Тероцефалия
| тероцефалы Временной диапазон: средняя пермь – средний триас Возможный таксон-потомок Cynodontia дожил до наших дней.
| |
|---|---|

| |
| Восстановление жизни двух представителей раннего тероцефалового рода Alopecognathus | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | Терапсида |
| Клэйд : | Эутериодонтия |
| Клэйд : | † Тероцефалия Метла , 1903 год. |
| Доплата | |
Тероцефалия — вымершая клада эутериодонтов - терапсидов (млекопитающих и их близких родственников) пермского и триасового периодов . Тероцефалы («звероголовые») названы в честь их больших черепов, которые, наряду со строением зубов, позволяют предположить, что они были хищниками . , не относящиеся к млекопитающим Как и другие синапсиды , тероцефалов когда-то описывали как « рептилий, подобных млекопитающим ». Тероцефалия — группа, наиболее тесно связанная с цинодонтами , давшими начало млекопитающим , и это родство подтверждается различными особенностями скелета. Действительно, было высказано предположение, что цинодонты могли произойти от тероцефалов и что признанные тероцефалы являются парафилетическими по отношению к цинодонтам.
Окаменелости тероцефалов многочисленны в Кару в Южной Африке , но они также были найдены в России , Китае , Танзании , Замбии и Антарктиде . Ранние окаменелости тероцефалов, обнаруженные в среднепермских отложениях Южной Африки, подтверждают гондванское происхождение этой группы, которая, по-видимому, быстро распространилась по Земле. Хотя почти каждая линия тероцефалов прекратила свое существование во время великого пермско-триасового вымирания , несколько представителей подгруппы под названием Eutherocephalia дожили до раннего триаса . Считается, что некоторые роды , принадлежащие к этой группе, обладают ядом , что делает их старейшими четвероногими, имеющими такие характеристики. Однако последние тероцефалы вымерли к началу среднего триаса , возможно, из-за изменения климата , а также конкуренции с цинодонтами и различными группами рептилий — в основном архозаврами и их близкими родственниками, включая архозавроморфов и архозавроформ .
Анатомия и физиология
[ редактировать ]
Подобно горгонопсиям и многим цинодонтам, большинство тероцефалов предположительно были хищниками . Ранние тероцефалы были во многих отношениях столь же примитивными, как и горгонопсии, но у них были некоторые продвинутые черты. Происходит расширение височного отверстия для прикрепления более широких приводящих мышц челюсти и редукция фаланг (костей пальцев рук и ног) до формулы фаланг млекопитающих. Наличие зарождающегося вторичного неба у продвинутых тероцефалов является еще одной особенностью, общей для млекопитающих. Открытие челюстных гребней у таких форм, как примитивные тероцефалы Glanosuchus , позволяет предположить, что по крайней мере некоторые тероцефалы могли быть теплокровными. [ 1 ]
К более поздним тероцефалам относились продвинутые Baurioidea , которые обладали некоторыми териодонтными характеристиками и имели высокую степень специализации. Например, у мелких бауриоидов и травоядных баурий отсутствовала окостеневшая заглазничная перемычка, отделяющая орбиту от височного отверстия, — состояние, типичное для примитивных млекопитающих. Эти и другие передовые особенности привели к давнему мнению, ныне отвергнутому, что иктидозавры и даже некоторые ранние млекопитающие произошли от бауриоидного тероцефалового ствола. Подобные характеристики млекопитающих, по-видимому, развивались параллельно среди ряда различных групп терапсидов, даже внутри Therocephalia. [ 1 ]

Для некоторых тероцефалов было предложено несколько более специализированных образов жизни. Многие мелкие формы, такие как иктидозухиды, интерпретировались как водные животные. Доказательства водного образа жизни включают склеротические кольца , которые, возможно, стабилизировали глаз под давлением воды, а также сильно развитые черепные суставы, которые могли поддерживать череп при поедании крупной рыбы и водных беспозвоночных. Один тероцефал, Nothogomphodon , имел большие саблевидные клыки и, возможно, питался крупными животными, включая других тероцефалов. Другие тероцефалы, такие как баурииды и наниктидопиды, имеют широкие зубы со множеством гребней, похожих на зубы млекопитающих, и, возможно, были травоядными . [ 2 ]
