дом
| дом Временной диапазон: ранняя пермь ,
| |
|---|---|

| |
| Скелет C. broilii в Полевом музее естественной истории | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | † Казеазаврия |
| Семья: | † Казеиды |
| Род: | † Дом Уиллистон , 1910 год. |
| Типовой вид | |
| Дом Бройла Уиллистон, 1910 год.
| |
Casea — род травоядных , казеидных синапсидов обитавших в поздней нижней перми ( кунгурском периоде ) на территории современного Техаса , США . Род представлен только типовым видом , Casea broilii названным Сэмюэлем Венделлом Уиллистоном в 1910 году. [ 1 ] Вид представлен черепом, связанным со скелетом ( голотип FMNH UC 656), вторым черепом (FMNH UC 698), частичным черепом с более сохранившимся зубным рядом, чем у предыдущих черепов (FMNH UC 1011), и несколькими неполные посткраниальные скелеты. [ 2 ] Позже были установлены еще три вида Casea , но сегодня они считаются недействительными или принадлежащими к разным родам. [ 3 ] [ 4 ] [ 5 ] Казеа представлял собой небольшое животное длиной около 1,20 м и весом около 20 кг. [ 6 ] [ 7 ]
Этимология
[ редактировать ]Название рода и видовой эпитет даны в честь палеонтологов Эрмин К. Кейс и Фердинанда Бройли . [ 1 ]
Описание
[ редактировать ]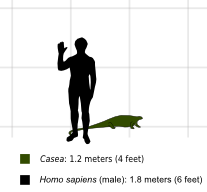
Череп
[ редактировать ]Череп , крышей черепа , , относительно небольшой по сравнению с размером тела, демонстрирует типичную морфологию казеидов с наклоненной вперед мордой украшенной множеством мелких ямок, и большим шишковидным отверстием . Внешние ноздри меньше, чем у более производных казеидов. Орбиты . очень большие, направлены наружу и немного вперед При взгляде сверху конец морды шире и более закруглен, чем у более продвинутых казеид. [ 8 ] [ 6 ] [ 9 ] Нёбо широкое, пластинчатое. Узкая межкрыловидная впадина разделяет задние отделы неба по средней линии. [ 3 ] Кости неба почти полностью покрыты зубами, самые крупные из них расположены по краям, а самые мелкие — в центре неба. Верхние челюсти имели по два зуба на каждой предчелюстной кости и по 11 зубов на каждой верхней челюсти . имелось по 12 зубов На каждой половине нижней челюсти , некоторые из них располагались на венечном возвышении , что является примитивным признаком. Первые шесть зубов верхней и нижней челюстей очень крепкие, конические, почти круглые у основания, но более закругленные на вершине и несколько сжатые медиально-латерально . Верхние челюсти вертикальные, а первые шесть зубов нижней челюсти направлены наружу и вперед под углом сорок градусов и более. FMNH UC 698 видно мало деталей На зубах голотипа FMNH UC 656 и паратипа из-за очень грубой подготовки этих экземпляров. Однако более тщательная подготовка верхних зубов экземпляра FMNH UC 1011 выявила наличие трехстворчатых коронок . [ 8 ] [ 6 ] [ 2 ]
Посткраниальный скелет
[ редактировать ]
Казея — казеид легкого телосложения с довольно короткими по сравнению с длиной животного конечностями. 24 В позвоночнике или 25 прекрестцовых позвонков , а в крестце — три позвонка. Хвост до конца не известен. У трех экземпляров сохранился сочлененный хвостовой ряд, включающий от 18 до 22 позвонков. Исходя из этого, общее количество хвостовых позвонков оценивается примерно в пятьдесят. [ 8 ] [ 6 ] Ребра . образуют бочкообразную грудную клетку , типичную для травоядных казеидов Таз представляет собой подвздошную кость с расширенным дорсальным краем, имеющую форму очень широкого веера. Его медиальная поверхность плоская и гладкая с небольшими пестринами по дорсальному краю. Крестцовые