Четвероногие
| Четвероногие Временной диапазон: Четвероногие позвоночные (tetrapods sensu lato ) возникли в эйфельском ярусе среднего девона. [2] | |
|---|---|
 | |
| По часовой стрелке сверху слева: Mercurana myristicapaulstris , кустарниковая лягушка; Dermophis mexicanus , безногая амфибия; Equus quagga , равнинная зебра; Sterna maxima , крачка (морская птица); Pseudotrapelus sinaitus , синайская агама; Tachyglossus aculeatus , ехидна с коротким клювом. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Инфрафилум: | челюстноротые |
| Клэйд : | Эугнатостома |
| Клэйд : | Телеостома |
| Суперкласс: | Четвероногие Хачек и Кори, 1896 г. [ Лорин ] [3] [4] |
| Подгруппы | |
| |
Четвероногий ( / ˈ t ɛ t r ə ˌ p ɒ d / ; [5] от древнегреческого τετρα- (тетра-) 'четыре' и ( poús) 'нога') — любое четырехконечное позвоночное животное надкласса Четвероногих ( / πούς t ɛ ˈ t r æ p ə d ə / ) . [6] К четвероногим относятся все современные и вымершие амфибии и амниоты , причем последние, в свою очередь, эволюционировали в две основные клады : зауропсиды ( рептилии , включая динозавров и, следовательно, птиц ) и синапсиды (вымершие пеликозавры , терапсиды и все современные млекопитающие , включая человека ). Некоторые четвероногие, такие как змеи , безногие ящерицы и червячи , в ходе эволюции стали безногими в результате мутаций гена Hox . [7] Тем не менее, эти группы без конечностей по-прежнему квалифицируются как четвероногие по своему происхождению, а у некоторых сохранилась пара рудиментарных шпор , которые являются остатками задних конечностей .
Четвероногие произошли от группы примитивных полуводных животных, известных как тетраподоморфы , которые, в свою очередь, произошли от древних лопастных рыб ( саркоптеригов ) около 390 миллионов лет назад, в период среднего девона . [8] Тетраподоморфы были переходными между лопастными рыбами и настоящими четвероногими с четырьмя конечностями, хотя большинство из них все еще соответствовали строению тела, ожидаемому от других лопастных рыб. Древнейшие окаменелости четвероногих позвоночных (четвероногих в широком смысле слова) — следы из среднего девона , а окаменелости тела стали распространены ближе к концу позднего девона , около 370—360 миллионов лет назад. четвероногих Все эти девонские виды принадлежали к стеблевой группе , а это означает, что они не имели прямого отношения к какой-либо современной группе четвероногих. Широкие анатомические дескрипторы, такие как «четвероногие» и «амфибия», могут приближаться к некоторым членам стволовой группы, но некоторые палеонтологи предпочитают более конкретные термины, такие как Stegocephali . Конечности развились до наземного передвижения , но к началу каменноугольного периода, 360 миллионов лет назад, несколько стволовых четвероногих экспериментировали с полуводным образом жизни, чтобы добывать пищу и кров на суше. Первые коронные — четвероногие (происшедшие от последних общих предков современных четвероногих) появились к визейскому веку раннего карбона . [1]
Конкретные водные предки четвероногих и процесс, посредством которого они колонизировали сушу Земли после выхода из воды, остаются неясными. Переход от строения тела, основанного жаберном на дыхании и хвоста с помощью водном передвижении , к такому, который позволяет животному выживать вне воды и передвигаться по суше, является одним из самых глубоких известных эволюционных изменений. [9] [10] Четвероногие имеют многочисленные анатомические и физиологические особенности, отличающиеся от их предков — водных рыб. К ним относятся отдельные структуры головы и шеи для питания и движений, добавочные скелеты ( , плечевой и тазовый пояс в частности ) для переноски веса и передвижения, более универсальные глаза для зрения, средние уши для слуха и более эффективные сердце и легкие для циркуляции и обмена кислорода. внешняя вода.
Stem-tetrapods and "fish-a-pods" were primarily aquatic. Modern amphibians, which evolved from earlier groups, are generally semiaquatic; the first stages of their lives are as waterborne eggs and fish-like larvae known as tadpoles, and later undergo metamorphosis to grow limbs and become partly terrestrial and partly aquatic. However, most tetrapod species today are amniotes, most of which are terrestrial tetrapods whose branch evolved from earlier tetrapods early in the Late Carboniferous. The key innovation in amniotes over amphibians is the amnion, which enables the eggs to retain their aqueous contents on land, rather than needing to stay in water. (Some amniotes later evolved internal fertilization, although many aquatic species outside the tetrapod tree had evolved such before the tetrapods appeared, e.g. Materpiscis.) Some tetrapods, such as snakes and caecilians, have lost some or all of their limbs through further speciation and evolution; some have only concealed vestigial bones as a remnant of the limbs of their distant ancestors. Others returned to being amphibious or otherwise living partially or fully aquatic lives, the first during the Carboniferous period,[11] others as recently as the Cenozoic.[12][13]
One fundamental subgroup of amniotes, the sauropsids, diverged into the reptiles: lepidosaurs (lizards, snakes, and the tuatara), archosaurs (crocodilians and dinosaurs, of which birds are a subset), turtles, and various other extinct forms. The remaining group of amniotes, the synapsids, include mammals and their extinct relatives. Amniotes include the only tetrapods that further evolved for flight—such as birds from among the dinosaurs, the extinct pterosaurs from earlier archosaurs, and bats from among the mammals.
Definitions
[edit]The precise definition of "tetrapod" is a subject of strong debate among paleontologists who work with the earliest members of the group.[14][15][16][17]
Apomorphy-based definitions
[edit]A majority of paleontologists use the term "tetrapod" to refer to all vertebrates with four limbs and distinct digits (fingers and toes), as well as legless vertebrates with limbed ancestors.[15][16] Limbs and digits are major apomorphies (newly evolved traits) which define tetrapods, though they are far from the only skeletal or biological innovations inherent to the group. The first vertebrates with limbs and digits evolved in the Devonian, including the Late Devonian-age Ichthyostega and Acanthostega, as well as the trackmakers of the Middle Devonian-age Zachelmie trackways.[8]
Defining tetrapods based on one or two apomorphies can present a problem if these apomorphies were acquired by more than one lineage through convergent evolution. To resolve this potential concern, the apomorphy-based definition is often supported by an equivalent cladistic definition. Cladistics is a modern branch of taxonomy which classifies organisms through evolutionary relationships, as reconstructed by phylogenetic analyses. A cladistic definition would define a group based on how closely related its constituents are. Tetrapoda is widely considered a monophyletic clade, a group with all of its component taxa sharing a single common ancestor.[16] In this sense, Tetrapoda can also be defined as the "clade of limbed vertebrates", including all vertebrates descended from the first limbed vertebrates.[17]
Crown group tetrapods
[edit]
* Under the apomorphy-based definition used by many paleontologists, tetrapods originate at the orange star ("First vertebrates with tetrapod limb")
* When restricted to the crown group, tetrapods originate at the "last common ancestor of recent tetrapods"
A portion of tetrapod workers, led by French paleontologist Michel Laurin, prefer to restrict the definition of tetrapod to the crown group.[14][18] A crown group is a subset of a category of animal defined by the most recent common ancestor of living representatives. This cladistic approach defines "tetrapods" as the nearest common ancestor of all living amphibians (the lissamphibians) and all living amniotes (reptiles, birds, and mammals), along with all of the descendants of that ancestor. In effect, "tetrapod" is a name reserved solely for animals which lie among living tetrapods, so-called crown tetrapods. This is a node-based clade, a group with a common ancestry descended from a single "node" (the node being the nearest common ancestor of living species).[16]
Defining tetrapods based on the crown group would exclude many four-limbed vertebrates which would otherwise be defined as tetrapods. Devonian "tetrapods", such as Ichthyostega and Acanthostega, certainly evolved prior to the split between lissamphibians and amniotes, and thus lie outside the crown group. They would instead lie along the stem group, a subset of animals related to, but not within, the crown group. The stem and crown group together are combined into the total group, given the name Tetrapodomorpha, which refers to all animals closer to living tetrapods than to Dipnoi (lungfishes), the next closest group of living animals.[19] Many early tetrapodomorphs are clearly fish in ecology and anatomy, but later tetrapodomorphs are much more similar to tetrapods in many regards, such as the presence of limbs and digits.
Laurin's approach to the definition of tetrapods is rooted in the belief that the term has more relevance for neontologists (zoologists specializing in living animals) than paleontologists (who primarily use the apomorphy-based definition).[17] In 1998, he re-established the defunct historical term Stegocephali to replace the apomorphy-based definition of tetrapod used by many authors.[20] Other paleontologists use the term stem-tetrapod to refer to those tetrapod-like vertebrates that are not members of the crown group, including both early limbed "tetrapods" and tetrapodomorph fishes.[21] The term "fishapod" was popularized after the discovery and 2006 publication of Tiktaalik, an advanced tetrapodomorph fish which was closely related to limbed vertebrates and showed many apparently transitional traits.
