Морфология двукрылых
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
Морфология двукрылых существенно отличается от более широкой морфологии насекомых . Двукрылые — очень большой и разнообразный отряд , состоящий в основном из насекомых малого и среднего размера. У них выдающиеся сложные глаза на подвижной голове и (максимум) одна пара функциональных перепончатых крыльев. [1] которые прикрепляются к сложному среднегруди. Вторая пара крыльев, на заднегруди, редуцирована до жужжальца . Фундаментальной особенностью отряда является его замечательная специализация с точки зрения формы крыльев и морфо-анатомической адаптации грудной клетки - особенности, которые придают особую подвижность его летающим формам. Нитевидные, стильчатые или аристатные усики коррелируют с таксонами Nematocera , Brachycera и Cyclorrhapha соответственно. [2] Он обнаруживает значительное морфологическое единообразие у низших таксонов , особенно на уровне рода и вида . Принципиальное значение в их систематике имеет конфигурация покровных щетинок, а также жилкование крыльев. Проявляет полный метаморфоз (яйцо, личинка, куколка, взрослая особь). [2] или голометаболическое развитие. [1] Личинки безногие, у Nematocera имеют головные капсулы с нижнечелюстным ротовым аппаратом. Однако личинки «высших мух» (Brachycera) безголовы, червеобразны и имеют только три возраста. Куколки у Nematocera закрытые, у Brachycera — грубые. [1] [2]
Взрослый
[ редактировать ] | 1 : охраняется ; 2 : переднее дыхалец (стигма) ; 3 : щит ; 4 : базовая стоимость ; 5 : калиптра ; 6 : блюдо ; 7 : крыловидный нерв (рёбра) ; 8 : крыло ; 9 : сжечь ; 10 : держись ; 11 : заднее дыхалец (стигма) ; 12 : бедра ; 13 : флейта ; 14 : шпора ; 15 : предплюсна ; 16 : проплевра ; 17 : ниц ; 18 : мезоплевра ; 19 : среднегрудинная кость ; 20 : метаплевра ; 21 : заднегруди ; 22 : сложный глаз ; 23 : хребет ; 24 : антенна ; 25 : верхнечелюстные щупики ; 26 : губа (нижняя) ; 27 : тазик ; 28 : псевдотрахея ; 29 : тип. |
Взрослые особи маленькие (< 2 millimetres (5⁄64 in)) to medium-sized insects (- < 10 millimetres (25⁄64 in)). Larger Diptera are rare, only certain families of Diptera Mydidae and Pantophthalmidae reach Размах крыльев 95–100 миллиметров ( 3 + 3 ⁄ 4 –4 дюйма), в то время как тропические виды Tipulidae были зарегистрированы с размахом крыльев более 100 миллиметров (4 дюйма). Они имеют тусклые или яркие цвета, однородные или пестрые, а иногда имитируют, как, например, у Syrphidae . Принципиальное значение для систематики имеет наличие и расположение прикреплённых покровных щетинок .
Внешний
[ редактировать ]Голова
[ редактировать ]В этом разделе есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|

Голова отличается от груди, с заметным сужением на шее. У «низших мух» ( Nematocera ) она прогнатная (голова ориентирована горизонтально ртом вперед), у «высших мух» ( Brachycera ) гипогнатная (голова ориентирована вертикально ртом вентрально).Форма черепной капсулы также варьируется. У Nematocera дорсально-вентральная часть головы выдвигается вперед от глаз за счет развития в длину наличника и субгенальной области (subgena), дистальный конец расширения - «ротовой аппарат».


У «высших» двукрылых голова имеет почти шаровидную форму, а лобно-клипеус представляет собой область, ограниченную сверху глазами и теменем. У Cyclorrhapha Schizophora особенно важным морфологическим элементом является наличие птилинального шва, образующегося в результате резорбции птилина после выхода из куколки. Шов разделяет две области: 1. верхняя — лобная область, имеющая непрерывность с вершиной, орбитальной областью и щекой, 2. нижняя — лицо или наличник , содержит место прикрепления усиков и заканчивается эпистомальный край, образующий верхнюю губу.
