Эустенодон
| Эустенодон Временной диапазон: Поздний девон
| |
|---|---|

| |
| Череп | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Саркоптеригии |
| Клэйд : | Тетраподоморфа |
| Клэйд : | Эотетраподиформные |
| Семья: | † Тристихоптериды |
| Род: | † Эустенодон Ярвик , 1952 год. |
| Разновидность | |

Эустенодон (по-гречески «сильный зуб» — eusthenes — означает «сильный», — odon означает «зуб») — вымерший род тристихоптеридных из тетраподоморфов позднего девона , существовавший между 383 и 359 миллионами лет назад ( фран - фамен ). . [ 3 ] [ 4 ] Они хорошо известны как космополитический род, останки которого были обнаружены в Восточной Гренландии , Австралии , Центральной России , Южной Африке , Пенсильвании и Бельгии . [ 5 ] [ 6 ] По сравнению с другими близкородственными родами Tristichopteridae клады , Eusthenodon был одной из крупнейших лопастно-плавниковых рыб (примерно 2,5 метра в длину) и одним из наиболее производных тристихоптерид наряду со своими близкими родственниками Cabonnichthys и Mandageria . [ 7 ] [ 4 ]
Большой размер, хищническая экология и эволюционно обусловленные черты, которыми обладает Eusthenodon , вероятно, способствовали его способности заселять и процветать в многочисленных местах по всему миру, упомянутых выше. Eusthenodon считается лишь одним из многих космополитических родов рыбной фауны « старого красного песчаника » верхнего девона. [ 3 ] [ 8 ] [ 9 ] В результате была выдвинута гипотеза, что диверсификация Eusthenodon морфологически и других сходных тристихоптерид не ограничивалась биогеографическими барьерами, а ограничивалась только их индивидуальной экологией и мобильностью. [ 9 ]
Большая часть останков евстенодона , обнаруженных в этих глобально распределенных местах, состояла в основном из черепных элементов и в основном неизвестна по полным скелетам. [ 5 ] [ 9 ] [ 8 ] Следовательно, большая часть доступной литературы, посвященной Eusthenodon, в первую очередь сосредоточена на тонкостях костей черепа, чтобы исследовать этот род, а другие делают выводы на основе известных признаков Tristichopteridae. [ 9 ] Йохансон и Альберг (1997) в своей оценке нового материала саркоптеригов представили такие выводы, предполагающие, что Eusthenodon, вероятно, обладал тем же трехфуркатным или ромбовидным хвостовым плавником с осевой лопастью, слегка повернутой дорсально, как это было у других тристихоптерид (названных Йохансоном эустеноптеридами). вместе с первым спинным плавником треугольной формы. [ 5 ]
История и именование
[ редактировать ]В 1952 году шведский палеонтолог Эрик Ярвик впервые описал первый вид — Eusthenodon wangsjoi из рода Eusthenodon . [ 10 ] Образец был извлечен в 1936 году из богатых окаменелостями отложений верхнедевонских отложений Восточной Гренландии, региона, который приобрел огромное внимание палеонтологов позвоночных после открытия ранних конечностей позвоночных Ichthyostega . [ 11 ] Настоящее название рода, Eusthenodon , относится к отчетливо большим бивням, присутствующим в верхней и нижней челюстях. [ 12 ]
Видовое название дано в честь Густава Венгсё как первооткрывателя. Первоначально Джарвик писал его как wängsjöi . Однако ICZN запрещает использование диакритических знаков в биномах. В 1962 году Воробьева изменила видовое название на waengsjoei . В 2009 году Дэниел Сниттинг и Хеннинг Блом отметили, что это ошибочное предположение о немецких умлаутов присутствии . Согласно статье 32.5.2.1 МКЗН правильное написание — wangsjoi . [ 13 ]
Описание
[ редактировать ]Череп
[ редактировать ]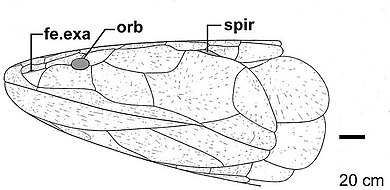
В своем первоначальном диагнозе первых останков Eusthenodon , опубликованном в 1952 году, Ярвик описывает особенности, присутствующие в останках Eusthenodon wangsjoi, включая те, которые являются важными признаками рыб-тристихоптерид (называемых Ярвиком ризодонтидами ), а также черты, уникальные для описанных останков. Видовые и диагностические признаки рода. [ 14 ] Заостренная голова Eusthenodon относительно большая по сравнению с другими близкородственными Osteolepiformes с короткими теменными щитками , которые способствуют расширению морды . [ 15 ] Лобноэтмоидальный щит крыши черепа у Eusthenodon заметно длиннее теменного щитка. [ 16 ] Соотношение длин лобно-этмоидального и теменного щитов использовалось палеонтологами как диагностический инструмент для различения таксонов , а в ряде случаев