Анатомия птиц
Анатомия птиц , или физиологическая структура тел птиц , демонстрирует множество уникальных приспособлений, в основном способствующих полету . У птиц легкий скелет и легкая, но мощная мускулатура , которая, наряду с кровеносной и дыхательной системами, способными к очень высокой скорости обмена веществ и снабжению кислородом , позволяет птице летать. Развитие клюва привело к эволюции специально приспособленной пищеварительной системы .
Скелетная система
[ редактировать ]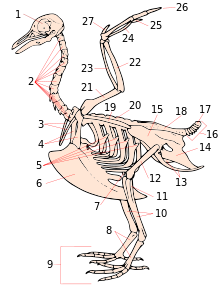
- череп
- шейные позвонки
- вилка
- коракоид
- крючковидные отростки ребер
- киль
- коленная чашечка
- предплюсна-плюсна
- цифры
- большеберцовая кость ( tibiotarsus )
- малоберцовая кость ( берцовая кость )
- бедренная кость
- седалищная кость ( безымянная )
- Лобок (без имени)
- его (безымянный)
- хвостовые позвонки
- пигостиль
- синсакрум
- лопатка
- спинные позвонки
- плечевая кость
- локтевая кость
- радиус
- Карпометакарпус
- Цифра III
- Цифра II
- Цифра I ( алула )

- Клюв
- Голова
- Ирис
- Ученик
- Мантия
- Малые кроющие
- лопатки
- Покрытия
- Тертиалы
- крупа
- Праймериз
- Вентиляция
- Бедро
- Тибио-тарзальное сочленение
- Тарсус
- Ноги
- большеберцовая кость
- Живот
- фланги
- Грудь
- Горло
- Плетень
- Полоска для глаз
У птиц многие кости полые ( пневматизированные ) с перекрещивающимися распорками или фермами для обеспечения структурной прочности . Количество полых костей варьируется у разных видов, хотя у крупных планирующих и парящих птиц их, как правило, больше. Дыхательные воздушные мешки часто образуют воздушные карманы внутри полуполых костей скелета птицы. [1] Кости ныряющих птиц часто менее полые, чем у неныряющих видов. Пингвины , гагары , [2] а тупики вообще лишены пневматизированных костей. [3] [4] У нелетающих птиц , таких как страусы и эму пневматизированы. , бедренные кости [5] и, в случае эму, пневматизированные шейные позвонки . [6]
Осевой скелет
[ редактировать ]Скелет птиц хорошо приспособлен к полету . Он чрезвычайно легкий, но достаточно прочный, чтобы выдерживать нагрузки при взлете, полете и приземлении. Одной из ключевых адаптаций является слияние костей в отдельные окостенения , такие как пигостиль . Из-за этого у птиц обычно меньше костей, чем у других наземных позвоночных. У птиц также нет зубов или даже настоящей челюсти , вместо этого у них есть клюв , который гораздо легче. В клюве многих птенцов имеется выступ, называемый яйцевым зубом , который облегчает их выход из околоплодного яйца . Он отпадает после проникновения в яйцо.

| Цвет | Позвоночный отдел |
|---|---|
| Розовый | Шейные позвонки |
| Апельсин | Грудные/спинные позвонки |
| Желтый | Синсакрум |
| Зеленый | Хвостовые позвонки |
| Синий | Пигостиль |
Позвоночник
[ редактировать ]В позвоночнике выделяют пять отделов позвонков :
Шейные позвонки
[ редактировать ]Шейные позвонки обеспечивают структурную поддержку шеи и насчитывают от 8 до 25 позвонков у некоторых видов лебедей ( Cygninae ) и других птиц с длинной шеей. Все шейные позвонки имеют прикрепленные поперечные отростки, кроме первого. Этот позвонок (С1) называется атлантом, он сочленяется с затылочными мыщелками черепа и не имеет отверстия, типичного для большинства позвонков. [7] Шея птицы состоит из множества шейных позвонков, что позволяет птицам обладать повышенной гибкостью. Гибкая шея позволяет многим птицам с неподвижными глазами более продуктивно двигать головой и концентрировать взгляд на предметах, находящихся близко или далеко на расстоянии. [8] У большинства птиц примерно в три раза больше шейных позвонков, чем у людей, что обеспечивает повышенную устойчивость во время быстрых движений, таких как полет, приземление и взлет. [9] Шея играет роль в покачивании головой, которое присутствует как минимум у 8 из 27 отрядов птиц, включая Columbiformes , Galliformes и Gruiformes . [10] Покачивание головой - это оптокинетическая реакция , которая стабилизирует окружение птицы, когда она чередуется между фазой толчка и фазой удержания. [11] Качание головы синхронно с движением ног, поскольку голова движется в соответствии с остальным телом. [11] Данные различных исследований показывают, что основной причиной покачивания головой у некоторых птиц является стабилизация окружающей среды, хотя неясно, почему в некоторых, но не во всех отрядах птиц наблюдается покачивание головой. [12]
Грудные позвонки
[ редактировать ]Грудных позвонков насчитывается от 5 до 10, причем первый грудной позвонок различим благодаря сращению прикрепленного к нему ребра с грудиной, тогда как ребра шейных позвонков свободны. [7] Передние грудные позвонки у многих птиц сращены и сочленяются с нотарием грудного пояса. [13]

Синсакрум
[ редактировать ]Синсакрум . состоит из одного грудного, шести поясничных, двух крестцовых и пяти крестцово-каудальных позвонков, сросшихся в одну окостеневшую структуру, которая затем срастается с подвздошной костью [14] Вне полета эта структура обеспечивает основную поддержку остальной части тела. [7] Подобно крестцу млекопитающих, синсакрум не имеет отчетливой формы диска, характерной для шейных и грудных позвонков. [15]
Хвостовые позвонки
[ редактировать ]Свободные позвонки, следующие сразу за сросшимися крестцово-каудальными позвонками синсакра, известны как хвостовые позвонки. У птиц от 5 до 8 свободных хвостовых позвонков. [7] Хвостовые позвонки обеспечивают структуру хвостов позвоночных и гомологичны копчику млекопитающих , лишенных хвостов. [16]
Пигостиль
[ редактировать ]У птиц последние 5–6 хвостовых позвонков срастаются, образуя пигостиль . [14] В некоторых источниках отмечается, что эту сросшуюся структуру могут составлять до 10 хвостовых позвонков. Эта структура обеспечивает точку крепления рулевых перьев, которые помогают контролировать полет. [7]

лопаточный пояс
[ редактировать ]Птицы — единственные живые позвоночные , у которых сросшиеся ключицы и килевидная грудная кость . [17] Килевидная грудина служит местом прикрепления мышц, используемых при полете или плавании. [17] Нелетающие птицы, такие как страусы , не имеют килевидной грудины и имеют более плотные и тяжелые кости по сравнению с летающими птицами. [18] У плавающих птиц широкая грудина, у ходячих птиц - длинная, а у летающих птиц грудина почти одинаковая по ширине и высоте. [19] Грудная клетка состоит из вилки (рычажной кости) и клювовидного отростка (воротничковой кости), которые вместе с лопаткой образуют грудной пояс ; боковая часть грудной клетки образована ребрами, сходящимися у грудины (средняя линия грудной клетки). [7]
Ребрышки
[ редактировать ]имеются крючковидные отростки У птиц на ребрах . Это крючкообразные выступы костей, которые помогают укрепить грудную клетку, перекрывая ребро позади них. Эта особенность также встречается у туатары ( Sphenodon ).
Череп
[ редактировать ]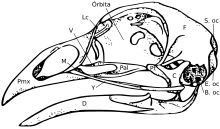
Череп состоит из пяти основных костей: лобной (макушка головы), теменной (затылок), предчелюстной и носовой (верхний клюв ) и нижней челюсти (нижний клюв). Череп нормальной птицы обычно весит около 1% от общей массы тела птицы. Глаз занимает значительную часть черепа и окружен склеротическим глазным кольцом — кольцом крошечных костей. Эта характеристика также наблюдается у их собратьев-рептилий.
Грубо говоря, черепа птиц состоят из множества мелких, не перекрывающихся костей. Считается, что педоморфоз , сохранение наследственного состояния у взрослых, способствовал эволюции птичьего черепа. По сути, черепа взрослых птиц будут напоминать молодые формы их предков -теропод-динозавров . [20] По мере развития птичьего происхождения и возникновения педоморфоза они потеряли заглазничную кость за глазом, эктоптеригоид в задней части неба и зубы. [21] [22] Структуры неба также сильно изменились с изменениями, в основном с редукцией, наблюдаемыми в крыловидной, небной и скуловой костях. Также произошло сокращение приводящих камер. [22] Все эти состояния наблюдались у ювенильных форм их предков. Предчелюстная , кость также гипертрофировалась образуя клюв, в то время как верхняя челюсть уменьшилась, о чем свидетельствуют оба исследования развития. [20] и палеонтологический [23] исследования. Это расширение клюва произошло одновременно с потерей функциональной руки и развитием точки в передней части клюва, напоминающей «палец». [22] играет Известно также, что предчелюстная кость большую роль в пищевом поведении рыб. [24] [25]
Строение черепа птиц имеет важное значение для их пищевого поведения. Птицы демонстрируют независимое движение костей черепа, известное как краниальный кинезис . Краниальный кинез у птиц встречается в нескольких формах, но все эти разновидности возможны благодаря анатомии черепа. Животные с крупными перекрывающимися костями (в том числе предки современных птиц) [26] [27] [28] имеют акинетический (некинетический) череп. [29] [30] По этой причине утверждалось, что педоморфный птичий клюв можно рассматривать как эволюционное нововведение. [22]
У птиц череп диапсидный , как и у рептилий, с предслезной ямкой (имеется у некоторых рептилий). Череп имеет единственный затылочный мыщелок . [31]
Аппендикулярный скелет
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( сентябрь 2018 г. ) |
Плечо состоит из лопатки (лопатки), клювовидного отростка и плечевой кости (плечо). Плечевая кость соединяется с лучевой и локтевой костью (предплечьем), образуя локоть. Запястье образуют «запястье» и « руку и пясть » птицы, а пальцы слиты вместе. Кости крыла очень легкие, поэтому птице легче летать.
