Спатицефал
| Спатицефал Временной диапазон: ранний – поздний карбон ,
| |
|---|---|

| |
| Восстановление жизни Spathicephalus mirus | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Саркоптеригии |
| Клэйд : | Тетраподоморфа |
| Клэйд : | Эльписстостегалия |
| Клэйд : | Стегоцефалы |
| Суперсемейство: | † Бафетоидея |
| Семья: | † Спатицефалиды Бомонт, 1977 год. |
| Род: | † Спатицефал Ватсон, 1929 год. |
| Разновидность | |
Spathicephalus — вымерший род стеблевых четвероногих (ранних четырехконечных позвоночных), живших в середине каменноугольного периода . Род включает два вида: типовой вид S. mirus из Шотландии, известный по двум преимущественно целым черепам и другому черепному материалу, и вид S. pereger из Новой Шотландии, известный по единственному фрагменту черепной таблицы . Судя по материалу S. mirus , внешний вид Spathicephalus не похож на внешний вид любого другого раннего четвероногого: уплощенный череп квадратной формы и челюсти, усеянные сотнями очень маленьких долотообразных зубов. Тем не менее, Spathicephalus разделяет некоторые анатомические особенности с семейством стеблевых четвероногих под названием Baphetidae , что побудило большинство палеонтологов, изучавших этот род, поместить его в более крупную группу под названием Baphetoidea , часто как часть собственного монотипического семейства Spathicephalidae . Считается, что спатицефалы питались водными беспозвоночными, сочетая всасывание и фильтрацию. .
История обучения
[ редактировать ]Типовой вид Spathicephalus , S. mirus , был назван палеонтологом Д. М. С. Уотсоном в 1929 году. Уотсон описал семь ископаемых экземпляров из обнажения Рамблс -Айронстоун в городе Лоанхед в Мидлотиане , Шотландия. Железняк ) и является датируется поздним намюрским этапом (самый ранний верхний карбон частью группы известнякового угля . Эти экземпляры были обнаружены в 1880-х годах и включают в себя практически полный череп с обнаженным нёбом , отпечаток нижней части крыши черепа , правую часть задней части черепа и различные фрагменты челюстей. [ 2 ] В то время спатицефалы и другие четвероногие из Намюра в Шотландии были одними из старейших известных четвероногих, предшествовавших более известным скоплениям четвероногих позднего карбона из Британских угольных мер . В ноябре 1974 года шотландский палеонтолог Стэнли П. Вуд обнаружил дополнительные фрагменты черепа и челюсти спатицефала в карьере ( карьер Дора) недалеко от города Кауденбит в Файфе . Вуд нашел эти окаменелости в слое алевролита того же возраста, что и железняк в Лоанхеде. [ 3 ]

Американский палеонтолог Дональд Бэрд назвал второй вид Spathicephalus , S. pereger , из Новой Шотландии в 1962 году. [ 2 ] Бэрд назвал S. pereger на основании отпечатка правой половины черепной таблицы, которую коллекционеры из Гарвардского музея сравнительной зоологии нашли на пляже между Пойнт-Эдвард и бухтой Китинг на острове Кейп-Бретон . Отпечаток сохранился в алевролитах формации Пойнт-Эдвард , которая датируется поздним верхним карбоном (что соответствует раннему намюрскому периоду в Европе), а это означает, что S. pereger предшествует S. mirus на несколько миллионов лет. [ 4 ]
Описание
[ редактировать ]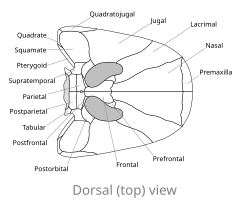



В отличие от большинства других ранних четвероногих с округлой или заостренной мордой, Spathicephalus mirus имеет уплощенный, почти идеально квадратной формы череп до 22 сантиметров (8,7 дюйма) в ширину и длину. [ 2 ] Квадратная форма обусловлена, прежде всего, расширением парных носовых костей вдоль средней линии морды. Кости предчелюстной кости на кончике рыла образуют весь передний край квадрата, а кости верхней челюсти образуют боковые края. Верхняя челюсть спатицефала необычна тем, что она тонкая (не более 1 сантиметра (0,39 дюйма) в толщину) по всей длине. Орбиты или глазницы большие, обращены прямо вверх и расположены близко друг к другу в задней части черепа. В отличие от примерно круглых орбит большинства четвероногих, орбиты спатицефалов имеют почковидную форму, поскольку они срослись с другой парой отверстий, называемых анторбитальными отверстиями (у других ранних четвероногих анторбитальные отверстия лежат перед орбитами и отделены от них). ). Черепная пластинка за орбитами очень мала по сравнению с таковыми у других четвероногих каменноугольного периода, но, как и у других четвероногих, в ее центре имеется небольшое отверстие, называемое шишковидным отверстием . Маленькая таблица «рога» отходят латерально от задней части черепной пластины, образуя височные вырезки . Поверхность крыши черепа покрыта выпуклостями и гребнями, палеонтологи Э. Х. Бомонт и Т. Р. Смитсон описывают это состояние как «пустулезный орнамент». [ 2 ]
