Ботриолепис
| Ботриолепис Временной диапазон: поздний девон.
~ | |
|---|---|

| |
| Ботриолепис канадский слепой | |

| |
| Восстановление жизни Bothriolepis canadensis после Бешара и др. (2014) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | † Плакодермии |
| Заказ: | † Антиарх |
| Семья: | † Ботриолепидиды |
| Род: | † Ботриолепис Эйхвальд , 1840 г. |
| Разновидность | |
|
| |
Ботриолепис (от греческого : βόθρος bóthros , «траншея» и греческого : λεπίς lepis, разнообразный род антиарховых эры плакодерм , живших в период от среднего до позднего девона палеозойской «чешуя») — широко распространенный, обильный и . Исторически Ботриолепис обитал во множестве палеосред, разбросанных по каждому палеоконтиненту , включая прибрежные морские и пресноводные условия. [ 1 ] Большинство видов Bothriolepis были охарактеризованы как относительно небольшие донные пресноводные детритофаги (организмы, которые получают питательные вещества, потребляя разлагающийся растительный / животный материал), в среднем около 30 сантиметров (12 дюймов) в длину. [ 2 ] Однако самый крупный вид, B. rex , имел предполагаемую длину тела 170 сантиметров (67 дюймов). Хотя он обширен и насчитывает более 60 видов, обитающих по всему миру, [ 3 ] сравнительно Bothriolepis не является чем-то необычно более разнообразным, чем большинство современных донных видов, обитающих сегодня. [ 4 ]
Классификация
[ редактировать ]Bothriolepis — род плакодерм, относящийся к отряду Antiarchi . Самые ранние антиарховые плакодермы впервые появились в силурийском периоде палеозойской эры и к девонскому периоду могли быть обнаружены на каждом палеоконтиненте. [ 5 ] Самые ранние представители Bothriolepis появляются в среднем девоне.
Антиархи, как и другие плакодермы, морфологически разнообразны и характеризуются костными пластинками, покрывающими их голову и переднюю часть туловища. [ 5 ] Ранние онтогенетические стадии плакодерм имели более тонкие костные пластинки как внутри головы, так и внутри туловищного щита, что позволяло легко различать ранние онтогенетические стадии плакодерм в летописи окаменелостей и таксоны, которые обладали полностью развитыми костными пластинками, но были небольшими по характеристикам. [ 5 ] Костные пластинки плакодермы обычно состояли из трех слоев: компактного базального пластинчатого костного слоя, среднего губчатого костного слоя и поверхностного слоя; [ 5 ] Ботриолепис можно классифицировать как плакодерму, поскольку он обладает этими слоями.
Плакодермы вымерли к концу девона. [ 5 ] Placodermi — парафилетическая группа клады Gnathostomata , включающая всех челюстных позвоночных. [ 5 ] Неясно, когда именно появились челюстноротые, но скудные ранние окаменелости указывают на то, что это было где-то в раннепалеозойскую эру. [ 6 ] Последний вид Bothriolepis вымер вместе с остальными Placodermi в конце девона.
Общая анатомия
[ редактировать ]
Голова
[ редактировать ]В голове ботриолеписа есть два отверстия : замочная скважина по средней линии на верхней стороне для глаз и ноздрей и отверстие для рта на нижней стороне возле переднего конца головы. Открытие сохранившихся структур, которые выглядят как носовые капсулы, подтверждает мнение о том, что внешние носовые отверстия располагались на дорсальной стороне головы возле глаз. [ 7 ] Кроме того, положение рта на вентральной стороне черепа соответствует типичной горизонтальной ориентации в состоянии покоя Bothriolepis . На черепе у него была особенность: отдельная перегородка кости ниже отверстия для глаз и ноздрей, окружающая носовые капсулы, называемая предглазничной выемкой.