Бедра состоят из таза, который включает в себя три основные кости: подвздошную (верхнюю часть бедра), седалищную (боковые части бедра) и лобковую (переднюю часть бедра). Они слиты в одну ( безымянную кость ). Безымянные кости имеют эволюционное значение, поскольку позволяют птицам откладывать яйца. Они встречаются в вертлужной впадине (тазовой впадине) и сочленяются с бедренной костью, которая является первой костью задней конечности.
Верхняя часть ноги состоит из бедренной кости. В коленном суставе бедренная кость соединяется с большеберцовой костью (голенью) и малоберцовой костью (боковой частью голени). Предплюсне -плюсневые кости образуют верхнюю часть стопы, пальцы составляют пальцы. Кости ног птиц самые тяжелые, что способствует низкому центру тяжести, что облегчает полет. Скелет птицы составляет всего около 5% от общей массы тела.
У них сильно удлиненный тетрадиатный таз , похожий на некоторые пресмыкающиеся. На задних конечностях имеется внутрипредплюсновый сустав, встречающийся и у некоторых рептилий. Наблюдается обширное сращение туловищных позвонков, а также сращение грудного пояса .
Крылья
[ редактировать ]Ноги
[ редактировать ]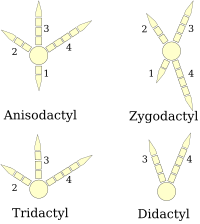
(схемы правой ноги)
Лапы птиц подразделяются на анизодактильные , зигодактильные , гетеродактильные , синдактильные или пампродактильные . [32] Анизодактиль — наиболее распространенное расположение пальцев у птиц: три пальца вперед и один назад. Это часто встречается у певчих и других птиц-насадников , а также у охотничьих птиц, таких как орлы , ястребы и соколы .
Синдактилия, встречающаяся у птиц, подобна анизодактилии, за исключением того, что второй и третий пальцы (внутренний и средний, направленные вперед), или три пальца, срослись вместе, как у опоясанного зимородка Ceryle alcyon . Это характерно для ракообразных ( зимородков , щурок , катунов и др.).
Зигодактильная (от греч. ζυγον , ярмо ) стопа имеет два пальца, обращенных вперед (вторая и третья цифры), и два назад (первая и четвертая цифры). Такое расположение наиболее распространено у древесных видов, особенно у тех, которые карабкаются по стволам деревьев или сквозь листву. Зигодактилия встречается у попугаев , дятлов (в том числе у мерцаний ), кукушек (в том числе у дорожных бегунов ) и некоторых сов . Были обнаружены следы зигодактиля, датируемые 120–110 млн лет назад (ранний мел ), за 50 миллионов лет до первых идентифицированных окаменелостей зигодактилей. [33]
Гетеродактилия похожа на зигодактилию, за исключением того, что цифры три и четыре направлены вперед, а цифры один и два — назад. Это встречается только у трогонов , тогда как пампродактиль — это конструкция, при которой все четыре пальца могут быть направлены вперед, а птицы могут поворачивать два внешних пальца назад. Это характерно для стрижей ( Apodidae ).
Эволюция
[ редактировать ]Изменение задних конечностей
[ редактировать ]Значительное сходство строения задних конечностей птиц и других динозавров связано с их способностью ходить на двух ногах, или прямохождением . [34] В XX веке преобладало мнение, что переход к прямохождению произошел за счет превращения передних конечностей в крылья. Современные ученые считают, что, наоборот, это было необходимым условием возникновения полета. [35]
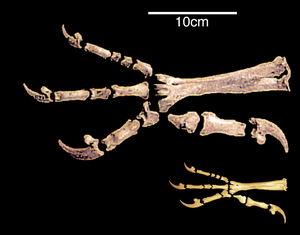
Переход к использованию для передвижения только задних конечностей сопровождался увеличением ригидности поясничного и крестцового отделов. Лобковые кости птиц и некоторых других двуногих динозавров обращены назад. Ученые связывают это со смещением центра тяжести тела назад. Причиной такого сдвига называют переход к прямохождению или развитие мощных передних конечностей, как у археоптерикса . [36] [37] Большой и тяжелый хвост двуногих динозавров, возможно, служил дополнительной опорой. Частичная редукция хвоста и последующее формирование пигостиля произошли за счет отклонения назад первого пальца задней конечности; у динозавров с длинным жестким хвостом развитие стопы протекало иначе. Этот процесс, по-видимому, происходил параллельно у птиц и некоторых других динозавров. Вообще, у птиц предковой является анизодактильная стопа, которая к тому же обладает лучшей хватательной способностью и позволяет уверенно передвигаться как по земле, так и по веткам. На этом фоне выделяются птерозавры, которые в процессе неудачных эволюционных изменений не смогли полноценно передвигаться на двух ногах, а вместо этого развили физические средства полета. [ нужны дальнейшие объяснения ] это принципиально отличалось от птиц. [37]
Изменения передних конечностей
[ редактировать ]Изменения задних конечностей не затронули расположение передних конечностей, которые у птиц оставались раздвинутыми по бокам, а у нептичьих динозавров перешли на парасагиттальную ориентацию. [36] В то же время передние конечности, освободившиеся от опорной функции, имели широкие возможности для эволюционных изменений. Сторонники гипотезы бега считают, что полет сформировался благодаря быстрому бегу, подпрыгиванию, а затем скольжению. Передние конечности могли использоваться для хватания после прыжка или в качестве «сетей для ловли насекомых», животные могли ими махать, помогая себе во время прыжка. Согласно древесной гипотезе, предки птиц лазили по деревьям с помощью передних конечностей и оттуда планировали, после чего приступали к полету. [38]
Мышечная система
[ редактировать ]
У большинства птиц имеется около 175 различных мышц, в основном управляющих крыльями, кожей и ногами. В целом мышечная масса птиц сосредоточена вентрально. Самыми крупными мышцами птицы являются грудные мышцы, или большая грудная мышца, которая управляет крыльями и составляет около 15–25% массы тела летающей птицы. Они обеспечивают мощный взмах крыльев, необходимый для полета. Мышца, расположенная глубоко (под) грудными мышцами, представляет собой супракоракоидеус, или малую грудную мышцу. Он поднимает крыло между взмахами крыльев. Обе группы мышц прикрепляются к килю грудины. Это примечательно, поскольку у других позвоночных есть мышцы, позволяющие поднимать верхние конечности, обычно прикрепленные к областям задней части позвоночника. Супракоракоид и грудные кости вместе составляют около 25–40% полной массы тела птицы. [39] Каудальнее грудных мышц и супракораковидной мышцы расположены внутренние и внешние косые мышцы живота, которые сжимают живот. Кроме того, имеются и другие мышцы живота, которые расширяют и сжимают грудную клетку, а также удерживают грудную клетку. Мышцы крыла, как видно на отмеченных изображениях, функционируют в основном при разгибании или сгибании локтя, перемещении крыла в целом или разгибании или сгибании отдельных пальцев. Эти мышцы регулируют крылья для полета и всех других действий. [39] Состав мышц различается у разных видов и даже внутри семейств. [40]

У птиц уникальные шеи, удлиненные и со сложной мускулатурой, поскольку они позволяют голове выполнять функции, для которых другие животные могут использовать грудные конечности. [39]

Кожные мышцы помогают птице в полете, регулируя перья, которые прикреплены к кожным мышцам, и помогают птице в маневрах полета, а также помогают в брачных ритуалах.