У многих мелких тероцефалов на морде есть небольшие ямки, которые, вероятно, поддерживают вибриссы (усы). В 1994 году российский палеонтолог Леонид Татаринов предположил, что эти ямки были частью системы электрорецепции водных тероцефалов. [ 3 ] Однако более вероятно, что эти ямки представляют собой увеличенные версии тех, которые, как считается, поддерживают усы или отверстия для кровеносных сосудов в мясистой губе. [ 2 ] Роды имеют некоторые структуры, позволяющие предположить, что у этих двух животных были органы для распространения яда. Euchambersia и Ichibengops , датируемые лопингом , особенно привлекают внимание палеонтологов, поскольку ископаемые черепа приписываемые им [ 4 ] [ 5 ]
Классификация
[ редактировать ]
Тероцефалы развились как одна из нескольких линий немлекопитающих терапсидов и имеют тесное родство с цинодонтами, которые включают млекопитающих и их предков. Большинство современных исследователей широко считают их родственной группой цинодонтов, объединенных в кладу Eutheriodontia . Однако некоторые исследователи предположили, что тероцефалы сами являются предками цинодонтов, что делает тероцефалов кладистически парафилетическими по сравнению с цинодонтами. Исторически сложилось так, что согласно этой гипотезе часто предполагается, что цинодонты произошли от тероцефалового семейства Whaitsiidae (или были наиболее близки к нему) , однако исследование 2024 года вместо этого обнаружило подтверждение сестринских отношений между цинодонтами и эвтероцефалиями. [ 6 ] Самые старые известные тероцефалы впервые появляются в летописи окаменелостей одновременно с другими основными группами терапсидов, включая горгонопсию , на которую они похожи во многих примитивных чертах. Например, многие ранние тероцефалы обладают длинными клыками, похожими на таковые у горгонопсов. Тероцефалы, однако, пережили горгонопсов, сохранившись до начала среднего триаса как мелкие плотоядные животные, похожие на ласку, и травоядные животные, похожие на цинодонтов. [ 7 ]
Хотя общее происхождение с цинодонтами (и, следовательно, с млекопитающими) объясняет многие сходства между этими группами, некоторые ученые считают, что другие сходства лучше объясняются конвергентной эволюцией , например, утратой заглазничной перемычки у некоторых форм, формулой фаланг млекопитающих. и некоторая форма вторичного неба у большинства таксонов. И тероцефалы, и цинодонты пережили пермско-триасовое массовое вымирание ; но, хотя тероцефалы вскоре вымерли, цинодонты подверглись быстрой диверсификации. У тероцефалов наблюдалось снижение скорости кладогенеза , а это означает, что после вымирания появилось несколько новых групп. Большинство триасовых линий тероцефалов зародилось в поздней перми и просуществовало лишь короткий период времени в триасе. [ 8 ] вымирает в позднеанизийском периоде . [ 9 ]
Таксономия
[ редактировать ]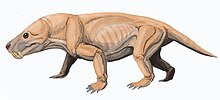
Тероцефалия была впервые названа и задумана Робертом Брумом в 1903 году как отряд , включающий в себя то, что он считал примитивными териодонтами, основанными в первую очередь на Scylacosaurus и Ictidosaurus . Однако его первоначальная концепция тероцефалии сильно отличалась от современной классификации, включая также различные роды горгонопсиев (включая горгонопсов ) и диноцефалов . С 1903 по 1907 год Брум добавил в эту группу больше родов тероцефалов, а также некоторых нетероцефалов, включая аномодонта Galechirus . Включение последнего подчеркнуло взгляд Брума на тероцефалов как на «примитивных» и предков других терапсидов, полагая, что аномодонты произошли от тероцефалоподобного предка, такого как Galechirus . Однако к 1908 году он считал включение его и некоторых других нетероцефалов в эту группу сомнительным. В 1913 году Брум восстановил Gorgonopsia в отличие от Therocephalia, но в течение многих десятилетий после этого он и другие исследователи все еще находились в замешательстве относительно того, какие роды к какой группе принадлежат. Ранг группы также варьировался от порядка, подотряда и инфраотряда в зависимости от предпочитаемой авторами систематики терапии. [ 10 ]