The two subclades of crown tetrapods are Batrachomorpha and Reptiliomorpha. Batrachomorphs are all animals sharing a more recent common ancestry with living amphibians than with living amniotes (reptiles, birds, and mammals). Reptiliomorphs are all animals sharing a more recent common ancestry with living amniotes than with living amphibians.[22] Gaffney (1979) provided the name Neotetrapoda to the crown group of tetrapods, though few subsequent authors followed this proposal.[17]
Biodiversity
[edit]Tetrapoda includes three living classes: amphibians, reptiles, and mammals. Overall, the biodiversity of lissamphibians,[23] as well as of tetrapods generally,[24] has grown exponentially over time; the more than 30,000 species living today are descended from a single amphibian group in the Early to Middle Devonian. However, that diversification process was interrupted at least a few times by major biological crises, such as the Permian–Triassic extinction event, which at least affected amniotes.[25] The overall composition of biodiversity was driven primarily by amphibians in the Palaeozoic, dominated by reptiles in the Mesozoic and expanded by the explosive growth of birds and mammals in the Cenozoic. As biodiversity has grown, so has the number of species and the number of niches that tetrapods have occupied. The first tetrapods were aquatic and fed primarily on fish. Today, the Earth supports a great diversity of tetrapods that live in many habitats and subsist on a variety of diets.[24] The following table shows summary estimates for each tetrapod class from the IUCN Red List of Threatened Species, 2014.3, for the number of extant species that have been described in the literature, as well as the number of threatened species.[26]
| Tetrapod group | Image | Class | Estimated number of described species[26] [a] | Number of species evaluated for Red List[26] | Share of described species evaluated for Red List[26] | Threatened species in Red List[26] | Best estimate of percent of threatened species[26] |
|---|---|---|---|---|---|---|---|
| Anamniotes lay eggs in water |  | Amphibians | 8,707 | 8,020 | 92% | 2,876 | 41% |
| Amniotes adapted to lay eggs on land |  | Reptiles | 12,060 | 10,254 | 85% | 1,848 | 21% |
 | Birds | 11,197 | 11,197 | 100% | 1,354 | 12% | |
 | Mammal | 6,631 | 5,980 | 90% | 1,339 | 26% | |
| Overall | 38,595 | 35,451 | 92% | 7,417 | |||
- ^ The estimates for amphibians, reptiles, birds and mammals were respectively taken from Amphibian Species of the World: An Online Reference (version 6.2, 1 December 2023), the Reptile Database (accessed: 0 December 2023), Handbook of the Birds of the World and BirdLife International digital checklist of the birds of the world (version 8; accessed: 11 December 2023) and the Mammal Diversity Database] (verison 1.11, released 15 April 2023; accessed 01 December 2023).[26]
Classification
[edit]
The classification of tetrapods has a long history. Traditionally, tetrapods are divided into four classes based on gross anatomical and physiological traits.[27] Snakes and other legless reptiles are considered tetrapods because they are sufficiently like other reptiles that have a full complement of limbs. Similar considerations apply to caecilians and aquatic mammals. Newer taxonomy is frequently based on cladistics instead, giving a variable number of major "branches" (clades) of the tetrapod family tree.
As is the case throughout evolutionary biology today, there is debate over how to properly classify the groups within Tetrapoda. Traditional biological classification sometimes fails to recognize evolutionary transitions between older groups and descendant groups with markedly different characteristics. For example, the birds, which evolved from the dinosaurs, are defined as a separate group from them, because they represent a distinct new type of physical form and functionality. In phylogenetic nomenclature, in contrast, the newer group is always included in the old. For this school of taxonomy, dinosaurs and birds are not groups in contrast to each other, but rather birds are a sub-type of dinosaurs.
History of classification
[edit]The tetrapods, including all large- and medium-sized land animals, have been among the best understood animals since earliest times. By Aristotle's time, the basic division between mammals, birds and egg-laying tetrapods (the "herptiles") was well known, and the inclusion of the legless snakes into this group was likewise recognized.[28] With the birth of modern biological classification in the 18th century, Linnaeus used the same division, with the tetrapods occupying the first three of his six classes of animals.[29] While reptiles and amphibians can be quite similar externally, the French zoologist Pierre André Latreille recognized the large physiological differences at the beginning of the 19th century and split the herptiles into two classes, giving the four familiar classes of tetrapods: amphibians, reptiles, birds and mammals.[30]
Modern classification
[edit]With the basic classification of tetrapods settled, a half a century followed where the classification of living and fossil groups was predominantly done by experts working within classes. In the early 1930s, American vertebrate palaeontologist Alfred Romer (1894–1973) produced an overview, drawing together taxonomic work from the various subfields to create an orderly taxonomy in his Vertebrate Paleontology.[31] This classical scheme with minor variations is still used in works where systematic overview is essential, e.g. Benton (1998) and Knobill and Neill (2006).[32][33] While mostly seen in general works, it is also still used in some specialist works like Fortuny et al. (2011).[34] The taxonomy down to subclass level shown here is from Hildebrand and Goslow (2001):[35]
- Superclass Tetrapoda – four-limbed vertebrates
- Class Amphibia – amphibians
- Subclass Ichthyostegalia – early fish-like amphibians (paraphyletic group outside leading to the crown-clade Neotetrapoda)
- Subclass Anthracosauria – reptile-like amphibians (often thought to be the ancestors of the amniotes)
- Subclass Temnospondyli – large-headed Paleozoic and Mesozoic amphibians
- Subclass Lissamphibia – modern amphibians
- Class Reptilia – reptiles
- Subclass Diapsida – diapsids, including crocodiles, dinosaurs, birds, lizards, snakes and turtles
- Subclass Euryapsida – euryapsids
- Subclass Synapsida – synapsids, including mammal-like reptiles-now a separate group (often thought to be the ancestors of mammals)
- Subclass Anapsida – anapsids
- Class Mammalia – mammals
- Subclass Prototheria – egg-laying mammals, including monotremes
- Subclass Allotheria – multituberculates
- Subclass Theria – live-bearing mammals, including marsupials and placentals
- Class Amphibia – amphibians
This classification is the one most commonly encountered in school textbooks and popular works. While orderly and easy to use, it has come under critique from cladistics. The earliest tetrapods are grouped under class Amphibia, although several of the groups are more closely related to amniotes than to modern day amphibians. Traditionally, birds are not considered a type of reptile, but crocodiles are more closely related to birds than they are to other reptiles, such as lizards. Birds themselves are thought to be descendants of theropod dinosaurs. Basal non-mammalian synapsids ("mammal-like reptiles") traditionally also sort under class Reptilia as a separate subclass,[27] but they are more closely related to mammals than to living reptiles. Considerations like these have led some authors to argue for a new classification based purely on phylogeny, disregarding the anatomy and physiology.
Evolution
[edit]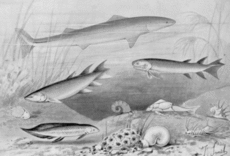

Ancestry
[edit]Tetrapods evolved from early bony fishes (Osteichthyes), specifically from the tetrapodomorph branch of lobe-finned fishes (Sarcopterygii), living in the early to middle Devonian period.



The first tetrapods probably evolved in the Emsian stage of the Early Devonian from Tetrapodomorph fish living in shallow water environments.[36][37]The very earliest tetrapods would have been animals similar to Acanthostega, with legs and lungs as well as gills, but still primarily aquatic and unsuited to life on land.
The earliest tetrapods inhabited saltwater, brackish-water, and freshwater environments, as well as environments of highly variable salinity. These traits were shared with many early lobed-finned fishes. As early tetrapods are found on two Devonian continents, Laurussia (Euramerica) and Gondwana, as well as the island of North China, it is widely supposed that early tetrapods were capable of swimming across the shallow (and relatively narrow) continental-shelf seas that separated these landmasses.[38][39][40]
Since the early 20th century, several families of tetrapodomorph fishes have been proposed as the nearest relatives of tetrapods, among them the rhizodonts (notably Sauripterus),[41][42] the osteolepidids, the tristichopterids (notably Eusthenopteron), and more recently the elpistostegalians (also known as Panderichthyida) notably the genus Tiktaalik.[43]
A notable feature of Tiktaalik is the absence of bones covering the gills. These bones would otherwise connect the shoulder girdle with skull, making the shoulder girdle part of the skull. With the loss of the gill-covering bones, the shoulder girdle is separated from the skull, connected to the torso by muscle and other soft-tissue connections. The result is the appearance of the neck. This feature appears only in tetrapods and Tiktaalik, not other tetrapodomorph fishes. Tiktaalik also had a pattern of bones in the skull roof (upper half of the skull) that is similar to the end-Devonian tetrapod Ichthyostega. The two also shared a semi-rigid ribcage of overlapping ribs, which may have substituted for a rigid spine. In conjunction with robust forelimbs and shoulder girdle, both Tiktaalik and Ichthyostega may have had the ability to locomote on land in the manner of a seal, with the forward portion of the torso elevated, the hind part dragging behind. Finally, Tiktaalik fin bones are somewhat similar to the limb bones of tetrapods.[44][45]
However, there are issues with positing Tiktaalik as a tetrapod ancestor. For example, it had a long spine with far more vertebrae than any known tetrapod or other tetrapodomorph fish. Also the oldest tetrapod trace fossils (tracks and trackways) predate it by a considerable margin. Several hypotheses have been proposed to explain this date discrepancy: 1) The nearest common ancestor of tetrapods and Tiktaalik dates to the Early Devonian. By this hypothesis, the lineage is the closest to tetrapods, but Tiktaalik itself was a late-surviving relic.[46] 2) Tiktaalik represents a case of parallel evolution. 3) Tetrapods evolved more than once.[47][48]
| Euteleostomi / Osteichthyes |
| ||||||||||||||||||||||||
| (bony vertebrates) |
History
[edit]Palaeozoic
[edit]Devonian stem-tetrapods
[edit]The oldest evidence for the existence of tetrapods comes from trace fossils: tracks (footprints) and trackways found in Zachełmie, Poland, dated to the Eifelian stage of the Middle Devonian, 390 million years ago,[8] although these traces have also been interpreted as the ichnogenus Piscichnus (fish nests/feeding traces).[49] The adult tetrapods had an estimated length of 2.5 m (8 feet), and lived in a lagoon with an average depth of 1–2 m, although it is not known at what depth the underwater tracks were made. The lagoon was inhabited by a variety of marine organisms and was apparently salt water. The average water temperature was 30 degrees C (86 F).[2][50] The second oldest evidence for tetrapods, also tracks and trackways, date from ca. 385 Mya (Valentia Island, Ireland).[51][52]
The oldest partial fossils of tetrapods date from the Frasnian beginning ≈380 mya. These include Elginerpeton and Obruchevichthys.[53] Some paleontologists dispute their status as true (digit-bearing) tetrapods.[54]
All known forms of Frasnian tetrapods became extinct in the Late Devonian extinction, also known as the end-Frasnian extinction.[55] This marked the beginning of a gap in the tetrapod fossil record known as the Famennian gap, occupying roughly the first half of the Famennian stage.[55]
The oldest near-complete tetrapod fossils, Acanthostega and Ichthyostega, date from the second half of the Fammennian.[56][57] Although both were essentially four-footed fish, Ichthyostega is the earliest known tetrapod that may have had the ability to pull itself onto land and drag itself forward with its forelimbs. There is no evidence that it did so, only that it may have been anatomically capable of doing so.[58][59]
The publication in 2018 of Tutusius umlambo and Umzantsia amazana from high latitude Gondwana setting indicate that the tetrapods enjoyed a global distribution by the end of the Devonian and even extend into the high latitudes.[60]

The end-Fammenian marked another extinction, known as the end-Fammenian extinction or the Hangenberg event, which is followed by another gap in the tetrapod fossil record, Romer's gap, also known as the Tournaisian gap.[61] This gap, which was initially 30 million years, but has been gradually reduced over time, currently occupies much of the 13.9-million year Tournaisian, the first stage of the Carboniferous period.[62]Tetrapod-like vertebrates first appeared in the Early Devonian period, and species with limbs and digits were around by the Late Devonian.[63] These early "stem-tetrapods" included animals such as Ichthyostega,[2] with legs and lungs as well as gills, but still primarily aquatic and poorly adapted for life on land. The Devonian stem-tetrapods went through two major population bottlenecks during the Late Devonian extinctions, also known as the end-Frasnian and end-Fammenian extinctions. These extinction events led to the disappearance of stem-tetrapods with fish-like features.[64] When stem-tetrapods reappear in the fossil record in early Carboniferous deposits, some 10 million years later, the adult forms of some are somewhat adapted to a terrestrial existence.[62][65] Why they went to land in the first place is still debated.