Глаза обычно очень заметны, но у Brachycera они достигают замечательного развития. У этого подотряда глаза заметно выпуклые и занимают большую часть головы. Пространство между двумя глазами иногда может сокращаться до узкой полосы, идущей от передней части затылочной области, или вовсе исчезать из-за непосредственного контакта глаз или их краев. Морфология сложного глаза характеризуется значительным количеством омматидий , порядка тысяч у мускоидов . Глазки, если они есть, расположены на макушке головы и расположены по углам треугольника в области, называемой стволматикум или глазковый треугольник.
Для систематики важно наличие, расположение и форма головных щетинок, и они имеют специфическую терминологию. Щетинки на голове: лобные щетинки, иногда называемые нижнеорбитальными, расположены на лобных пластинках лба, напоминающие небольшую аллею, идущую от основания усиков к темени и окаймляющую с боков срединную лобную полосу. Иногда они располагаются ниже, вдоль лобной части, ниже места прикрепления усиков и на большем или меньшем расстоянии. Орбитальные щетинки расположены на вершинных пластинках лба и обычно ограничены его верхней половиной. Они могут быть расположены продольными рядами, называемыми внутренними и внешними щетинками глазницы. (Лобную и теменную пластинки лба можно визуализировать по расположению лобных и орбитальных щетинок); глазковые щетинки расположены на вершине между глазками; наружная и внутренняя вертикальные щетинки расположены на границе между макушкой и затылком и возле верхнего угла глаз; поствертикальные щетинки расположены позади глазков на затылке, вблизи срединной линии головы; вибриссы обычно располагаются в небольшом количестве вдоль лицевых отделов дугообразного шва, у края ротовой полости; иногда они поднимаются по шву на большее или меньшее расстояние, иногда почти до места прикрепления усиков; ложные вибриссы-щетинки располагаются по краю ротовой полости.
Иногда терминология противоречива. Например, у Acalyptraae обычно имеются две щетинки, более или менее крепкие, расположенные вдоль заднего края глазкового треугольника. Эти щетинки называются «поствертикальными щетинками» в старой литературе, начиная с девятнадцатого века, и этот термин иногда используется в современной литературе. Стейскал (1976) предложил название «постоцеллярная щетина», принятый термин в « Руководстве по неарктическим двукрылым» (McAlpine, JF, 1981) и « Руководстве по палеарктическим двукрылым» (Бернхард Мерц, Жан-Поль Хэнни, 2000) и, следовательно, этот термин широко встречается в литературе, посвященной этим двум фундаментальным работам. Две другие щетинки, присутствующие только у некоторых семейств Acalyptratae , расположены сзади и латерально от глазкового треугольника и в старой литературе называются «внутренними затылочными». Стейскал (1976) использует название «паравертикальные щетинки», и это же название используется в основной номенклатуре двух цитируемых руководств. По-русски боковые части лба называются орбитами. В английском языке эту часть чаще всего называют «фронталией», «парафронталией» или «лобной орбитой», тогда как простой термин «орбита» относится к краю сложного глаза. Срединная часть нижней части головы, или лица, часто несет в нижних углах пару (или несколько пар) крупных щетинок (щетинок), называемых вибриссами, а иногда несколько или даже целый ряд вдоль гребня, идущего вверх от затылка. вибриссы. Последние щетинки в русских текстах называются еще вибриссальными щетинками, а не лицевыми щетинками.
Усики делятся на два основных морфологических типа , которые лежат в основе различия между двумя подотрядами. Все усики мух состоят из трех отделов: скапуса, ножки и жгутика. Собственные мышцы имеются только на скапусе и ножке. [3] У Nematocera они многочленные, нитевидные или перистые, состоят из 7—15 недифференцированных экземпляров. У Brachycera усики состоят из до шести члеников, из которых первые три хорошо развиты. У большинства семейств Brachycera третий сегмент увеличен, а более апикальные сегменты редуцированы до придатка, называемого стилусом, когда он жесткий, и аристой, когда он похож на щетину.
Ротовой аппарат насекомых в зависимости от систематической группы имеет различное строение. Ротовой аппарат видоизменён и объединен в сосательный хоботок, весьма изменчивый по строению. Предковым состоянием является хобот колюще-сосущего типа, более видоизмененный хобот образует различные рашпильные или губчатые жидкости. Лабеллум . является одним из таких губчатых органов У некоторых видов нефункциональный ротовой аппарат взрослых особей.
 Nematocera: голова Culicidae . Перистые нитевидные усики, пронзающие присасывающиеся ротовые части.
