Горгонопсия
| Горгонопсианцы Временной диапазон: средняя пермь — поздняя пермь ,
| |
|---|---|

| |
| Скелет Inostrancevia alexandri в Музее науки , Тренто, Италия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | Терапсида |
| Клэйд : | Териодонтия |
| Клэйд : | † Горгонопсия Сили , 1895 г. |
| Роды | |
Горгонопсия (от греческого Gorgon , мифологического зверя, и óps «аспект») — вымершая клада саблезубых существовавшая терапсидов, от средней до верхней перми , возможно, даже до раннего триаса , примерно между 265 и 252 миллионами лет назад. . Для них характерен длинный и узкий череп, а также удлиненные верхние, а иногда и нижние клыки и резцы, которые, вероятно, использовались как рубящее и колющее оружие. Постклыковые зубы обычно редуцированы или отсутствуют. Для охоты на крупную добычу они, возможно, использовали тактику «укус и отступление», устраивая засаду и нанося изнурительный укус цели, а также преследуя ее на безопасном расстоянии до того, как травмы истощат ее, после чего горгонопсиан хватал животное и наносил удар. убийственный укус. У них был бы непомерный зевок, возможно, более 90°, без необходимости расшатывать челюсть.
С течением времени они заметно увеличивались в размерах: от небольших черепов длиной 10–15 см (4–6 дюймов) в средней перми до медвежьих пропорций до 60 см (2 фута) в верхней перми. Последние горгонопсии, Rubidgeinae , были самыми крепкими в группе и могли наносить особенно сильные укусы. Считается, что горгонопсианы были полностью наземными животными и могли ходить полупрямой походкой, обладая таким же наземным двигательным диапазоном, что и современные крокодилы . Они, возможно, были более подвижными, чем их жертвы, но, вероятно, были инерционными гомеотермами, а не эндотермами, в отличие от современных тероцефалов и цинодонтов , и поэтому, вероятно, были сравнительно менее активными. Хотя горгонопсианы были способны поддерживать довольно высокую температуру тела, неясно, имели ли они также потовые железы или мех (а также усы и связанные с ними структуры). Их мозг напоминал мозг современных рептилий , а не современных млекопитающих . Большинство видов могли быть преимущественно дневные (активные в течение дня), хотя некоторые могли быть сумеречными (активными на рассвете или в сумерках) или ночными (активными ночью). Считается, что у них было бинокулярное зрение , теменный глаз (который улавливает солнечный свет и поддерживает циркадный ритм ), острое обоняние, функциональный вомероназальный орган («орган Якобсона») и, возможно, рудиментарная барабанная перепонка .
Все основные группы терапсидов произошли 275 миллионов лет назад от предка- пеликозавра (плохо определенная группа, включающая все синапсиды , не являющиеся терапсидами). Захват терапсидов у пеликозавров произошел в средней перми, когда мир постепенно становился более засушливым. Горгонопсии стали высшими хищниками своей среды обитания после капитанского массового вымирания , которое привело к гибели диноцефалов и некоторых крупных тероцефалов после средней перми. Несмотря на существование в пермском периоде единого континента, Пангеи , горгонопсии были обнаружены только в супергруппе Кару (в основном в Южной Африке, но также в Танзании, Замбии и Малави), формации Моради в Нигере, на западе России и в Турфанский бассейн в Синьцзяне , Китай , с вероятными остатками, известными из формации Кундарам в бассейне Пранхита-Годавари в Индии . Эти места представляли собой полузасушливые районы с обильным сезонным количеством осадков. горгонопсий Все роды очень похожи по внешнему виду, и, следовательно, многие виды были названы на основе незначительных и, вероятно, возрастных различий с момента их открытия в конце 19 века, и эта группа подверглась нескольким таксономическим пересмотрам.
Большинство горгонопсиев вымерло во время фазы пермско-триасового вымирания, произошедшего в самом конце перми, когда крупная вулканическая активность (которая привела к образованию Сибирских ловушек ) и, как следствие, массивный всплеск парниковых газов вызвали быстрое засушивание из-за температуры. пик, кислотные дожди , частые лесные пожары и потенциальное разрушение озонового слоя . Однако некоторые более мелкие таксоны, такие как Cyonosaurus , возможно, дожили до раннего триаса . Большие хищные ниши были захвачены архозаврами ( а именно крокодилами и динозаврами ) в мезозое .
