775 .
- ^ Степанова А, Конрад С, Герреро-Кастильо С, Манфреди Г, Ваннуччи С, Арнольд С, Галкин А (сентябрь 2019 г.). «Дезактивация митохондриального комплекса I после гипоксии-ишемии в незрелом мозге» . Журнал церебрального кровотока и метаболизма . 39 (9): 1790–1802. дои : 10.1177/0271678X18770331 . ПМК 6727140 . ПМИД 29629602 .
- ^ Монкада С., Ерусалимский Дж.Д. (март 2002 г.). «Модулирует ли оксид азота выработку энергии и апоптоз митохондрий?». Обзоры природы. Молекулярно-клеточная биология . 3 (3): 214–20. дои : 10.1038/nrm762 . ПМИД 11994742 . S2CID 29513174 .
- ^ Jump up to: а б Член парламента Мерфи (январь 2009 г.). «Как митохондрии производят активные формы кислорода» . Биохимический журнал . 417 (1): 1–13. дои : 10.1042/BJ20081386 . ПМК 2605959 . ПМИД 19061483 .
- ^ Хансфорд Р.Г., Хог Б.А., Милдазин В. (февраль 1997 г.). «Зависимость образования H2O2 митохондриями сердца крысы от наличия субстрата и возраста донора». Журнал биоэнергетики и биомембран . 29 (1): 89–95. дои : 10.1023/A:1022420007908 . ПМИД 9067806 . S2CID 7501110 .
- ^ Степанова А, Конрад С, Манфреди Г, Спрингетт Р, Тен В, Галкин А (март 2019 г.). «Зависимость производства активных форм кислорода митохондриями мозга от уровня кислорода является линейной, за исключением случаев, когда оно ингибируется антимицином А» . Журнал нейрохимии . 148 (6): 731–745. дои : 10.1111/jnc.14654 . ПМК 7086484 . ПМИД 30582748 .
- ^ Jump up to: а б Степанова А, Каль А, Конрад С, Тен В, Старков А.С., Галкин А (декабрь 2017 г.). «Обратный перенос электронов приводит к потере флавина из митохондриального комплекса I: потенциальный механизм ишемического реперфузионного повреждения головного мозга» . Журнал церебрального кровотока и метаболизма . 37 (12): 3649–3658. дои : 10.1177/0271678X17730242 . ПМЦ 5718331 . ПМИД 28914132 .
- ^ Мюллер Ф.Л., Лю Ю., Абдул-Гани М.А., Лустгартен М.С., Бхаттачарья А., Джанг Ю.К., Ван Реммен Х. (январь 2008 г.). «Высокие темпы продукции супероксида в митохондриях скелетных мышц, дышащих как на комплексных I-, так и на комплексных II-связанных субстратах». Биохимический журнал . 409 (2): 491–9. дои : 10.1042/BJ20071162 . ПМИД 17916065 .
- ^ Сахни П.В., Чжан Дж., Сосунов С., Галкин А., Ниацецкая З., Старков А. и др. (февраль 2018 г.). «Метаболиты цикла Кребса и преимущественное окисление сукцината после неонатального гипоксически-ишемического повреждения головного мозга у мышей» . Педиатрические исследования . 83 (2): 491–497. дои : 10.1038/пр.2017.277 . ПМК 5866163 . ПМИД 29211056 .
- ^ Эстерхази Д., Кинг М.С., Яковлев Г., Херст Дж. (март 2008 г.). «Продуцирование активных форм кислорода комплексом I (НАДН:убихиноноксидоредуктаза) из Escherichia coli и сравнение с ферментом из митохондрий» . Биохимия . 47 (12): 3964–71. дои : 10.1021/bi702243b . ПМИД 18307315 .
- ^ Куссмаул Л., Херст Дж. (май 2006 г.). «Механизм производства супероксида НАДН:убихиноноксидоредуктазой (комплекс I) из митохондрий бычьего сердца» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (20): 7607–12. Бибкод : 2006PNAS..103.7607K . дои : 10.1073/pnas.0510977103 . ПМЦ 1472492 . ПМИД 16682634 .
- ^ Галкин А, Брандт У (август 2005 г.). «Образование супероксидного радикала чистым комплексом I (НАДН:убихиноноксидоредуктаза) из Yarrowia lipolytica» . Журнал биологической химии . 280 (34): 30129–35. дои : 10.1074/jbc.M504709200 . ПМИД 15985426 .