Другой отличительной чертой S. mirus является зубной ряд; Его челюсти покрыты сотнями маленьких, долотообразных, близко расположенных зубов. Каждый из этих маргинальных зубов имеет диаметр около 3 миллиметров (0,12 дюйма) в поперечном сечении и образует непрерывный ряд вдоль верхней и нижней челюстей. В то время как на верхней челюсти они направлены прямо вниз, на нижней челюсти маргинальные зубы слегка наклонены внутрь (мезиально). Зубы на костях сошника образуют на небе второй параллельный ряд, не такой обширный, как краевой зубной ряд. Эти зубы диаметром около 1 миллиметра (0,10 см) меньше маргинальных зубов. Остальная часть неба имеет тонкий пустулезный орнамент более мелкого масштаба, чем на крыше черепа. Это уникальное состояние среди ранних четвероногих, многие из которых имеют более обширный орнамент на небе, включая костные зубцы, дополнительные зубные ряды и небные клыки. Зубной ряд спатицефала настолько необычен, что легко можно идентифицировать отдельные фрагменты челюстей с находящимися в них зубами. [ 2 ]
Хотя вид S. pereger известен только по частичной крыше черепа, палеонтолог Дональд Бэрд с уверенностью отнес его к Spathicephalus , поскольку формы его костей близко соответствовали соответствующим костям S. mirus . Одним из немногих отличий, отличающих S. pereger от S. mirus, является сетчатый или «сотовый» орнамент на крыше черепа, характеризующийся ямками и бороздками. Пустулезный орнамент, наблюдаемый у S. mirus, на самом деле редко встречается среди ранних четвероногих ( плагиозаврид темноспондилы и очень ранние четвероногие Ichthyostega - одни из немногих, у кого они есть), тогда как сетчатый орнамент S. pereger более типичен. [ 2 ]
Эволюционные отношения
[ редактировать ]Спатицефал — один из первых представителей группы четвероногих, в которую входят все позвоночные с четырьмя конечностями. Согласно самым последним исследованиям филогении ранних четвероногих , он принадлежит к кладе или эволюционной группе внутри четвероногих, называемой Baphetoidea , но находится за пределами подгруппы бафетоидов, называемой Baphetidae , которые образуют «ядро» Baphetoidea. Самые последние исследования эволюционных взаимоотношений четвероногих показывают, что бафетоиды относятся к четвероногим стволовой группы , а это означает, что они ответвились от других четвероногих до появления самого недавнего общего предка современных четвероногих ( амфибий , рептилий , птиц и млекопитающих ). Палеонтологи признали, что спатицефал был близким родственником бафетид с тех пор, как Д. М. С. Уотсон впервые описал его в 1929 году. С бафетидами спатицефала связывают две основные особенности : предглазничные отверстия, сросшиеся с глазницами, и закрытое небо, образованное в основном парой широких крыловидных костей. . Уотсон поместил Spathicephalus в Loxommatidae, семейство, которое он назвал двенадцатью годами ранее, включив в него так называемых бафетид. Хотя название Baphetidae существует с 1865 года и имеет приоритет над Loxommatidae, палеонтологи на протяжении большей части двадцатого века называли этих четвероногих локсоммидами. В обзоре ранних четвероногих 1947 года палеонтолог Альфред Ромер назвал спатицефала «гротескным типом локсоммид», считая его необычный череп свидетельством примитивного положения внутри группы. [ 5 ]
Спатицефалы не были включены в современный филогенетический анализ взаимоотношений ранних четвероногих до 2000-х годов. В 2009 году палеонтологи Анджела Милнер , Эндрю Милнер и Стиг Уолш включили анатомические характеристики S. mirus в анализ взаимоотношений бафетоидов. Анализ поместил Spathicephalus сразу за Baphetidae как сестринский таксон группы, и этот результат они использовали для обоснования его помещения в отдельное семейство Spathicephalidae. Анализ также показал, что Eucritta melanolimnetes , более древний вид из визейской стадии Шотландии, является самым базальным представителем Baphetoidea. Ниже представлена кладограмма , показывающая результаты их анализа: [ 6 ]
