Тиоредоксинредуктаза
| Тиоредоксинредуктаза | |
|---|---|
| Идентификаторы | |
| Символ | ? |
| ИнтерПро | ИПР005982 |
| PROSITE | PS00573 |
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1зоф / СКОПе / СУПФАМ |
Тиоредоксинредуктазы ( TR , TrxR ) ( EC 1.8.1.9 ) представляют собой ферменты, восстанавливающие тиоредоксин (Trx). [ 1 ] Были идентифицированы два класса тиоредоксинредуктазы: один класс у бактерий и некоторых эукариот и один у животных. В бактериях TrxR также катализирует восстановление глутаредоксиноподобных белков, известных как NrdH. [ 2 ] [ 3 ] [ 4 ] Оба класса представляют собой флавопротеины , функционирующие как гомодимеры. Каждый мономер содержит простетическую группу FAD , домен связывания НАДФН и активный центр, содержащий окислительно-восстановительную дисульфидную связь . [ 5 ]
Сотовая роль
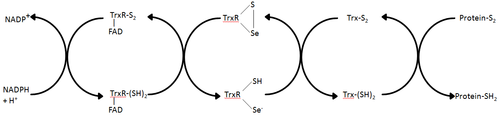
[ редактировать ]Тиоредоксинредуктазы – ферменты, катализирующие восстановление тиоредоксина. [ 1 ] и, следовательно, они являются центральным компонентом тиоредоксиновой системы. Вместе с тиоредоксином (Trx) и НАДФН наиболее общее описание этой системы представляет собой систему восстановления дисульфидных связей в клетках. Электроны отбираются у НАДФН через TrxR и переносятся в активный центр Trx, что приводит к восстановлению дисульфидов белка или других субстратов. [ 6 ] Система Trx существует во всех живых клетках и имеет эволюционную историю, связанную с ДНК как генетическим материалом, защитой от окислительного повреждения из-за метаболизма кислорода и передачей окислительно-восстановительных сигналов с использованием таких молекул, как перекись водорода и оксид азота. [ 7 ] [ 8 ]
Разнообразие
[ редактировать ]Два класса тиоредоксинредуктазы развились независимо:
- Высокомолекулярный тип (MW = ~ 55 000), содержащий остаток селеноцистеина в активном центре, был идентифицирован у высших эукариот, включая человека. Этот TxR связан с глутатионредуктазой , трипанотионредуктазой , редуктазой ртути и липоамиддегидрогеназой . [ 5 ]
- Низкомолекулярный тип (MW = ~ 35 000) идентифицирован у архей, бактерий и других эукариев. [ 5 ]
Эти два класса TrxR имеют лишь ~20% идентичности последовательностей в той части первичной последовательности, где их можно надежно выровнять. [ 5 ] Конечная реакция обоих классов TrxR идентична, но механизм действия каждого из них различен. [ 9 ]
У людей экспрессируются три изофермента тиоредоксинредуктазы: тиоредоксинредуктаза 1 (TrxR1, цитозольная), тиоредоксинредуктаза 2 (TrxR2, митохондриальная), тиоредоксинредуктаза 3 (TrxR3, специфичная для семенников). [ 10 ] Каждый изозим кодируется отдельным геном:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Структура
[ редактировать ]кишечная палочка
[ редактировать ]В ThxR E. coli имеется два связывающих домена: один для FAD , а другой для NADPH . Соединение между этими двумя доменами представляет собой двухцепочечный антипараллельный β-лист . [ 11 ] Каждый домен по отдельности очень похож на аналогичные домены в глутатионредуктазе и липоамиддегидрогеназе , но относительная ориентация этих доменов в ThxR повернута на 66 градусов. [ 11 ] Это становится важным в механизме действия фермента, который описан ниже. ThxR гомодимеризуется, при этом граница раздела между двумя мономерами образована тремя альфа-спиралями и двумя петлями. [ 11 ] Каждый мономер может отдельно связывать молекулу тиоредоксина .