Carboniferous
[edit]
During the early Carboniferous, the number of digits on hands and feet of stem-tetrapods became standardized at no more than five, as lineages with more digits died out (exceptions within crown-group tetrapods arose among some secondarily aquatic members). By mid-Carboniferous times, the stem-tetrapods had radiated into two branches of true ("crown group") tetrapods, one ancestral to modern amphibians and the other ancestral to amniotes. Modern amphibians are most likely derived from the temnospondyls, a particularly diverse and long-lasting group of tetrapods. A less popular proposal draws comparisons to the "lepospondyls", an eclectic mixture of various small tetrapods, including burrowing, limbless, and other bizarrely-shaped forms. The reptiliomorphs (sometimes known as "anthracosaurs") were the relatives and ancestors of the amniotes (reptiles, mammals, and kin). The first amniotes are known from the early part of the Late Carboniferous. All basal amniotes had a small body size, like many of their contemporaries, though some Carboniferous tetrapods evolved into large crocodile-like predators, informally known as "labyrinthodonts".[66][67] Amphibians must return to water to lay eggs; in contrast, amniote eggs have a membrane ensuring gas exchange out of water and can therefore be laid on land.
Amphibians and amniotes were affected by the Carboniferous rainforest collapse (CRC), an extinction event that occurred around 307 million years ago. The sudden collapse of a vital ecosystem shifted the diversity and abundance of major groups. Amniotes and temnospondyls in particular were more suited to the new conditions. They invaded new ecological niches and began diversifying their diets to include plants and other tetrapods, previously having been limited to insects and fish.[68]
Permian
[edit]
In the Permian period, amniotes became particularly well-established, and two important clades filled in most terrestrial niches: the sauropsids and the synapsids. The latter were the most important and successful Permian land animals, establishing complex terrestrial ecosystems of predators and prey while acquiring various adaptations retained by their modern descendants, the mammals. Sauropsid diversity was more subdued during the Permian, but they did begin to fracture into several lineages ancestral to modern reptiles. Amniotes were not the only tetrapods to experiment with prolonged life on land. Some temnospondyls, seymouriamorphs, and diadectomorphs also successfully filled terrestrial niches in the earlier part of the Permian. Non-amniote tetrapods declined in the later part of the Permian.
The end of the Permian saw a major turnover in fauna during the Permian–Triassic extinction event. There was a protracted loss of species, due to multiple extinction pulses.[69] Many of the once large and diverse groups died out or were greatly reduced.
Mesozoic
[edit]The diapsid reptiles (a subgroup of the sauropsids) strongly diversified during the Triassic, giving rise to the turtles, pseudosuchians (crocodilian ancestors), dinosaurs, pterosaurs, and lepidosaurs, along with many other reptile groups on land and sea. Some of the new Triassic reptiles would not survive into the Jurassic, but others would flourish during the Jurassic. Lizards, turtles, dinosaurs, pterosaurs, crocodylomorphs, and plesiosaurs were particular beneficiaries of the Triassic-Jurassic transition. Birds, a particular subset of theropod dinosaurs capable of flight via feathered wings, evolved in the Late Jurassic. In the Cretaceous, snakes developed from lizards, rhynchocephalians (tuataras and kin) declined, and modern birds and crocodilians started to establish themselves.
Among the characteristic Paleozoic non-amniote tetrapods, few survived into the Mesozoic. Temnospondyls briefly recovered in the Triassic, spawning the large aquatic stereospondyls and the small terrestrial lissamphibians (the earliest frogs, salamanders, and caecilians). However, stereospondyl diversity would crash at the end of the Triassic. By the Late Cretaceous, the only surviving amphibians were lissamphibians. Many groups of synapsids, such as anomodonts and therocephalians, that once comprised the dominant terrestrial fauna of the Permian, also became extinct during the Triassic. During the Jurassic, one synapsid group (Cynodontia) gave rise to the modern mammals, which survived through the rest of the Mesozoic to later diversify during the Cenozoic. The Cretaceous-Paleogene extinction event at the end of the Mesozoic killed off many organisms, including all the non-avian dinosaurs and nearly all marine reptiles. Birds survived and diversified during the Cenozoic, similar to mammals.
Cenozoic
[edit]Following the great extinction event at the end of the Mesozoic, representatives of seven major groups of tetrapods persisted into the Cenozoic era. One of them, a group of semiaquatic reptiles known as the Choristodera, became extinct 11 million years ago for unclear reasons.[70] The seven Cenozoic tetrapods groups are:
- Lissamphibia: frogs, salamanders, and caecilians
- Mammalia: monotremes, marsupials, placentals,and †multituberculates
- Lepidosauria: tuataras and lizards (including amphisbaenians and snakes)
- Testudines: turtles
- Crocodilia: crocodiles, alligators, caimans and gharials
- Aves: birds
- †Choristodera (extinct)
Phylogeny
[edit]Stem group
[edit]Stem tetrapods are all animals more closely related to tetrapods than to lungfish, but excluding the tetrapod crown group. The cladogram below illustrates the relationships of stem-tetrapods. All these lineages are extinct except for Dipnomorpha and Tetrapoda; from Swartz, 2012:[71]
| Rhipidistia |
| ||||||
Crown group
[edit]Crown tetrapods are defined as the nearest common ancestor of all living tetrapods (amphibians, reptiles, birds, and mammals) along with all of the descendants of that ancestor.
Включение некоторых вымерших групп в крону четвероногих зависит от родственных связей современных амфибий, или лисамфибий . В настоящее время существуют три основные гипотезы происхождения лисамфибий. Согласно гипотезе темноспондилов (TH), лиссамфибии наиболее тесно связаны с диссорофоидными темноспондилами , что делает темноспондилов четвероногими. В гипотезе лепоспондилов (LH) лиссамфибии являются сестринским таксоном лизорофовых лепоспондилов , что делает лепоспондилов четвероногими, а темноспондилов стеблевыми четвероногими. Согласно полифилетической гипотезе (PH), лягушки и саламандры произошли от диссорофоидных темноспондилов, тогда как червяги произошли от лепоспондилов микрозавров, в результате чего как лепоспондилы, так и темноспондилы стали настоящими четвероногими. [72] [73]
Происхождение современных земноводных
[ редактировать ]Темноспондильная гипотеза (ТН)
[ редактировать ]Эта гипотеза существует в нескольких вариантах, в большинстве из которых лиссамфибии произошли от диссорофоидных темноспондилов, обычно с акцентом на амфибамид и бранхиозавридов. [74]
Гипотеза темноспондилов в настоящее время является предпочитаемой или точкой зрения большинства, поддерживаемой Рутой и др . (2003a,b), Рутой и Коутсом (2007), Коутсом и др. (2008), Сигурдсеном и Грином (2011) и Шохом (2013, 2014). . [73] [75]
Кладограмма изменена по данным Коутса, Руты и Фридмана (2008). [76]
Кронная группа четвероногих |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Лепоспондильная гипотеза (ЛГ)
[ редактировать ]Кладограмма изменена на основе Лорина, «Как позвоночные покинули воду» (2010). [77]
| Стегоцефалия | † стеблевые четвероногие общая группа Lissamphibia общая группа Амниота | |
| («Четвероногие») |

Гипотеза полифилии (ПГ)
[ редактировать ]По этой гипотезе батрахии (лягушки и саламандра) произошли от диссорофоидных темноспондилов, а червячие - от лепоспондилов микрозавров. Есть два варианта: один разработан Кэрроллом , [78] другой Андерсона. [79]
Кладограмма модифицирована по Шоху, Фробишу (2009). [80]
| Четвероногие |
| ||||||||||||||||||||||||

Анатомия и физиология
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Июль 2015 г. ) |
Рыба-предок четвероногих, тетраподоморф, обладала чертами, схожими с чертами, унаследованными ранними четвероногими, включая внутренние ноздри и большой мясистый плавник, построенный на костях, которые могли дать начало конечностям четвероногих. Чтобы размножиться в наземной среде , животным пришлось преодолеть определенные трудности. Их тела нуждались в дополнительной поддержке, поскольку плавучесть больше не имела значения. Удержание воды теперь стало важным, поскольку она больше не была живой матрицей и могла легко потеряться в окружающей среде. Наконец, животным нужны были новые сенсорные системы ввода, чтобы иметь возможность разумно функционировать на суше.