Возможные близкие родственники
[ редактировать ]Загадочный каменноугольный вид четвероногих Doragnathus woodi может быть связан со Spathicephalus . опубликованных в 1993 и 1994 годах, российский палеонтолог О. А. Лебедев предположил, что Doragnathus был младшим синонимом Spathicephalus В двух статьях , . [ 7 ] [ 8 ] Doragnathus woodi , названный Смитсоном в 1980 году из карьера Дора в Файфе, Шотландия (того самого рудника, где палеонтолог Стэнли П. Вуд обнаружил останки S. mirus ), известен только по фрагментам верхней и нижней челюсти. [ 9 ] Сами челюсти очень напоминают челюсти других четвероногих каменноугольного периода, но многочисленные очень маленькие, плотно расположенные, заостренные и изогнутые внутрь ( лингвально ) краевые зубы, которые они содержат, уникальны. Учитывая, что Doragnathus и Spathicephalus были единственными известными в то время четвероногими каменноугольными животными с очень маленькими и плотно расположенными краевыми зубами, Лебедев утверждал, что эти два вида представляют один и тот же таксон. Поскольку материал Doragnathus был меньше, чем любой экземпляр S. mirus , известный в то время, он также предположил, что Doragnathus представляет собой юную форму Spathicephalus и что изогнутые заостренные зубы первого трансформировались в прямые, долотообразные зубы Spathicephalus. последнее во время разработки. Однако эта гипотеза была опровергнута Бомонтом и Смитсоном, которые в 1998 году сообщили о челюсти S. mirus , которая была такой же маленькой, как и материал Doragnathus , но имела прямые зубы в форме долота. Бомонт и Смитсон также указали, что Doragnathus отличается от Spathicephalus. в наличии дополнительного зубного ряда вдоль парасимфезиальной пластинки и венечных отростков (серии костей, образующих внутренние части нижней челюсти ранних четвероногих, утраченных позднее в эволюции четвероногих). [ 2 ]
Другое стеблевое четвероногие, Sigournea multidentata из раннего карбона США, также может быть связано со Spathicephalus . Названный в 2006 году по визейского возраста месторождению трещин в Айове, Сигурнея немного старше Спатицефала . Он похож как на спатицефала , так и на дорагната, имея множество маленьких, плотно расположенных зубов. Sigournea отличается от Spathicephalus и напоминает Doragnathus наличием заостренных, а не долотообразных краевых зубов и второго ряда зубов в нижних челюстях, а от обоих таксонов отличается наличием на внутренней поверхности нижней челюсти отверстия, называемого экзомекелиевым окном . Однако, учитывая, что сигурнея известна только по одной нижней челюсти, ее родство с другими четвероногими остается неопределенным. [ 10 ]
Милнер и др. (2009) предположили, что Doragnathus woodi и Sigournea multidentata могут принадлежать к Spathicephalidae, хотя они не включили эти два вида в свой филогенетический анализ. [ 6 ] Более ранний филогенетический анализ, опубликованный Марчелло Рутой и Джоном Р. Болтом в 2008 году, включал Doragnathus woodi , Sigournea multidentata и Spathicephalus mirus , но не обнаружил, что все три вида группируются вместе. Вместо этого они нашли хорошую поддержку для Sigournea группировки с Occidens portlocki из самого раннего каменноугольного периода ( турне ) Ирландии и Doragnathus группировки с Greererpeton burkemorani из визеанской эпохи Западной Вирджинии в Соединенных Штатах. [ 11 ]
Пищевое поведение
[ редактировать ]Причудливая морфология черепа спатицефала предполагает, что его пищевые привычки сильно отличались от привычек других четвероногих каменноугольного периода. Большинство стеблевых четвероногих, включая бафетид, скорее всего, были рыбоядными (рыбоядными) из-за их больших зубов и глубоких черепов, которые служили точками крепления для сильных и быстродействующих челюстных мышц. Маленькие долотообразные зубы спатицефала не подходили для ловли рыбы. Более того, уплощенная форма его черепа означает, что депрессор нижней челюсти , мышцы, которые прикрепляются к задней части черепа и отвечают за открытие нижней челюсти, не имел бы много места для закрепления и, следовательно, имел бы плохое механическое преимущество . [ 2 ]