- ^ Чоу А.П., Ли С., Фицморис А.Г., Бронштейн Дж.М. (август 2010 г.). «Механизмы ингибирования протеасом, индуцированного ротеноном» . Нейротоксикология . 31 (4): 367–72. дои : 10.1016/j.neuro.2010.04.006 . ПМЦ 2885979 . ПМИД 20417232 .
- ^ Эстевес А.Р., Лу Дж., Родова М., Оньянго И., Лези Э., Дубинский Р., Лайонс К.Е., Пава Р., Бернс Дж.М., Кардосо С.М., Свердлов Р.Х. (май 2010 г.). «Митохондриальное дыхание и связанные с дыханием белки в клеточных линиях, созданных в результате митохондриального переноса Паркинсона» . Журнал нейрохимии . 113 (3): 674–82. дои : 10.1111/j.1471-4159.2010.06631.x . ПМИД 20132468 .
- ^ Галкин А (ноябрь 2019). «Ишемия/реперфузия головного мозга и повреждение митохондриального комплекса I». Биохимия. Биохимия . 84 (11): 1411–1423. дои : 10.1134/S0006297919110154 . ПМИД 31760927 . S2CID 207990089 .
- ^ Каль А., Степанова А., Конрад С., Андерсон С., Манфреди Г., Чжоу П. и др. (май 2018 г.). «Критическая роль флавина и глутатиона в комплексной I-опосредованной биоэнергетической недостаточности при ишемии/реперфузионном повреждении головного мозга» . Гладить . 49 (5): 1223–1231. дои : 10.1161/СТРОКЕАХА.117.019687 . ПМК 5916474 . ПМИД 29643256 .
- ^ Jump up to: а б Степанова А, Сосунов С, Ниацецкая З, Конрад С, Старков АА, Манфреди Г и др. (сентябрь 2019 г.). «Редокс-зависимая потеря флавина митохондриальным комплексом I при ишемии/реперфузионном повреждении головного мозга» . Антиоксиданты и окислительно-восстановительная сигнализация . 31 (9): 608–622. дои : 10.1089/ars.2018.7693 . ПМК 6657304 . ПМИД 31037949 .
- ^ Андреацца AC, Шао Л., Ван Дж. Ф., Янг Л.Т. (апрель 2010 г.). «Активность митохондриального комплекса I и окислительное повреждение митохондриальных белков в префронтальной коре пациентов с биполярным расстройством» . Архив общей психиатрии . 67 (4): 360–8. doi : 10.1001/archgenpsychiatry.2010.22 . ПМИД 20368511 .
- ^ Моран М., Ривера Х., Санчес-Араго М., Бласкес А., Меринеро Б., Угальде С., Аренас Х., Куэсва Х.М., Мартин М.А. (май 2010 г.). «Взаимодействие митохондриальной биоэнергетики и динамики в сложных фибробластах с дефицитом I» . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1802 (5): 443–53. дои : 10.1016/j.bbadis.2010.02.001 . ПМИД 20153825 .
- ^ Бинукумар Б.К., Бал А., Кандималла Р., Сункария А., Гилл К.Д. (апрель 2010 г.). «Нарушение митохондриального энергетического обмена и дисфункция печени после хронического воздействия дихлофоса». Токсикология . 270 (2–3): 77–84. дои : 10.1016/j.tox.2010.01.017 . ПМИД 20132858 .
- ^ Сабатер, Б. (19 ноября 2021 г.). «На грани ненужности: гены хлоропластов » . Международный журнал молекулярных наук . 22 (22): 12505. doi : 10.3390/ijms222212505 . ПМЦ 8621559 . ПМИД 34830386 .
Внешние ссылки
[ редактировать ]- Институт науки и технологий Австрии (ISTA): Группа Сазанова MRC МБУ Группа Сазанова
- Интерактивная молекулярная модель НАДН-дегидрогеназы (требуется MDL Chime )
- Домашняя страница Комплекса I
- Страница новостей комплекса I в Facebook
- Электрон + Транспорт + Комплекс + I в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
На момент редактирования в этой статье используется контент из «3.D.1 Семейство H+ или Na+-транслокирующих НАДН-дегидрогеназ (NDH)» , которое лицензируется таким образом, что разрешается повторное использование в соответствии с непортированной лицензией Creative Commons Attribution-ShareAlike 3.0 . но не под GFDL . Все соответствующие условия должны быть соблюдены.