Череп
[ редактировать ]У ранних четвероногих мозг занимал только половину черепа. Остальное было заполнено жировой тканью или жидкостью, которая давала мозгу пространство для роста по мере его адаптации к жизни на суше. [81] Структуры неба и челюстей тетраморфов были аналогичны таковым у ранних четвероногих, и их зубной ряд также был схожим: зубы в форме лабиринта располагались на небе в виде ямок и зубов. Основное различие между ранними тетраподоморфными рыбами и ранними четвероногими заключалось в относительном развитии передней и задней частей черепа ; морда гораздо менее развита, чем у большинства ранних четвероногих, а заглазничный череп исключительно длиннее, чем у амфибий. Примечательной характеристикой, отличающей череп четвероногих от черепа рыбы, является относительная длина передней и задней частей. У рыбы была длинная задняя часть, а передняя короткая; таким образом, орбитальные пустоты располагались ближе к переднему концу. У четвероногих передняя часть черепа удлинилась, в результате чего орбиты оказались дальше назад по черепу.
Шея
[ редактировать ]У тетраподоморфных рыб, таких как Eusthenopteron , часть тела, которая позже стала шеей, была покрыта множеством покрывающих жабры костей, известных как оперкулярный ряд . Эти кости функционировали как часть насосного механизма, прогоняющего воду через рот и мимо жабр. Когда рот открывался для приема воды, жаберные створки закрывались (включая жаберные кости), обеспечивая тем самым попадание воды только через рот. Когда рот закрывался, жаберные створки открывались, и вода выталкивалась через жабры.
У Acanthostega , базального четвероногого, жаберные кости исчезли, хотя подлежащие жаберные дуги все еще присутствуют. Помимо глазного ряда, у Acanthostega также утрачены кости, покрывающие горло (горловой ряд). Объединенную оперкулярную серию и гулярную серию иногда называют оперкуло-гулярной или оперкулогулярной серией. Другие кости области шеи, утраченные у Acanthostega (а позже и у четвероногих), включают внелопаточную серию и супраклеитральную серию. Оба набора костей соединяют плечевой пояс с черепом. С утратой этих костей четвероногие обзавелись шеей, позволяющей голове вращаться несколько независимо от туловища. Это, в свою очередь, потребовало более прочных связей мягких тканей между головой и туловищем, включая мышцы и связки, соединяющие череп с позвоночником и плечевым поясом. Кости и группы костей также были консолидированы и укреплены. [82]
У четвероногих каменноугольного периода шейный сустав (затылок) служил точкой поворота позвоночника относительно задней части черепа. У тетраподоморфных рыб, таких как Eusthenopteron , такого шейного сустава не существовало. Вместо этого хорда (стержень из протохряща) входила в отверстие в задней части черепной коробки и продолжалась до ее середины. Acanthostega имела то же устройство, что и Eusthenopteron , и, следовательно, не имела шейного сустава. Шейный сустав развился независимо у разных линий ранних четвероногих. [83]
Кажется, что все четвероногие держат шею в максимально возможном вертикальном положении, когда находятся в нормальной, настороженной позе. [84]
Зубной ряд
[ редактировать ]
У четвероногих была структура зубов, известная как «плидентин», характеризующаяся складчатостью эмали, как видно на поперечном сечении. Более крайняя версия, обнаруженная у ранних четвероногих, известна как «лабиринтодонт» или «лабиринтодонт плидентин». Этот тип строения зубов независимо развился у нескольких видов костных рыб, как лучепёрых, так и лопастных, у некоторых современных ящериц, а также у ряда тетраподоморфных рыб. Складывание, по-видимому, развивается, когда в маленькой челюсти вырастает клык или большой зуб, и прорезывается, когда он еще слаб и незрел. Складывание придает дополнительную прочность молодому зубу, но дает мало преимуществ, когда зуб зрелый. Такие зубы связаны с питанием мягкой добычи у молоди. [85] [86]
Осевой скелет
[ редактировать ]При переходе из воды на сушу позвоночник должен был противостоять изгибу, вызванному весом тела, и обеспечивать подвижность там, где это необходимо. Раньше он мог гнуться по всей длине. Точно так же парные придатки раньше не были связаны с позвоночником, но медленно укрепляющиеся конечности теперь передавали свою поддержку оси тела.
Пояса
[ редактировать ]Плечевой пояс был отсоединен от черепа, что привело к улучшению наземного передвижения. ранних саркоптеригов Клейтрум сохранился в виде ключицы , а межключица была хорошо развита и лежала на нижней стороне груди. У примитивных форм две ключицы и межключичная мышца могли вырасти вентрально таким образом, что образовали широкую грудную пластину. Верхняя часть пояса имела плоскую лопаточную лопатку (плечевую кость) , ниже которой располагалась суставная впадина, служившая поверхностью сочленения плечевой кости, а вентрально располагалась большая плоская клювовидная пластинка, завернутая к средней линии.
Тазовый . пояс также был намного больше, чем простая пластинка рыб, и вмещал больше мышц Он простирался далеко дорсально и соединялся с позвоночником одним или несколькими специализированными крестцовыми ребрами . Задние ноги были в некоторой степени специализированы: они не только поддерживали вес, но и обеспечивали движение. Дорсальным продолжением таза была подвздошная кость , а широкая вентральная пластинка состояла из лобковой кости спереди и седалищной кости сзади. Три кости встречаются в одной точке в центре тазового треугольника, называемого вертлужной впадиной , обеспечивая поверхность сочленения бедренной кости.
Конечности
[ редактировать ]Мясистые лопастные плавники, опирающиеся на кости, по-видимому, были наследственной чертой всех костистых рыб ( Osteichthyes ). У предков лучепёрых рыб ( Actinopterygii ) плавники развивались в другом направлении. Предки четвероногих четвероногих получили дальнейшее развитие лопастных плавников. Парные плавники имели кости, явно , локтевой гомологичные плечевой и лучевой кости в передних плавниках, а также бедренной кости , голени и малоберцовой кости в брюшных плавниках. [87]
Парные плавники ранних саркоптеригов были меньше конечностей четвероногих, но строение скелета было очень сходным в том смысле, что ранние саркоптериги имели одну проксимальную кость (аналог плечевой или бедренной кости ), две кости в следующем сегменте (предплечье или голень) ), а также неравномерное деление плавника, примерно сравнимое со строением запястья / предплюсны и фаланг руки .
Передвижение
[ редактировать ]В типичной позе ранних четвероногих плечо и нога вытянулись почти горизонтально от его тела, а предплечье и голень вытянулись вниз от верхнего сегмента под почти прямым углом . Вес тела не был сосредоточен на конечностях, а был перенесен на 90 градусов наружу и вниз через нижние конечности, которые касались земли. животного Большая часть силы была использована для того, чтобы просто поднять свое тело с земли для ходьбы, что, вероятно, было медленным и трудным. В такой позе он мог делать лишь короткие широкие шаги. Это было подтверждено окаменевшими следами, найденными в каменноугольных породах .
Кормление
[ редактировать ]У ранних четвероногих была широкая зияющая челюсть со слабыми мышцами, позволяющими ее открывать и закрывать. В челюсти располагались небные, сошковые (верхние) и венечные (нижние) клыки средних размеров, а также ряды более мелких зубов. Это отличалось от более крупных клыков и маленьких краевых зубов более ранних рыб-тетраподоморфов, таких как Eusthenopteron . Хотя это указывает на изменение привычек питания, точная природа этих изменений неизвестна. Некоторые учёные предлагают перейти на донное питание или кормление на мелководье (Альберг и Милнер, 1994). Другие предлагают способ питания, сравнимый с режимом питания японской гигантской саламандры, которая использует как всасывание, так и прямое кусание, чтобы поедать мелких ракообразных и рыбу. Изучение этих челюстей показывает, что они использовались для питания под водой, а не на суше. [88]
У более поздних наземных четвероногих появились два способа смыкания челюстей: статический и кинетический инерционный (также известный как щелканье). В статической системе челюстные мышцы расположены таким образом, что челюсти испытывают максимальную силу в сомкнутом или почти сомкнутом состоянии. В кинетической инерционной системе максимальная сила применяется, когда челюсти широко открыты, в результате чего челюсти закрываются с большой скоростью и импульсом. Хотя кинетическая инерционная система иногда встречается у рыб, она требует специальных приспособлений (например, очень узких челюстей), чтобы справляться с высокой вязкостью и плотностью воды, которая в противном случае препятствовала бы быстрому смыканию челюстей.
четвероногих Язык построен из мышц, которые когда-то управляли жаберными отверстиями. Язык прикреплен к подъязычной кости , которая когда-то была нижней половиной пары жаберных перемычек (второй пары после тех, которые превратились в челюсти). [89] [90] [91] Язык не развивался до тех пор, пока жабры не начали исчезать. У Acanthostega все еще были жабры, так что это было более позднее развитие. У животных, питающихся водным путем, пища поддерживается водой и может буквально всплывать (или засасываться) в рот. На суше язык становится важным.
Дыхание
[ редактировать ]На эволюцию дыхания ранних четвероногих повлияло событие, известное как «угольный разрыв», период продолжительностью более 20 миллионов лет, произошедший в среднем и позднем девоне, когда уровень кислорода в атмосфере был слишком низким для поддержания лесных пожаров. [92] В это время рыбы, обитающие в бескислородных водах (с очень низким содержанием кислорода), находились под эволюционным давлением, требующим развития способности дышать воздухом. [93] [94] [95]
Ранние четвероногие, вероятно, использовали четыре метода дыхания : легкие , жабры , кожное дыхание (кожное дыхание) и дыхание через слизистую оболочку пищеварительного тракта, особенно через рот.