-
 Структура E. coli тиоредоксина, связанного с димером ThxR
Структура E. coli тиоредоксина, связанного с димером ThxR -
 Структура ThxR E. coli с меченными простетическими группами FAD и NADPH
Структура ThxR E. coli с меченными простетическими группами FAD и NADPH


млекопитающее
[ редактировать ]Структура TrxR млекопитающих аналогична структуре E. coli . Он содержит домен связывания FAD и NADPH , а также интерфейс между двумя субъединицами мономера. У ThxR млекопитающих имеется вставка в домене связывания FAD между двумя альфа-спиралями, которая образует небольшую пару бета-цепей. [ 12 ] Активный дисульфид фермента расположен на одной из этих спиралей, и, таким образом, активная дисульфидная связь расположена в домене FAD , а не в домене NADPH , как у E. coli и других прокариот . [ 12 ]
-
 Структура простетических групп ThxR FAD и NADPH человека
Структура простетических групп ThxR FAD и NADPH человека

Механизм
[ редактировать ]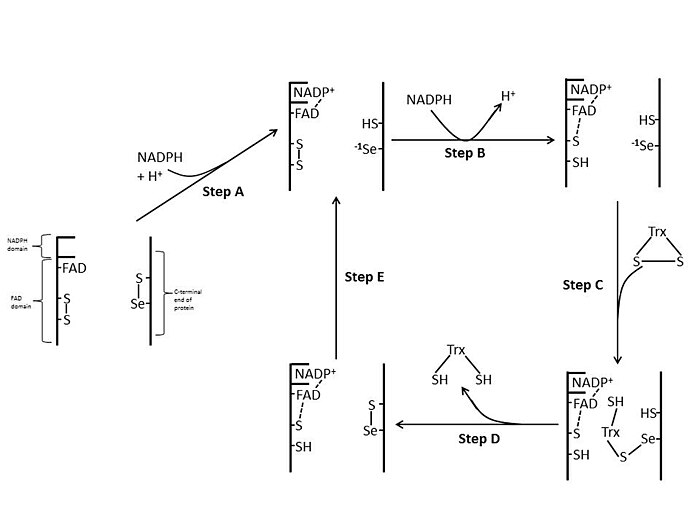
кишечная палочка
[ редактировать ]В E. coli ThxR пространственная ориентация доменов FAD и НАДФН такова, что редокс-активные кольца FAD и НАДФН не находятся в непосредственной близости друг от друга. [ 1 ] Когда домен FAD E. coli поворачивается на 66 градусов, а домен NADPH остается фиксированным, две простетические группы перемещаются в тесный контакт, позволяя электронам переходить от НАДФН к FAD, а затем к дисульфидной связи в активном центре. [ 1 ] [ 15 ] Консервативными остатками активного центра в E. coli являются -Cys-Ala-Thr-Cys-. [ 1 ]
млекопитающее
[ редактировать ]TrxR млекопитающих имеют гораздо более высокую гомологию последовательностей с глутатионредуктазой, чем E. coli . [ 1 ] Остатки Cys в активном центре в домене FAD и связанном домене NADPH находятся в непосредственной близости, что устраняет необходимость поворота на 66 градусов для переноса электрона, обнаруженного в E. coli . Дополнительной особенностью механизма млекопитающих является наличие остатка селеноцистеина на С-конце белка, необходимого для каталитической активности. Консервативными остатками в активном центре млекопитающих являются -Cys-Val-Asn-Val-Gly-Cys-. [ 1 ]
Методы обнаружения
[ редактировать ]Тиоредоксинредуктазу можно определить количественно различными методами, такими как анализ DTNB с использованием реагента Эллмана . Серия флуоресцентных зондов TRFS на основе дисульфида продемонстрировала избирательное обнаружение TrxR. [ 16 ] [ 17 ] [ 18 ] [ 19 ] Мафири синтезировал первый диселенидный зонд, который был применен для обнаружения TrxR. [ 20 ] [ 21 ] Другие методы обнаружения включают иммунологические методы и анализ селеноцистин-тиоредоксинредуктазы (анализ SC-TR).
Клиническое значение
[ редактировать ]Лечение рака
[ редактировать ]Поскольку активность этого фермента необходима для роста и выживания клеток, он является хорошей мишенью для противоопухолевой терапии. Кроме того, активность фермента повышается при некоторых типах рака, включая злокачественную мезотелиому . [ 22 ] [ 23 ] Например, мотексафин гадолиний (MGd) — новый химиотерапевтический агент, избирательно воздействующий на опухолевые клетки, приводящий к гибели клеток и апоптозу за счет ингибирования тиоредоксинредуктазы и рибонуклеотидредуктазы .