Челюсть
[ редактировать ]Новый образец из формации Гого в бассейне Каннинг в Западной Австралии предоставил доказательства морфологических особенностей висцеральных элементов челюсти Bothriolepis . Используя образец, становится очевидным, что ментальная пластинка (кожная кость, образующая верхнюю часть челюсти) антиархов гомологична суборбитальной пластинке, обнаруженной у других плакодерм. Нижняя челюсть состоит из дифференцированной пластинчатой и кусающей частей. Рядом с нижнечелюстным суставом расположены прелатеральная и инфрапрелатеральная пластинки, которые обе являются костями, несущими каналы. У небноквадрата отсутствует высокий глазничный отросток, и он прикреплялся только к вентральной части подбородочной пластинки, что доказывает, что решетчатая область черепной коробки (область черепа, разделяющая мозг и полость носа) на самом деле была глубже, чем первоначально предполагалось. [ 8 ] Помимо перечисленного выше образца из формации Гого, обнаружено еще несколько экземпляров с ротовым аппаратом, удерживаемым в естественном положении перепонкой, закрывающей ротовую область и прикрепляющейся к боковому и переднему краям головы. [ 9 ] У ботриолеписа есть челюсть, две половинки которой разделены и у взрослой особи функционально независимы. [ 9 ]

Ствол
[ редактировать ]У ботриолеписа был тонкий ствол, вероятно, покрытый мягкой кожей без чешуек и пятен. В состоянии покоя, по-видимому, в основном стабильной была ориентация дорсальной поверхностью вверх, о чем свидетельствует плоская поверхность на вентральной стороне. [ 1 ] Очертания туловища позволяют предположить, что там могла присутствовать хорда , окруженная перепончатой оболочкой. [ 9 ] однако прямых доказательств этого нет, поскольку хорда состоит из мягких тканей, которые обычно не сохраняются в летописи окаменелостей. Подобно другим антиархам , грудной щит Ботриолеписа был прикреплен к его тяжелобронированной голове. Его коробчатый корпус был заключен в броневые листы, обеспечивающие защиту от хищников. К вентральной поверхности туловища прикреплена большая тонкая круглая пластина, отмеченная глубокими линиями и поверхностными гребнями. Эта пластинка лежит чуть ниже отверстия клоаки . [ 9 ]
Дермальный скелет
[ редактировать ]Скелет дермы состоит из трех слоев: поверхностного пластинчатого слоя, губчатой губки и компактного базального пластинчатого слоя. Даже в раннем онтогенезе эти слои проявляются у экземпляров Bothriolepis canadensis . Компактные слои развиваются первыми. [ 10 ] Предполагается, что поверхностный слой имеет зубчики, которые могли состоять из клеточной кости. [ 11 ]
Плавники и хвост
[ редактировать ]У Ботриолеписа была пара длинных шипообразных грудных плавников , сочлененных у основания и расположенных чуть больше половины длины. Эти шипообразные плавники, вероятно, использовались для того, чтобы поднять тело со дна; его тяжелая броня заставила бы его быстро затонуть, как только он потерял бы инерцию движения вперед. [ 2 ] [ 12 ] Возможно, он также использовал свои грудные плавники, чтобы выбрасывать на себя осадок (ил, песок или что-то еще). Первоначально считалось, что помимо грудных плавников у него есть два спинных плавника, но в 1996 году было опровергнуто существование низкого удлиненного переднего спинного плавника, и теперь считается, что у него есть только высокий закругленный задний спинной плавник. [ 13 ] Хвостовой хвост был удлиненным и заканчивался узкой полосой, но, к сожалению, редко сохраняется в окаменелостях. [ 9 ] У ботриолеписа отсутствовали брюшные плавники. Ранние антиархи, такие как Parayunnanolepis , имели брюшные плавники, что предполагает вторичную потерю брюшных плавников у Bothriolepis . [ 14 ]
Мягкая анатомия