Nematocera: голова Culicidae . Перистые нитевидные усики, пронзающие присасывающиеся ротовые части. Брахицера: Muscoidea . Антенна с аристой
Брахицера: Muscoidea . Антенна с аристой Брахицера: Muscoidea. Сосущий ротовой аппарат
Брахицера: Muscoidea. Сосущий ротовой аппарат



грудная клетка
[ редактировать ]| Морфология грудной клетки мускоидных двукрылых праволатеральный (слева), спинной (справа) | |
 |  |
| 1 : мезопрескутум; 2 : плечевая мозоль; 3 : нотоплейрон; 4 : среднеспинка; 5 : задняя мозоль; 6 : мезоскутеллюм; 7 : поперечный шов; 8 : заднещитик; 9 : заднеспинка; 10 : база крыла; 11 : мезоплевротергит , или латеротергит, или кататергит; 12 : недоуздок ; 13 : стигма; 14 : метаплевры ; 15 : метакокса; 16 : гипоплеврон или мерон ; 17 : мезококса; 18 : эпимеральный шов; 19 : мезоэпимерон или анэпимерон; 20 : вентрально мезоэпистернум или катепистерно; 21 : шов эпистерно-прекоксальный; 22 : прококса; 23 : плевральный шов ; 24 : дорсальный мезоэпистерн или анэпистерн; 25 : проплейрон. Хетотаксия (щетинки): а : акростикальная; dc : дорсоцентральный; ф : посмертный; ом : плечевой; ps : предшовный; НП : нотоплевральный; ia : интралярный (постшовный); sa : супралярный (постшовный); па : посталярный; psct : щиток. Терминология McAlpine по сравнению с другой терминологией Эквиваленты: – постпронотум = плечевая мозоль или плечевая кость; анэпистернум = мезоплеврон; проэпистернум = проплеврон; проэпимерон = нет эквивалента ; анэпимерон = птероплеврон; katepisternum = грудиноплеврон; катепимерон = нет эквивалента ; мерон = гипоплеврон; большая ампула = нет эквивалента ; латеротергит = нет эквивалента ; медиотергит = нет эквивалента ; постпронотум = плечевая мозоль или плечевая кость; анэпистернум = мезоплеврон; проэпистернум = проплеврон; проэпимерон = нет эквивалента ; анэпимерон = птероплеврон; katepisternum = грудиноплеврон; катепимерон = нет эквивалента ; мерон = гипоплеврон; большая ампула = нет эквивалента ; латеротергит = нет эквивалента ; медиотергит = нет эквивалента | |
Таксономически важные щетинки на груди.
[ редактировать ]
Плевры: 4. проплевры, 5. мезоплевры, 6. метаплевры.
Грудина: 7. переднегруди, 8. мезостернум, 9. заднегруди.
- акростальные ( а ) щетинки, прилежащие к срединной продольной оси щитка. Они могут быть нерегулярными или располагаться в два или более рядов. Важны число рядов, количество щетинок в каждом ряду, размер и толщина. Во многих группах акростальные щетинки заменены щетинками или волосками.
- прескутеллярный ( psc ) — две акростальные щетинки, более развитые, чем остальные акростальные щетинки, прикрепляющиеся впереди предщитовидного шва.
- дорсоцентрально ( dc ) эти щетинки расположены вдоль двух рядов, прилегающих к акросталиям и за их пределами.
- постплечевые ( ph ) щетинки выровнены в предшовной области и параллельны шву, отделяющему щиток от плечевых мозолей.
- плечевые ( hm ), иногда называемые постпронотальными щетинками, находятся на плечевых мозолях.
- предшовный ( пс )
- нотоплевральные ( np ) щетинки на нотоплевроне
- интралярные (постшовные) ( ia ) более или менее правильно расположенные щетинки вблизи дорсоцентрального ряда. Позиция четко не определена.
- надшовные (постшовные) ( sa ) эти щетинки ограничены в количестве и расположены от преалярной мозоли до надсупралярной области.
- посталярные ( па ) ограничены в количестве, располагаются на посталалярной мозоли у латерального края щитка позади места прикрепления крыла.
- щитковые ( psct ) щетинки на щитике. Они могут быть краевыми или на дорсальной стороне щитка (так называемого диска).
плевры Таксономическое значение имеет также хетотаксия . Признаками, принимаемыми во внимание, являются наличие или отсутствие, количество и положение щетинок и групп волосков на коже.