Мышц на туловище и хвосте всего несколько, но они очень сильные и необходимы птице. К ним относятся латеральная мышца хвоста и мышца, поднимающая хвост, которые контролируют движение хвоста и раскрытие прямоугольников, придавая хвосту большую площадь поверхности, что помогает удерживать птицу в воздухе, а также помогает при повороте. [39]
Мышечный состав и адаптация различаются в зависимости от теорий мышечной адаптации в зависимости от того, произошла ли эволюция полета сначала в результате взмахов руками или скольжения. [41]
Покровная система
[ редактировать ]
Весы
[ редактировать ]Чешуя птиц , как и клювы, когти и шпоры, состоит из кератина. Они встречаются в основном на пальцах ног и лапках (голенях птиц), обычно до большеберцово-предплюсневого сустава, но у некоторых птиц их можно обнаружить и дальше по ногам. У многих орлов и сов ноги оперены до пальцев ног (но не включая их). [42] [43] [44] Чешуя большинства птиц существенно не перекрывается, за исключением случаев зимородков и дятлов . Первоначально считалось, что чешуя и щитки птиц гомологичны чешуе и щиткам рептилий; [45] однако более поздние исследования показывают, что чешуя у птиц изменилась после эволюции перьев. [46] [47] [48]
Эмбрионы птиц начинают развитие с гладкой кожей. На ступнях роговой слой или самый внешний слой кожи может ороговеть, утолщаться и образовывать чешуйки. Эти шкалы можно разделить на;
- Канцелла – мелкие чешуйки, которые на самом деле представляют собой всего лишь утолщение и затвердение кожи, испещренное неглубокими бороздками.
- Щиток — чешуя, которая не так велика, как щитки, например, на хвостовой курицы или задней части плюсны .
- Щитки – самые крупные чешуи, обычно на передней поверхности плюсны и дорсальной поверхности пальцев стопы.
Ряды щитков на передней части плюсны можно назвать «акрометатарзием» или «акротарзием».
Ретикулы расположены на латеральной и медиальной поверхностях (боках) стопы и первоначально считались отдельными чешуйками. Однако гистологические и эволюционные исследования в этой области показали, что в этих структурах отсутствует бета-кератин (отличительный признак чешуи рептилий) и они полностью состоят из альфа-кератина. [47] [49] Это, наряду с их уникальной структурой, привело к предположению, что на самом деле это почки перьев, задержанные на ранних стадиях развития. [47]
В совокупности чешуйчатый покров на лапах птиц называется подотека.
Тельца Хербста и знания
[ редактировать ]На клювах многих куликов есть тельца Гербста , которые помогают им находить добычу, спрятанную под мокрым песком, путем обнаружения малейшей разницы давления в воде. [50] Все современные птицы умеют перемещать части верхней челюсти относительно черепной коробки. Однако это более заметно у некоторых птиц и легко обнаруживается у попугаев. [51]
Область между глазом и клювом на боковой стороне головы птицы называется лором . Иногда эта область лишена перьев, а кожа может быть окрашена, как у многих видов семейства бакланов .
Клюв
[ редактировать ]Клюв, клюв или рострум — это внешняя анатомическая структура птиц, которая используется для еды, прихорашивания , манипулирования объектами, убийства добычи, борьбы, поиска пищи, ухаживания и кормления детенышей. Хотя клювы значительно различаются по размеру, форме и цвету, их основная структура схожа. Два костных выступа — верхняя и нижняя челюсти — покрыты тонким ороговевшим слоем эпидермиса, известным как рамфотека . У большинства видов к дыхательной системе ведут два отверстия, известные как ноздри.
Дыхательная система
[ редактировать ]


Из-за высокой скорости обмена веществ, необходимой для полета, птицам требуется большое количество кислорода. Их высокоэффективная дыхательная система помогает им удовлетворить этот спрос.
Хотя у птиц есть легкие, они представляют собой довольно жесткие структуры, которые не расширяются и не сжимаются, как у млекопитающих, рептилий и многих земноводных. Вместо этого структуры, которые действуют как мехи, вентилирующие легкие, представляют собой воздушные мешки , которые распределены по большей части тела птиц. [52] Воздушные мешки перемещают воздух в одном направлении через парабронхи ригидных легких. [53] [54] Основным механизмом однонаправленных потоков в легких птиц является необратимость потока при высоких числах Рейнольдса, проявляющаяся в асимметричности соединений и их петлеобразующей связности. [55]
Хотя легкие птиц меньше, чем у млекопитающих сопоставимого размера, воздушные мешки составляют 15% от общего объема тела, тогда как у млекопитающих альвеолы , выполняющие функцию мехов, составляют только 7% от общего объема тела. [56] В целом легкие птиц имеют площадь дыхательной поверхности примерно на 15% больше, объем крови в легочных капиллярах - в 2,5-3 раза больше, а гематогазовый барьер на 56-67% тоньше, чем в легких млекопитающих аналогичного типа. масса тела. [57] [58] Стенки воздухоносных мешков плохо кровоснабжаются и поэтому не играют прямой роли в газообмене .
У птиц нет диафрагмы , и поэтому они используют межреберные и брюшные мышцы для расширения и сжатия всей торако-брюшной полости, таким образом ритмично изменяя объемы всех своих воздушных мешков в унисон (иллюстрация справа). Активной фазой дыхания у птиц является выдох, требующий сокращения дыхательных мышц. [54] Расслабление этих мышц вызывает вдох.
Три различных набора органов осуществляют дыхание — передние воздушные мешки (межключичные, шейные и передние грудные), легкие и задние воздушные мешки (задние грудные и брюшные полости). Обычно в системе имеется девять воздушных мешков; [54] однако это число может варьироваться от семи до двенадцати, в зависимости от вида птиц. У воробьинообразных семь воздушных мешков, поскольку ключичные воздушные мешки могут соединяться или сливаться с передними грудными мешками.
При вдохе окружающий воздух первоначально поступает к птице через ноздри, откуда нагревается, увлажняется и фильтруется в носовых ходах и верхних отделах трахеи. [56] Оттуда воздух попадает в нижнюю часть трахеи и продолжает движение сразу за сиринкс , после чего трахея разветвляется на два главных бронха , идущие к двум легким. Первичные бронхи входят в легкие и становятся внутрилегочными бронхами, которые отдают ряд параллельных ветвей, называемых вентробронхами, а немного дальше — эквивалентный набор дорсобронхов. [59] Концы внутрилегочных бронхов отводят воздух в задние воздушные мешки на каудальном конце птицы. Каждая пара дорсо-вентробронхов соединена большим количеством параллельных микроскопических воздушных капилляров (или парабронхов), в которых происходит газообмен . [59] При вдохе птицы трахеальный воздух поступает через внутрилегочные бронхи в задние воздушные мешки, а также в дорсальные бронхи (но не в вентробронхи, отверстия которых во внутрилегочные бронхи, как считалось ранее, были плотно закрыты при вдохе). [59] Однако более поздние исследования показали, что аэродинамика бронхиальной архитектуры направляет вдыхаемый воздух от отверстий вентробронхов в продолжение внутрилегочного бронха в сторону дорсобронхов и задних воздухоносных мешков. [53] [60] ). Из дорсобронхов воздух поступает через парабронхи (и, следовательно, через газообменник) в вентробронхи, откуда воздух может выйти только в расширяющиеся передние воздушные мешки. Так, во время вдоха как задний, так и передний воздушные мешки расширяются. [59] задние воздушные мешки наполняются свежим вдыхаемым воздухом, а передние воздушные мешки наполняются «отработанным» (бедным кислородом) воздухом, который только что прошел через легкие.
Считалось, что во время выдоха внутрилегочные бронхи плотно сужаются между областью, где разветвляются вентробронхи, и областью, где разветвляются дорсобронхи. [59] Но теперь считается, что более сложные аэродинамические элементы имеют тот же эффект. [53] [60] Таким образом, сокращающиеся задние воздушные мешки могут опорожняться только в дорсобронхи. Отсюда свежий воздух из задних воздухоносных мешков поступает через парабронхи (в том же направлении, что и при вдохе) в вентробронхи. Воздушные пути, соединяющие вентробронхи и передние воздушные мешки с внутрилегочными бронхами, открываются во время выдоха, позволяя бедному кислородом воздуху из этих двух органов выходить через трахею наружу. [59] Таким образом, насыщенный кислородом воздух течет постоянно (в течение всего дыхательного цикла) в одном направлении через парабронхи. [1]

Кровоток через легкие птицы расположен под прямым углом к потоку воздуха через парабронхи, образуя перекрестно-токовую обменную систему (см. рисунок слева). [59] [61] Парциальное давление кислорода в парабронхах снижается по их длине по мере диффундирования О 2 в кровь. Кровеносные капилляры, выходящие из теплообменника вблизи входа воздушного потока, поглощают больше О 2 , чем капилляры, выходящие из околовыходного конца парабронхов. При смешивании содержимого всех капилляров конечное парциальное давление кислорода смешанной легочной венозной крови выше, чем у выдыхаемого воздуха, [59] [61] но тем не менее вдвое меньше, чем вдыхаемый воздух, [59] таким образом достигается примерно такое же системное парциальное давление кислорода в артериальной крови, как у млекопитающих с их легкими сильфонного типа . [59]
Трахея представляет собой область мертвого пространства : бедный кислородом воздух, содержащийся в ней в конце выдоха, первым возвращается в задние воздушные мешки и легкие. По сравнению с дыхательными путями млекопитающих объем мертвого пространства у птиц в среднем в 4,5 раза больше, чем у млекопитающих того же размера. [59] [56] Птицы с длинной шеей неизбежно будут иметь длинные трахеи, и поэтому им придется дышать глубже, чем млекопитающим, чтобы учесть больший объем мертвого пространства. У некоторых птиц (например, лебедя-кликуна , Cygnus cygnus , белой колпицы , Platalea leucorodia , американского журавля , Grus americana и шлемоносного курасса , Pauxi pauxi ) трахея, длина которой у некоторых журавлей может достигать 1,5 м, [59] скручивается взад и вперед внутри тела, резко увеличивая вентиляцию мертвого пространства. [59] Назначение этой необычной особенности неизвестно.