[ редактировать ]Структуры, состоящие из мягких тканей, обычно не сохраняются в окаменелостях, поскольку они легко разрушаются и разлагаются гораздо быстрее, чем твердые ткани, а это означает, что в летописи окаменелостей часто не хватает информации о внутренней анатомии ископаемых видов. Однако иногда может произойти сохранение структур мягких тканей, если отложения заполняют внутренние структуры организма после его смерти. Статья Роберта Денисона под названием «Мягкая анатомия ботриолеписа » исследует формы и органы ботриолеписа . [ 7 ] Эти внутренние структуры сохранились, когда различные типы отложений, окружающих внешнюю часть животного, заполнили внутренние панцири (таким образом могли сохраниться только органы, сообщающиеся с внешней средой). В разных разрезах Ботриолеписа были идентифицированы три различных типа отложений : первый - бледный зеленовато-серый песчаник средней текстуры, в основном состоящий из кальцита; второй похожий, но более тонкий осадок, сохранивший многие формы органов; и третий отчетливый мелкозернистый алевролит, состоящий из кварца, слюды и других минералов, но без кальцита. [ 7 ] Эти отложения помогли сохранить следующие внутренние элементы:
Пищеварительная система
[ редактировать ]
В целом пищеварительную систему ботриолеписа , включающую органы, участвующие в приеме пищи, пищеварении и удалении отходов, можно охарактеризовать как простую и прямую, в отличие от системы человека. Начинается на переднем конце организма небольшой ротовой полостью, расположенной над задним участком пластинок верхней челюсти. Кзади от рта пищеварительная система простирается в более широкую и уплощенную в дорсо-вентральном направлении область, называемую глоткой , из которой возникают жабры и легкие. Пищевод , который также характеризуется как уплощенная в дорсо-вентральном направлении трубка, простирается ото рта в желудок и приводит к уплощенной эллипсоидной структуре. Эта структура может быть гомологична переднему концу кишечника, встречающемуся у других рыб. [ 7 ] Плоскостность этих структур могла быть преувеличена, когда ископаемые образцы подверглись тектонической деформации в течение геологического времени. Кишка начинается узко на переднем конце, расширяется в поперечном направлении, а затем снова сужается кзади по направлению к цилиндрической прямой кишке, которая заканчивается непосредственно в пределах заднего конца панциря туловища. Пищеварительная система примитивна по своей природе и не имеет расширенного отдела желудка, но специализирована самостоятельно приобретенным сложным спиральным клапаном, сравнимым с таковым у пластиножаберных и многих костных рыб и сходным с таковым у некоторых акул. Единственная складка ткани, свернутая вокруг своей оси, образует этот специализированный спиральный клапан. [ 7 ]
Жабры
[ редактировать ]Предполагается, что жабры Ботриолеписа относятся к примитивному типу, однако их строение до сих пор недостаточно изучено. С латеральной стороны они окружены оперкулярной складкой и находятся в пространстве под латеральной частью головного щита, простираясь медиально под нейрокраниум . По сравнению с жабрами рыб нормальной формы, жаберная область у Bothriolepis расположена более дорсально, спереди более скучена и в целом относительно короткая и широкая. [ 7 ]
Парные вентральные мешки
[ редактировать ]Кзади от туловищного панциря отходят парные вентральные мешки, доходящие до переднего конца спиральной кишки. Мешки, по-видимому, берут начало в глотке в виде одной срединной трубки, которая затем расширяется кзади и в конечном итоге разделяется на два мешка, которые могут быть гомологичны легким некоторым двуногим и четвероногим . [ 7 ] Была выдвинута гипотеза, что эти легкие в сочетании с суставчатыми руками и жестким поддерживающим скелетом позволили Ботриолепису путешествовать по суше. Кроме того, как Роберт Денисон [ 7 ] Согласно утверждениям, поскольку нет никаких доказательств связи между внешней ноздрей и ртом, Ботриолепис, вероятно, дышал так же, как современные двоякодышащие рыбы, то есть располагая рот над поверхностью воды и заглатывая воздух.