Жабры
[ редактировать ]Ранние четвероногие Acanthostega имели по крайней мере три, а возможно и четыре пары жаберных перемычек, каждая из которых содержала глубокие бороздки в том месте, где можно было бы ожидать найти афферентную жаберную артерию. Это убедительно свидетельствует о наличии функциональных жабр. [96] Некоторые водные темноспондилы сохранили внутренние жабры, по крайней мере, до ранней юры. [97] Признаки наличия прозрачных рыбоподобных внутренних жабр присутствуют у архегозавра . [98]
Легкие
[ редактировать ]Легкие возникли как дополнительная пара мешков в горле, позади жаберных мешков. [99] Вероятно, они присутствовали у последнего общего предка костистых рыб. У некоторых рыб они превратились в плавательные пузыри для поддержания плавучести . [100] [101] Легкие и плавательный пузырь гомологичны (происходят от общей предковой формы), как и легочная артерия (которая доставляет лишенную кислорода кровь от сердца к легким) и артерии, снабжающие плавательный пузырь. [102] Воздух вводился в легкие посредством процесса, известного как буккальная накачка . [103] [104]
У древнейших четвероногих выдох, вероятно, осуществлялся с помощью мышц туловища (торакоабдоминальной области). Вдыхание ребрами было либо примитивным для амниот, либо развилось независимо, по крайней мере, у двух разных линий амниот. У земноводных он не встречается. [105] [106] Мускулистая диафрагма уникальна для млекопитающих. [107]
Стремление отдачи
[ редактировать ]Хотя широко распространено мнение, что четвероногие вдыхали посредством буккального накачивания (накачивания ртом), согласно альтернативной гипотезе, аспирация (вдыхание) происходила посредством пассивной отдачи экзоскелета , аналогично современной примитивной лучепёрой рыбе Polypterus . Эта рыба вдыхает через дыхало (дыхало) — анатомическую особенность, присущую ранним четвероногим. Выдох осуществляется за счет мышц туловища. Во время выдоха костные чешуйки в верхней части грудной клетки вдавливаются. Когда мышцы расслаблены, костные чешуйки возвращаются в исходное положение, создавая значительное отрицательное давление внутри туловища, что приводит к очень быстрому всасыванию воздуха через дыхалец. [108] [109] [110]
Кожное дыхание
[ редактировать ]Кожное дыхание, известное как кожное дыхание , часто встречается у рыб и земноводных и происходит как в воде, так и вне воды. У некоторых животных водонепроницаемые барьеры препятствуют газообмену через кожу. Например, кератин кожи человека, чешуя рептилий, чешуя современных белковых рыб затрудняют газообмен. Однако у ранних четвероногих чешуя состояла из сильно васкуляризированной кости, покрытой кожей. По этой причине считается, что ранние четвероногие могли в значительной степени осуществлять кожное дыхание. [111]
Метаболизм углекислого газа
[ редактировать ]Хотя дышащие воздухом рыбы могут поглощать кислород через легкие, легкие, как правило, неэффективны для выделения углекислого газа. У четвероногих способность легких выделять CO 2 возникла постепенно и не была полностью достигнута до эволюции амниот. То же ограничение применимо и к кишечному дыханию воздухом (КИШ), т. е. дыханию слизистой оболочкой пищеварительного тракта. [112] Кожа четвероногих была эффективна как для поглощения кислорода, так и для выделения CO 2 , но лишь до определенного момента. По этой причине ранние четвероногие могли страдать хронической гиперкапнией (высоким уровнем CO 2 в крови ). Это не редкость для рыб, обитающих в водах с высоким содержанием CO 2 . [113] Согласно одной из гипотез, «скульптурные» или «украшенные» кожные кости крыши черепа, обнаруженные у ранних четвероногих, могли быть связаны с механизмом облегчения респираторного ацидоза (кислоты крови, вызванной избытком CO 2 ) посредством компенсаторного метаболического алкалоза . [114]
Тираж
[ редактировать ]Ранние четвероногие, вероятно, имели трехкамерное сердце , как и современные земноводные, а также лепидозавровые и хелониевые рептилии, у которых насыщенная кислородом кровь из легких и дезоксигенированная кровь из дышащих тканей поступает через отдельные предсердия и направляется через спиральный клапан в сердце. соответствующий сосуд — аорта для насыщенной кислородом крови и легочная вена для дезоксигенированной крови. Спиральный клапан необходим для сведения к минимуму смешивания двух типов крови, что позволяет животному иметь более высокий уровень метаболизма и быть более активным, чем в противном случае. [115]
Чувства
[ редактировать ]обоняние
[ редактировать ]Разница в плотности воздуха и воды приводит к тому, что запахи (определенные химические соединения, обнаруживаемые хеморецепторами ) ведут себя по-разному. Животному, впервые вышедшему на сушу, было бы трудно обнаружить такие химические сигналы, если бы его сенсорный аппарат развился в контексте обнаружения в воде. Вомероназальный орган также впервые появился в носовой полости для обнаружения феромонов из биологических субстратов на суше, хотя впоследствии он был утерян или превратился в рудиментарный у некоторых линий, таких как архозавры и катарины , но расширился у других, таких как лепидозавры . [116]
Система боковой линии
[ редактировать ]У рыб есть система боковой линии , которая улавливает колебания давления в воде. Такое давление невозможно обнаружить в воздухе, но на черепе ранних четвероногих были обнаружены бороздки для органов чувств боковой линии, что позволяет предположить либо водную, либо преимущественно водную среду обитания . она исчезла Современные земноводные, ведущие полуводный образ жизни, демонстрируют эту особенность, тогда как у высших позвоночных .
Зрение
[ редактировать ]Изменения в глазах произошли потому, что поведение света на поверхности глаза в воздушной и водной среде различается из-за разницы в показателе преломления , поэтому фокусное расстояние линзы изменилось , чтобы функционировать на воздухе. Глаз теперь находился в относительно сухой среде , а не в воде, поэтому веки развились, а слезные протоки развились, чтобы производить жидкость для увлажнения глазного яблока.
Ранние четвероногие унаследовали набор из пяти опсинов палочек и колбочек позвоночных , известных как опсины . [117] [118] [119]
У первых позвоночных присутствовало четыре конусных опсина, унаследованных от предков беспозвоночных:
- LWS / MWS (чувствительность к длинным и средним волнам) — зеленый, желтый или красный
- SWS1 (коротковолновая чувствительность) - ультрафиолет или фиолетовый - теряется у однопроходных (утконос, ехидна)
- SWS2 (чувствительный к коротким волнам) — фиолетовый или синий — отсутствует у териан (плацентарных млекопитающих и сумчатых).
- RH2 (родопсин - как опсин колбочек) - зеленый - теряется отдельно у земноводных и млекопитающих, сохраняется у рептилий и птиц.
У первых челюстных позвоночных присутствовал единственный палочковый опсин, родопсин, унаследованный от предка бесчелюстных позвоночных:
- RH1 (родопсин) — сине-зеленый — используется для ночного видения и цветокоррекции в условиях низкой освещенности.
Баланс
[ редактировать ]Четвероногие сохранили балансирующую функцию внутреннего уха от предков рыб.
Слух
[ редактировать ]воздуха Вибрации не могли вызвать пульсации в черепе, как в настоящем органе слуха . Дыхальца из соединительной ткани сохранились в виде слуховой вырезки , в конечном итоге закрытой барабанной перепонкой , тонкой плотной мембраной , также называемой барабанной перепонкой (однако эта и слуховая вырезка были утрачены у предковых амниот , и позже барабанные перепонки были получены независимо).
Гиомандибула рыбы мигрировала вверх из положения, поддерживающего челюсть, и уменьшалась в размерах , образуя колумеллу . Расположенная между барабанной перепонкой и черепной коробкой в полости, заполненной воздухом, колумелла теперь была способна передавать вибрации от внешней части головы внутрь. Таким образом, колумелла стала важным элементом системы согласования импеданса , связывая воздушные звуковые волны с рецепторной системой внутреннего уха. Эта система развивалась независимо в пределах нескольких различных линий земноводных .
Ухо согласования импеданса должно было соответствовать определенным условиям для работы. Колумелла должна была располагаться перпендикулярно барабанной полости, быть маленькой и достаточно легкой, чтобы уменьшить ее инерцию , и подвешиваться в полости, наполненной воздухом. У современных видов, чувствительных к частотам выше 1 кГц , основание колумеллы составляет 1/20 площади барабанной перепонки. Однако у ранних амфибий колумелла была слишком большой, из-за чего площадь подножки была слишком большой, что мешало слышать высокие частоты. Таким образом, похоже, что они могли слышать только звуки высокой интенсивности и низкой частоты, а колумелла, скорее всего, просто поддерживала черепную коробку, прижатую к щеке.
Только в раннем триасе, примерно через сто миллионов лет после завоевания суши, барабанное среднее ухо развилось (независимо) у всех четвероногих. [120] Примерно пятьдесят миллионов лет спустя (поздний триас) у млекопитающих колумелла еще больше сократилась и превратилась в стремечко .
См. также
[ редактировать ]- Форма тела
- Геологическая шкала времени
- Шестиногий
- Морские четвероногие
- Осьминог
- Доисторическая жизнь
- Четвероногие § Четвероногие против четвероногих
Ссылки
[ редактировать ]- ^ Jump up to: а б Марьянович, Д. (2021). «Создание калибровочной колбасы на примере повторной калибровки транскриптомного древа времени челюстных позвоночных» . Границы генетики . 12 . 521693. дои : 10.3389/fgene.2021.521693 . ПМК 8149952 . ПМИД 34054911 .
- ^ Jump up to: а б с Недзведский, Гжегож; Шрек, Питер; Наркевич, Катажина; Наркевич, Марек; Альберг, Пер Э. (7 января 2010 г.). «Следы четвероногих из раннего среднего девона Польши». Природа . 463 (7277): 43–48. Бибкод : 2010Natur.463...43N . дои : 10.1038/nature08623 . ISSN 0028-0836 . ПМИД 20054388 . S2CID 4428903 .