- anepisternum или mesopleuron - анэпистернальные или мезоплевральные щетинки
- katepisternum или sternopleuron - катепистернальные или грудиноплевральные щетинки.
- проэпистернум и проэпимерон - проэпистернальные и проэпимеральные щетинки, или проплевральные щетинки
- анэпимерон (pteropleuron) – анэпимеральные или птероплевральные щетинки.
- мерон (гипоплеврон) – меральные или гипоплевральные щетинки.
Фундаментальной особенностью двукрылых является замечательная эволюционная специализация, достигающаяся в форме крыльев и морфоанатомической адаптации грудной клетки. За исключением редких бескрылых форм, двукрылые обычно крылаты и используют крылья как основное средство передвижения.
Крылья
[ редактировать ]Уровень специализации – анатомической, функциональной и морфологической – таков, что в целом эти насекомые летают, зачастую исключительно хорошо, с особым акцентом на ловкость. Все двукрылые оснащены только одной парой функциональных крыльев, находящихся на среднегруди (спереди). Крылья на заднегруди преобразованы в жужжальца или коромысла. От этой характеристики произошло название отряда, от греческого dipteros , что означает «два крыла». Вследствие такого морфологического строения среднегруди представляет собой сегмент большего развития и сложности, тогда как переднегруди и заднегруди значительно редуцированы.
Жужжальца — это булавовидные органы, используемые для балансировки насекомого в полете и состоящие из проксимальной части, соединенной с механосенсорным органом. Гомологию между крыльями и жужжальцами демонстрирует четырехкрылый мутант плодовой мушки Drosophila melanogaster . Развитие жужжальца варьирует в зависимости от систематической группы: у Tipulidae они тонкие, но длинные и хорошо заметные, а у большинства других групп обычно скрыты крыльями. У Calyptraae, к которому относятся наиболее развитые двукрылые, жужжальца защищены калиптрами (небольшими перепонками над жужжальцами).
Среднегрудное крыло целиком перепончатое, совершенно прозрачное и бесцветное или несет зональную пигментацию, полезную для распознавания. Его поверхность разделена на три области: наиболее развитой является крыловая (основная лётная) область, поддерживаемая крепкими жилками крыльев; сзади — анальная область; и, наконец, в заднем отделе, проксимальном, имеется расширенная лобовидная алула. Алула, также называемая подмышечной долей, представляет собой широкую лопасть на проксимальном заднем крае стебля крыла. Он продолжается с верхним суппортом и дистально (обычно) отделен от крыла углублением, называемым алюлярным вырезом. Алули — это недавно приобретенный признак двукрылых (Hennig, 1973), а алюли обычно отсутствуют или слабо развиты у Nematocera (за исключением Anisopodidae), но присутствуют и относительно велики у Brachycera. У высших двукрылых между алулой и грудной клеткой находится верхняя калиптра, а также тегула . Калиптры находятся чуть ниже места соединения крыла с грудной клеткой и являются частью подмышечной перепонки крыльев некоторых двукрылых - две базальные доли называются калиптерами (также называемыми чешуйками, чешуйками). Проксимальная доля называется нижним калиптером (или базалиптером, или чешуйкой грудной клетки). Он возникает из борозды между щитиком и постнотумом в виде узкой перепончатой связки и заканчивается там, где более дистальная доля, называемая верхним калиптером (или дистикалиптером, или чешуйчатой костью), резко загибается над ней (калиптральная складка). Верхний суппорт обычно больше нижнего, но в некоторых группах ( Tabanidae , Acroceridae и многие Calyptraae), нижний чашелистик крупнее верхнего. Калиптральная бахрома представляет собой бахрому волосков вдоль заднего края каждого калиптера. Тегула [4] - это самая проксимальная пластинка у основания реберного края (также называемая реберной пластинкой или погоном). Рядом с ним (дистально) находится басикоста.
Система жилкования упрощена, но является представителем системы Комстока-Нидхэма , которая была задумана в конце девятнадцатого века для точного определения терминологии морфологии крыльев насекомых. У двукрылых жилки крыльев реберные, подреберные, радиальные, медиальные и локтевые. Кроме того, имеются две анальные вены, из которых вторая, называемая также подмышечной, отделяет анальную область от алулы. Детали жилок крыльев, поперечные жилки и форма клеток являются важными характеристиками для определения таксономических групп, в том числе на видовом уровне.