Несмотря на оригинальную интерпретацию, представленную Денисоном в 1941 году, не все палеонтологи согласны с тем, что плакодермы, такие как Ботриолепис, действительно обладали легкими. Например, в своей статье «Легкие» у плакодерм, стойкого палеобиологического мифа, связанного с предвзятыми интерпретациями окружающей среды , Д. Гуже предполагает, что, хотя следы некоторых органов пищеварения могут быть очевидны в осадочных структурах, нет никаких доказательств, подтверждающих наличие легких. в образцах из формации Эскуминак в Канаде, на которых было основано первоначальное утверждение. Он отмечает, что распространение ботриолеписа по всему миру ограничено строго морской средой, и поэтому считает, что наличие легких у ботриолеписа сомнительно. Дальнейшее исследование окаменелостей, вероятно, необходимо, чтобы прийти к выводу о наличии легких у Ботриолеписа . [ 15 ]
Кормление
[ редактировать ]Считается, что ботриолепис , как и все другие антиархи, питался, напрямую заглатывая грязь и другие мягкие отложения, чтобы переварить детрит , мелкие микроорганизмы, водоросли и другие формы органических веществ в проглоченных отложениях. Кроме того, расположение рта на брюшной стороне головы также позволяет предположить, что Ботриолепис, вероятно, питался снизу. Считается, что регулярное присутствие «углеродистого материала в пищеварительном тракте» указывает на то, что большая часть его рациона состояла из растительного материала. [ 7 ]
Распределение
[ редактировать ]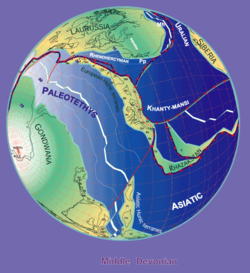
Стампфли и Борель, 2000 г.
Окаменелости ботриолеписа встречаются в слоях среднего и позднего девона (от 387 до 360 миллионов лет назад). [ 12 ] Поскольку окаменелости обнаружены в пресноводных отложениях, предполагается, что ботриолепис провел большую часть своей жизни в пресноводных реках и озерах, но, вероятно, мог проникать и в соленую воду , поскольку его ареал, по-видимому, соответствовал континентальным береговым линиям девона. Большие группы экземпляров Bothriolepis были обнаружены в Азии, Европе, Австралии ( формация Гого и песчаник Мандагери ), [ 9 ] [ 16 ] Африка ( месторождение фермы Ватерлоо ) [ 17 ] Пенсильвания (формация Катскилл), [ 1 ] Квебек ( формация Эскуминак ), [ 1 ] Вирджиния (Чемунг), [ 18 ] Колорадо, [ 18 ] Формация Куш ( Бояка , Колумбия), [ 19 ] и по всему миру.