служило единственным отличительным признаком, разделяющим две группы (как это видно при разделении клад Eusthenopteron и Тристихоптер ). [ 8 ] [ 5 ] Среди эустеноптерид (тристихоптерид) существует тенденция, показывающая все более высокие значения этого соотношения у более производных родов, при этом Eusthenodon обладает самым высоким значением с соотношением лобноэтмоидального щита к теменному щиту 2,30. [ 8 ] Дальнейшее увеличение длины морды у многих видов четвероногих может также быть еще одним свидетельством, подтверждающим тенденцию к все более длинным лобноэтмоидальным щиткам, присутствующим в последующих кладах, тесно связанных с эвстеноптеридами, включая поздние эоподы. [ 8 ] Орбитальные отверстия , в которых расположены маленькие глаза евстенодона, были заметно меньше по размеру по сравнению с размером более крупного лобно-этмоидального щита. [ 12 ] [ 8 ] Расположенная кзади от глазничного окна, задняя надглазничная кость распространяется вниз вдоль окна и соприкасается со слезной костью . [ 17 ] В отличие от других Osteolepiformes, у которых также имеется задняя надглазничная кость, простирающаяся вентрально вдоль глазничного окна, контакт задней надглазничной кости со слезной костью является диагностическим признаком Eusthenodon и приводит к отделению скуловой и заглазничной костей от места сближения. глазничное окно. [ 14 ]
Положение и относительные размеры дополнительных окон, присутствующих у Eusthenodon , включая экзонарное окно, шишковидное отверстие и шишковидное отверстие, являются дополнительными диагностическими признаками этого рода. [ 12 ] [ 8 ] [ 4 ] Треугольное шишковидное окно хорошо известно у Eusthenodon краю или соприкасающимся с ним своим большим размером и характерным задним хвостом окна, приближающимся к заднему лобному . [ 12 ] [ 8 ] Напротив, шишковидное отверстие значительно меньше по размеру и расположено отчетливо кзади как от центра радиации лобной, так и заглазничной кости лобно-решетчатого щита. [ 12 ] При осмотре Eusthenodon черепа сверху можно увидеть окно exonarina, расположенное высоко и сбоку на морде. [ 12 ]
Из трех височных костей, составляющих теменной щит, присутствующих у Osteolepiformes ( межвисочной , надвисочной и вневисочной), наличие вневисочной кости в «постспиракулярном» положении определяется как смещение кости из латерального положения в надвисочное. в более задне-латеральное положение является важным и диагностическим признаком клады Tristichopteridae. [ 8 ] [ 17 ] Вневисочная кость, присутствующая у Eusthenodon, отличается своим полным постспиракулярным положением, в результате чего отсутствует контакт между надвисочной и вневисочной костями - состояние, которое, как известно, существует только у Eusthenodon . [ 8 ] Одна теория, объясняющая тенденцию, наблюдаемую в смещении вневисочной кости назад у более продвинутых рыб, предполагает, что изменение пропорций головы способствовало более обтекаемой форме тела и улучшало его маневренность и скорость в водной среде. [ 8 ]
Наружная щечная пластинка хорошо документирована у Eusthenodon: она в 3,5 раза длиннее теменного щита и в 3,0 раза длиннее теменного щита и в 3,0 раза длиннее его высоты. [ 10 ] Щечная пластинка и нижняя челюсть у Eusthenodon значительно длиннее, чем у любых других Osteolepiformes . У евстенодона нижняя челюсть уменьшается по высоте при движении от заднего конца к переднему и значительно ниже по высоте в передней части. [ 10 ]
Зубной ряд
[ редактировать ]Как следует из названия, у евстенодона большие бивни, которые выступают из верхней и нижней челюстей черепа. [ 17 ] В частности, вдоль средней линии морды на предчелюстной кости появляются два больших и толстых зуба . [ 5 ] Судя по неполному материалу, собранному после открытия евстенодона, самые большие бивни, по оценкам, имели длину не менее 50 миллиметров. [ 10 ] Эти два зуба уплощены в передне-заднем направлении и имеют характерные острые режущие края. [ 8 ] [ 4 ] В исследовании, представленном Гаэлем Клементом в 2009 году, в котором был описан недавно обнаруженный комплекс тристихоптерид, было обнаружено, что увеличенные зубы преимущественно находились на одной линии с зубным рядом предчелюстной кости и не встречались парами. [ 9 ] Следовательно, увеличенные предчелюстные зубы были описаны как «псевдо клыки», а не как настоящие клыки, которые , как считалось ранее, присутствовали у Eusthenodon . [ 