Кардиомиопатия
[ редактировать ]Дилатационная кардиомиопатия ( ДКМП ) является распространенным диагнозом в случаях застойной сердечной недостаточности . Тиоредоксинредуктазы являются важными белками для регулирования клеточного окислительно-восстановительного баланса и смягчения повреждений, вызванных активными формами кислорода, образующимися в результате окислительного фосфорилирования в митохондриях . Инактивация митохондриального TrxR2 у мышей приводит к истончению стенок желудочков сердца и неонатальной смерти. [ 10 ] Кроме того, две мутации в гене TrxR2 обнаружены у пациентов с диагнозом ДКМП, а не в контрольной популяции. Предполагается, что патологическое влияние этих мутаций заключается в нарушении способности контролировать окислительное повреждение кардиомиоцитов . [ 24 ]
Антибиотик
[ редактировать ]Недавно были проведены исследования, показавшие, что низкомолекулярная тиоредоксинредуктаза может быть мишенью для новых антибиотиков (таких как ауранофин или эбселен). [ 25 ] ) Это особенно верно для Mycobacterium Haemophilum и может быть использовано для бактерий, устойчивых к антибиотикам. [ 26 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г Мустачич Д., Повис Дж. (февраль 2000 г.). «Тиоредоксинредуктаза» . Биохимический журнал . 346 Ч. 1 (1): 1–8. дои : 10.1042/0264-6021:3460001 . ПМК 1220815 . ПМИД 10657232 .
- ^ Джордан А., Аслунд Ф., Понтис Э., Райхард П., Холмгрен А. (июль 1997 г.). «Характеристика Escherichia coli NrdH. Глутаредоксиноподобный белок с тиоредоксинподобным профилем активности» . Журнал биологической химии . 272 (29): 18044–50. дои : 10.1074/jbc.272.29.18044 . ПМИД 9218434 .
- ^ Пхулера С., Манде СК (июнь 2013 г.). «Кристаллическая структура микобактерии туберкулеза NrdH при размере 0,87 Å предполагает возможный способ ее активности». Биохимия . 52 (23): 4056–65. дои : 10.1021/bi400191z . ПМИД 23675692 .
- ^ Пхулера С., Акиф М., Сардесай А.А., Манде СК (01 января 2014 г.). «Окислительно-восстановительные белки микобактерий туберкулеза» . Журнал Индийского института науки . 94 (1): 127–138. ISSN 0970-4140 .
- ^ Перейти обратно: а б с д Хирт Р.П., Мюллер С., Эмбли Т.М., Кумбс Г.Х. (июль 2002 г.). «Разнообразие и эволюция тиоредоксинредуктазы: новые перспективы». Тенденции в паразитологии . 18 (7): 302–8. дои : 10.1016/S1471-4922(02)02293-6 . ПМИД 12379950 .
- ^ Перейти обратно: а б Холмгрен А., Лу Дж. (май 2010 г.). «Тиоредоксин и тиоредоксинредуктаза: текущие исследования с особым упором на болезни человека» . Связь с биохимическими и биофизическими исследованиями . 396 (1): 120–4. дои : 10.1016/j.bbrc.2010.03.083 . ПМИД 20494123 .
- ^ Мейер Ю., Бьюкенен Б.Б., Виньольс Ф., Райххельд Дж.П. (2009). «Тиоредоксины и глутаредоксины: объединяющие элементы в окислительно-восстановительной биологии». Ежегодный обзор генетики . 43 : 335–67. doi : 10.1146/annurev-genet-102108-134201 . ПМИД 19691428 .
- ^ Лиллиг CH, Холмгрен А (январь 2007 г.). «Тиоредоксин и родственные молекулы - от биологии к здоровью и болезням». Антиоксиданты и окислительно-восстановительная сигнализация . 9 (1): 25–47. дои : 10.1089/ars.2007.9.25 . ПМИД 17115886 .
