Тазовая шпора

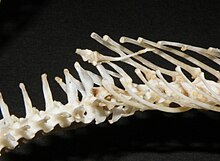
Тазовые шпоры (также известные как рудиментарные ноги) — это внешние выступы, обнаруживаемые вокруг клоаки у некоторых надсемейств змей , принадлежащих к большему инфраотряду Alethinophidia . [1] Эти шпоры состоят из остатков бедренной кости, которая затем покрывается роговичной шпорой или когтеобразной структурой. [1] Эта бедренная кость происходит от предковых задних конечностей, обнаруженных у самого недавнего общего предка современных змей и других рептилий клады Toxicofera , многие из которых имеют полностью функциональные передние и задние конечности. [2] В связи с тем, что шпоры происходят от наследственного состояния функциональных ног, но больше не предназначены специально для передвижения, эти структуры соответствуют критериям, позволяющим считаться рудиментарными . [3] Тем не менее, использование этих структур было тщательно задокументировано. Виды, имеющие внешние шпоры, имеют соответствующие мышцы, неврологические структуры и васкуляризацию, обеспечивающие независимое движение. [3] Шпоры более выражены и заметны у самцов, и их можно было наблюдать во время ухаживания. [4] Шпоры специально используются самцами для захвата и стимуляции самок во время ухаживания и спаривания. У некоторых видов самцы также используют свои шпоры, чтобы вступать в бой друг с другом. [3]
Ископаемая пластинка
[ редактировать ]Летопись окаменелостей змей невелика. [5] Тем не менее, многочисленные окаменелости документируют прогрессирование развития безногих представителей подотряда Serpentes . Вид вымершей змеи Najash rioegrina был впервые описан в 2006 году и предложен как самый ранний ветвящийся таксон подотряда Serpentes . [6] Окаменелости были найдены в районе Патагонии в Аргентине и датированы верхним меловым периодом . У Наджаша рионегрины были обнаружены крестец, тазовый пояс и крепкие структуры задних конечностей за пределами грудной клетки, что привело исследователей к выводу, что эти задние конечности были пригодны для передвижения. [6] Значение этой находки велико, так как в этот период времени существовало еще три известных вида змей на ногах: Pachyrhachispromaticus , Haasiophis terrasanctus и Eupodophis descouensi , но все они были предсказаны как морские виды, и у всех из них отсутствовала область крестца. найден у N. rionegrina . [7] [6] Таким образом, палеонтологи с помощью филогенетического анализа пришли к выводу, что N. rionegrina является самым древним таксоном среди Serpentes , а три ранее описанных вымерших вида были более тесно связаны с современными змеями, принадлежащими к Alethinophidia . [6]
Недавний анализ многочисленных записей окаменелостей подтвердил эти выводы и дополнительно продемонстрировал редукцию структур таза и задних конечностей в этих линиях. [5] [2] Дополнительные доказательства того, что эти структуры являются плезиоморфными , можно найти в тазу некоторых современных таксонов, таких как Candoia carinata или Eunectes murinus . Эти таксоны обладают трехлучевым тазом, который можно наблюдать и в скелетах современных ящериц . [2]
Распределение
[ редактировать ]Наличие тазовых шпор у современных видов Serpentes ограничено. У большинства представителей Scolecophidia нет шпор, как и у представителей самой густонаселенной группы алетинофидий — Caenophidia . Однако шпоры присутствуют среди базальных клад Alethinophidia , в том числе у Booidea и Pythonoidea . [8] среди Amerophidia и среди одного представителя Uropeltoidea , Cylindrophiidae . Эти базальные клады иногда называют примитивными змеями, поскольку они считаются самыми ранними расходящимся таксонами Alethinophidia , клады, которая включает большинство описанных ныне живущих видов змей. [9] Присутствие и использование шпор у Booidea и Pythonoidea хорошо задокументировано — в этих надсемействах шпоры можно наблюдать как инструменты для ухаживания и конкуренции между самцами, и они имеют половой диморфизм . [3]