Сайт формации Кэтскилл
[ редактировать ]Формация Катскилл (верхний девон, фаменский ярус), расположенная в округе Тиога , штат Пенсильвания , является местом обитания крупной выборки мелких особей Bothriolepis . Образец был собран из серии каменных плит, состоявших из частичных или полных сочлененных внешних скелетов. Было обнаружено более двухсот особей, расположенных близко друг к другу, практически без перекрытия. много информации о характеристиках молодых особей Ботриолеписа Из этого образца можно получить . Морфометрическое исследование, проведенное Джейсоном Даунсом и соавторами, выявило определенные характеристики, указывающие на молодость Bothriolepis , в том числе умеренно большую голову и умеренно большие глазничные отверстия — обе характеристики также были признаны Эриком Стенсио в 1948 году у самых маленьких B. canadensis особей . . [ 1 ] Несколько других особенностей, которые Стенсио отметил как характерные для молодых особей, также можно увидеть в выборке из Катскилла. Эти особенности включают «нежные кожные кости с орнаментом, состоящим из непрерывных анастомозирующих гребней, а не бугорков, дорсальный щиток туловища, более узкий, чем длинный, с непрерывным и выраженным дорсальным срединным гребнем, а также предсрединную пластинку, которая шире, чем длинна». [ 1 ] B. nitida и B.minor также описаны с этого сайта. [ 20 ]
Разновидность
[ редактировать ]Палеонтология позвоночных во многом зависит от способности различать разные виды единым образом как внутри конкретного рода, так и среди всех организмов. Род Bothriolepis не является исключением из этого принципа. Ниже перечислены некоторые известные виды Bothriolepis ; Всего было названо более шестидесяти видов, и вполне вероятно, что значительная часть из них допустима из-за космополитической природы Bothriolepis . [ 3 ]
Ботриолепис канадский
[ редактировать ]
Bothriolepis canadensis — таксон, который часто служит модельным организмом для отряда Antiarchi из-за огромной выборки полных, неповрежденных экземпляров, найденных в формации Эскуминак в Квебеке , Канада . [ 1 ] Из-за огромного размера выборки этот вид часто используется для сравнения данных о росте недавно приобретенных экземпляров Bothriolepis , в том числе обнаруженных в упомянутой выше формации Катскилл. Это сравнение позволяет исследователям определить, представляют ли недавно найденные образцы молодые особи или новый вид «Ботриолепис».
B. canadensis был впервые описан в 1880 году Дж. Ф. Уайтавсом с использованием ограниченного количества изуродованных образцов. Следующим, кто предложил реконструкцию вида, был У. Паттен, опубликовавший свои открытия в 1904 году после открытия нескольких экземпляров, хорошо сохранившихся в 3-D. В 1948 году Э. Стенсио опубликовал подробное описание анатомии B. canadensis , используя обилие материала, которое в конечном итоге стало наиболее широко принятым описанием этого вида. После публикации Стенсио многие другие предоставили реконструированные модели B. canadensis с измененными аспектами анатомии, включая модифицированный одиночный спинной плавник Везины, а в последнее время - реконструкции Арсено и др . на основе образцов с небольшими тафономическими искажениями. В настоящее время модель Арсено и др . считается наиболее точным, хотя до сих пор ведется много споров о различных аспектах внешней анатомии этого вида. Несмотря на неопределенность, B. canadensis по-прежнему считается одним из самых известных видов. [ 13 ]
Внешний скелет Bothriolepis canadensis представлен клеточной дермальной костной тканью и характеризуется отчетливой горизонтальной зональностью или стратификацией. [ 10 ] Модельная рыба имеет среднюю общую длину 43,67 сантиметра (17,19 дюйма) и среднюю длину кожного панциря 15,53 сантиметра (6,11 дюйма), что составляет 35,6% от расчетной общей длины. [ 13 ] Как и многие антиархи, B. canadensis также имел узкие грудные плавники, гетероцеркальный хвостовой плавник (то есть хорда доходит до верхней доли хвостового хвоста) и большой спинной плавник, который, вероятно, не играл важной роли в движении, а вместо этого действовал скорее стабилизатор. [ 13 ]
Ботриолепис африканский
[ редактировать ]