Другая группа плоскоголовых водных четвероногих, называемая плагиозавридами, темноспондилами, хотя и имеет лишь отдаленное родство, возможно, напоминала спатицефалов по своему пищевому поведению . Считается, что плагиозавриды, жившие в триасовый период , были донными рыбоядными животными, которые компенсировали свою слабую депрессорную нижнюю челюсть мощными приводящими мышцами , которые быстро закрывали бы нижнюю челюсть. Возможно, они отдыхали на дне озер и рек с открытой пастью и защелкивали ее, чтобы поймать рыбу. [ 12 ] В 1947 году палеонтолог Альфред Ромер предположил, что спатицефал был донным рыбоядным животным, но в отличие от плагиозавридов у него были слабые приводящие мышцы. [ 5 ] У спатицефалов , плагиозавридов и многих других ранних четвероногих пара отверстий в задней части неба, называемых подвисочными ямками, выполняет функцию якоря для приводящих мышц. Эти ямки у плагиозавридов большие, поэтому считается, что они имели мощные аддукторы. Однако у спатицефала подвисочные ямки меньшего размера, а это означает, что у него должны были быть слабые аддукторы и депрессоры нижней челюсти. В 1998 году палеонтологи Э. Х. Бомонт и Т. Р. Смитсон выдвинули гипотезу, что спатицефалы питались водными беспозвоночными, сочетая всасывание и фильтрацию. Чтобы питаться, он должен был лежать на дне озера или реки, слегка приоткрыв челюсти и подъязычным аппаратом закрывая горло . Когда группа мелких беспозвоночных приближалась, она опускала подъязычную мышцу, открывая горло и вызывая быстрое снижение давления внутри рта. Затем вода текла в рот, унося с собой добычу. Даже имея слабые приводящие мышцы, спатицефал мог сомкнуть челюсти до того, как добыча ускользнет, а затем снова поднять подъязычную мышцу, чтобы вытолкнуть воду обратно, одновременно фильтруя беспозвоночных рядами маленьких зубов. [ 2 ]
Ссылки
[ редактировать ]- ^ Тимоти Р. Смитсон; Майкл А.Э. Браун; Сара Дж. Дэвис; Джон Э.А. Маршалл; Дэвид Миллуорд; Стиг А. Уолш; Дженнифер А. Клак (2017). «Новый Миссисипский четвероногий из Файфа, Шотландия, и его экологический контекст» . Статьи по палеонтологии . 3 (4): 547–557. дои : 10.1002/spp2.1086 . hdl : 2381/40472 .
- ^ Jump up to: а б с д и ж г час я Бомонт, Э.Г.; Смитсон, Т.Р. (1998). «Морфология черепа и взаимоотношения аберрантной каменноугольной амфибии Spathicephalus mirus Watson» . Зоологический журнал Линнеевского общества . 122 (1–2): 187–209. дои : 10.1111/j.1096-3642.1998.tb02529.x .
- ^ Эндрюс, С.М.; Браун, MAE; Панчен, Алабама; Вуд, СП (1977). «Открытие амфибий в намюре (верхний карбон) Файфа». Природа . 265 (5594): 529. дои : 10.1038/265529a0 . S2CID 4186803 .
- ^ Бэрд, Д. (1962). «Рахитомная амфибия Spathicephalus из Миссисипи Новой Шотландии» . Бревиора . 157 : 1–9.
- ^ Jump up to: а б Ромер, А.С. (1947). «Обзор Лабиринтодонтии» . Вестник Музея сравнительной зоологии . 99 (1): 1–368.
- ^ Jump up to: а б Милнер, AC; Милнер, Арканзас; Уолш, Ю.А. (2009). «Новый экземпляр Baphetes из Ныржан, Чехия, и внутренние родственные связи Baphetidae» . Акта Зоология . 90 : 318–334. дои : 10.1111/j.1463-6395.2008.00340.x .
- ^ Лебедев О.А.; Клак, Дж. А. (1993). «Четвероногие верхнего девона из Андреевки, Тульская область, Россия» (PDF) . Палеонтология . 36 (3): 721–734.
- ^ Альберг, ЧП; Лукшевичс, Э.; Лебедев О. (1994). «Первые находки четвероногих из девона (верхнего фамена) Латвии». Философские труды Королевского общества B: Биологические науки . 343 (1305): 303. doi : 10.1098/rstb.1994.0027 .
- ^ Смитсон, TR; Клак, Дж. А. (2013). «Аппендикулярные элементы скелета четвероногих из раннего карбона Шотландии». Comptes Рендус Палевол . 12 (7–8): 405. doi : 10.1016/j.crpv.2013.06.006 .
- ^ Болт, младший; Ломбард, RE (2006). « Sigournea multidentata , новое стволовое четвероногие из Верхней Миссисипи штата Айова, США». Журнал палеонтологии . 80 (4): 717. doi : 10.1666/0022-3360(2006)80[717:SMANST]2.0.CO;2 . S2CID 130082060 .
- ^ Рута, М.; Болт, младший (2008). «Брахиопоид Hadrokkosaurus bradyi из раннего среднего триаса Аризоны и филогенетический анализ признаков нижней челюсти у темноспондильных амфибий» . Acta Palaeontologica Polonica . 53 (4): 579. doi : 10.4202/app.2008.0403 .
- ^ Дамиани, Р.; Шох, Р.Р.; Хеллрунг, Х.; Вернебург, Р.; Гастоу, СП (2009). «Плагиозаврид темноспондил Plagiosuchus pustuliferus (Amphibia: Temnospondyli) из среднего триаса Германии: анатомия и функциональная морфология черепа» . Зоологический журнал Линнеевского общества . 155 (2): 348. doi : 10.1111/j.1096-3642.2008.00444.x .