В то же время небольшие «продвинутые» тероцефалы, которые теперь классифицируются как Baurioidea, часто рассматривались как принадлежащие к отдельной подгруппе терапсидов, отличной от тероцефалов, Bauriamorpha . [ 11 ] Бауриаморфы классифицировались отдельно от тероцефалов на протяжении многих десятилетий, хотя часто предполагалось, что они произошли от тероцефалов параллельно с цинодонтами, каждый из которых обычно принадлежал к разным группам тероцефалов. [ 10 ] Включение бауриоидов в состав Therocephalia было твердо установлено только в 1980-х годах, а именно Кемпом (1982) и Хопсоном и Баргусеном (1986). [ 12 ] [ 13 ]
С тех пор, как группа была названа, были предложены различные подгруппы и клады тероцефалов, хотя их состав и номенклатура часто были крайне нестабильными, а некоторые ранее признанные клады тероцефалов оказывались искусственными или основанными на сомнительных таксонах. Это привело к тому, что некоторые распространенные имена в тероцефалической литературе, иногда используемые десятилетиями, были заменены менее известными именами, имеющими приоритет. Например, Scaloposauridae были основаны на окаменелостях с преимущественно ювенильными характеристиками и, вероятно, представлены неполовозрелыми экземплярами из других разрозненных семейств тероцефалов.
В другом примере название «Pristerognathidae» широко использовалось для группы базальных тероцефалов на протяжении большей части 20-го века, но с тех пор было признано, что название Scylacosauridae имеет прецедент для этой группы. Более того, сфера охвата «Pristerognathidae» была нестабильной и по-разному ограничивалась отдельной подгруппой ранних тероцефалов (наряду с другими, такими как Lycosuchidae, Alopecodontidae и Ictidosauridae), а также охватывала всю совокупность ранних тероцефалов. [ 10 ] Точно так же в литературе 20-го века для тероцефалов, соответствующих семейству Adkidnognathidae, использовались различные названия, включая Annatherapsididae, Euchambersiidae (самое старое доступное название) и Moschorhinidae, и их члены часто имели запутанное отношение к вайтсидам. Консенсус относительно названия и содержания Akidnognathidae был достигнут только в 21 веке, утверждая, что группа семейного уровня создана на основе самого старого подлежащего упоминанию рода и, таким образом, Akidnognathidae имеет прецедент для этой группы неваицоидных эвтероцефалов. [ 11 ]
С другой стороны, некоторые группы, которые раньше считались искусственными, оказались действительными. Аберрантное семейство тероцефалов Lycosuchidae, когда-то идентифицированное по наличию множественных функциональных клыкообразных зубов , было предложено представлять собой неестественную группу на основе исследования замены клыков у ранних тероцефалов, проведенного Ван ден Хевером в 1980 году. [ 14 ] Однако последующий анализ выявил дополнительные синапоморфии, подтверждающие монофилию этой группы (включая отсроченную замену клыкообразных), и Lycosuchidae в настоящее время считается действительной базальной кладой внутри Therocephalia. [ 15 ] Однако большинство родов, входящих в группу, с тех пор были объявлены сомнительными, и теперь в нее входят только Lycosuchus и Simorhinella . [ 16 ]

Вместо этого современная таксономия тероцефалов основана на филогенетическом анализе видов тероцефалов, который последовательно распознает две группы ранних тероцефалов (Lycosuchidae и Scylacosauridae), в то время как более производные тероцефалы образуют кладу Eutherocephalia. Некоторые анализы показали, что сцилакозавриды ближе к эвтероцефалам, чем к ликозухидам, и поэтому были объединены в кладу Scylacosauria , в то время как другие предположили, что они являются сестринскими таксонами друг друга. основные клады, соответствующие семействам Akidnognathidae , Chthonosauridae , Hofmeyriidae , Whaitsiidae Внутри Eutherocephalia выделяются , а также различные субклады, сгруппированные под Baurioidea. Однако, хотя отдельные группы тероцефалов широко признаны действительными, взаимоотношения между ними часто плохо подтверждаются. [ 17 ] [ 18 ] [ 19 ] Таким образом, существует несколько названных клад более высокого уровня, объединяющих несколько субкладов, за исключением Whaitsiioidea (объединяющих Hofmeyriidae и Whaitsiidae) и Baurioidea.