9 ] Анализ горизонтального поперечного сечения первого клыка показывает простой и неправильно сложенный ортодонт. [ 5 ] В полости пульпы обнаруживается остеодентин. [ 8 ] [ 5 ] Наличие увеличенных псевдо-клыков на предчелюстной кости у Eusthenodon подтвердило его филогенетическое положение в кладе Tristichopteridae, поскольку аналогичные модели зубных рядов обнаружены у других близкородственных производных тристихоптерид. [ 9 ] Количество маленьких заостренных зубов вдоль зубного ряда также подтверждает тенденции развития зубных рядов с течением времени, поскольку у более производных родов обнаруживается большее количество зубов по сравнению с более примитивными видами, такими как Eusthenopteron . [ 8 ]
Несмотря на наличие набора предчелюстных ложных клыков, Eusthenodon и другие крупные тристихоптериды филогенетического происхождения демонстрируют сложный передний зубной ряд и характерные увеличенные зубные клыки. [ 5 ] [ 9 ] Слегка вогнутое зубчатое поле парасфеноидной кости приподнято у примитивных тристихоптерид, тогда как у Eusthenodon оно заметно утоплено . Кроме того, наличие характерной лезвиевидной вертикальной пластинки, присутствующей на переднем венечном отростке, существует у большинства других тристихоптерид, но отсутствует у производных родов, таких как Eusthenodon . [ 9 ] У тристихоптерид передний и средний венец несут по крайней мере одну пару клыков, тогда как у Eusthenodon задний венчик имеет две пары клыков. [ 7 ] [ 9 ] Кроме того, известно, что маргинальные венечные зубы присутствуют практически у всех других тристихоптерид (за исключением известного отсутствия у одного рода Cabonnicthys) , однако у Eusthenodon и близкородственного Mandageria в передней части передней части зуба отсутствует значительное количество маргинальных венечных зубов. челюсть. [ 8 ] Эта редукция маргинальных короноидных зубов подтверждает филогенетическую ассоциацию Eusthenodon , Mandageria и Cabonnichthys и служит производной характеристикой поздних тристихоптерид. [ 8 ] Эустенодон обладает небольшой парасимфизальной пластинкой, прикрепленной к селезенке посредством небольшого прикрепления пластинки к передней части мезиальной пластинки. [ 9 ] [ 14 ] Форма и размер парасимфизальной пластинки, представленной у Eusthenodon, присутствуют у всех тристихоптерид и являются диагностической характеристикой семейства. [ 9 ] [ 10 ]
Весы
[ редактировать ]В соответствии с признаками, описанными Бергом (1955) как важные диагностические признаки Tristichopteridae, Eusthenodon обладает пропорционально крупными, отчетливо круглыми чешуями без космина , которые имеют сетчатый рисунок гребней с редким появлением независимых бугорков. [ 18 ] [ 10 ] [ 19 ] Кроме того, каждая из этих круглых безкосминовых чешуй включает проксимальный центральный прикрепительный выступ, также характерный для Tristichopteridae. [ 8 ] [ 10 ] В отличие от большинства других тристихоптерид, в орнаменте чешуи Eusthenodon имеются гребни, образующие отчетливую сеть, тогда как чешуя Eusthenopteron, как правило, имеет орнамент из значительно более коротких гребней, присутствующих в не полностью сросшихся бугорках. [ 10 ] [ 8 ] Область перекрытия чешуек у Eusthenodon также больше, чем у Eusthenopteron . [ 10 ]
Классификация
[ редактировать ]Эустенодон принадлежит к семейству Tristichopteridae, подразделению отряда Osteolepiformes в рамках более крупного класса Sarcopterygii. [ 8 ] Sarcopterygii - основная клада, которая произошла от лучепёрых Actinopterygii с развитием лопастных плавников.
Филогения Tristichopteridae была описана Гаэлем Клементом, Дэниелом Сниттингом и П.Е. Альбергом (2008) после проведения максимально экономного анализа взаимосвязей внутри клады: [ 9 ]
Ссылки
[ редактировать ]- ^ Даунс, JP; Барбоза, Дж.; Даешлер, Э.Б. (2021). «Новый вид Eusthenodon (Sarcopterygii, Tristichopteridae) из верхнего девона (фамен) Пенсильвании, США, и обзор таксономии Eusthenodon ». Журнал палеонтологии позвоночных . 41 (3): e1976197. Бибкод : 2021JVPal..41E6197D . дои : 10.1080/02724634.2021.1976197 . S2CID 240453731 .
- ^ Даунс, JP; Осатчук, М.М.; Гудчайлд, ОА; Даешлер, Э.Б. (2023). «Второй вид Eusthenodon (Tristichopteridae, Sarcopterygii) из верхнедевонской (фаменской) формации Катскилл в Пенсильвании, США, и обзор глобального распространения Eusthenodon ». Журнал палеонтологии позвоночных . дои : 10.1080/02724634.2023.2201627 .