Таксономически важные условия жилкования крыльев
[ редактировать ]- ребра ( C ), подкоста ( Sc ), лучевая кость ( R ), средняя часть ( M ), кубит ( Cu ) и анальная вена ( A ). Было много разногласий относительно гомологии жилок позади подкосты у двукрылых, и некоторые авторы просто нумеруют продольные жилки спереди назад. Помимо продольных жилок имеются три основные поперечные жилки, обычно называемые поперечными жилками. Это внутренняя или передняя поперечная жилка, задняя или дисковая поперечная жилка и анальная поперечная жилка. Основными клетками крыла, используемыми в классификации, являются дискальные, анальные и вторые базальные клетки.
Наиболее часто встречающиеся термины, используемые в идентификационных ключах Diptera :
- клеточная чашка . Также называется задней кубитальной клеткой и часто называется анальной клеткой. Важным признаком является форма клеточной чашки.
- реберный разрыв . Они часто встречаются, особенно у Schizophora. Они представляют собой ослабления косты, их количество от одного до трех. Обычно эти разрывы располагаются проксимальнее места прикрепления подреберья. Чаще всего разрыв происходит проксимальнее места соединения подкосты с ребром или воображаемой точки, где она могла бы присоединиться к ребре в случае завершения. Подобный разрыв, который встречается почти так же часто, расположен немного дистальнее плечевой поперечной вены (поперечная вена h). Третий, гораздо менее частый, разрыв находится немного проксимальнее поперечной жилки h, обозначая короткий, утолщенный, сильно щетинистый участок у основания ребра (костагиума Сеги ). Переломы называются реберными, плечевыми hb и подреберными sb . Реберные разрывы у Nematocera и нижних Brachycera редки. Считается, что они являются точками сгибания крыльев во время полета, а также связаны со складками куколки на крыле.
- субкоста . Вторая продольная жилка крыла, позади ребра. Он может достичь Косты, исчезнуть раньше Косты или присоединиться к R1 до того, как достигнет Косты. см. [1] (= вспомогательная жилка многих авторов)
- дисковая клетка . Закрытая ячейка (d, dm) в центре крыла, окаймленная медиальными жилками и закрытая поперечной жилкой ММ у многих Nematocera. Если она ограничена жилкой CuA1 и закрыта поперечной жилкой DM-Cu (как у большинства нижних Brachycera и всех высших двукрылых), то это строго диско-медиальная клетка (но ее все же часто называют дисковой клеткой).
- базальная радиальная или первая базальная (br) клетка. Эта клетка спереди отграничена базальным отделом лучевой кости и радиальным сектором, дистально — радиально-медиальной поперечной жилкой.
- базально-медиальная или вторая базальная (bm) клетка. Эта клетка ограничена спереди базальным отделом задней медии и дистально медиально-локтевой поперечной жилкой.
- субапикальная клетка . Клетка между жилками R4+5, M1+2, rm и апикальным краем крыла. Если M1+2 заканчивается на краю крыла, субапикальная ячейка открыта. Если M1+2 изгибается в сторону R4+5 и соединяется с R4+5 перед краем крыла, ячейка закрывается.
- анальная жилка. Могут быть одна или две анальные жилки (затем А1 и А2), доходящие или не доходящие до края крыла.
Щиток почти всегда отчетливый, но намного меньше мезоскутума (и находится непосредственно позади него). Макрохеты щитка играют важную роль в систематике.
 Схема радиальных вилок Limoniidae : Nematocera
Схема радиальных вилок Limoniidae : Nematocera цилиндрротомы Крыльевые жилки : Nematocera.
цилиндрротомы Крыльевые жилки : Nematocera.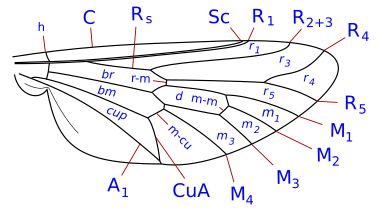 Tabanidae Жилки крыльев : Brachycera.