Филогения
[ редактировать ]Ранние филогенетические анализы тероцефалов, такие как анализ Хопсона и Баргузена (1986) и ван ден Хевера (1994), выявили и подтвердили многие из упомянутых выше подтаксонов тероцефалов в филогенетическом контексте. Однако отношения более высокого уровня было трудно разрешить, особенно между субкладами Eutherocephalia (т.е. Hofmeyriidae, Akidnognathidae, Whaitsiidae и Baurioidea). Например, Хопсон и Баргузен (1986) смогли обнаружить эвтероцефалию только как неразрешенную политомию . [ 13 ] Несмотря на эти недостатки, последующие дискуссии о взаимоотношениях тероцефалов основывались почти исключительно на этом анализе. [ 11 ] Более поздние анализы были сосредоточены на взаимоотношениях ранних цинодонтов, а именно Абдала (2007) и Бота и др. (2007) включили некоторые тероцефалические таксоны и подтвердили существование Eutherocephalia, но также обнаружили, что цинодонты являются сестринским таксоном тероцефалического тероцефала Theriognathus , что делает Therocephalia парафилетическим. [ 20 ] [ 21 ]
Более поздний филогенетический анализ тероцефалов, начатый Хаттенлокером (2009), подчеркивает использование более широкого выбора таксонов и признаков тероцефалов. Такой анализ подтвердил, что Therocephalia является сестринской кладой цинодонтов, а монофилия Therocephalia была подтверждена последующими исследователями. [ 11 ] [ 7 ]
Ниже представлена кладограмма, модифицированная на основе анализа, опубликованного Кристианом А. Сидором, Зоя. Т. Кулик и Адам К. Хуттенлокер в 2022 году упрощенно, чтобы проиллюстрировать взаимоотношения основных признанных тероцефалических субкладов. [ 22 ] Он основан на матрице данных, впервые опубликованной Huttenlocker et al. (2011), [ 8 ] и представляет собой широкие топологии, обнаруженные в других итерациях этого набора данных, таких как Sigurdsen et al. (2012), Хаттенлокер и др. (2014 г.) и Лю и Абдала (2022 г.). [ 23 ] [ 24 ] [ 17 ] Пример лабильности этих отношений продемонстрирован Лю и Абдалой (2023), которые обнаружили альтернативную топологию с Chthonosauridae, глубоко гнездящимися внутри Akidnognathidae. [ 25 ]
| Терапсида |
| ||||||||||||||||||||||||||||||
Ниже представлена модифицированная кладограмма Pusch et al. (2024), анализируя взаимоотношения тероцефалов и ранних цинодонтов. Их анализ был сосредоточен на включении эндокраниальных характеристик, чтобы помочь определить взаимоотношения тероцефалов и цинодонтов, чтобы дополнить предыдущие анализы, которые почти полностью опирались на поверхностные черепные и зубные характеристики, подверженные конвергентной эволюции, и, как таковые, включают только таксоны с доступными применимыми данными. Из них можно было включить только четырех тероцефалов. Однако каждая из них представляет четыре основные группы в филогении тероцефалов: две «базальные тероцефалы» Lycosurus (Lycosuchidae) и Alopecognathus (Scylacosauridae) и два производных члена Eutherocephalia, Olivierosuchus (Akidnognathidae) и Theriognathus (Whaitsiidae). [ 6 ]
Примечательно, что их анализ последовательно показал, что цинодонты и эвтероцефалы являются сестринскими таксонами, а базальные тероцефалы Lycosuchus и сцилакозавриды занимают более базальное положение, что делает тероцефалов такими, какими их традиционно считали парафилетическими. Это отличается от предыдущих предложений о парафилетических тероцефалиях, которые обычно считали цинодонтов наиболее близкими к производным тероцефалам ваицидид. [ 6 ]
|
Традиционные тероцефалы |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Рубидж, бакалавр наук; Сидор, Калифорния (2001). «Эволюционные закономерности пермо-триасовых терапсидов» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 32 : 449–480. doi : 10.1146/annurev.ecolsys.32.081501.114113 . Архивировано из оригинала (PDF) 21 марта 2012 г.