- ^ Jump up to: а б Блом, Хеннинг; Клак, Дженнифер; Альберг, Пер. (2007). «Девонские позвоночные из Восточной Гренландии: обзор состава и распространения фауны». Геодиверситас . 29 : 119–132 – через ResearchGate.
- ^ Jump up to: а б с д Клемент, Гаэль (2002). «Крупные Tristichopteridae (Sarcopterygii, Tetrapodomorpha) из позднефаменской формации Эвье в Бельгии» . Палеонтология . 45 (3): 577–593. Бибкод : 2002Palgy..45..577C . дои : 10.1111/1475-4983.00250 . ISSN 0031-0239 .
- ^ Jump up to: а б с д и ж г час Альберг, Пер Э.; Йохансон, Зерина (15 декабря 1997 г.). «Второй тристихоптерид (Sarcopterygii, Osteolepiformes) из верхнего девона Кановиндры, Новый Южный Уэльс, Австралия, и филогения Tristichopteridae». Журнал палеонтологии позвоночных . 17 (4): 653–673. Бибкод : 1997JVPal..17..653A . дои : 10.1080/02724634.1997.10011015 . ISSN 0272-4634 .
- ^ Лебедев О.А.; Захаренко Г.В.; Силантьев В.В.; Евдокимова, И.О. (2018). «Новые находки рыб в нижних слоях фамена (верхнего девона) Центральной России и местообитания хованщинских комплексов позвоночных» . Эстонский журнал наук о Земле . 67 (1): 59. дои : 10.3176/earth.2018.04 . ISSN 1736-4728 .
- ^ Jump up to: а б Альберг, Пер Э.; Йохансон, Зерина (15 декабря 1997 г.). «Второй тристихоптерид (Sarcopterygii, Osteolepiformes) из верхнего девона Кановиндры, Новый Южный Уэльс, Австралия, и филогения Tristichopteridae». Журнал палеонтологии позвоночных . 17 (4): 653–673. Бибкод : 1997JVPal..17..653A . дои : 10.1080/02724634.1997.10011015 . ISSN 0272-4634 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Борген, Ульф Дж.; Накрем, Ханс А. (30 сентября 2016 г.). «Морфология, филогения и систематика остеолепиформных рыб». Окаменелости и слои . Серия «Окаменелости и слои». 61 : 1–481. дои : 10.1002/9781119286448.ch1 . ISBN 9781119286431 . ISSN 2637-6032 .
- ^ Jump up to: а б с д и ж г час я дж к л м КЛЕМАН, ГАЭЛЬ; СНИТИНГ, ДЭНИЭЛ; АЛЬБЕРГ, ПЕР ЭРИК (2009). «Новый тристихоптерид (Sarcopterygii, Tetrapodomorpha) из формации Эвье верхнего фамена (верхний девон) Бельгии» (PDF) . Палеонтология . 52 (4): 823–836. Бибкод : 2009Palgy..52..823C . дои : 10.1111/j.1475-4983.2009.00876.x . ISSN 0031-0239 . Архивировано (PDF) из оригинала 20 июля 2018 г.
- ^ Jump up to: а б с д и ж г час я Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. стр. 54–68. OCLC 952685457 .
- ^ Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. п. 6. ОКЛК 952685457 .
- ^ Jump up to: а б с д и ж Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. п. 54. OCLC 952685457 .
- ^ Дэниел Сниттинг и Хеннинг Блом, 2009, «Исправление названий таксонов, содержащих диакритические знаки - примеры палеозойских позвоночных», Журнал палеонтологии позвоночных , 29 (1): 269-270
- ^ Jump up to: а б с Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. стр. 54–68. OCLC 952685457 .
- ^ Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. п. 55. OCLC 952685457 .
- ^ Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. п. 54. OCLC 952685457 .
- ^ Jump up to: а б с Ярвик, Эрик (1952). О рыбьевидном хвосте у ихтиостегидных стегоцефалов: с описаниями нового стегоцефала и нового кистеперого из верхнего девона Восточной Гренландии . Том. 114. К.А. Райтцель. стр. 54–68. OCLC 952685457 .
- ^ Йохансон, З.; Ричи, А. (1 января 2000 г.). «Рипидистианы (Sarcopterygii) из Хантерского алевролита (поздний фамен) недалеко от Гренфелла, Новый Южный Уэльс, Австралия» . Ископаемый рекорд . 3 (1): 111–136. doi : 10.5194/fr-3-111-2000 . ISSN 2193-0074 .
- ^ Берг, Л.С. (1958). Перевод стр. 161-288 из «Системы современных и ископаемых видов рыб и рыб» Берга, 1955 г. OCLC 1081960984 .