Описание
[ редактировать ]
Раньше горгонопсиды в средней перми были довольно маленькими, с длиной черепа 10–15 см (4–6 дюймов). [ 1 ] тогда как некоторые более поздние роды достигли массивных медвежьих размеров, самым крупным из которых была Inostrancevia , достигающая 3,5 м (11 футов) в длину и 300 кг (660 фунтов) массы тела. [ 2 ] Тем не менее, мелкие горгонопсии оставались многочисленными до полного исчезновения (хотя мелкие виды на самом деле могут представлять собой молодые экземпляры других таксонов). [ 1 ]
Как и другие пермские терапсиды, горгонопсии развили несколько характеристик млекопитающих. К ним могла относиться парасагиттальная походка (конечности были ориентированы вертикально и двигались параллельно позвоночнику) в отличие от раскидистой походки амфибий и более ранних синапсидов. Такое изменение походки у терапсидов, возможно, было связано с уменьшением размера хвоста и формулы фаланг. [ 3 ] (количество костей на палец, которое у горгонопсов составляло 2,3.4.5,3, как у рептилий [ 4 ] ). Другие разработки включали фиброзную пластинчатую кортикальную кость и глубоко посаженные зубы. [ 3 ] Как и у рептилий, у горгонопсов отсутствует вторичное небо, отделяющее ротовую полость от носовой полости и препятствующее жеванию.
Череп
[ редактировать ]Анатомия горгонопсов невероятно мало различается. [ 5 ] Многие виды отличаются неясными пропорциональными различиями, и, следовательно, более мелкие виды могут фактически представлять собой молодь более крупных таксонов. Примечательно, что сошник на кончике морды различается у разных видов по степени его расширения, а также по положению, степени расширения и форме трех гребней. [ 1 ] Обычно они имеют длинный и узкий череп. [ 4 ] У молодых Rubidgea морда, по-видимому, была шире, чем длинна. [ 6 ] В отличие от эутериодонтов, затылочная кость (в задней части черепа) прямоугольная (ширина больше высоты) и вогнутая, а не треугольная. [ 7 ] : 279

Мозг горгонопсий, как и другие , не относящиеся к млекопитающим терапсиды , лишен расширения неокортекса , имеет относительно крупный по сравнению с передним мозгом задний мозг, большой эпифизарный нерв (обнаруживается у существ с теменным глазом на макушке), увеличенный гипофиз и общая вытянутая форма; в целом напоминающий мозг рептилии . [ 8 ] Мозговая коробка также была скорее рептильной, сравнительно меньше и не такой толстой, как у млекопитающих. [ 7 ] : 280 Флоккулюс вестибулоокулярным , доля мозжечка , пропорционально велика и связана с рефлексом (который стабилизирует взгляд при движении головы). Судя по ориентации полукружных каналов уха (которые должны быть ориентированы параллельно земле), голова горгонопсии GPIT/RE/7124 была бы наклонена вперед примерно на 41°, увеличивая перекрытие между поля зрения двух глаз и улучшение бинокулярного зрения — полезно для хищника. В отличие от рептилий и млекопитающих , полукружные каналы плоские, вероятно, потому, что они вклинены между опистотической ( внутренней ушной костью) и надзатылочной костями. [ 8 ]
Зубы
[ редактировать ]
Как и многие млекопитающие, горгонопсии были гетеродонтами с четко выраженными резцами , клыками и постклыковыми зубами, премолярам и гомологичными коренным зубам . [ 1 ] У них было пять резцов на верхней челюсти (у большинства первые три были одинакового размера, а последние два были короче) и четыре на нижней. [ 7 ] : 17–18
У большинства горгонопсий резцы были крупными, а верхние клыки были удлинены в сабли, во многом как у более поздних саблезубых кошек . У некоторых горгонопсий были исключительно длинные верхние клыки, такие как Inostrancevia , а у некоторых из них был выступ на нижней челюсти, который закрывал кончик клыка, когда пасть была закрыта. Обычно считается, что сабли использовались как колющее или рубящее оружие, для чего требовалось чрезвычайно широкое раскрытие зева. И верхние, и нижние клыки Рубиджеи были удлиненными, и животному понадобился бы еще больший зевок. [ 9 ] Зубчатый рисунок горгонопсов был больше всего похож на зубчатый рисунок тероподовых динозавров , чем на другие синапсиды. [ 10 ] Небо также имеет бугристость и гребни, на которых часто имеются функциональные зубы, которые, возможно, использовались для удержания борющейся добычи, отвлекая эти мощные силы от хрупких клыков. Подобные гребни были идентифицированы на махайродонте Homotherium . [ 7 ] : 280 Постклыковые зубы уменьшились как в размере, так и в количестве; многие рубиджины (поздние горгонопсии) не имели постклыков на нижней челюсти, [ 11 ] а у Клеландины их совсем не было. [ 6 ]