Воздух проходит через легкие в одном направлении как во время выдоха, так и во время вдоха, вызывая, за исключением бедного кислородом воздуха мертвого пространства, остающегося в трахее после выдоха и вдыхаемого в начале вдоха, практически не смешивать новый воздух, богатый кислородом, с отработанным. бедный кислородом воздух (как это происходит в легких млекопитающих ), меняющийся только (от богатого кислородом к бедному кислородом) по мере его движения (однонаправленно) через парабронхи.
В легких птиц нет альвеол , как в легких млекопитающих. Вместо этого они содержат миллионы узких проходов, известных как парабронхи, соединяющих дорсобронхи с вентробронхами на обоих концах легких. Воздух течет вперед (каудально краниально) через параллельные парабронхи. Эти парабронхи имеют сотовые стенки. Клетки сот представляют собой тупиковые воздушные пузырьки, называемые предсердиями , которые выступают радиально из парабронхов. Предсердия путем простой диффузии являются местом газообмена . [62] Кровоток вокруг парабронхов (и их предсердий) образует перекрестный газообменник (см. схему слева). [59] [61]
У всех видов птиц, за исключением пингвинов, в легких есть небольшой участок, посвященный «неопульмональным парабронхам». Эта неорганизованная сеть микроскопических трубок ответвляется от задних воздухоносных мешков и беспорядочно открывается как в дорсо-, так и в вентробронхи, а также непосредственно во внутрилегочные бронхи. В отличие от парабронхов, в которых воздух движется однонаправленно, поток воздуха в неопульмональных парабронхах двунаправленный. Неопульмональные парабронхи никогда не составляют более 25% общей газообменной поверхности птиц. [56]

Чтобы птицы издавали звук, они используют орган, расположенный над легкими, называемый сиринксом , который состоит из колец трахеи, сирингеальных мышц, барабанной перепонки и внутренних костных структур, которые способствуют производству звука. Затем воздух проходит через этот орган, что приводит к вокализации птиц. Звук может производиться за счет движения барабанной перепонки. Высота звука также может быть изменена путем открытия и закрытия барабанной перепонки, что позволяет увеличивать или уменьшать громкость звука. [63]
Кровеносная система
[ редактировать ]четырёхкамерное У птиц сердце , [64] совместно с млекопитающими и некоторыми рептилиями (главным образом крокодилами ). Эта адаптация обеспечивает эффективную транспортировку питательных веществ и кислорода по всему телу, обеспечивая птиц энергией для полета и поддержания высокого уровня активности. Сердце колибри с рубиновым горлом бьется до 1200 раз в минуту (около 20 ударов в секунду). [65]
Пищеварительная система
[ редактировать ]

Обрезать
[ редактировать ]
имеется мускулистый мешочек, У многих птиц вдоль пищевода называемый зобом . Зобы одновременно смягчают пищу и регулируют ее поток через систему, временно сохраняя ее. Размер и форма зобов у птиц весьма различны. [66] Члены семейства Columbidae , такие как голуби , производят питательное молоко , которое выкармливают детенышей путем срыгивания . [67]
провентрикулюс
[ редактировать ]Птичий желудок состоит из двух органов: преджелудка и желудка , которые работают вместе во время пищеварения . Прожелудочек представляет собой трубку в форме стержня, которая находится между пищеводом и желудком и выделяет соляную кислоту и пепсиноген в пищеварительный тракт . [67] Кислота превращает неактивный пепсиноген в активный протеолитический фермент пепсин , который разрушает специфические пептидные связи, обнаруженные в белках , с образованием набора пептидов , которые представляют собой аминокислотные цепи , которые короче, чем исходный пищевой белок. [68] [69] Желудочные соки (соляная кислота и пепсиноген) смешиваются с содержимым желудка посредством мышечных сокращений желудка. [70]
желудок
[ редактировать ]Желудок состоит из четырех мышечных полос, которые вращаются и измельчают пищу, перемещая пищу из одной области желудка в другую. Желудок некоторых видов травоядных птиц, таких как индейка и перепела, [66] содержит небольшие кусочки песка или камня, называемые гастролитами , которые птица проглатывает, чтобы облегчить процесс измельчения и выполнять функцию зубов . Использование желудочных камней - это сходство, обнаруженное между птицами и динозаврами , которые оставили гастролиты в качестве следов окаменелостей . [67]
Кишечник
[ редактировать ]Частично переваренное и измельченное содержимое желудка, теперь называемое комком, поступает в кишечник , где поджелудочной железы и ферменты кишечника завершают переваривание перевариваемой пищи. Продукты пищеварения затем всасываются через слизистую кишечника в кровь. Кишечник заканчивается через толстую кишку отверстием или клоакой , которая служит общим выходом для почечных и кишечных экскрементов, а также для откладки яиц. [71] Однако, в отличие от млекопитающих, многие птицы не выделяют объемные части (грубые корма) непереваренной пищи (например, перья, мех, фрагменты костей и семенную шелуху) через клоаку, а срыгивают их в виде пищевых гранул . [72] [73]
Питьевое поведение
[ редактировать ]Птицы пьют тремя основными способами: используя силу тяжести, сосание и язык. Жидкость также получают из пищи.
Большинство птиц не могут глотать за счет «сосущего» или «перекачивающего» действия перистальтики пищевода ( как это делают люди ) и пьют, неоднократно поднимая голову после наполнения рта, чтобы жидкость могла течь под действием силы тяжести. Этот метод обычно описывается как «потягивание» или «опрокидывание». [74] Заметным исключением является семейство голубей и голубей Columbidae ; фактически, по словам Конрада Лоренца в 1939 году:
порядок можно узнать по единственной поведенческой характеристике, а именно по тому, что при питье вода накачивается за счет перистальтики пищевода, которая происходит без исключения внутри отряда. Однако единственная другая группа, которая демонстрирует такое же поведение, Pteroclidae , помещается рядом с голубями именно по этому, несомненно, очень древнему признаку. [75]
Хотя это общее правило все еще остается в силе, с тех пор были отмечены некоторые исключения в обоих направлениях. [74] [76]
Кроме того, специализированные нектара любители , такие как нектарники ( Nectariniidae ) и колибри ( Trochilidae ), пьютс помощью выступающих рифленых или корытообразных языков, а попугаи ( Psittacidae ) лакают воду. [74]
У многих морских птиц возле глаз есть железы, которые позволяют им пить морскую воду. Излишняя соль выводится из ноздрей. Многие пустынные птицы получают необходимую им воду исключительно из пищи. Устранение азотистых отходов, таких как мочевая кислота, снижает физиологическую потребность в воде, [77] поскольку мочевая кислота не очень токсична, и поэтому ее не нужно разбавлять в таком большом количестве воды. [78]
Репродуктивная и мочеполовая системы
[ редактировать ]


У самцов птиц есть два семенника увеличиваются в сотни раз , которые в период размножения и производят сперму . [79] Семенники у птиц обычно асимметричны, у большинства птиц левое семенник больше. [80] Самки птиц в большинстве семейств имеют только один функциональный яичник (левый), соединенный с яйцеводом, хотя на эмбриональной стадии у каждой самки присутствуют два яичника. У некоторых видов птиц есть два функциональных яичника, а у киви всегда сохраняются оба. [81] [82] У птиц нет мужских добавочных желез . [83]
У большинства самцов птиц нет фаллоса . У самцов видов без фаллоса сперма хранится в семенных клубочках внутри клоакального выступа до совокупления. Во время совокупления самка отводит хвост в сторону, а самец либо садится на самку сзади или спереди (как у вязолистки ), либо приближается к ней очень близко. Затем клоаки соприкасаются, и сперма может попасть в репродуктивные пути самки. Это может произойти очень быстро, иногда менее чем за полсекунды. [84]
Сперма хранится для хранения спермы самки в канальцах в течение периода от недели до более 100 дней. [85] в зависимости от вида. Затем яйцеклетки будут оплодотворяться индивидуально по мере того, как они покидают яичники, прежде чем скорлупа кальцинируется в яйцеводе . После того как яйцо отложено самкой, эмбрион продолжает развиваться в яйце вне тела самки.

Многие водоплавающие и некоторые другие птицы, например страус и индейка , обладают фаллосом . [86] Похоже, это наследственное состояние среди птиц; большинство птиц потеряли фаллос. [87] Считается, что длина связана с конкуренцией сперматозоидов у видов, которые обычно спариваются много раз за сезон размножения; сперма, отложенная ближе к яичникам, с большей вероятностью достигнет оплодотворения. [88] [89] Более длинные и сложные фаллосы, как правило, встречаются у водоплавающих птиц, самки которых имеют необычные анатомические особенности влагалища (например, тупиковые мешочки и завитки по часовой стрелке). Эти вагинальные структуры можно использовать для предотвращения проникновения мужского фаллоса (который скручивается против часовой стрелки). У этих видов совокупление часто бывает насильственным, и сотрудничество самки не требуется; способность самки предотвращать оплодотворение может позволить ей выбрать отца для своего потомства. [89] [90] [91] [92] В отсутствие совокупления фаллос спрятан в отделении проктодеума клоаки, прямо внутри вентиляционного отверстия.