ребра образуют единый, непрерывный контакт с подвздошной костью, который образуется за счет перекрытия первого и второго крестцовых ребер, а также между вторым и третьим ребрами. [ 8 ] [ 6 ] [ 7 ] Голень дистального отличается умеренно увеличенным проксимальным концом, причем последний несколько шире конца , как у Eocasea . У более производных казеидов проксимальный конец большеберцовой кости значительно увеличен. [ 7 ] Неполный скелет Casea broilii (FMNH UR 2514), найденный в типовом местонахождении и описанный только в 2014 году, демонстрирует таранную кость, все еще сочлененную с большеберцовой костью. Таким образом, в отличие от описаний Ромера, Прайса и Олсона, таранная кость Casea broilii представляет собой удлиненный элемент, у которого сочленение малоберцовой кости отделено от сочленения с большеберцовой костью длинной шейкой. Эта разница в интерпретации является результатом неправильной идентификации частичной и изолированной стопы (FMNH UC 657), приписываемой Casea , но, вероятно, принадлежащей другому животному. [ 7 ]

Географический и стратиграфический ареал
[ редактировать ]Все экземпляры Casea broilii происходят из одного ископаемого кармана, известного как Cacops костный пласт , расположенного в округе Бэйлор , штат Техас . [ 10 ] [ 8 ] Это место, открытое и раскопанное Полом Миллером в 1909 и 1910 годах, сегодня больше недоступно, поскольку оно было затоплено в 1920-х годах после строительства плотины озера Кемп . [ 11 ] Карман окаменелостей имел длину около 3 м, ширину 1,8 м и толщину 60 см. [ 10 ] [ 8 ] Это была часть уровня красной глины , вкрапленного между пластами верхней формации Арройо и более грубыми отложениями нижней формации Вейл (две формации группы Клир-Форк ). [ 10 ] [ 8 ] [ 7 ] Фауны аммоноидей, обнаруженные в морских слоях, присутствующих у основания и вершины группы Клир-Форк, указывают на то, что три формации (Арройо, Вале и Чоза), слагающие ее, представляют собой относительно короткую геологическую продолжительность, соответствующую части кунгурского периода . [ 12 ] [ 13 ] Местоположение и фаунистический состав костного пласта Какопс указывают на существование особой фауны, обитавшей в географическом районе, далеком от пойм и дельт прибрежных районов, где обитала хорошо известная фауна из более классических нижнепермских местонахождений Северной Америки. [ 14 ] Экземпляры Casea ассоциировались с очень многочисленными экземплярами панцирных и полностью наземных амфибий Cacops (перечислено более 50 особей, в том числе десять черепов, отсюда и название костного пласта), десятком скелетов Varanopidae Varanops и фрагментами Seymouria. и Капторин . Многие кости были покрыты очень тонким слоем сцементированной глины; другие были более или менее сцементированы в узелковые массы. Большинство скелетов лежало на брюхе, но некоторые были найдены и на спине. У большинства скелетов конечности были сочленёнными. У других фаланги стоп были более или менее разбросаны, а хвост или целые конечности разъединены. [ 10 ] [ 8 ] Судя по положению скелетов и условиям захоронения, после смерти животных тела подверглись очень малому воздействию. По мнению Уиллистона, эти животные погибли бы в луже со стоячей и вечной водой. Трупы, сложенные друг на друга последовательными слоями, соответствуют накоплению, растянувшемуся на несколько поколений. [ 10 ] Однако никаких тафономических исследований памятника не было опубликовано, а учитывая сегодняшнюю недоступность местности, сказать больше сложно.