Горгонопсианы были полифиодонтами , и зубы у них росли непрерывно на протяжении всей жизни человека. [ 12 ] Как и у некоторых терапсидов, пока существовал один функционирующий клык, другой клык вырастал, чтобы заменить его, когда он неизбежно ломался. Левая и правая стороны челюстей не обязательно должны были быть синхронными, поэтому, например, первый клык на левой стороне мог функционировать, в то время как первый клык на правой стороне все еще рос. [ 12 ] Такой метод мог быть использован для того, чтобы всегда иметь набор функциональных клыков, поскольку наличие одного клыка или его отсутствие серьезно затрудняло бы охоту, а выращивание таких больших зубов занимало много времени. С другой стороны, поскольку функциональный клык обычно находится в лунке переднего зуба (а не в равном количестве в каждой лунке), вполне возможно, что замена клыка происходила конечное число раз, и в конечном итоге у животного остался бы единственный зуб. , постоянный набор функциональных клыков в этих лунках. В 1984 году британские палеонтологи Дорис и Кеннет Кермак предположили, что клыки росли до размера черепа и постоянно ломались, пока животное не перестало расти, и что у горгонопсов наблюдалась ранняя версия конечной замены зубов, наблюдаемая у многих млекопитающих. Схема замещения остальных зубов неясна. [ 9 ] Постклыковые зубы заменялись медленнее, чем остальные зубы, вероятно, из-за отсутствия у них функционального значения. [ 1 ]
Посткраниум
[ редактировать ]
Все семь шейных позвонков (в области шеи) одинакового размера, за исключением последнего, который короче и ниже; есть один атлас и одна ось . Как и у саблезубых кошек, шея длинная, с хорошо развитой мускулатурой, что было бы особенно полезно, когда клыки были втоплены в животное. Как и другие ранние синапсиды, горгонопсианы имеют единственный затылочный мыщелок , а сочленение (суставы) шейных позвонков в целом рептильное, что позволяет двигать головой из стороны в сторону, но ограничивает движение вверх и вниз. Последний шейный отдел по форме больше напоминает спинные позвонки . [ 7 ] : 291–293
Спинные кости имеют форму катушки и все выглядят примерно одинаково друг с другом. Остистые отростки круто отходят от центра и имеют острые кили на передней и задней сторонах. В отличие от эутериодонтов, горгонопсианы не имеют выраженных поясничных позвонков . Тем не менее, спинные кости, относящиеся к этой серии, похожи на поясничные отделы саблезубых кошек с круто ориентированными зигопофазами, что полезно для стабилизации нижней части спины, особенно при удержании борющейся добычи. [ 7 ] : 293–295
три Крестцовых позвонков , и ряд прикрепляется к тазу первым позвонком. Таз рептильный, с разделенными подвздошной , седалищной и лобковой костью . Бедренная кость имеет слегка S-образную форму, короткая, но длиннее и тоньше плечевой кости . У большинства большеберцовая и малоберцовая кость сильно изогнуты друг к другу, причем большеберцовая кость более прочная, чем малоберцовая. [ 7 ] : 295–299 Сустав между лодыжкой и пяточной костями мог быть несколько подвижным. Пятый палец как на руках, так и на ногах не был прикреплен к запястью / предплюсне , а вместо этого соединялся непосредственно с локтевой /пяточной костью. [ 13 ]
Таксономия
[ редактировать ]Местонахождения, содержащие ископаемые
[ редактировать ]
В 1876 году первые останки горгонопсов были идентифицированы в группе Бофорта в супергруппы Кару Южной Африке биологом и палеонтологом Ричардом Оуэном . Он классифицировал окаменелости как Gorgonops torvus , объединив греческое слово Горгона , мифологическое животное, со словом óps ( ὄψ ), что означает «аспект». [ 14 ] В Африке горгонопсии также были обнаружены в обнажениях Кару в долине Рухуху в Танзании, в долине Верхней Луангвы в Замбии и Чивете в в Малави . [ 7 ] : 7