Tabanidae Жилки крыльев : Brachycera.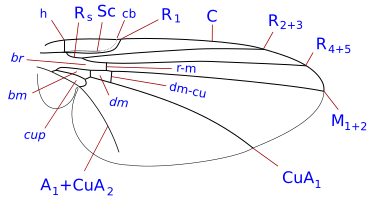 Phytomyzinae Крыльевые жилки
Phytomyzinae Крыльевые жилки Phoridae (редуцированное жилкование) Крыльевые жилки
Phoridae (редуцированное жилкование) Крыльевые жилки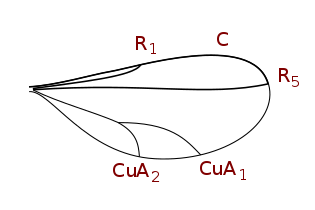 Cecidomyiinae жилкование крыльев (редуцированное жилкование)
Cecidomyiinae жилкование крыльев (редуцированное жилкование)






Относительно тонкие ножки имеют точно расположенные щетинки, которые также участвуют в хетотаксии. Бедра и голени могут нести комбинации дорсальных, антеродорсальных, заднедорсальных, вентральных, антеровентральных и задне-вентральных щетинок. Положение, количество, размер и наклон этих щетинок важны в систематике высших мух. Нога сгибается (голень на бедре) в дорсально-вентральной плоскости. Тыльная сторона большеберцовой кости (особенно) и бедренной кости часто определяется двойной линией очень маленьких щетинок. Еще одна важная щетина — преапикальная на большеберцовой кости (наличие или отсутствие важно на уровне семьи).
Живот
[ редактировать ]Морфология брюшка у обоих полов в значительной степени определяется морфоанатомической адаптацией в зависимости от размножения. В целом, 10 уритов (один из сегментов брюшной полости или постбрюшного отдела) редуцируются до меньшего количества уритов из-за структурных изменений первого и последнего уритов. Обычно наблюдается атрофия первого урита и слияние 2° и 3° уротергитов. Тергиты и стерниты хорошо отличаются друг от друга, но часто наблюдается дифференцированное развитие, когда тергиты перекрывают стерниты; крайний случай - когда расширения тергита вентрально сливаются, образуя трубчатую структуру или кольцо. У самок последние мочеточники истончаются и растягиваются, образуя гибкий телескопический яйцеклад. Эта морфологическая адаптация часто сопровождается склеротизацией терминального восьмого уха, благодаря чему яйцеклад может проникать через ткани организма, в которых будут размещаться яйца и личинки. У самцов последние мочевые пути претерпевают сложную трансформацию, образуя устройство, интегрированное с гениталиями, называемое гипопигием. Степень и характер структурных изменений варьируют в зависимости от систематической группы, но обычно сопровождаются развитием долей девятого уротергита в щипцы (эпандрий) и IX уростерн (гипандрий). Иногда наблюдается перекручивание оси брюшка, приводящее к изменению положения эпандрия и гипандрия.
 Брюшко Sarcophaga carnaria
Брюшко Sarcophaga carnaria Брюшко Каллифоры
Брюшко Каллифоры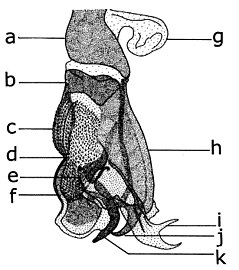 Гениталии самца Afridigalia adrianponti
Гениталии самца Afridigalia adrianponti



Внутренний
[ редактировать ]Пищеварительная система
[ редактировать ]Передняя кишка
[ редактировать ]Сокращение отдела P4 передней кишки модулируется серотонином , который является экзогенным для самой передней кишки и поступает вместо этого из гемолимфы . В частности, серотонин увеличивает скорость сокращений. Liscia et al., 2012 продемонстрировали, что этот серотонин, вероятно, возникает в серотониновых нервных сплетениях грудно -абдоминального синганглия . Если зоб наполнен и серотонин достигает P4, P4 сжимается и выталкивает содержимое в P2, и в зависимости от активации сфинктера в это время P2 выталкивает его в среднюю кишку для переваривания или срыгивает его в пустоту. [5]
Личинка
[ редактировать ]Большинство личинок двукрылых обитают в водной среде, в разлагающихся органических субстратах и в других организмах (грибах, животных, растениях). Поэтому их морфологическая структура имеет существенное упрощение.
Личинка двукрылых безногих (без ног), но иногда, особенно у водных личинок, имеет придатки, похожие на псевдоподии. Голова обычно лишена глаз, имеет жевательный ротовой аппарат, видоизмененные усики с числом сегментов до шести, более или менее развитые или редуцированные до сосочков. Голова может быть: четко отделена от грудной клетки (эвцефальные личинки), неотличима от остального тела (микроцефалическая) или затоплена в грудной клетке (криптоцефальная).