- ^ Хачек, Б.; Кори, CJ (1896). зоотомия в пятнадцати лекциях ( Элементарная на немецком языке). Йена: Густав Фишер.
- ^ де Кейрос, К.; Кантино, PD; Готье, Ж.А., ред. (2020). « Четвероногие Б. Хатчек и К. Дж. Кори 1896 [М. Лорин], преобразованное название клады». Филонимы: дополнение к Филокоду . Бока-Ратон: CRC Press. стр. 759–764. ISBN 978-1-138-33293-5 .
- ^ «четвероногий» . Dictionary.com Полный (онлайн). nd
- ^ «четвероногие» . Словарь Merriam-Webster.com . Проверено 30 декабря 2022 г.
- ^ Ди-Пои, Николя; Монтойя-Бургос, Хуан И.; Миллер, Хилари; и др. (04 марта 2010 г.). «Изменения в структуре и функции Hox-генов в ходе эволюции чешуйчатого строения тела» . Природа . 464 (7285): 99–103. Бибкод : 2010Natur.464...99D . дои : 10.1038/nature08789 . ПМИД 20203609 . S2CID 205219752 . Проверено 6 июля 2023 г.
- ^ Jump up to: а б с Наркевич, Катажина; Наркевич, Марек (январь 2015 г.). «Возраст старейших следов четвероногих из Зачелме, Польша». Летайя . 48 (1): 10–12. Бибкод : 2015Лета..48...10Н . дои : 10.1111/лет.12083 . ISSN 0024-1164 .
- ^ Лонг Дж. А., Гордон М. С. (сентябрь – октябрь 2004 г.). «Величайший шаг в истории позвоночных: палеобиологический обзор перехода рыб от четвероногих» . Физиол. Биохим. Зоол . 77 (5): 700–19. дои : 10.1086/425183 . ПМИД 15547790 . S2CID 1260442 . Архивировано из оригинала 12 апреля 2016 г. Проверено 9 апреля 2011 г. в формате PDF. Архивировано 29 октября 2013 г. на Wayback Machine.
- ^ Шубин Н. (2008). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Нью-Йорк: Книги Пантеона. ISBN 978-0-375-42447-2 .
- ^ Лорин 2010 , стр. 163
- ^ Кановиль, Аврора; Лорен, Мишель (июнь 2010 г.). «Эволюция микроанатомии плечевой кости и образа жизни амниот и некоторые комментарии к палеобиологическим выводам» . Биологический журнал Линнеевского общества . 100 (2): 384–406. дои : 10.1111/j.1095-8312.2010.01431.x .
- ^ Лорен, Мишель ; Кановиль, Аврора; Кильяк, Александра (август 2009 г.). «Использование палеонтологических и молекулярных данных супердеревьев для сравнительных исследований: на примере микроанатомии бедренной кости лисамфибий» . Журнал анатомии . 215 (2): 110–123. дои : 10.1111/j.1469-7580.2009.01104.x . ПМК 2740958 . ПМИД 19508493 .
- ^ Jump up to: а б Лорен, Мишель (01 марта 2002 г.). «Филогения четвероногих, происхождение земноводных и определение названия четвероногих» . Систематическая биология . 51 (2): 364–369. дои : 10.1080/10635150252899815 . ISSN 1076-836X . ПМИД 12028737 .
- ^ Jump up to: а б Андерсон, Джейсон С. (1 сентября 2002 г.). «Использование известных имен в филогенетической номенклатуре: ответ Лорину» . Систематическая биология . 51 (5): 822–827. дои : 10.1080/10635150290102447 . ISSN 1076-836X . ПМИД 12396594 .
- ^ Jump up to: а б с д Рута, М.; Коутс, Мичиган; Квик, DLJ (2003). «Возвращение к ранним взаимоотношениям четвероногих» (PDF) . Биологические обзоры . 78 (2): 251–345. дои : 10.1017/S1464793102006103 . ПМИД 12803423 . S2CID 31298396 .
- ^ Jump up to: а б с д Лорен, Мишель; Андерсон, Джейсон С. (1 февраля 2004 г.). Саймон, Крис (ред.). «Значение названия четвероногих в научной литературе: обмен» . Систематическая биология . 53 (1): 68–80. дои : 10.1080/10635150490264716 . ISSN 1076-836X . ПМИД 14965901 . S2CID 15922260 .
- ^ Кейрос, Кевин де; Кантино, Филип Д.; Готье, Жак А. (2020). «Stegocephali ED Cope 1868 [М. Лорин], преобразованное название клады». В Де Кейруше, Кевин; Кантино, Филип; Готье, Жак (ред.). Филонимы: дополнение к PhyloCode (1-е изд.). Бока-Ратон: CRC Press. дои : 10.1201/9780429446276 . ISBN 9780429446276 . S2CID 242704712 .
- ^ Clack 2012 , стр. 87–9.
- ^ Лорен, Мишель; Жирондо, Марк; де Риклес, Арманд (2000). «Ранняя эволюция четвероногих» (PDF) . Тенденции в экологии и эволюции . 15 (3): 118–123. дои : 10.1016/S0169-5347(99)01780-2 . ISSN 0169-5347 . ПМИД 10675932 . Архивировано из оригинала (PDF) 22 июля 2012 г. Проверено 8 июня 2015 г.
- ^ Лаурин 2010 , с. 9
- ^ Бентон 2009 , с. 99
- ^ Марьянович Д., Лаурин М. (2008). «Оценка доверительных интервалов для стратиграфических диапазонов высших таксонов: случай Lissamphibia» (PDF) . Acta Palaeontologica Polonica . 53 (3): 413–432. дои : 10.4202/app.2008.0305 . S2CID 53592421 . Архивировано (PDF) из оригинала 29 октября 2013 г. Проверено 17 января 2013 г.
- ^ Jump up to: а б Сахни, С.; Бентон, MJ; Ферри, Пенсильвания (август 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Уорд, ПД; Бота, Дж.; Бьюик, Р.; Кок, Миссури; Эрвин, Д.Х.; Гарриссон, GH; Киршвинк, Дж.Л.; Смит, Р. (4 февраля 2005 г.). «Резкое и постепенное вымирание наземных позвоночных поздней перми в бассейне Кару, Южная Африка» (PDF) . Наука . 307 (5710): 709–714. Бибкод : 2005Sci...307..709W . CiteSeerX 10.1.1.503.2065 . дои : 10.1126/science.1107068 . ПМИД 15661973 . S2CID 46198018 . Архивировано из оригинала (PDF) 13 августа 2012 года . Проверено 28 октября 2017 г.
- ^ Jump up to: а б с д и ж г час «Сводная статистика» . Красный список исчезающих видов МСОП . 2023.1 . Проверено 5 февраля 2024 г. Таблица 1а: Количество видов, оцененное по отношению к общему числу описанных видов, и количество видов, находящихся под угрозой исчезновения, по основным группам организмов
- ^ Jump up to: а б Ромер, А.С. (1949). Тело позвоночного . Филадельфия: У. Б. Сондерс. (2-е изд. 1955 г.; 3-е изд. 1962 г.; 4-е изд. 1970 г.)
- ^ Ллойд, Германия (1961). «Развитие теории классификации животных Аристотеля». Фронезис . 6 (1): 59–81. дои : 10.1163/156852861X00080 . JSTOR 4181685 .
- ^ Линней, Чарльз (1758). Система природы по трем царствам природы: по классам, порядкам, родам, видам, с признаками, различиями, синонимами, местами (на латыни) ( 10-е изд .). Стокгольм: Лоуренс Сальвиус. Архивировано из оригинала 10 октября 2008 г. Проверено 13 января 2018 г.
- ^ Латрейля Латрейель, Пенсильвания (1804 г.): Nouveau Dictionnaire à Histoire Naturelle, xxiv., цитируется в «Естественных семьях животного мира» , изложенных кратко и в аналитическом порядке , 1825 г.
- ^ Смит, CH (2005). «Ромер, Альфред Шервуд (США, 1894–1973)» . Архивировано 12 октября 2008 г. в Wayback Machine . Университет Западного Кентукки
- ^ Бентон, MJ (1998). «Качество летописи окаменелостей позвоночных». стр. 269–303 в Доноване, С.К. и Поле, CRC (ред.), Адекватность летописи окаменелостей , рис. 2. Нью-Йорк: Wiley.
- ^ Нил, JD (редактор) (2006). Физиология репродукции Нобила и Нила (3-е изд.). Том 2. Академическая пресса . п. 2177.
- ^ Фортуни, Дж.; Болет, А.; Селлес, АГ; Картанья, Дж.; Галобарт, А. (2011). «Новые сведения о пермских и триасовых позвоночных животных Пиренейского полуострова с упором на Пиренейский и Каталонский бассейны» (PDF) . Журнал иберийской геологии . 37 (1): 65–86. дои : 10.5209/rev_JIGE.2011.v37.n1.5 . Архивировано (PDF) из оригинала 17 мая 2011 г. Проверено 4 декабря 2012 г.
- ^ Хильдебранд, М.; Дж. Э. Гослоу-младший (2001). Анализ строения позвоночных . больной. Виола Хильдебранд. Нью-Йорк: Уайли. п. 429. ИСБН 978-0-471-29505-1 .
- ^ Clack 2012 , стр. 125–6.
- ^ МакГи 2013 , с. 92
- ^ Клак 2012 , с. 132
- ^ Лорин 2010 , стр. 64–8
- ^ Штайер, 2012 , стр. 37–8.
- ^ Клак 2012 , с. 76
- ^ МакГи 2013 , с. 75
- ^ МакГи 2013 , стр. 74–75.
- ^ Clack 2012 , стр. 82–4.
- ^ Штайер, 2012 , стр. 17–23.