Таксономия
[ редактировать ]В настоящее время род Casea включает только вид Casea broilii . В прошлом к этому роду относились еще три вида, но сегодня они представляют собой отдельные роды и/или считаются недействительными. В 1954 году Эверетт К. Олсон сообщил о двух новых видах, обнаруженных в группе Клир-Форк в Техасе : Casea nicholsi и C. halselli . [ 15 ] В 1974 году Дениз Сигоно-Рассел и Дональд Э. Рассел установили вид Casea rutena для экземпляра из южной Франции . [ 16 ] Эти три вида известны по следующему материалу:
- Casea nicholsi представлена двумя экземплярами из верхней части формации Вейл в округе Нокс . Голотип (FMNH UR 86) состоит из частичного базикрания и нижней челюсти (в основном сохранившихся в виде естественного слепка), значительной части позвоночного столба (кроме хвоста), части таза, элементов передних конечностей и части стопа. Единственный упомянутый экземпляр (FMNH UR 85) был обнаружен в непосредственной связи с голотипом и, как и он, состоит из частичного черепа, большой части позвоночного столба без хвоста, части плечевых костей, таза, бедренной кости. и конец малоберцовой кости. [ 15 ] [ 2 ]
- Casea halselli происходит из более поздних слоев и была названа в честь очень фрагментарного скелета, найденного в средней части формации Чоза в округе Фоард . Он состоит из тазового пояса с повреждением подвздошной кости, частичной левой бедренной и большеберцовой костей, головки правой бедренной кости, четырех хвостовых позвонков и фрагментов поясничных позвонков. [ 15 ] [ 2 ]
- Casea rutena происходит из нижнепермского бассейна Родез в Аверон департаменте в Окситания регионе , на юге Франции . Он известен по частичному скелету, состоящему из почти полного черепа, шейных позвонков, плечевых костей, полного левого предплечья с полной сочлененной левой кисти и части правой руки. [ 16 ] [ 4 ]
В 2008 году первый филогенетический анализ Caseidae впервые выявил парафилию рода Casea , французского вида Casea rutena , представляющего отдельный и более производный род, до сих пор не названный в этом исследовании. [ 3 ] Три года спустя Casea rutena была исключена из рода Casea и помещена в новый род Euromycter с новой комбинацией Euromycter rutenus . [ 4 ] В 2015 году другое исследование, опубликованное Романо и Никосией, снова признало род Casea парафилетическим. Casea nicholsi идентифицируется как таксон, более тесно связанный с родами Caseoides и Caseopsis, чем с Casea broilii . Таким образом, C. nicholsi определенно принадлежит к другому роду, который, однако, недостаточно известен, чтобы получить название. Романо и Никосия также считают фрагментарный вид Casea halselli nomen dubium , хотя важные различия с типовым видом в форме и прочности бедренных и большеберцовых костей позволяют предположить, что они принадлежат к другому роду, кроме Casea . [ 5 ] По мнению Вернебурга и его коллег, C. halselli представляет собой проблемный таксон неопределенного, возможно, родственного сфенакоморфам. [ 17 ]
Филогения
[ редактировать ]
В первом филогенетическом анализе Caseidae, опубликованном в 2008 году, Casea broilii занимает базальное положение внутри казеид, но, однако, является более производным, чем Oromycter . [ 3 ]
Ниже представлена первая кладограмма Caseidae, опубликованная Мэддином и др. в 2008 году. [ 3 ]
Филогенетический анализ, проведенный Бенсоном, показывает аналогичное положение для Casea broilii . Этот анализ также подтверждает парафилию рода Casea . [ 18 ]
Ниже филогенетический анализ Caseasauria, опубликованный Бенсоном в 2012 году. [ 18 ]
Исследование, опубликованное в 2015 году Romano & Nicosia и включающее почти все Caseidae (за исключением Alierasaurus ronchii из Сардинии , считающегося слишком фрагментарным), показывает аналогичную позицию для Casea broilii . C. nicholsi обнаружен как более производный таксон, более близкий к родам Caseoides и Caseopsis, чем к Casea broilii . [ 5 ]
Ниже представлен самый щедрый филогенетический анализ, опубликованный Romano & Nicosia в 2015 году. [ 5 ]