- ^ Арскотт Л.Д., Громер С., Ширмер Р.Х., Беккер К., Уильямс CH (апрель 1997 г.). «Механизм тиоредоксинредуктазы из плаценты человека подобен механизмам липоамиддегидрогеназы и глутатионредуктазы и отличается от механизма тиоредоксинредуктазы из Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (8): 3621–6. Бибкод : 1997PNAS...94.3621A . дои : 10.1073/pnas.94.8.3621 . ЧВК 20490 . ПМИД 9108027 .
- ^ Перейти обратно: а б Конрад М., Якупоглу С., Морено С.Г., Липпль С., Банжак А., Шнайдер М., Бек Х., Хацопулос А.К., Джаст Ю., Синовац Ф., Шмаль В., Чиен К.Р., Вурст В., Борнкамм Г.В., Брилмайер М. (ноябрь 2004 г.). «Основная роль митохондриальной тиоредоксинредуктазы в кроветворении, развитии сердца и функции сердца» . Молекулярная и клеточная биология . 24 (21): 9414–23. дои : 10.1128/MCB.24.21.9414-9423.2004 . ПМК 522221 . ПМИД 15485910 .
- ^ Перейти обратно: а б с Уильямс CH (октябрь 1995 г.). «Механизм и структура тиоредоксинредуктазы Escherichia coli» . Журнал ФАСЭБ . 9 (13): 1267–76. дои : 10.1096/fasebj.9.13.7557016 . hdl : 2027.42/154540 . ПМИД 7557016 . S2CID 26055087 .
- ^ Перейти обратно: а б Сандалова Т., Чжун Л., Линдквист Ю., Холмгрен А., Шнайдер Г. (август 2001 г.). «Трехмерная структура тиоредоксинредуктазы млекопитающих: значение для механизма и эволюции селеноцистеин-зависимого фермента» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (17): 9533–8. Бибкод : 2001PNAS...98.9533S . дои : 10.1073/pnas.171178698 . ПМК 55487 . ПМИД 11481439 .
- ^ Чжун Л., Арнер Э.С., Холмгрен А. (май 2000 г.). «Структура и механизм тиоредоксинредуктазы млекопитающих: активный центр представляет собой окислительно-восстановительно-активный селенолтиол / селененилсульфид, образованный из консервативной последовательности цистеин-селеноцистеин» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (11): 5854–9. Бибкод : 2000PNAS...97.5854Z . дои : 10.1073/pnas.100114897 . ЧВК 18523 . ПМИД 10801974 .
- ^ Беккер К., Херольд-Менде С., Парк Дж.Дж., Лоу Г., Ширмер Р.Х. (август 2001 г.). «Человеческая тиоредоксинредуктаза эффективно ингибируется комплексами (2,2':6',2''-терпиридин)платины(II). Возможные последствия для новой противоопухолевой стратегии». Журнал медицинской химии . 44 (17): 2784–92. дои : 10.1021/jm001014i . ПМИД 11495589 .
- ^ Леннон BW, Williams CH (август 1997 г.). «Восстановительная полуреакция тиоредоксинредуктазы Escherichia coli». Биохимия . 36 (31): 9464–77. дои : 10.1021/bi970307j . ПМИД 9235991 .
- ^ Ли Х, Чжан Б., Ян С., Ли Дж., Ван С., Вэй Икс и др. (июнь 2019 г.). «Быстрый и специфичный флуоресцентный зонд для тиоредоксинредуктазы, который действует за счет расщепления дисульфидной связи» . Природные коммуникации . 10 (1): 2745. Бибкод : 2019NatCo..10.2745L . дои : 10.1038/s41467-019-10807-8 . ПМК 6588570 . ПМИД 31227705 .
- ^ Ма Х, Чжан Дж, Чжан Цз, Лю Ю, Фан Дж (октябрь 2016 г.). «Зонд с быстрым откликом и красной эмиссией тиоредоксинредуктазы млекопитающих». Химические коммуникации . 52 (81): 12060–12063. дои : 10.1039/C6CC04984B . ПМИД 27709154 .
- ^ Чжао Дж., Цюй Ю., Гао Х., Чжун М., Ли Х., Чжан Ф. и др. (ноябрь 2020 г.). «Потеря функции тиоредоксинредуктазы в модели инсульта у мышей, выявленная с помощью двухфотонного флуоресцентного зонда». Химические коммуникации . 56 (90): 14075–14078. дои : 10.1039/D0CC05900E . ПМИД 33107534 . S2CID 225082279 .