- ^ Жанвье, Филипп; Клеман, Гаэль (7 января 2010 г.). «Палеонтология: происхождение мутных четвероногих». Природа . 463 (7277): 40–41. Бибкод : 2010Natur.463...40J . дои : 10.1038/463040a . ISSN 0028-0836 . ПМИД 20054387 . S2CID 447958 .
- ^ МакГи 2013 , стр. 79–81.
- ^ Клак 2012 , с. 126
- ^ Лукас, Спенсер Г. (2015). «Тинопус и критический обзор следов девонских четвероногих». Ихнос . 22 (3–4): 136–154. Бибкод : 2015Ично..22..136Л . дои : 10.1080/10420940.2015.1063491 . S2CID 130053031 .
- ^ Наркевич, Марек; Грабовский, Яцек; Наркевич, Катажина; Недзведский, Гжегож; Реталлак, Грегори Дж.; Шрек, Питер; Де Влишоувер, Дэвид (15 февраля 2015 г.). «Палеообстановка доломитов Эйфеля с самыми ранними тропами четвероногих (горы Святого Креста, Польша)». Палеогеография, Палеоклиматология, Палеоэкология . 420 : 173–192. Бибкод : 2015PPP...420..173N . дои : 10.1016/j.palaeo.2014.12.013 . ISSN 0031-0182 .
- ^ Стоссель, И. (1995) Открытие новой тропы девонских четвероногих на юго-западе Ирландии. Журнал Геологического общества, Лондон, 152, 407–413.
- ^ Стоссел, И., Уильямс, Э.А. и Хиггс, К.Т. (2016) Ихнология и среда отложения следов тетрапод на острове Валентия в среднем девоне, юго-запад Ирландии. Палеогеография, Палеоклиматология, Палеоэкология, 462, 16–40.
- ^ Clack 2012 , стр. 117–8.
- ^ Лаурин 2010 , с. 85
- ^ Jump up to: а б МакГи, 2013 , стр. 103–4.
- ^ Каллиер, В.; Клак, Дж.А.; Альберг, ЧП (2009). «Контрастные траектории развития самых ранних известных передних конечностей четвероногих». Наука . 324 (5925): 364–367. Бибкод : 2009Sci...324..364C . дои : 10.1126/science.1167542 . ISSN 0036-8075 . ПМИД 19372425 . S2CID 28461841 .
- ^ Клак 2012 , стр. 147.
- ^ Клак 2012 , стр. 159.
- ^ Пирс, Стефани Э.; Клак, Дженнифер А.; Хатчинсон, Джон Р. (2012). «Трехмерная подвижность суставов конечностей у ранних четвероногих ихтиостега» . Природа . 486 (7404): 523–6. Бибкод : 2012Natur.486..523P . дои : 10.1038/nature11124 . ISSN 0028-0836 . ПМИД 22722854 . S2CID 3127857 .
- ^ Гесс, Роберт; Альберг, Пер Эрик (8 июня 2018 г.). «Фауна четвероногих из девонского полярного круга» . Наука . 360 (6393): 1120–1124. Бибкод : 2018Sci...360.1120G . дои : 10.1126/science.aaq1645 . ПМИД 29880689 . S2CID 46965541 .
- ^ МакГи 2013 , стр. 214–5.
- ^ Jump up to: а б Классенс, Леон; Андерсон, Джейсон С.; Смитсон, Тим; Мански, Крис Ф.; Мейер, Таран; Клак, Дженнифер (27 апреля 2015 г.). «Разнообразная фауна четвероногих у подножия Ромерс-Гэп » . ПЛОС ОДИН . 10 (4): e0125446. Бибкод : 2015PLoSO..1025446A . дои : 10.1371/journal.pone.0125446 . ISSN 1932-6203 . ПМЦ 4411152 . ПМИД 25915639 .
- ^ МакГи 2013 , с. 78
- ^ МакГи 2013 , стр. 263–4.
- ^ «Исследовательский проект: Среднепалеозойский биотический кризис: определение траектории эволюции четвероногих» . Архивировано из оригинала 12 декабря 2013 г. Проверено 6 апреля 2014 г.
- ^ Джордж Р. Макги-младший (12 ноября 2013 г.). Когда вторжение на сушу провалилось: наследие девонского вымирания . Издательство Колумбийского университета. ISBN 9780231160568 . Архивировано из оригинала 08 августа 2020 г. Проверено 25 апреля 2020 г.
- ^ Холл, Брайан К. (15 сентября 2008 г.). Плавники в конечности: эволюция, развитие и трансформация . Издательство Чикагского университета. ISBN 9780226313405 . Архивировано из оригинала 9 августа 2020 г. Проверено 25 апреля 2020 г.
- ^ Сахни, С., Бентон, М.Дж. и Фалькон-Лэнг, Х.Дж. (2010). «Разрушение тропических лесов спровоцировало диверсификацию пенсильванских четвероногих в Еврамерике». Геология . 38 (12): 1079–1082. Бибкод : 2010Geo....38.1079S . дои : 10.1130/G31182.1 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сахни, Сарда и Бентон, Майкл Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества B: Биологические науки . 275 (1636): 759–765. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Бёме М., Спасов Н., Фусс Дж., Трешер А., Дин А.С., Прието Дж. и др. (ноябрь 2019 г.). «Дополнительная информация: новая миоценовая обезьяна и передвижение предка человекообразных обезьян и человека» (PDF) . Природа . 575 (7783): 489–493. Бибкод : 2019Natur.575..489B . дои : 10.1038/s41586-019-1731-0 . ПМИД 31695194 . S2CID 207888156 .
- ^ Шварц, Б. (2012). «Морской стебель-четвероногий из девона западной части Северной Америки» . ПЛОС ОДИН . 7 (3): e33683. Бибкод : 2012PLoSO...733683S . дои : 10.1371/journal.pone.0033683 . ПМК 3308997 . ПМИД 22448265 .
- ^ Сигурдсен, Тронд; Грин, Дэвид М. (июнь 2011 г.). «Происхождение современных земноводных: переоценка» . Зоологический журнал Линнеевского общества . 162 (2): 457–469. дои : 10.1111/j.1096-3642.2010.00683.x . ISSN 0024-4082 .
- ^ Jump up to: а б Бентон, Майкл (4 августа 2014 г.). Палеонтология позвоночных . Уайли. п. 398. ИСБН 978-1-118-40764-6 . Архивировано из оригинала 19 августа 2020 года . Проверено 2 июля 2015 г.
- ^ Наринс, Питер М.; Фэн, Альберт С.; Фэй, Ричард Р. (11 декабря 2006 г.). Слух и звуковая коммуникация у амфибий . Springer Science & Business Media. п. 16. ISBN 978-0-387-47796-1 . Архивировано из оригинала 20 августа 2020 года . Проверено 2 июля 2015 г.
- ^ С. Блэр Хеджес; Судхир Кумар (23 апреля 2009 г.). Древо времени жизни . ОУП Оксфорд. стр. 354–. ISBN 978-0-19-156015-6 . Архивировано из оригинала 18 августа 2020 года . Проверено 13 марта 2016 г.
- ^ Коутс, Майкл И.; Рута, Марчелло; Фридман, Мэтт (2008). «Со времен Оуэна: изменение взглядов на раннюю эволюцию четвероногих». Ежегодный обзор экологии, эволюции и систематики . 39 (1): 571–592. doi : 10.1146/annurev.ecolsys.38.091206.095546 . ISSN 1543-592X . JSTOR 30245177 .
- ^ Лорин 2010 , стр. 133
- ^ Кэрролл, Роберт Л. (2007). «Палеозойское происхождение саламандр, лягушек и червяг» . Зоологический журнал Линнеевского общества . 150 (с1): 1–140. дои : 10.1111/j.1096-3642.2007.00246.x . ISSN 0024-4082 .
- ^ Андерсон, Джейсон С. (декабрь 2008 г.). «Фокусный обзор: происхождение современных амфибий». Эволюционная биология . 35 (4): 231–247. Бибкод : 2008EvBio..35..231A . дои : 10.1007/s11692-008-9044-5 . ISSN 0071-3260 . S2CID 44050103 .
- ^ Фробиш, Северная Каролина; Шох, Р.Р. (2009). «Тестирование влияния миниатюризации на филогению: палеозойские диссорофоидные амфибии» . Систематическая биология . 58 (3): 312–327. дои : 10.1093/sysbio/syp029 . ISSN 1063-5157 . ПМИД 20525586 .
- ^ Возвышение четвероногих: как наши ранние предки оставили воду, чтобы ходить по суше
- ^ Clack 2012 , стр. 29, 45–6.
- ^ Клак 2012 , стр. 207, 416.
- ^ Тейлор, член парламента (2014). «Количественная оценка влияния межпозвонкового хряща на нейтральную позу шеи динозавров-зауроподов» . ПерДж . 2 : е712. дои : 10.7717/peerj.712 . ПМЦ 4277489 . ПМИД 25551027 .
- ^ Clack 2012 , стр. 373–4.
- ^ Шмидт-Киттлер, Норберт; Фогель, Клаус (6 декабря 2012 г.). Конструктивная морфология и эволюция . Springer Science & Business Media. стр. 151–172. ISBN 978-3-642-76156-0 . Архивировано из оригинала 19 августа 2020 года . Проверено 15 июля 2015 г.
- ^ Менье, Франсуа Ж.; Лорен, Мишель (январь 2012 г.). «Микроанатомическое и гистологическое исследование длинных костей плавников девонского саркоптерига Eusthenopteron foordi ». Акта Зоология . 93 (1): 88–97. дои : 10.1111/j.1463-6395.2010.00489.x .
- ^ Нинан, Дж. М.; Рута, М.; Клак, Дж.А.; Рэйфилд, EJ (22 апреля 2014 г.). «Биомеханика питания у Acanthostega и при переходе рыба-четвероногие» . Труды Королевского общества B: Биологические науки . 281 (1781): 20132689. doi : 10.1098/rspb.2013.2689 . ISSN 0962-8452 . ПМЦ 3953833 . ПМИД 24573844 .