После вылупления яиц родители оказывают им разную степень заботы с точки зрения еды и защиты. Скороразвитые птицы могут самостоятельно позаботиться о себе в течение нескольких минут после вылупления; Вылупившиеся птенцы altricial беспомощны, слепы и голы и требуют длительного родительского ухода. Птенцы многих наземно-гнездящихся птиц, таких как куропатки и кулики, часто способны бежать практически сразу после вылупления; таких птиц называют гнездящимися . Однако молодые особи, гнездящиеся в норах, часто совершенно неспособны выжить без посторонней помощи. Процесс, при котором птенец приобретает перья до тех пор, пока не сможет летать, называется «оперением».
Некоторые птицы, такие как голуби, гуси и журавли с красной короной , остаются со своими партнерами на всю жизнь и могут регулярно производить потомство.
Почка
[ редактировать ]Птичьи почки функционируют почти так же, как и более тщательно изученные почки млекопитающих , но с некоторыми важными изменениями; хотя большая часть анатомии по своей конструкции осталась неизменной, в ходе их эволюции произошли некоторые важные модификации.
Трехраздельные почки располагаются на двусторонней стороне позвоночного столба и соединяются с нижними отделами желудочно-кишечного тракта. [93] В зависимости от вида птиц корковое вещество составляет около 71–80% массы почки, а мозговое вещество намного меньше и составляет около 5–15% массы. Кровеносные сосуды и другие трубки составляют остальную массу.
Уникальным для птиц является наличие двух разных типов нефронов (функциональной единицы почки): оба нефрона, как у рептилий, расположены в коре головного мозга; и нефроны млекопитающих, расположенные в мозговом веществе.Нефроны рептилий более многочисленны, но не имеют характерных петель Генле, наблюдаемых у млекопитающих. Ввиду отсутствия петли Генле у птиц способность концентрировать воду у них не сильно зависит от нее. Реабсорбция воды полностью зависит от копродеума и прямой кишки. [20]
Моча, собранная почками, опорожняется в клоаку через мочеточники, а затем в толстую кишку посредством обратной перистальтики .


Нервная система
[ редактировать ]Мозг
[ редактировать ]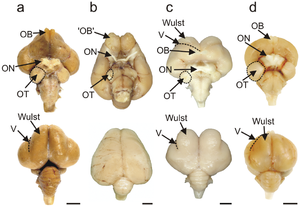
Птичий мозг — центральный орган нервной системы птиц. Птицы обладают большим и сложным мозгом , который обрабатывает , интегрирует и координирует информацию, полученную из окружающей среды, и принимает решения о том, как реагировать остальными частями тела. у всех хордовых , птичий мозг находится в костях черепа головы Как и .
Мозг птицы разделен на несколько отделов, каждый из которых выполняет свою функцию. Головной мозг или телэнцефалон разделен на два полушария и контролирует высшие функции. В конечном мозге преобладает большой мантий , который соответствует млекопитающих коре головного мозга и отвечает за когнитивные функции птиц. Паллий состоит из нескольких основных структур: гиперпаллия, дорсальной выпуклости паллия, встречающейся только у птиц, а также нидопаллия, мезопаллия и архипаллия. Ядерная структура конечного мозга птиц, в которой нейроны распределены в трехмерно расположенных кластерах, без крупномасштабного разделения белого и серого вещества , хотя существуют слоистые и столбчатые связи. Структуры мантии связаны с восприятием , обучением и познанием . Под паллием расположены два компонента субпаллия: полосатое тело и паллидум . Субпаллиум соединяет различные части конечного мозга и играет важную роль в ряде критических форм поведения. В задней части конечного мозга располагаются таламус , средний мозг и мозжечок . соединяет Задний мозг остальную часть головного мозга со спинным мозгом.
Размер и структура птичьего мозга позволяют птицам совершать заметные действия, такие как полет и вокализация . Специальные структуры и пути объединяют слуховые и зрительные чувства, сильные у большинства видов птиц, а также обычно более слабые обонятельные и тактильные чувства. Социальное поведение , широко распространенное среди птиц, зависит от организации и функций мозга. Некоторые птицы демонстрируют сильные познавательные способности, обусловленные уникальной структурой и физиологией птичьего мозга.У птиц большое соотношение массы мозга к массе тела. Это отражается в развитом и сложном интеллекте птиц .
Зрение
[ редактировать ]У птиц острое зрение — у хищников ( хищных птиц ) зрение в восемь раз острее, чем у человека — благодаря более высокой плотности фоторецепторов в сетчатке (до 1 000 000 на квадратный мм у Бутеоса по сравнению с 200 000 у человека), большому количеству нейронов в зрительных нервах — второй набор глазных мышц, отсутствующий у других животных, и, в некоторых случаях, углубленная ямка , увеличивающая центральную часть поля зрения. Многие виды, в том числе колибри и альбатросы , имеют по две ямки в каждом глазу. Многие птицы способны улавливать поляризованный свет.
Слух
[ редактировать ]Птичье ухо приспособлено улавливать небольшие и быстрые изменения высоты звука пения птиц . Общая форма барабанной перепонки птиц овулярная, слегка коническая. морфологические различия в среднем ухе Между видами наблюдаются . Косточки зеленых вьюрков , черных дроздов , певчих дроздов и домашних воробьев пропорционально короче таковых у фазанов , крякв и морских птиц . У певчих птиц сиринкс позволяет соответствующим владельцам создавать сложные мелодии и звуки. Среднее ухо птиц состоит из трех полукружных каналов, каждый из которых заканчивается ампулой и соединяется, соединяясь с макулой-саккулюсом и лагеной , от которых отходит улитка — прямая короткая трубка, идущая к наружному уху. [94]
Вкус
[ редактировать ]Птицы произошли от предка, который утратил вкусовые рецепторы типа T1R2, которые позволяют другим животным, например аллигаторам, ощущать сладкий вкус. После того, как многие птицы адаптировались к диете с высоким содержанием сахара, они модифицировали свои вкусовые рецепторы умами (T1R1-T1R3), чтобы они также могли ощущать сладкий вкус. TR2, используемый для определения горького вкуса, у птиц снижен. [95] [96]
Иммунная система
[ редактировать ]Иммунная система птиц аналогична иммунной системе других челюстных позвоночных. У птиц есть как врожденная , так и адаптивная иммунная система. Птицы подвержены опухолям , иммунодефициту и аутоиммунным заболеваниям.
Бурса Фабриция
[ редактировать ]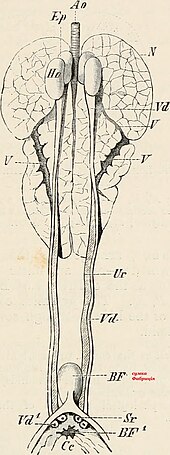
Функция
[ редактировать ]Бурса Фабрициуса , также известная как клоакальная бурса, представляет собой лимфоидный орган , который способствует выработке В-лимфоцитов во время гуморального иммунитета . Бурса Фабрициуса присутствует на ювенильной стадии, но скручивается. Например, бурса не видна после полового созревания у разных видов воробьев . [97] Для сравнения: у млекопитающих В-лимфоциты развиваются в костном мозге.
Анатомия
[ редактировать ]Бурса Фабрициуса представляет собой круглый мешочек, соединенный с верхней дорсальной стороной клоаки . Бурса состоит из множества складок, известных как складки, которые выстланы более чем 10 000 фолликулами, окруженными соединительной тканью и окруженными мезенхимой . Каждый фолликул состоит из коркового вещества, окружающего мозговое вещество. В коре находятся сильно уплотненные В-лимфоциты , тогда как в мозговом веществе лимфоциты располагаются рыхло. [97] Продолговатый мозг отделен от просвета эпителием , что способствует транспорту эпителиальных клеток в просвет бурсы. Вокруг каждого фолликула расположено 150 000 В-лимфоцитов. [98]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Ритчисон, Гэри. «Орнитология (Био 554/754): Дыхательная система птиц» . Университет Восточного Кентукки . Проверено 27 июня 2007 г.
- ^ Гир, HT (1952). «Воздушные мешки гагары» (PDF) . Аук . 69 (1): 40–49. дои : 10.2307/4081291 . JSTOR 4081291 . Проверено 21 января 2014 г.
- ^ Смит, Натан Д. (2011). «Масса тела и экология кормления предсказывают эволюционные закономерности пневматическости скелета в разнообразной кладе водоплавающих птиц» . Эволюция . 66 (4): 1059–1078. дои : 10.1111/j.1558-5646.2011.01494.x . ПМИД 22486689 . S2CID 42793145 .
- ^ Фастовский, Дэвид Э.; Вейшампель, Дэвид Б. (2005). Эволюция и вымирание динозавров (2-е изд.). Кембридж, Нью-Йорк, Мельбурн, Мадрид, Кейптаун, Сингапур, Сан-Паулу: Издательство Кембриджского университета. ISBN 978-0-521-81172-9 . Проверено 21 января 2014 г.