В филогенетическом анализе, опубликованном в 2022 году Вернебургом и его коллегами, Casea broilii занимает место между Oromycter и Casea nicholsi . Последний занимает более базальное положение, чем в кладограмме Романо и Никосии, восстанавливаясь как более базальный таксон, чем род Euromycter . [ 17 ]
Ниже представлена кладограмма, опубликованная Вернебургом и его коллегами в 2022 году. [ 17 ]
Палеобиология
[ редактировать ]
Кормление и диета
[ редактировать ]Казея представляет собой одно из первых крупных и весьма успешных травоядных среди наземных синапсид. [ 19 ] Среди позвоночных эту стратегию питания можно разделить на множество категорий, включая фоливорность, плодоядность и зерноядность, но среди ранних наземных позвоночных она питается листьями, стеблями, корнями и корневищами. Травоядные животные используют массивное дробление зубных рядов на нёбе и нижней челюсти. [ 3 ] Казеиды принадлежат к самой базальной кладе синапсидов Caseasauria, в которую входят также мелкие плотоядные эотиридиды. [ 3 ] У казеидов на травоядность указывает наличие массивной грудной клетки в грудном и спинном отделах, а также расширенное туловище, простирающееся назад до тазового пояса, с большими ребрами, сросшимися с поясничными позвонками. Это говорит о том, что эта стратегия питания возникла где-то между поздним пенсильванским периодом и ранней перми. [ 3 ] Некоторые казеиды демонстрируют стоматологическую специализацию: в краевом прикусе присутствуют листообразные большие зубцы. [ 20 ]
Передвижение
[ редактировать ]Передвижение Casea включает в себя крестец из трех позвонков в ранних синапсидах и не имеет очевидной связи с размером тела. ЛеБлан и Рейс утверждают, что эта сакральная анатомия была связана с более эффективным передвижением по земле, чем с повышенной нагрузкой на ногу. [ 7 ] Избирательное давление на перенос веса или более эффективные стили передвижения и все более наземный образ жизни, возможно, способствовали повторному приобретению трех крестцовых позвонков у Synapsida. [ 7 ] Развитие прикрепления третьего крестцового ребра к тазу у Synapsids может подтвердить эту гипотезу. [ 7 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Уиллистон, Юго-Запад (1910). «Новые пермские рептилии: Рахитомные позвонки» . Журнал геологии . 18 (7): 585–601. Бибкод : 1910JG.....18..585W . дои : 10.1086/621786 . S2CID 129153998 .
- ^ Jump up to: а б с д Олсон, ЕС (1968). «Семейство Caseidae». Филдиана: Геология . 17 : 225–349.
- ^ Jump up to: а б с д и ж г час Мэддин, ХК; Сидор, Калифорния; Рейс, Р.Р. (2008). «Краниальная анатомия тектона эннатозавров (Synapsida: Caseidae) из средней перми России и эволюционные связи Caseidae». Журнал палеонтологии позвоночных . 28 (1): 176. doi : 10.1671/0272-4634(2008)28[160:CAOETS]2.0.CO;2 . S2CID 44064927 .
- ^ Jump up to: а б с Рейс, РР; Мэддин, ХК; Фрёбиш, Дж.; Фальконне, Дж. (2011). «Новый крупный казеид (Synapsida, Caseasauria) из перми Родеза (Франция), включая переоценку «Casea» rutena Sigogneau-Russell & Russell, 1974» . Георазнообразие . 33 (2): 227–246. дои : 10.5252/g2011n2a2 . S2CID 129458820 .
- ^ Jump up to: а б с д Романо, М.; Никосия, У. (2015). «Кладистический анализ Caseidae (Caseasauria, Synapsida): использование метода взвешивания пробелов для включения таксонов на основе неполных экземпляров» . Палеонтология . 58 (6): 1109–1130. дои : 10.1111/пала.12197 . S2CID 86489484 .
- ^ Jump up to: а б с д и Ромер, А.С.; Прайс, Л.И. (1940). «Обзор пеликозаврии». Специальный доклад Геологического общества Америки . Специальные статьи Геологического общества Америки. 28 : 1–538. дои : 10.1130/SPE28-p1 .
- ^ Jump up to: а б с д и ж г час ЛеБлан, ARH; Рейс, Р.Р. (2014). «Новый посткраниальный материал раннего казеида Casea broilii Williston, 1910 (Synapsida: Caseidae) с обзором эволюции крестца палеозойских синапсидов немлекопитающих» . ПЛОС ОДИН . 9 (12): e115734. Бибкод : 2014PLoSO...9k5734L . дои : 10.1371/journal.pone.0115734 . ПМЦ 4278720 . ПМИД 25545624 .