- ^ Jump up to: а б Ивахненко, М.Ф. (2011). «Пермские и триасовые тероцефалы (Eutherapsida) Восточной Европы». Палеонтологический журнал . 45 (9): 981–1144. дои : 10.1134/S0031030111090012 . S2CID 128958135 .
- ^ Татаринов, Л.П. (1994). «О сохранении рудиментарного рострального трубчатого комплекса кистокрылых у териодонтов и о возможном развитии электрорецепторных систем у некоторых представителей этой группы». Доклады Академии наук . 338 (2): 278–281.
- ^ Бенуа, Дж.; Нортон, Луизиана; Менеджер, PR; Рубидж, бакалавр наук (2017). «Переоценка ядовитой способности Euchambersia mirabilis (Therapsida, Therocephalia) с использованием методов микроКТ-сканирования» . ПЛОС ОДИН . 12 (2): e0172047. Бибкод : 2017PLoSO..1272047B . дои : 10.1371/journal.pone.0172047 . ПМК 5302418 . ПМИД 28187210 .
- ^ Полевой музей (13 августа 2015 г.). «Доисторический хищник, получивший прозвище «лицо со шрамом», обнаружен в Замбии» (пресс-релиз). Наука Дейли .
- ^ Jump up to: а б с Пуш, ЛК; Каммерер, CF; Фрёбиш, Дж. (2024). «Происхождение и эволюция Cynodontia (Synapsida, Therapsida): переоценка филогении и систематики самых ранних представителей этой клады с использованием технологий 3D-визуализации» . Анатомическая запись . дои : 10.1002/ar.25394 . ПМИД 38444024 .
- ^ Jump up to: а б Ангельчик, Кеннет Д.; Каммерер, Кристиан Ф. (2018). «Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». В Захосе, Фрэнк Э.; Ашер, Роберт Дж. (ред.). Эволюция, разнообразие и систематика млекопитающих . Берлин: Де Грюйтер. стр. 160–162. ISBN 9783110275902 .
- ^ Jump up to: а б Хаттенлокер, АК; Сидор, Калифорния; Смит, RMH (2011). «Новый экземпляр Promoschorhynchus (Therapsida: Therocephalia: Akidnognathidae) из нижнего триаса Южной Африки и его влияние на выживание териодонтов на границе пермо-триаса». Журнал палеонтологии позвоночных . 31 (2): 405–421. дои : 10.1080/02724634.2011.546720 . S2CID 129242450 .
- ^ Грюнерт, Хенрик Рихард; Броклхерст, Нил; Фрёбиш, Йорг (25 марта 2019 г.). «Разнообразие и неравенство тероцефалий: закономерности макроэволюции через два массовых вымирания» . Научные отчеты . 9 (5063): 5063. Бибкод : 2019НатСР...9.5063Г . doi : 10.1038/s41598-019-41628-w . ПМК 6433905 . ПМИД 30911058 .
- ^ Jump up to: а б с Ван ден Хевер, Дж. (1987). Сравнительная и функциональная морфология черепа ранних Therocephalia (Amniota: Therapsida) (кандидатская диссертация). Университет Стелленбоша.
- ^ Jump up to: а б с д Хаттенлокер, А. (2009). «Исследование кладистических отношений и монофилии тероцефаловых терапсидов (Amniota: Synapsida)» . Зоологический журнал Линнеевского общества . 157 (4): 865–891. дои : 10.1111/j.1096-3642.2009.00538.x .
- ^ Кемп, Т.С. (1982). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. ISBN 9780124041202 .
- ^ Jump up to: а б Хопсон, Дж.А.; Баргузен, Х (1986). «Анализ терапевтических отношений». В Хоттоне, Северная Каролина; Маклин, доктор медицинских наук; Рот, Джей-Джей; Рот, ЕС (ред.). Экология и биология млекопитающих рептилий . Вашингтон: Издательство Смитсоновского института. стр. 83–106.
- ^ ван ден Хевер, JA (1980). «О достоверности тероцефалового семейства Lycosuchidae (Reptilia, Therapsida)» . Анналы Южноафриканского музея . 81 : 111–125.