- ^ Безюденхаут, Эй Джей; Грёневальд, HB; Соли, Дж. Т. (1999). «Анатомическое исследование дыхательных воздушных мешков у страусов» (PDF) . Ондерстепоорт Журнал ветеринарных исследований . 66 (4). Ветеринарный институт Ондерстепорта: 317–325. ПМИД 10689704 . Проверено 21 января 2014 г.
- ^ Ведель, Мэтью Дж. (2003). «Пневматичность позвонков, воздушные мешки и физиология динозавров-зауроподов» (PDF) . Палеобиология . 29 (2): 243–255. Бибкод : 2003Pbio...29..243W . doi : 10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2 . S2CID 46619244 . Проверено 21 января 2014 г.
- ^ Jump up to: а б с д и ж Маршалл, Эй Джей (1960). Биология и сравнительная физиология птиц: Том I. Виктория, Австралия: Академическая пресса. стр. 245–251. ISBN 978-1-4832-3142-6 .
- ^ Телекоммуникации, интерактивные медиа – Образовательная Небраска. «Проект Клюв: Адаптации: Скелетная система: Шейные позвонки» . Projectbeak.org . Проверено 16 марта 2018 г.
- ^ Хогенбум, Мелисса. «Как птицы видят прямо» . Проверено 14 апреля 2018 г.
- ^ «Почему голуби кивают головой при ходьбе? Тайны повседневности: забавные научные факты из Библиотеки Конгресса» . www.loc.gov . Проверено 14 апреля 2018 г.
- ^ Jump up to: а б Трое, Николаус; Фрост, Барри (февраль 2000 г.). «Покачивание головой у голубей: насколько стабильна фаза удержания?» (PDF) . Журнал экспериментальной биологии . 203 (Часть 5): 935–940. дои : 10.1242/jeb.203.5.935 . ПМИД 10667977 .
- ^ Фрост, Би Джей (1978). «Оптокинетическая основа покачивания головой голубя». Журнал экспериментальной биологии . 74 : 187–195. CiteSeerX 10.1.1.556.8783 . дои : 10.1242/jeb.74.1.187 .
- ^ Сторер, Роберт В. (1982). «Сросшиеся грудные позвонки у птиц» . Журнал Института орнитологии имени Ямашина . 14 (2–3): 86–95. дои : 10.3312/jyio1952.14.86 . ISSN 1883-3659 .
- ^ Jump up to: а б Рашид, Дана Дж; Чепмен, Сьюзен С; Ларссон, Ханс CE; Орган, Крис Л; Бебин, Анн-Гаэль; Мерцдорф, Криста С; Брэдли, Роджер; Хорнер, Джон Р. (2014). «От динозавров к птицам: хвост эволюции» . ЭвоДево . 5 (1): 25. дои : 10.1186/2041-9139-5-25 . ISSN 2041-9139 . ПМК 4304130 . ПМИД 25621146 .
- ^ Кардонг, Кеннет В. (2019). Позвоночные животные: сравнительная анатомия, функции, эволюция (8-е изд.). Нью-Йорк, штат Нью-Йорк. стр. 313–321. ISBN 978-1-259-70091-0 . OCLC 1053847969 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Накацукаса, Масато (май 2004 г.). «Приобретение прямохождения: миоценовые записи гоминоидов и современные аналоги двуногих протогоминид» . Журнал анатомии . 204 (5): 385–402. дои : 10.1111/j.0021-8782.2004.00290.x . ISSN 0021-8782 . ПМК 1571308 . ПМИД 15198702 .
- ^ Jump up to: а б «Скелет» . fsc.fernbank.edu . Проверено 16 марта 2018 г.
- ^ «Нелетающие птицы» . СКЕЛЕТЫ: Музей остеологии (Оклахома-Сити) . Архивировано из оригинала 27 июня 2018 г. Проверено 2 мая 2018 г.
- ^ ДЮЗЛЕР, А.; ОЗГЕЛЬ, Ö.; ДУРСУН, Н. (2006). «Морфометрический анализ грудины у птиц» (PDF) . Турецкий журнал ветеринарии и зоотехники . 30 : 311–314. ISSN 1303-6181 . Архивировано из оригинала (PDF) 12 ноября 2013 г. Проверено 1 марта 2013 г.
- ^ Jump up to: а б с Бхуллар, Бхарт-Анджан С.; Маруган-Лобон, Хесус; Расимо, Фернандо; Бевер, Гейб С.; Роу, Тимоти Б.; Норелл, Марк А.; Абжанов, Архат (27 мая 2012 г.). «У птиц педоморфные черепа динозавров». Природа . 487 (7406): 223–226. Бибкод : 2012Natur.487..223B . дои : 10.1038/nature11146 . ISSN 1476-4687 . ПМИД 22722850 . S2CID 4370675 .
- ^ Лушар, Антуан; Вирио, Лоран (2011). «От морды до клюва: потеря зубов у птиц». Тенденции в экологии и эволюции . 26 (12): 663–673. Бибкод : 2011TEcoE..26..663L . дои : 10.1016/j.tree.2011.09.004 . ПМИД 21978465 .
- ^ Jump up to: а б с д Бхуллар, Бхарт-Анджан С.; Хэнсон, Майкл; Фаббри, Маттео; Причард, Адам; Бевер, Гейб С.; Хоффман, Ева (01 сентября 2016 г.). «Как сделать череп птицы: основные переходы в эволюции птичьего черепа, педоморфоза и клюва как суррогатной руки» . Интегративная и сравнительная биология . 56 (3): 389–403. дои : 10.1093/icb/icw069 . ISSN 1540-7063 . ПМИД 27371392 .
- ^ Хуан, Цзяндун; Ван, Ся; Ху, Юаньчао; Лю, Цзя; Петейя, Дженнифер А.; Кларк, Джулия А. (15 марта 2016 г.). «Новый орнитурин из раннего мела Китая проливает свет на эволюцию раннего экологического и краниального разнообразия птиц» . ПерДж . 4 : е1765. дои : 10.7717/peerj.1765 . ISSN 2167-8359 . ПМЦ 4806634 . ПМИД 27019777 .
- ^ ЛАУДЕР, ДЖОРДЖ В. (1 мая 1982 г.). «Закономерности эволюции механизма питания актиноптеригиевых рыб» . Американский зоолог . 22 (2): 275–285. дои : 10.1093/icb/22.2.275 . ISSN 1540-7063 .
- ^ Шеффер, Бобб; Розен, Донн Эрик (1961). «Основные адаптивные уровни в эволюции механизма питания актиноптеригиев» . Американский зоолог . 1 (2): 187–204. дои : 10.1093/icb/1.2.187 . JSTOR 3881250 .
- ^ Симонетта, Альберто М. (1 сентября 1960 г.). «О механическом влиянии птичьего черепа и их влиянии на эволюцию и классификацию птиц». Ежеквартальный обзор биологии . 35 (3): 206–220. дои : 10.1086/403106 . ISSN 0033-5770 . S2CID 85091693 .
- ^ Ван, М.; Стидхэм, штат Техас; Ли, З.; Сюй, Х.; Чжоу, З. (2021). «Птица мелового периода с черепом динозавра проливает свет на эволюцию черепов птиц» . Природные коммуникации . 12 (1): 3890. Бибкод : 2021NatCo..12.3890W . дои : 10.1038/s41467-021-24147-z . ПМЦ 8222284 . ПМИД 34162868 .
- ^ Хм.; Сансалоне, Г.; Роу, С.; Макдональд, П.Г.; О'Коннор, Дж. К.; Ли, З.; Сюй, Х; Чжоу, З. (2019). «Эволюция сошника и ее значение для краниального кинезиса у Паравеса» . ПНАС . 116 (39): 19571–8. Бибкод : 2019PNAS..11619571H . дои : 10.1073/pnas.1907754116 . ПМЦ 6765239 . ПМИД 31501339 .
- ^ Лингхэм-Солиар, Теагартен (30 января 1995 г.). «Анатомия и функциональная морфология крупнейшей известной морской рептилии Mosasaurus hoffmanni (Mosasauridae, Reptilia) из верхнего мела, верхнего маастрихта в Нидерландах». Фил. Пер. Р. Сок. Лонд. Б. 347 (1320): 155–180. Бибкод : 1995РСТБ.347..155Л . дои : 10.1098/rstb.1995.0019 . ISSN 0962-8436 .
- ^ Холлидей, Кейси М.; Уитмер, Лоуренс М. (2008). «Краниальный кинезис у динозавров: внутричерепные суставы, мышцы-протракторы и их значение для эволюции и функции черепа у диапсид». Журнал палеонтологии позвоночных . 28 (4): 1073–1088. Бибкод : 2008JVPal..28.1073H . дои : 10.1671/0272-4634-28.4.1073 . S2CID 15142387 .
- ^ Винг, Леонард В. (1956) Естественная история птиц . Компания Рональд Пресс.