- ^ Лю Ю, Ма Х, Чжан Л, Цуй Ю, Лю С, Фан Дж (февраль 2016 г.). «Небольшой молекулярный зонд выявил снижение активности митохондриальной тиоредоксинредуктазы на модели болезни Паркинсона». Химические коммуникации . 52 (11): 2296–9. дои : 10.1039/c5cc09998f . ПМИД 26725656 .
- ^ Мафирейи Т.Дж., Лоуз М., Бассетт Дж.В., Кэссиди П.Б., Эскобедо Дж.О., Стронгин Р.М. (август 2020 г.). «Диселенидный флуоресцентный зонд для обнаружения тиоредоксинредуктазы» . Ангеванде Хеми . 59 (35): 15147–15151. Бибкод : 2020АнгЧ.13215259М . дои : 10.1002/ange.202004094 . ПМЦ 9438933 . ПМИД 32449244 . S2CID 229142596 .
- ^ Мафирейи Т.Дж., Эскобедо Дж.О., Стронгин Р.М. (29 марта 2021 г.). «Флуорогенные зонды для активности тиоредоксинредуктазы» . Результаты по химии . 3 : 100127. doi : 10.1016/j.rechem.2021.100127 . ISSN 2211-7156 .
- ^ Нильсонн Г, Сан Х, Нюстрем С, Рундлёф АК, Потамиту Фернандес А, Бьёрнштедт М, Добра К (сентябрь 2006 г.). «Селенит индуцирует апоптоз в клетках саркоматоидной злокачественной мезотелиомы посредством окислительного стресса». Свободно-радикальная биология и медицина . 41 (6): 874–85. doi : 10.1016/j.freeradbiomed.2006.04.031 . hdl : 10616/47514 . ПМИД 16934670 .
- ^ Калос К., Сойни Й., Сайли М., Койстинен П., Какко С., Пяаккё П., Холмгрен А., Киннула В.Л. (май 2001 г.). «Повышающая регуляция тиоредоксина и тиоредоксинредуктазы при злокачественной мезотелиоме плевры человека» . Международный журнал рака . 95 (3): 198–204. doi : 10.1002/1097-0215(20010520)95:3<198::AID-IJC1034>3.0.CO;2-F . ПМИД 11307155 .
- ^ Сиббинг Д., Пфойфер А., Перишич Т., Маннес А.М., Фриц-Вольф К., Анвин С., Синнер М.Ф., Гигер С., Глёкнер С.Дж., Вихманн Х.Е., Креммер Е., Шефер З., Вальх А., Хинтерзеер М., Нэбауэр М., Кеаб С., Кастрати А., Шёмиг А., Мейтингер Т., Борнкамм Г.В., Конрад М., фон Бекерат Н. (май 2011 г.). «Мутации в митохондриальном гене тиоредоксинредуктазы TXNRD2 вызывают дилатационную кардиомиопатию» . Европейский кардиологический журнал . 32 (9): 1121–33. doi : 10.1093/eurheartj/ehq507 . hdl : 11858/00-001M-0000-0024-1F10-3 . ПМИД 21247928 .
- ^ Маршалл А.С., Кидд С.Е., Ламонт-Фридрих С.Дж., Аренц Г., Хоффманн П., Коад Б.Р., Брюнинг Дж.Б. (март 2019 г.). «Тиоредоксинредуктаза Aspergillus fumigatus» . Антимикробные средства и химиотерапия . 63 (3). дои : 10.1128/AAC.02281-18 . ПМК 6395915 . ПМИД 30642940 .
- ^ Харбут М.Б., Вильчез С., Луо X, Хенслер М.Е., Го Х., Ян Б. и др. (апрель 2015 г.). «Ауранофин оказывает бактерицидное действие широкого спектра, воздействуя на тиол-окислительно-восстановительный гомеостаз» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (14): 4453–8. Бибкод : 2015PNAS..112.4453H . дои : 10.1073/pnas.1504022112 . ПМЦ 4394260 . ПМИД 25831516 .
Внешние ссылки
[ редактировать ]- Тиоредоксин + редуктаза + (НАДФН) в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)