Хотя это наиболее известные таксоны, имеющие шпоры, существуют доказательства присутствия окостеневших рудиментарных структур и у других таксонов. Публикация 2019 года предоставила доказательства существования подобных окостеневших структур у видов Liotyphlops beui инфраотряда Scolecophidia . [2] Члены этой сестринской группы Scolecophidia плохо изучены из-за их загадочной природы и, как правило, небольшого размера, роющих и червеобразных. [8] Это исследование 2019 года является первым описанным появлением этих структур в семействе Anomalepididae , которое является одним из трех семейств Scolecophidia . [2]
Важность социального поведения
[ редактировать ]Многочисленные исследования были проведены по использованию тазовых шпор самцами надсемейства Booidea и Pythonoidea . Половой диморфизм шпор был официально описан Уильямом Х. Стикелем и Люсиль Ф. Стикель в 1946 году для рода Enygrus (более известного сегодня как Candoia ). [10] Авторы этого исследования отметили, что другие специалисты в этой области уже делали подобные наблюдения и раньше, но формально не исследовали эту тему специально. Выяснилось, что у самцов шпоры значительно крупнее, а у самок шпоры значительно короче, а иногда и отсутствуют снаружи. [10]
Доминирование
[ редактировать ]С тех пор исследования показали, что тазовые шпоры используются для демонстрации доминирования у многих видов. Использование шпор наряду с укусами наблюдалось в демонстрациях между самцами в находящейся в неволе группе индийских питонов ( Python molurus ), которые впоследствии сформировали линейную иерархию доминирования . Положение в этой иерархии напрямую коррелировало с количеством успешных спариваний самца. [11] Другое исследование обнаружило использование шпор мадагаскарскими удавами ( Sanzinia madagascariensis ), древесным видом. У этого вида исследователи не наблюдали поклевки. [12] Фактически, авторы заметили, что головы дерущихся самцов часто находились вне поля зрения друг друга. Вместо этого наблюдаемые самцы крепко сжимали друг друга задними областями своего тела, ориентируя при этом свои шпоры перпендикулярно телу . [12] Авторы предполагают, что эта форма боя шпорами адаптивна к древесной среде обитания вида. [12] Стоит также отметить, что борьба между самцами у видов, у которых нет шпор, таких как виды Caenophidia , опирается на особый репертуар поведения, который отличается от видов, имеющих шпоры. [13]
Поведение во время ухаживания
[ редактировать ]Подобно описанным наблюдениям за половым диморфизмом до их публикации, Уильям Х. Стикель и Люсиль Ф. Стикель также отметили, что другие наблюдения были сделаны за использованием тазовых шпор самцами на самках во время спаривания. [10] С тех пор эти наблюдения были официально исследованы и опубликованы для нескольких видов.
Одно из таких исследований обнаружило динамическое использование шпор самцами во время разных фаз ухаживания и спаривания у бирманских питонов ( Python molurus bivittatus ), при этом движения шпор вперед и назад различаются по скорости волнистости в зависимости от фазы. [14] Кроме того, самцы использовали шпоры, чтобы лучше расположить или приспособить клоаку самки для спаривания. Авторы отмечают, что использование шпор для боя у этого вида также можно наблюдать между самцами. [14] Отдельное исследование показало аналогичные результаты у алмазных питонов ( Morelia spilota ), вида, у которого не наблюдается никаких приступов борьбы между самцами. [15] У этого вида наблюдалось одновременное спаривание нескольких самцов с одной и той же самкой, при этом самцы не вступали в бой. Вместо этого было замечено, что самцы использовали шпоры, чтобы манипулировать хвостом самки. [15] Подтверждая эти результаты на одном из видов Нового Света , исследование, проведенное в 2023 году, задокументировало использование шпор при спаривании краснохвостыми удавами Boa constrictor . [3] Было замечено, что самцы неоднократно меняли ориентацию шпор между горизонтальным и вертикальным положениями, что, по мнению авторов, стимулировало мышечные сокращения у самок, что позволяло лучше выравнивать особей для спаривания. [3]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Пау, Ф.Х. (ред.). (2004). Герпетология (3-е изд.). Прентис Холл.