- ^ Проктор, Н.С. и Линч, П.Дж. (1998) Руководство по орнитологии: структура и функции птиц . Издательство Йельского университета. ISBN 0300076193
- ^ Локли, МГ; Ли, Р.; Харрис, доктор юридических наук; Мацукава, М.; Лю, М. (2007). «Самые ранние зигодактильные ноги птиц: данные по следам, похожим на дорожных бегунов раннего мела» (PDF) . Naturwissenschaften . 94 (8): 657–665. Бибкод : 2007NW.....94..657L . дои : 10.1007/s00114-007-0239-x . ПМИД 17387416 . S2CID 15821251 .
- ^ Гейтси, С.М. (2002). «19. Локомоторная эволюция на пути к современным птицам» . В Кьяппе, штат Луизиана; Уитмер, Л.М. (ред.). Мезозойские птицы: над головами динозавров . Издательство Калифорнийского университета. стр. 432–447. ISBN 978-0-520-20094-4 .
- ^ Дю Брюль, Э. Ллойд (1962). «Общий феномен прямохождения» . Американский зоолог . 2 (2): 205–208. дои : 10.1093/icb/2.2.205 .
- ^ Jump up to: а б Курочкин, Е. Н. (1995). «Краткий обзор мезозойских птиц и ранней эволюции класса Aves». Археоптерикс: Ежегодный журнал Друзей музея Айхштетт Юра . 13 :47–66. ISSN 0933-288X . OCLC 85132179 .
- ^ Jump up to: а б Лонг, Калифорния; Чжан, врач общей практики; Джордж, ТФ; Лонг, CF (2003). «Физическая теория, происхождение полета и синтез, предложенные для птиц». Журнал теоретической биологии . 224 (1): 9–26. Бибкод : 2003JThBi.224....9L . дои : 10.1016/S0022-5193(03)00116-4 . ПМИД 12900201 .
- ^ Богданович И.А. (2007). «Еще раз о происхождении птиц и полета: «беглый» или «древесный»?» (PDF) . Вестник Зоологии . 41 (3): 283–4.
- ^ Jump up to: а б с д Проктор, Ноубл С., Линч, Патрик Дж. (1993). Руководство по орнитологии . Нью-Хейвен и Лондон: Издательство Йельского университета. стр. 149–170. ISBN 978-0-300-07619-6 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Пикассо, Мариана Б.Дж.; Мосто, Мария К. (2018). «Миология крыльев каракаров (Aves, Falconiformes): мышечные особенности, связанные с полетным поведением» (PDF) . Зоология позвоночных . 68 (2): 177–190.
- ^ Тобальске, Брет В. (2016). «Эволюция птичьего полета: мышцы и ограничения производительности» . Философские труды Королевского общества B: Биологические науки . 371 (1704): 20150383. doi : 10.1098/rstb.2015.0383 . ПМЦ 4992707 . ПМИД 27528773 .
- ^ Фергюсон-Лис, Джеймс; Кристи, Дэвид А. (2001). Хищники мира . Лондон: Кристофер Хелм. стр. 67–68. ISBN 978-0-7136-8026-3 .
- ^ Тарботон, Уорик; Эразмус, Руди (1998). Совы и совы в Южной Африке . Кейптаун: Издательство Струик. п. 10. ISBN 1-86872-104-3 .
- ^ Оберприлер, Ульрих; Силли, Бургер (2002). Руководство по идентификации хищников для Южной Африки . Парклендс: Случайный дом. п. 8. ISBN 978-0-9584195-7-4 .
- ^ Лукас, Альфред М. (1972). Анатомия птиц – покровы . Ист-Лансинг, Мичиган, США: Проект анатомии птиц Министерства сельского хозяйства США, Университет штата Мичиган. стр. 67, 344, 394–601.
- ^ Сойер, Р.Х.; Кнапп, Л.В. (2003). «Развитие птичьей кожи и эволюционное происхождение перьев». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 298 (1): 57–72. Бибкод : 2003JEZB..298...57S . дои : 10.1002/jez.b.26 . ПМИД 12949769 .
- ^ Jump up to: а б с Дуайи, Д. (2009). «Новый сценарий эволюционного происхождения волос, перьев и птичьей чешуи» . Дж. Анат . 214 (4): 587–606. дои : 10.1111/j.1469-7580.2008.01041.x . ПМК 2736124 . ПМИД 19422430 .
- ^ Чжэн, X.; Чжоу, З.; Ван, X.; Чжан, Ф.; Чжан, X.; Ван, Ю.; Сюй, X. (2013). «Задние крылья базальных птиц и эволюция перьев ног». Наука . 339 (6125): 1309–12. Бибкод : 2013Sci...339.1309Z . CiteSeerX 10.1.1.1031.5732 . дои : 10.1126/science.1228753 . ПМИД 23493711 . S2CID 206544531 .
- ^ Штеттенхайм, Питер Р. (2000). «Покровная морфология современных птиц — обзор» . Американский зоолог . 40 (4): 461–477. дои : 10.1093/icb/40.4.461 .
- ^ Пирсма, Теунис; ван Алст, Рене; Курк, Карин; Берхудт, Герман; Лео Р.М. Маас (1998). «Новый механизм восприятия давления для обнаружения добычи у птиц: использование принципов динамики морского дна?» . Труды: Биологические науки . 265 (1404): 1377–83. дои : 10.1098/rspb.1998.0445 . ПМЦ 1689215 .
- ^ Зуси, Р.Л. (1984). «Функциональный и эволюционный анализ ринхокинеза у птиц». Смитсоновский вклад в зоологию . 395 (395): 1–40. дои : 10.5479/si.00810282.395 . hdl : 10088/5187 .
- ^ Колдер, Уильям А. (1996). Размер, функция и история жизни . Минеола, Нью-Йорк: Публикации Courier Dove. п. 91. ИСБН 978-0-486-69191-6 .
- ^ Jump up to: а б с Майна, Джон Н. (2005). Система легких воздушных мешков развития, строения и функций птиц; с 6 столами . Берлин: Шпрингер. стр. 3.2–3.3 «Легкие», «Система дыхательных путей (бронхиол)» 66–82. ISBN 978-3-540-25595-6 .
- ^ Jump up to: а б с Краутвальд-Юнгханнс, Мария-Элизабет; и др. (2010). Диагностическая визуализация экзотических домашних животных: птиц, мелких млекопитающих, рептилий . Германия: Издательство Manson. ISBN 978-3-89993-049-8 .
- ^ Нгуен, Куинь М.; Оза У., Ананд; Абуэзи, Джоанна; Сунь, Гуаньхуа; Чилдресс, Стивен; Фредерик, Кристина; Ристроф, Лейф (19 марта 2021 г.). «Выпрямление потока в петлевых сетевых моделях легких птиц» . Письма о физических отзывах . 126 (11): 114501. arXiv : 2103.11237 . Бибкод : 2021PhRvL.126k4501N . doi : 10.1103/PhysRevLett.126.114501 . ПМИД 33798375 . S2CID 232307000 .
- ^ Jump up to: а б с д Уиттоу, Дж. Кози (2000). Птичья физиология Стерки . Сан-Диего, Калифорния: Academic Press. стр. 233–241 . ISBN 978-0-12-747605-6 .
- ^ Майна, Джон Н. (ноябрь 2006 г.). «Развитие, строение и функция нового органа дыхания, системы легких и воздушных мешков птиц: идти туда, куда не ступало ни одно другое позвоночное животное» . Биологические обзоры . 81 (04): 545. doi : 10.1017/S1464793106007111 . ISSN 1464-7931 .
- ^ Майна, JN (октябрь 2008 г.). «Функциональная морфология дыхательной системы птиц, системы легкое – воздушный мешок: эффективность, основанная на сложности» . Страус . 79 (2): 117–132. дои : 10.2989/страус.2008.79.2.1.575 . ISSN 0030-6525 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Ричсон, Г. «BIO 554/754 - Орнитология: дыхание птиц» . Департамент биологических наук Университета Восточного Кентукки . Проверено 23 апреля 2009 г.
- ^ Jump up to: а б Стерки, П.Д. (1976). Стерки, П.Д. (ред.). Птичья физиология . Нью-Йорк: Springer Verlag. п. 201. дои : 10.1007/978-1-4612-4862-0 . ISBN 978-1-4612-9335-4 . S2CID 36415426 .
- ^ Jump up to: а б с д Скотт, Грэм Р. (2011). «Комментарий: Повышенная производительность: уникальная физиология птиц, летающих на больших высотах» . Журнал экспериментальной биологии . 214 (15): 2455–2462. дои : 10.1242/jeb.052548 . ПМИД 21753038 .
- ^ «Птичьи легкие» . Архивировано из оригинала 11 марта 2007 года.
- ^ Гросс, ВБ (1 июля 1964 г.). «Произведение голоса курицей» . Птицеводство . 43 (4): 1005–1008. дои : 10.3382/ps.0431005 . ISSN 0032-5791 .
- ^ Синн-Хэнлон, Джанет. «Сравнительная анатомия куриного сердца» . Университет Иллинойса.
- ^ Осборн, июнь (1998 г.). Колибри с рубиновым горлом . Издательство Техасского университета. п. 14 . ISBN 978-0-292-76047-9 .