- ^ Jump up to: а б с д и ж г час Уиллистон, Юго-Запад (1911). Американские пермские позвоночные . Издательство Чикагского университета. стр. 1–146.
- ^ Ангельчик, К.Д.; Каммерер, CF (2018). «Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». В Захосе, FE; Ашер, Р.Дж. (ред.). Справочник по зоологии: эволюция, разнообразие и систематика млекопитающих . Берлин: де Грюйтер. стр. 137 (рис. 5.6: D, H). ISBN 978-3-11-027590-2 .
- ^ Jump up to: а б с д и Уиллистон, Юго-Запад (1910). « Cacops , Desmospondylus ; новые роды пермских позвоночных» . Бюллетень Геологического общества Америки . 21 (1): 249–284. Бибкод : 1910GSAB...21..249W . дои : 10.1130/GSAB-21-249 .
- ^ Болт, младший (1977). « Какопы (амфибия: Labyrinthodontia) из местонахождения Форт-Силл, нижняя пермь Оклахомы» . Филдиана: Геология . 37 : 61–73.
- ^ Лукас, СГ (2006). «Глобальная биостратиграфия и биохронология пермских четвероногих». В Лукасе, СГ; Кассинис, Г.; Шнайдер, JW (ред.). Неморская пермская биостратиграфия и биохронология . Лондон: Геологическое общество, специальная публикация 265. стр. 65–93. ISBN 978-1-86239-206-9 .
- ^ Шнайдер, Дж.В.; Лукас, СГ; Шольце, Ф.; Фойгт, С.; Маркетти, Л.; Кляйн, Х.; Оплюштил, С.; Вернебург, Р.; Голубев В.К.; Баррик, Дж. Э.; Немировска, Т.; Рончи, А.; Дэй, Миссури; Силентьев В.В.; Рёсслер, Р.; Сэйбер, Х.; Линнеманн, У.; Жаринова В.; Шен, СЗ. (2020). «Континентальная биостратиграфия позднего палеозоя – начала мезозоя — ссылки на стандартную глобальную хроностратиграфическую шкалу». Палеомир . 29 (2): 186–238. дои : 10.1016/j.palwor.2019.09.001 . S2CID 210316208 .
- ^ Кемп, Т.С. (1982). «Пеликозавры». В Кемпе, Т.С. (ред.). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. стр. 64–68. ISBN 978-1-86239-206-9 .
- ^ Jump up to: а б с Олсон, ЕС (1954). «Фауна Долины и Чозы: 7. Pelycosauria, семейство Caseidae». Филдиана: Геология . 10 : 193–204.
- ^ Jump up to: а б Сигоно-Рассел, Д.; Рассел, Д.Э. (1974). «Изучение первых казеид (Reptilia, Pelycosauria) из Западной Европы» . Вестник Национального музея естественной истории . Серия 3. 38 (230): 145–215.
- ^ Jump up to: а б с Вернебург, Р.; Шпиндлер, Ф.; Фальконне, Дж.; Стейер, Дж.-С.; Виани-Лиод, М.; Шнайдер, JW (2022). «Новый казеидный синапсид из перми (Гваделупы) бассейна Лодев (Окситания, Франция)» (PDF) . Палеовертебрата . 45 (45(2)-e2): e2. дои : 10.18563/pv.45.2.e2 . S2CID 253542331 .
- ^ Jump up to: а б Бенсон, RBJ (2012). «Взаимоотношения базальных синапсид: краниальные и посткраниальные морфологические перегородки предполагают разные топологии». Журнал систематической палеонтологии . 10 (4): 601–624. дои : 10.1080/14772019.2011.631042 . S2CID 84706899 .
- ^ Броклхерст, Нил; Романо, Марко; Фробиш, Йорг (2017). «Анализ главных компонентов как альтернативный подход к морфометрическим признакам: филогения казеидов на примере». Дриада. дои : 10.5061/dryad.qg91m .
- ^ Уиллистон, Юго-Запад (1913). «Черепа араеосцелиса и казеи, пермских рептилий». Журнал геологии . 21 (8): 673–689. Бибкод : 1913JG.....21..743W . дои : 10.1086/622122 . JSTOR 30058406 . S2CID 140622280 .