Ботриолепис африканский [ 17 ] вид Bothriolepis , известный из самых высоких палеоширот и описанный из отложений, первоначально заложенных в пределах позднего девона Антарктического круга. Останки были извлечены исключительно из одного углеродистого сланца вблизи вершины позднего девона, фаменского периода , формации Витпоорт (группа Виттеберг), обнаженной на дороге, проходящей к югу от Маханда/ Грэмстаун в Южной Африке. Это место, лагерштетте фермы Ватерлоо, интерпретируется как представляющее собой задний барьер прибрежной лагуны с морским и речным влиянием. [ 21 ] Гесс заметил, что ботриолепис был менее многочисленным на участке фермы Ватерлоо, чем в большинстве мест обитания ботриолеписа , хотя здесь представлена полная онтогенетическая серия. Длина брони головы и туловища колебалась в пределах 20–300 миллиметров (0,79–11,81 дюйма), что, исходя из пропорций двух самых маленьких особей (у которых сохранились отпечатки хвоста), соответствует полной длине тела, варьирующейся от 52 до 780 миллиметров ( 2,0–30,7 дюйма). [ 22 ] Согласно оригинальному описанию, Bothriolepis africana считался наиболее близким к Bothriolepis barretti. [ 23 ] позднего живе Антарктиды . из Сходство между ними было использовано, чтобы предположить происхождение Bothriolepis africana из среды Восточной Гондваны . [ 17 ]

Ботриолепис колораденсис
[ редактировать ]Впервые описанный Истманом в 1904 году, этот вид был обнаружен в современном Колорадо . Существует вероятность, что этот вид похож, если не идентичен, на B. nitida , однако, поскольку доступный материал о B. coloradensis фрагментирован, невозможно сравнить эти два вида с какой-либо степенью уверенности. [ 18 ]
Ботриолепис нида
[ редактировать ]
Этот вид, обнаруженный на территории современной Пенсильвании , был первоначально описан Дж. Лейди в 1856 году. Как упоминалось выше, существует много споров относительно различения B. nitida и B. Virginiensis , однако они основаны на доказательствах, представленных Weems (2004). , [ 18 ] Есть несколько отличительных черт, характерных для каждого вида. B. nitida имеет максимальную длину головного щитка 65 миллиметров (2,6 дюйма), узкую и неглубокую тройчатую предглазничную выемку, передне-срединно-дорсальную (AMD) пластинку, которая шире, чем ее длина, и вентральный грудной щиток, имеющий выпуклую форму. боковые границы. [ 18 ]
Ботриолепис рекс
[ редактировать ]Первоначально описано Downs et al. (2016), Bothriolepis rex происходит из формации Нордстранд-Пойнт на острове Элсмир, Канада. Длина тела B. rex оценивается в 1,7 метра (5,6 футов), и поэтому это самый крупный из известных видов Bothriolepis . Его броня особенно толстая и плотная, даже если принять во внимание его размер. Даунс и др. (2016) предполагают, что это могло не только защищать животное от крупных хищников, но и служить балластом, предотвращающим всплывание этого крупного донного обитателя на поверхность.
Ботриолепис виргинский
[ редактировать ]Первоначально описано Weems et al. в 1981 году этот вид, Bothriolepis Virginiensis , произошел из «Чемунга», недалеко от Винчестера, штат Вирджиния . Некоторые черты, обнаруженные у B. Virginiensis , можно также обнаружить у других видов Bothriolepis (особенно B. nitida ), включая бороздки задней косой головной сенсорной линии, которые сходятся относительно далеко вперед на выйной пластинке, относительно удлиненное глазничное окно и низкое переднее отверстие. срединно-дорсальный гребень. [ 18 ] Характеристики, которые отличают B. Virginiensis от других видов, включают, помимо прочего, сросшиеся швы на голове, сросшиеся элементы в дистальных сегментах грудных плавников взрослых особей и длинную премедиальную пластинку относительно длины головного щита. [ 18 ]
В настоящее время ведется много споров о том, виды B. Virginiensis и B. nitida можно ли отличить друг от друга. Томсон и Томас заявляют, что пять видов Bothriolepis из Соединенных Штатов ( B. nitida, B.minor, B. Virginiensis, B. darbiensis и B. coloradensis ) невозможно четко отличить друг от друга. [ 4 ] И наоборот, Уимс утверждает, что существует несколько черт, которые отличают эти виды друг от друга, в том числе несколько, перечисленных выше. [ 18 ]
Ботриолепис euungae
[ редактировать ]
Этот вид описан в 1998 году из песчаника Мандагери в Кановиндре , где он известен по большому количеству образцов плакодермы, собранных в одном месте. Ботриолепис — одна из наиболее распространенных рыб на участке Кановиндра наряду с Ремиголеписом , к 1998 году обнаружено более 1300 особей. От всех других видов этот вид отличается редуцированным передним отростком субмаргинального, отделенным от заднего отростка субмаргинального широкий открытый вырез. Длина брони головы и туловища составляла 77,6–190 миллиметров (3,06–7,48 дюйма). [ 16 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Даунс, JP; Крисвелл, Кентукки; Даешлер, Э.Б. (октябрь 2011 г.). «Массовая смертность молодых антиархов ( Botriolepis sp.) из формации Катскилл (верхний девон, фаменский ярус), округ Тайога, Пенсильвания». Труды Академии естественных наук Филадельфии . 161 (161): 191–203. дои : 10.1635/053.161.0111 . S2CID 129501544 .