- ^ ван ден Хевер, JA (1994). «Краниальная анатомия ранней тероцефалии (амниоты: Therapsida)». Анналы Стелленбосского университета . 1 . ISBN 978-0-7972-0498-0 .
- ^ Абдала, Ф.; Каммерер, CF; Дэй, Миссури; Джира, С.; Рубидж, бакалавр наук (2014). «Морфология взрослых тероцефалов Simorhinella baini из средней перми Южной Африки, а также таксономия, палеобиогеография и временное распространение Lycosuchidae». Журнал палеонтологии . 88 (6): 1139–1153. дои : 10.1666/13-186 . ISSN 0022-3360 . S2CID 129323281 .
- ^ Jump up to: а б Лю, Дж.; Абдала, Ф. (2022). «Символическая южноафриканская тероцефаловая эвкамерсия в Китае: новое звено в расселении позднепермских позвоночных по Пангее» . Письма по биологии . 18 (7): 20220222. doi : 10.1098/rsbl.2022.0222 . ПМК 9278400 . PMID 35857894 .
- ^ Каммерер, CF; Масютин, В. (2018). «Новый тероцефал ( Gorynychus masyutinae gen. et sp. nov.) из пермского местонахождения Котельнич, Кировская область, Россия» . ПерДж . 6 . е4933. дои : 10.7717/peerj.4933 . ПМК 5995100 . ПМИД 29900076 .
- ^ Лю, Дж.; Абдала, Ф. (2019). «Фауна четвероногих верхней перми формации Наобаогоу в Китае: 3. Jiufengia jiai gen. и sp. nov., крупный тероцефал акидногнатид» . ПерДж . 7 . е6463. дои : 10.7717/peerj.6463 . ISSN 2167-8359 . ПМК 6388668 . ПМИД 30809450 .
- ^ Абдала, Ф. (2007). «Переописание Platycraniellus elegans (Therapsida, Cynodontia) из нижнего триаса Южной Африки и кладистические отношения эутериодонтов» . Палеонтология . 50 (3): 591–618. Бибкод : 2007Palgy..50..591A . дои : 10.1111/j.1475-4983.2007.00646.x . S2CID 83999660 .
- ^ Бота, Дж.; Абдала, Ф.; Смит, RMH (2007). «Старейший цинодонт: новые сведения о происхождении и разнообразии цинодонтов» . Зоологический журнал Линнеевского общества . 149 : 477–492. дои : 10.1111/j.1096-3642.2007.00268.x .
- ^ Сидор, Калифорния; Кулик, З.Т.; Хуттенлокер, АК (2022 г.). «Новый бауриаморфный тероцефал добавляет новый компонент в комплекс четвероногих нижнего триаса формации Фремоу (Трансантарктический бассейн) в Антарктиде». Журнал палеонтологии позвоночных . 41 (6): e2081510. дои : 10.1080/02724634.2021.2081510 . S2CID 250663346 .
- ^ Сигурдсен, Т.; Хаттенлокер, АК; Модесто, СП; Роу, ТБ; Дамиани, Р. (2012). «Переоценка морфологии и палеобиологии тероцефалов Tetracynodon darti (Therapsida) и филогенетических взаимоотношений Baurioidea». Журнал палеонтологии позвоночных . 32 (5): 1113–1134. дои : 10.1080/02724634.2012.688693 . S2CID 84457790 .
- ^ Хаттенлокер, АК (2014). «Уменьшение размеров тела у немлекопитающих Euteriodont Therapsids (Synapsida) во время массового вымирания в конце перми» . ПЛОС ОДИН . 9 (2). е87553. Бибкод : 2014PLoSO...987553H . дои : 10.1371/journal.pone.0087553 . ПМК 3911975 . ПМИД 24498335 .
- ^ Лю, Дж.; Абдала, Ф. (2023). «Укрепились связи наземной фауны поздней перми: первый тероцефал ваицоид из Китая». Палеонтология африканская . 56 : 16–35. hdl : 10539/35706 .
Внешние ссылки
[ редактировать ] СМИ, связанные с тероцефалией , на Викискладе?
СМИ, связанные с тероцефалией , на Викискладе?  Данные, относящиеся к тероцефалии , в Wikispecies
Данные, относящиеся к тероцефалии , в Wikispecies