- ^ Jump up to: а б Орнитология, Корнельская лаборатория. «Все об анатомии птиц от Академии птиц» . academy.allaboutbirds.org . Проверено 11 мая 2018 г.
- ^ Jump up to: а б с Захер, Мостафа (2012). «Анатомические, гистологические и гистохимические адаптации пищеварительного тракта птиц к их пищевым привычкам: I-Coturnix coturnix» . Журнал наук о жизни . 9 : 253–275.
- ^ Страйер, Люберт (1995). В: Биохимия (4-е изд.). Нью-Йорк: WH Freeman. стр. 250–1. ISBN 0-7167-2009-4 .
- ^ Моран, Эдвин (2016). «Желудочное переваривание белка посредством действия панкреозимов оптимизирует кишечные формы для всасывания, образования муцина и целостности ворсинок» . Наука и технология кормов для животных . 221 : 284–303. doi : 10.1016/j.anifeedsci.2016.05.015 .
- ^ Свихус, Биргер (2014). «Функция пищеварительной системы» . Журнал прикладных исследований птицеводства . 23 (2): 306–314. дои : 10.3382/январь 2014-00937 .
- ^ Сторер, Трейси И.; Узергер, Р.Л.; Стеббинс, Роберт С.; Нибаккен, Джеймс В. (1997). Общая зоология (6-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 750–751 . ISBN 978-0-07-061780-3 .
- ^ Тарботон, Уорик; Эразмус, Руди (1998). Совы и совы в Южной Африке . Кейптаун: Издательство Струик. стр. 28–29. ISBN 1-86872-104-3 .
- ^ Кемп, Алан; Кемп, Мэг (1998). Сасоль Хищные птицы Африки и ее островов . Лондон: New Holland Publishers (UK) Ltd., с. 332. ИСБН 1-85974-100-2 .
- ^ Jump up to: а б с Кейд, Том Дж.; Гринвальд, Льюис И. (1966). «Питьевое поведение мышевидных птиц в пустыне Намиб, Южная Африка» (PDF) . Аук . 83 (1): 126–8. дои : 10.2307/4082984 . JSTOR 4082984 .
- ^ К. Лоренц, Переговоры. Немецкий. Зоол. Гес., 41 [Зоол. Нет. Приложение 12]: 69–102, 1939.
- ^ Кейд, Том Дж.; Уиллоуби, Эрнест Дж.; Маклин, Гордон Л. (1966). «Питьевое поведение рябчиков в пустынях Намиб и Калахари, Африка» (PDF) . Аук . 83 (1): 124–6. дои : 10.2307/4082983 . JSTOR 4082983 .
- ^ Маклин, Гордон Л. (1996). Экофизиология пустынных птиц . Спрингер. ISBN 3-540-59269-5 .
- ^ Элфик, Джонатан (2016). Птицы: полное руководство по их биологии и поведению . Буффало, Нью-Йорк: Firefly Books. стр. 53–54. ISBN 978-1-77085-762-9 .
- ^ Исследование сезонных изменений в семенниках птиц. Архивировано 16 марта 2009 г. в Wayback Machine. Александр Уотсон, J. Physiol. 1919;53;86–91, «зеленушка ( Carduelis chromis )», «В начале лета (май и июнь) они размером с целую горошину, а в начале зимы (ноябрь) не больше булавочной головки»
- ^ Лейк, Пенсильвания (1981). «Мужские половые органы». В Кинге АС; Маклелланд Дж. (ред.). Форма и функции у птиц . Том. 2. Нью-Йорк: Академ. стр. 1–61. ISBN 978-0-12-407502-3 .
- ^ Кински, ФК (1971). «Постоянное наличие парных яичников у киви (Apteryx) с некоторым обсуждением этого состояния у других птиц». Журнал орнитологии . 112 (3): 334–357. Бибкод : 1971JOrni.112..334K . дои : 10.1007/BF01640692 . S2CID 28261057 .
- ^ Фитцпатрик, Флорида (1934). «Односторонние и двусторонние яичники хищных птиц» (PDF) . Бюллетень Уилсона . 46 (1): 19–22. JSTOR 4156262 .
- ^ Хайман, Либби Генриетта (15 сентября 1992 г.). Сравнительная анатомия позвоночных Хаймана . Издательство Чикагского университета. ISBN 978-0-226-87013-7 .
- ^ Линч, Уэйн; Линч, фотографии Уэйна (2007). Совы США и Канады: полное руководство по их биологии и поведению . Балтимор: Издательство Университета Джонса Хопкинса. п. 151. ИСБН 978-0-8018-8687-4 .
- ^ Биркхед, TR; А. П. Моллер (1993). «Половой отбор и временное разделение репродуктивных событий: данные о хранении спермы у рептилий, птиц и млекопитающих». Биологический журнал Линнеевского общества . 50 (4): 295–311. дои : 10.1111/j.1095-8312.1993.tb00933.x .
- ^ Джеймисон, генеральный менеджер Барри (14 октября 2011 г.). Репродуктивная биология и филогения птиц, Часть A: Филогения, морфология, гормоны и оплодотворение . ЦРК Пресс. ISBN 978-1-4398-4275-1 .
- ^ Эррера, AM; Шустер, С.Г.; Перритон, CL; Кон, MJ (2013). «Основы развития редукции фаллоса в ходе эволюции птиц» . Современная биология . 23 (12): 1065–74. Бибкод : 2013CBio...23.1065H . дои : 10.1016/j.cub.2013.04.062 . ПМИД 23746636 .
- ^ Маккракен, КГ (2000). «20-сантиметровый колючий пенис аргентинской озерной утки (Oxyura vittata)» (PDF) . Аук . 117 (3): 820–5. doi : 10.1642/0004-8038(2000)117[0820:TCSPOT]2.0.CO;2 . S2CID 5717257 .
- ^ Jump up to: а б Арнквист, Г.; И. Даниэльссон (1999). «Копулятивное поведение, морфология гениталий и успех мужского оплодотворения у водомерок». Эволюция . 53 (1): 147–156. дои : 10.2307/2640927 . JSTOR 2640927 . ПМИД 28565197 .
- ^ Эберхард, В. (2010). «Эволюция гениталий: теории, доказательства и новые направления». Генетика . 138 (1): 5–18. дои : 10.1007/s10709-009-9358-y . ПМИД 19308664 . S2CID 1409845 .
- ^ Хоскен, диджей; П. Стокли (2004). «Половой отбор и генитальная эволюция» (PDF) . Тенденции в экологии и эволюции . 19 (2): 87–93. CiteSeerX 10.1.1.509.2660 . дои : 10.1016/j.tree.2003.11.012 . ПМИД 16701234 . Архивировано из оригинала (PDF) 12 октября 2017 г. Проверено 26 августа 2018 г.
- ^ Бреннан, PLR; РО Прум; К.Г. Маккракен; доктор медицинских наук Соренсон; Р.Э. Уилсон; Т. Р. Биркхед (2007). «Коэволюция морфологии мужских и женских половых органов водоплавающих птиц» . ПЛОС ОДИН . 2 (5): е418. Бибкод : 2007PLoSO...2..418B . дои : 10.1371/journal.pone.0000418 . ПМК 1855079 . ПМИД 17476339 .
- ^ Лиерц, Майкл (январь 2003 г.). «Заболевания почек птиц: патогенез, диагностика и терапия». Ветеринарные клиники Северной Америки: практика экзотических животных . 6 (1): 29–55. дои : 10.1016/S1094-9194(02)00029-4 . ISSN 1094-9194 . ПМИД 12616833 .
- ^ Миллс, Роберт (март 1994 г.). «Прикладная сравнительная анатомия среднего уха птиц» . Журнал Королевского медицинского общества . 87 (3): 155–6. дои : 10.1177/014107689408700314 . ПМЦ 1294398 . ПМИД 8158595 .
- ^ Никнафс, Шахрам; Наварро, Марта; Шнайдер, Ева Р.; Роура, Евгений (2023). «Система птичьего вкуса» . Границы в физиологии . 14 : 1235377. doi : 10.3389/fphys.2023.1235377 . ПМЦ 10516129 . PMID 37745254 .
- ^ «Птицы потеряли пристрастие к сладкому, колибри вернули себе» . Национальное географическое общество . 21 августа 2014 г.
- ^ Jump up to: а б Андерсон, Тед Р. (2006). Биология вездесущего домового воробья: от генов к популяциям . Нью-Йорк: Издательство Оксфордского университета. стр. 390 . ISBN 978-0-19-530411-4 . OCLC 922954367 .
- ^ Надь, Н; Мадьяр, А (1 марта 2001 г.). «Развитие фолликул-ассоциированного эпителия и секреторных дендритных клеток в фабричной сумке цесарки (Numida meleagris), изученное с помощью новых моноклональных антител» . Анатомическая запись . 262 (3): 279–292. doi : 10.1002/1097-0185(20010301)262:3<279::aid-ar1038>3.0.co;2-i . ПМИД 11241196 .
Внешние ссылки
[ редактировать ]- Дыхательная система и органы дыхания птиц.
- Черепа и скелеты птиц
- Дыхательная система птиц
- Гистология дыхательной системы птиц

| Базы данных органов управления : Национальные |
|---|