- ^ Перейти обратно: а б с д и Палчи, А., Хатчинсон, Миннесота, Колдуэлл, М.В., Смит, К.Т., и Ли, М.Ю. (2019). Гомологии и эволюционная редукция таза и задних конечностей у змей с первым сообщением об окостеневших тазовых остатках у аномалепидида (Liotyphlops beui). Зоологический журнал Линнеевского общества , zlz098. дои : 10.1093/zoolinnean/zlz098
- ^ Перейти обратно: а б с д и ж Анзай, Р.К., Элеутерио, Н.Ф., Оливейра Лима, Т.Д., Манфио, Р.Х., и Алмейда Сантос, SMD (2023). Использование тазовых шпор во время ухаживания и спаривания краснохвостого удава Boa constrictor. Герпетологический вестник , 163 , 35–36. дои : 10.33256/hb163.3536
- ^ Бошо, Р. (Ред.). (1994). Змеи: естественная история . Стерлинг Паб.
- ^ Перейти обратно: а б Гарберольо, Ф.Ф., Апестегия, С., Симоэнс, Т.Р., Палчи, А., Гомес, Р.О., Нидам, Р.Л., Ларссон, HCE, Ли, М.С.И, и Колдуэлл, М.В. (2019). Новые черепа и скелеты меловой многоногой змеи Наджаша и эволюция строения тела современной змеи. Достижения науки , 5 (11), eaax5833. два : 10.1126/sciadv.aax5833
- ^ Перейти обратно: а б с д Апестегия С. и Захер Х. (2006). Наземная змея мелового периода с крепкими задними конечностями и крестцом. Природа , 440 (7087), 1037–1040. дои : 10.1038/nature04413
- ^ Беллэрс, А.Д., и Андервуд, Г. (1951). Происхождение змей. Биологические обзоры , 26 (2), 193–237. дои : 10.1111/j.1469-185X.1951.tb00646.x
- ^ Перейти обратно: а б Змеи (Змеи) . (без даты). Получено 10 октября 2023 г. с https://www.ucl.ac.uk/museums-static/obl4he/vertebratediversity/serpentes_snakes.html .
- ^ Ли, М.С.И., Хугалл, А.Ф., Лоусон, Р., и Скэнлон, Дж.Д. (2007). Филогения змей (Serpentes): объединение морфологических и молекулярных данных с учетом вероятности, байесовского анализа и анализа экономии. Систематика и биоразнообразие , 5 (4), 371–389. дои : 10.1017/S1477200007002290
- ^ Перейти обратно: а б с Стикель, WH, и Стикель, LF (1946). Половой диморфизм тазовых шпор энгруса. Копейя , 1946 (1), 10. дои : 10.2307/1438812
- ^ Баркер, Д.Г., Мерфи, Дж.Б., и Смит, К.В. (1979). Социальное поведение в содержащейся в неволе группе индийских питонов Python molurus (Serpentes, boidae) с формированием линейной социальной иерархии. Копейя , 1979 (3), 466. дои : 10.2307/1443224
- ^ Перейти обратно: а б с Карпентер, СиСи, Мерфи, Дж.Б., и Митчелл, Лос-Анджелес (1978). Боевые поединки с использованием шпор у мадагаскарского удава (Sanzinia madagascariensis). Герпетология , 34 (2), 207–212. JSTOR 3891678
- ^ Джиллингем, JC (1980). Коммуникативное и боевое поведение черной крысиной змеи (Elaphe obsoleta). Герпетология , 36 (2), 120–127. JSTOR 3891474
- ^ Перейти обратно: а б Джиллингем, Дж. К., и Чемберс, Дж. А. (1982). Использование тазовых шпор и ухаживания у бирманского питона, python molurus bivittatus. Копейя , 1982 (1), 193. дои : 10.2307/1444292
- ^ Перейти обратно: а б Слип, ди-джей, и Шайн, Р. (1988). Репродуктивная биология и система спаривания алмазных питонов morelia spilota (Serpentes: Boidae). Герпетология , 44 (4), 396–404. JSTOR 3892404
Эта о змее статья незавершена . Вы можете помочь Википедии, расширив ее . |
Эта анатомии позвоночных статья, посвященная , незавершена . Вы можете помочь Википедии, расширив ее . |