- ^ Jump up to: а б Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 33. ISBN 978-1-84028-152-1 .
- ^ Jump up to: а б Янг, GC (2010). «Плакодермы (бронированные рыбы): доминирующие позвоночные девона». Ежегодный обзор наук о Земле и планетах . 38 : 523–550. Бибкод : 2010AREPS..38..523Y . doi : 10.1146/annurev-earth-040809-152507 . S2CID 86268051 .
- ^ Jump up to: а б Томсон, Канзас; Томас, Б. (август 2001 г.). «О статусе видов Bothriolepis (Placodermi, Antiarchi) в Северной Америке». Журнал палеонтологии позвоночных . 21 (4): 679–686. doi : 10.1671/0272-4634(2001)021[0679:otsoso]2.0.co;2 . S2CID 86104844 .
- ^ Jump up to: а б с д и ж Йохансон, Зерина; Тринайстик, Кейт (2014). «Ископаемые онтогенезы: вклад онтогенеза плакодермы в наше понимание эволюции ранних челюстноротых» . Палеонтология . 57 (3): 505–516. Бибкод : 2014Palgy..57..505J . дои : 10.1111/пала.12093 . S2CID 85673117 .
- ^ Бразо, М. (2009). «Мозговая коробка и челюсти девонского акантода и современного происхождения челюстноротых» (PDF) . Природа . 457 (7227): 305–308. Бибкод : 2009Natur.457..305B . дои : 10.1038/nature07436 . hdl : 10044/1/17971 . ПМИД 19148098 . S2CID 4321057 .
- ^ Jump up to: а б с д и ж г час я Денисон, Р.Х. (сентябрь 1941 г.). «Мягкая анатомия Ботриолеписа ». Журнал палеонтологии . 15 (5): 553–561.
- ^ Янг, GC (1984). «Реконструкция челюстей и черепной коробки девонской плакодермы Fish Bothriolepis ». Палеонтология . 27 (3): 635–661.
- ^ Jump up to: а б с д и ж Паттен, В. (июль 1904 г.). «Новые факты о ботриолеписе ». Биологический бюллетень . 7 (2): 113–124. дои : 10.2307/1535537 . JSTOR 1535537 .
- ^ Jump up to: а б Даунс, JP; Донохью, PCJ (2009). «Скелетная гистология Bothriolepis canadensis (Placodermi, Antiarchi) и эволюция скелета у происхождения челюстных позвоночных». Журнал морфологии . 270 (11): 1364–1380. дои : 10.1002/jmor.10765 . ПМИД 19533688 . S2CID 31585571 .
- ^ Джайлз, С. (2013). «Гистология дермальных скелетов «плакодермы»: значение для природы предков челюстноротых» . Журнал морфологии . 274 (6): 627–644. дои : 10.1002/Jmor.20119 . ПМК 5176033 . ПМИД 23378262 .
- ^ Jump up to: а б «Музей Эпохи Рыб – Окаменелости» . Музей возраста рыб, Новый Южный Уэльс, Австралия.