 Молофилус , Типулиды. Червеобразная личинка не имеет ложноножек и рубцов, но лопасти и дыхальца (две) на заднем конце идентифицируют ее как Diptera.
Молофилус , Типулиды. Червеобразная личинка не имеет ложноножек и рубцов, но лопасти и дыхальца (две) на заднем конце идентифицируют ее как Diptera. Личинка Anopheles (эуцефальные личинки)
Личинка Anopheles (эуцефальные личинки) Culex restuans Личинка
Culex restuans Личинка Личинка Muscidae (микроцефалическая)
Личинка Muscidae (микроцефалическая) Stratiomyidae Личинка
Stratiomyidae Личинка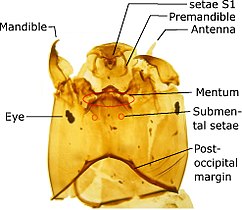 ортокладиины Голова
ортокладиины Голова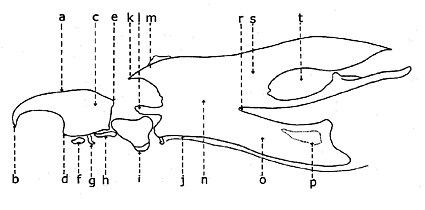 Цефалоглоточный аппарат
Цефалоглоточный аппарат Eristalis Личинка с дыхательным сифоном
Eristalis Личинка с дыхательным сифоном








В зависимости от количества и положения дыхалец трахеи можно выделить следующие типы дыхательных аппаратов.
- апнейстический, с отсутствием рыльцев ( дыхалец )
- метапневстический, только с одной парой рыльцев. Они находятся на животе.
- амфипнейстический, с двумя парами рыльцев. По паре на переднегруди, одна пара на брюшке.
- голопневстический, с двумя парами грудных рыльцев и восемью брюшными парами.
Наиболее частый тип, встречающийся в совокупности Brachycera, является амфипневстическим, тогда как другие типы встречаются преимущественно у водных личинок.Личинки Brachycera Cyclorrhapha имеют червеобразный вид с незначительной дифференциацией частей тела (голова, грудь, брюшко) до такой степени, что их обычно ошибочно называют червями. Эти личинки имеют цилиндро-коническую форму, более широкую в брюшке. Упрощенный ротовой аппарат, представленный двумя челюстями, имеющими форму крючка, и рядом внутренних головных склеритов, образующихся в сложном цефалоглоточном аппарате в отличие от других жевательных ротовых аппаратов, крючки цефалоглоточного аппарата снабжены движениями в вертикальной плоскости.
Особые морфологические приспособления наблюдаются у личинок, приспособленных к жизни в водной среде или в качестве эндопаразитоидов: например, у сапропагических водных личинок Eristalis , имеющих длинный дыхательный сифон, позволяющий им жить в слякотных или вонючих водах, а у Tachinidae имеют дыхательные трубки, ведущие в трахеи хозяина или за пределы тела хозяина.
Красный
[ редактировать ]Куколки двукрылых могут быть выпуклыми, экзаратными и коарктными. У куколок Obtect очертания крыльев и ног видны, но плотно прижаты к остальной части тела, так как все окутано единственной кутикулой; куколки exarate имеют придатки, окруженные собственной кутикулой, и поэтому их можно отделить от остальной части насекомого. Коарктатные куколки развиваются внутри кожи личинки.
Куколки Cyclorrhapha , подотряда Brachycera , имеют коарктатные куколки в куколке (футляре, образующемся в результате затвердевания личиночной кожицы), образовавшемся в результате модификации морфологического и биохимического экзувия последней личиночной стадии. По способу открытия пупария во время вылета имаго различают две большие систематические группы: Aschiza и Schizophora .
Куколки Obtect обычно свободны и незащищены, за исключением куколок Simuliidae , которые защищены боццолетти, построенными из остатков, склеенных между собой шелком.
В последней фазе жизни куколки двукрылых становятся подвижными.
 Обтектная куколка Anopheles
Обтектная куколка Anopheles Пупарий Ceratitis capitata
Пупарий Ceratitis capitata


Ссылки и дальнейшее чтение
[ редактировать ]Эта статья во многом основана на переводе итальянской страницы о Diptera .