- ^ Клак 2012 , с. 49 212
- ^ Батлер, Энн Б.; Ходос, Уильям (2 сентября 2005 г.). Сравнительная нейроанатомия позвоночных: эволюция и адаптация . Джон Уайли и сыновья. п. 38. ISBN 978-0-471-73383-6 . Архивировано из оригинала 18 августа 2020 года . Проверено 11 июля 2015 г.
- ^ Клаудсли-Томпсон, Джон Л. (6 декабря 2012 г.). Разнообразие земноводных и рептилий: Введение . Springer Science & Business Media. п. 117. ИСБН 978-3-642-60005-0 . Архивировано из оригинала 18 августа 2020 года . Проверено 11 июля 2015 г.
- ^ Клак, Дж. А. (2007). «Сравнительная биология» . Интегративная и сравнительная биология . 47 (4): 510–523. дои : 10.1093/icb/icm055 . ISSN 1540-7063 . ПМИД 21672860 .
- ^ МакГи 2013 , стр. 111, 139–41.
- ^ Скотт, AC; Гласспул, Эй-Джей (18 июля 2006 г.). «Многообразие палеозойских пожарных систем и колебания концентрации кислорода в атмосфере» . Труды Национальной академии наук . 103 (29): 10861–10865. Бибкод : 2006PNAS..10310861S . дои : 10.1073/pnas.0604090103 . ISSN 0027-8424 . ПМЦ 1544139 . ПМИД 16832054 .
- ^ Клак 2012 , стр. 140.
- ^ Клак 2012 , стр. 166.
- ^ Сьюс, Ханс-Дитер ; Фрейзер, Николас К. (13 августа 2013 г.). Триасовая жизнь на суше: Великий переход . Издательство Колумбийского университета. п. 85. ИСБН 978-0-231-50941-1 . Архивировано из оригинала 20 августа 2020 года . Проверено 21 июля 2015 г.
- ^ Вицманн, Флориан; Брейнерд, Элизабет (2017). «Моделирование физиологии водных темноспондилов Archegosaurus decheni из ранней перми Германии» . Ископаемый рекорд . 20 (2): 105–127. Бибкод : 2017FossR..20..105W . doi : 10.5194/fr-20-105-2017 .
- ^ Клак 2012 , стр. 23.
- ^ Лорин 2010 , стр. 36–7
- ^ МакГи 2013 , стр. 68–70.
- ^ Вебстер, Дуглас; Вебстер, Молли (22 октября 2013 г.). Сравнительная морфология позвоночных . Эльзевир Наука. стр. 372–5. ISBN 978-1-4832-7259-7 . Архивировано из оригинала 19 августа 2020 года . Проверено 22 мая 2015 г.
- ^ Бентон 2009 , с. 78
- ^ Клак 2012 , стр. 238.
- ^ Clack 2012 , стр. 73–4.
- ^ Брейнерд, Элизабет Л.; Оверкович, Томаш (2006). «Функциональная морфология и эволюция аспирационного дыхания четвероногих» . Респираторная физиология и нейробиология . 154 (1–2): 73–88. дои : 10.1016/j.resp.2006.06.003 . ISSN 1569-9048 . ПМИД 16861059 . S2CID 16841094 . Архивировано из оригинала 04 сентября 2020 г. Проверено 24 ноября 2018 г.
- ^ Меррелл, Эллисон Дж.; Кардон, Габриэль (2013). «Развитие диафрагмы — скелетной мышцы, необходимой для дыхания млекопитающих» . Журнал ФЭБС . 280 (17): 4026–4035. дои : 10.1111/февраль 12274 . ISSN 1742-464X . ПМЦ 3879042 . ПМИД 23586979 .
- ^ Эванс, Дэвид Х.; Клэйборн, Джеймс Б. (15 декабря 2005 г.). Физиология рыб, третье издание . ЦРК Пресс. п. 107. ИСБН 978-0-8493-2022-4 . Архивировано из оригинала 19 августа 2020 года . Проверено 28 июля 2015 г.
- ^ Грэм, Джеффри Б.; Вегнер, Николас К.; Миллер, Лорен А.; Еврей Кори Дж.; Лай, Н. Чин; Берквист, Рэйчел М.; Фрэнк, Лоуренс Р.; Лонг, Джон А. (январь 2014 г.). «Дыхательное воздушное дыхание у многокрылых рыб и его значение для воздушного дыхания стеблевых четвероногих» . Природные коммуникации . 5 : 3022. Бибкод : 2014NatCo...5.3022G . дои : 10.1038/ncomms4022 . ISSN 2041-1723 . ПМИД 24451680 . Архивировано из оригинала 04 сентября 2020 г. Проверено 24 ноября 2018 г.
- ^ Викариус, Мэтью К.; Сир Жан-Ив (апрель 2009 г.). «Покровный скелет четвероногих: происхождение, эволюция и развитие» . Журнал анатомии . 214 (4): 441–464. дои : 10.1111/j.1469-7580.2008.01043.x . ISSN 0021-8782 . ПМК 2736118 . ПМИД 19422424 .
- ^ Clack 2012 , стр. 233–7.
- ^ Нельсон, Дж. А. (март 2014 г.). «Ломать ветер, чтобы выжить: рыбы, которые дышат воздухом кишкой». Журнал биологии рыб . 84 (3): 554–576. Бибкод : 2014JFBio..84..554N . дои : 10.1111/jfb.12323 . ISSN 0022-1112 . ПМИД 24502287 .
- ^ Клак 2012 , с. 235
- ^ Янис, СМ; Девлин, К.; Уоррен, Делавэр; Витцманн, Ф. (август 2012 г.). «Кожная кость у ранних четвероногих: палеофизиологическая гипотеза адаптации к наземному ацидозу» . Труды Королевского общества B: Биологические науки . 279 (1740): 3035–3040. дои : 10.1098/rspb.2012.0558 . ISSN 0962-8452 . ПМЦ 3385491 . ПМИД 22535781 .
- ^ Clack 2012 , стр. 235–7.
- ^ Понселе, Г., и Шимелд, С.М. (2020). Эволюционное происхождение обонятельной системы позвоночных. Открытая биол. 10:200330. дои: 10.1098/rsob.200330
- ^ Дэвид М. Хант; Марк В. Хэнкинс; Шон П. Коллин; Н. Джастин Маршалл (4 октября 2014 г.). Эволюция визуальных и невизуальных пигментов . Спрингер. стр. 165–. ISBN 978-1-4614-4355-1 . Архивировано из оригинала 18 августа 2020 года . Проверено 13 марта 2016 г.
- ^ Ставенга, генеральный директор; де Грип, WJ; Пью, EN (30 ноября 2000 г.). Молекулярные механизмы зрительной трансдукции . Эльзевир. п. 269. ИСБН 978-0-08-053677-4 . Архивировано из оригинала 20 августа 2020 года . Проверено 14 июня 2015 г.
- ^ Лазарева Ольга Ф.; Симидзу, Тору; Эдвард А. Вассерман (19 апреля 2012 г.). Как животные видят мир: сравнительное поведение, биология и эволюция зрения . ОУП США. п. 459. ИСБН 978-0-19-533465-4 . Архивировано из оригинала 19 августа 2020 года . Проверено 14 июня 2015 г.
- ^ Кристенсен, Кристиан Бек; Лауридсен, Хенрик; Кристенсен-Дальсгаард, Якоб; Педерсен, Майкл; Мэдсен, Питер Теглберг (2015). «Лучше, чем рыба на суше? Слышать метаморфозы у саламандр» . Труды Королевского общества B: Биологические науки . 282 (1802). дои : 10.1098/рспб.2014.1943 . ПМЦ 4344139 . ПМИД 25652830 . Архивировано из оригинала 22 апреля 2016 г. Проверено 20 января 2016 г.
Дальнейшее чтение
[ редактировать ]- Бентон, Майкл (5 февраля 2009 г.). Палеонтология позвоночных (3-е изд.). Джон Уайли и сыновья. п. 1. ISBN 978-1-4051-4449-0 . Проверено 10 июня 2015 г.
- Клак, Дж. А. (2012). Набирает силу: происхождение и эволюция четвероногих (2-е изд.). Блумингтон, Индиана, США: Издательство Университета Индианы. ISBN 9780253356758 .
- Лорен, Мишель (2010). Как позвоночные покинули воду . Издательство Калифорнийского университета. ISBN 978-0-520-26647-6 . Проверено 26 мая 2015 г.
- МакГи, Джордж Р. младший (2013). Когда вторжение на сушу провалилось: наследие девонского вымирания . Издательство Колумбийского университета. ISBN 978-0-231-16057-5 . Проверено 2 мая 2015 г.
- Стейер, Себастьян (2012). Земля до динозавров . Издательство Университета Индианы. п. 59. ИСБН 978-0-253-22380-7 . Проверено 1 июня 2015 г.
- Клак, Дженнифер А. (2009). «Переход от плавника к конечностям: новые данные, интерпретации и гипотезы из палеонтологии и биологии развития». Ежегодный обзор наук о Земле и планетах . 37 (1): 163–179. Бибкод : 2009AREPS..37..163C . doi : 10.1146/annurev.earth.36.031207.124146 .
- Холл, Брайан К., изд. (2007). Плавники в конечности: эволюция, развитие и трансформация . Чикаго: Издательство Чикагского университета. ISBN 978-0-226-31340-5 .
- Лонг Дж.А., Янг Г.К., Холланд Т., Сенден Т.Дж., Фицджеральд Э.М. (ноябрь 2006 г.). «Исключительная девонская рыба из Австралии проливает свет на происхождение четвероногих». Природа . 444 (7116): 199–202. Бибкод : 2006Natur.444..199L . дои : 10.1038/nature05243 . ПМИД 17051154 . S2CID 2412640 .
- Бентон, Майкл (2005). Палеонтология позвоночных (3-е изд.). Издательство Блэквелл.







| Базы данных органов управления : Национальные |
|---|