- ^ Jump up to: а б с д Бешар, И.; Арсено, Ф.; Клотье, Р.; Керр, Дж. (2014). «Девонская рыба Bothriolepis canadensis с помощью трехмерных цифровых изображений». Электронная палеонтология . 17 (1).
- ^ Шарест, Франция; Йохансон, Зерина; Клотье, Ришар (2018). «Потеря в процессе становления: отсутствие брюшных плавников и наличие педоморфных тазовых поясов у антиарховой плакодермы позднего девона (челюстной стебель-гнатостом)» . Письма по биологии . 14 (6): 20180199. doi : 10.1098/rsbl.2018.0199 . ISSN 1744-9561 . ПМК 6030608 . ПМИД 29899132 .
- ^ Гуже, Д. (2011). « Легкие» у плакодерм — стойкий палеобиологический миф, связанный с предвзятыми интерпретациями окружающей среды». Comptes Рендус Палевол . 10 (5–6): 323–329. дои : 10.1016/j.crpv.2011.03.008 .
- ^ Jump up to: а б Йохансон, Зерина (25 ноября 1998 г.). «Верхнедевонская рыба Bothriolepis (Placodermi: Antiarchi) из близ Кановиндры, Новый Южный Уэльс, Австралия» . Записи Австралийского музея . 50 (3): 315–348. дои : 10.3853/j.0067-1975.50.1998.1289 . ISSN 0067-1975 .
- ^ Jump up to: а б с ЛОНГ, Дж.А., АНДЕРСОН, М.Э., ГЕСС, Р.В. и ХИЛЛЕР, Н. (1997). Новые рыбы-плакодермы из позднего девона Южной Африки. Журнал палеонтологии позвоночных 17,253–268.
- ^ Jump up to: а б с д и ж г час Уимс, RE (март 2004 г.). « Botriolepis viginiensis , действительный вид рыб-плакодерм, отделимый от Bothriolepis nitida ». Журнал палеонтологии позвоночных . 24 (1): 245–250. Бибкод : 2004JVPal..24..245W . дои : 10.1671/20 . S2CID 85572685 .
- ^ Жанвье, Филипп; Вильярроэль А, Карлос (1998). «Девонские рыбы массива Флореста (Бояка, Колумбия). Таксономические, биостратиграфические, биогеографические и экологические аспекты» . Колумбийская геология . 23 :3–18 . Проверено 31 марта 2017 г.
- ^ Саллан, Лорен Коул; Коутс, Майкл И. (2010). «Вымирание в конце девона и узкое место в ранней эволюции современных челюстных позвоночных» . Труды Национальной академии наук . 107 (22): 10131–10135. Бибкод : 2010PNAS..10710131S . дои : 10.1073/pnas.0914000107 . ISSN 0027-8424 . ПМК 2890420 . ПМИД 20479258 .
- ^ Гесс, Роберт В.; Уитфилд, Алан К. (14 февраля 2020 г.). «Этуарная рыба и эволюция четвероногих: данные из устьевого озера Гондваны в позднем девоне (фамене) и южноафриканского голоценового эквивалента». Биологические обзоры. doi:10.1111/brv.12590. ПМИД 32059074
- ^ GESS, RW (2011). Модели фаменского биоразнообразия Гондваны в высоких широтах - данные из южноафриканской формации Витпоорт (Cape Supergroup, WittebergGroup). Кандидатская диссертация: Университет Витватерсранда, Йоханнебург.
- ^ ЯНГ, Г. (1984). Реконструкция челюстей и черепной коробки девонской плакодермальной рыбы Bothriolepis. Палеонтология 27, 635–661.
- ^ Геннатас, Лу-Андреа (28 марта 2022 г.). Геохронология и происхождение позднедевонского рыбного пласта кановиндры, Лахланского орогена (дипломная работа). Университет Маккуори. дои : 10.25949/19432913.v1 .