- Браун, Б.В., Боркент, А., Камминг, Дж.М., Вуд, Д.М., Вудли, Н.Э. и Зумбадо, М. (редакторы) 2009 Руководство по центральноамериканским двукрылым . Том 1 NRC Research Press, Оттава ISBN 978-0-660-19833-0
- Капинера, Джон Л. (редактор), Энциклопедия энтомологии Springer Verlag, 2008 г. ISBN 978-1-4020-6242-1 .
- Коллесс, Д.Х. и Макэлпайн, Д.К., 1991 г. Diptera (мухи), стр. 717–786. В: Отдел энтомологии. Организация Содружества научных и промышленных исследований, Канберра (спонсор), «Насекомые Австралии». Мельбурнский университет. Пресс, Мельбурн.
- Гриффитс, GCD Филогенетическая классификация Diptera Cyclorrhapha с особым упором на строение постабдомена самцов . Сер. Энт. 8, 340 стр. [Др. В. Джанк, Невада, Гаага] (1972).
- Вилли Хенниг . Личиночные формы двукрылых . 3-я часть. Акад.-Верлаг, Берлин. 185 стр., 3 пл. 1948 год
- Макэлпайн, Дэвид К., 1958 Ключ к австралийским семействам Acalptrate Diptera (Insecta). Отчеты Австралийского музея 24 (12) 183-190 , полный текст и рисунки в формате pdf.
- Макэлпайн, Дж. Ф. 1981. Морфология и терминология. В: Макэлпайн, Дж. П. и др. (ред.): Руководство по неарктическим двукрылым, том. 1 Оттава: Исследовательский отдел, Сельское хозяйство Канады, Монография 27. ISBN 0660107317 формате pdf скачать инструкцию в
- Бернхард Мерц, Жан-Поль Хэнни, 2000 г. Морфология и терминология взрослых двукрылых (кроме терминалий) . В: Ласло Папп, Бела Дарвас. Вклад в руководство по палеарктическим двукрылым . Том 1: Общая и прикладная диптерология. Будапешт, Science Herald, 2000: 22–51. ISBN 963-04-8839-6 .
- КГВ Смит, 1989 г. Знакомство с незрелыми стадиями британских мух. Diptera Larvae, с примечаниями о яйцах, пупариях и куколках. Справочники по идентификации британских насекомых , том 10, часть 14. Скачать руководство в формате pdf (две части, основной указатель текста и рисунков)
- Хунфу, Чжу, 1949 г. Как узнать неполовозрелых насекомых; иллюстрированный ключ для определения отрядов и семейств многих неполовозрелых насекомых с предложениями по их сбору, выращиванию и изучению, автор: Х. Ф. Чу. На фото ключевая серия о природе Дубьюк, Айова, WC Brown Co. Полный текст онлайн здесь.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Капинера, Джон Л. (2008). Энциклопедия энтомологии (2-е изд.). Springer Science & Business Media. п. 1481 . ISBN 9781402062421 .
- ^ Перейти обратно: а б с Мейер, Джон Р. (28 марта 2016 г.). «Двукрылые: настоящие мухи/комары/комары/мошки» . Общая энтомология . Государственный университет Северной Каролины . Проверено 5 февраля 2018 г.
- ^ Десси, Джанкарло (28 мая 2019 г.). «Морфология и анатомия взрослых: усики» . Мухи [Насекомое: двукрылые] . Проверено 27 августа 2020 г.
- ^ «Больше информации о: Тегула» . Diptera.info .
- ^ Стоффолано, Джон Г .; Хазелтон, Аарон Т. (7 января 2013 г.). «Взрослый урожай двукрылых: уникальный и забытый орган». Ежегодный обзор энтомологии . 58 (1). Годовые обзоры : 205–225. doi : 10.1146/annurev-ento-120811-153653 . ISSN 0066-4170 . ПМИД 23317042 .
Внешние ссылки
[ редактировать ]![]() СМИ, связанные с анатомией двукрылых, на Викискладе?
СМИ, связанные с анатомией двукрылых, на Викискладе?
- Анатомия мухи .
- Глоссарий тефритид
- Джанкарло Десси
- Рисунок Жилкование крыла
- Водные двукрылые Монголии
- MDFRC Водные двукрылые Австралии
- «Мир насекомых» Иллюстрированная статья (на французском языке)