Горгонопсии были впервые идентифицированы в России в 1890-х годах в местонахождении Соколки на Северной Двине в Сибири под руководством русского палеонтолога Владимира Прохоровича Амалицкого . В посмертной публикации он был описан как Inostrancevia alexandri , и это один из самых известных и крупнейших горгонопсов. С тех пор были описаны еще лишь несколько русских таксонов: Православлевия , Зауроктонус , Вяткогогоргон , Сухогоргон и Ночница . [ 15 ] Один из названных русских родов, Леогоргон , сейчас считается nomen dubium . [ 6 ]
Горгонопсианы явно отсутствуют за пределами этих двух областей. [ 15 ] В 1979 году китайский палеонтолог Ян Чжунцзянь описал китайского горгонопсина Wangwusaurus tayuensis на основе зубов из позднепермской формации Цзиюань . [ 16 ] но в 1981 году палеонтологи Дениз Сигоньо-Рассел и Ай-Лин Сан обнаружили, что назначенный материал представляет собой случайное собрание, из которых только два имеют хотя бы отдаленное сходство с горгонопсией. [ 17 ] В 2011 году неполный набор зубов был обнаружен в местности в пределах Турфанской впадины , Синьцзян , на северо-западе страны. Ископаемый материал, хотя и тонкий, описан в 2022 году палеонтологами Цзюнь Лю и Ван Яном и подтверждает, что он произошел от горгонопсиана, датируемого верхней перми, который на самом деле жил на территории современного Китая. [ 18 ] В 2003 году индийские палеонтологи Сангамитра Рэй и Сасвати Бандиопадьяй отнесли несколько фрагментов черепа из позднепермской формации Кундарам к горгонопсу среднего размера, [ 19 ] хотя характеристики горгонопсиев также были зарегистрированы у некоторых тероцефалов. [ 15 ] В 2008 году в позднепермской формации Моради в Нигере (одной из немногих низкоширотных формаций, содержащих четвероногих) был обнаружен рубиджейновый фрагмент верхней челюсти и клык. Это первое свидетельство существования низкоширотной горгонопсии. . [ 20 ]
Классификация
[ редактировать ]
После открытия Оуэн предположил, что горгонопс и несколько других описанных им таксонов из супергруппы Кару были хладнокровными рептилиями, несмотря на то, что у них были зубы, напоминающие зубы хищных млекопитающих. Он предложил отнести их всех к недавно созданному отряду Theriodontia (который он поместил в класс Reptilia). Он решил разделить Theriodontia на семейства , основанные на анатомии ноздрей (костных ноздрей): «Mononarialia» для тех, у кого в черепе есть одно отверстие для носа, как у млекопитающих, «Binarialia» для тех, у кого есть два отверстия, как у рептилий, и «Тектинариалия» для Горгонопса , потому что его отверстие было затенено толстой костяной крышей. [ 14 ] ( tectus означает в переводе с латыни «покрытый, крытый, палубный»). [ 21 ] В 1890 году английский натуралист Ричард Лидеккер сделал горгонопса типовым видом семейства Gorgonopsidae. [ б ] [ 22 ] Британский палеонтолог Гарри Сили в 1895 году полагал, что у горгонопса отсутствует отверстие в височной кости (височное окно), что является диагностическим признаком Theriodontia, и поэтому возвел Gorgonopsidae в Gorgonopsia, отличный от Theriodontia. Он отнес все южноафриканские материалы, несущие черты как рептилий, так и млекопитающих, к отряду «Theriosuchia» и считал подотрядами Gorgonopsia и Theriodontia. его [ 23 ] Американский палеонтолог Генри Фэйрфилд Осборн полностью переработал классификацию Reptilia в 1903 году и выделил две основные группы: Diapsida и Synapsida . [ 24 ] а в 1905 году южноафриканский палеонтолог Роберт Брум создал третью группу, Therapsida , для размещения «млекопитающих рептилий», включая Theriodontia. Он также оспорил утверждение Сили и отнес Горгонопса обратно к Териодонтии, но поместил его в свою недавно созданную подгруппу Тероцефалия , растворив Горгонопсию. [ 25 ] В 1913 году, особенно в свете почти полного черепа G. torvus, обнаруженного преподобным Джоном Х. Уэйтсом, Брум восстановил статус горгонопсии. [ 26 ]

Число южноафриканских родов быстро росло в 20-м веке, и его возглавлял главным образом Брум, чьи обширные исследования терапсидов Кару — с начала его карьеры в стране в 1897 году до его смерти в 1951 году — привели к описанию 57 южноафриканских родов. голотипов горгонопсий экземпляры и 29 родов. Многие таксоны Брума позже были признаны недействительными. [ 27 ] Многие другие современные исследователи создали совершенно новые виды или роды на основе единичных экземпляров. [ 7 ] : 57 Следовательно, Gorgonopsia стала предметом большого таксономического хаоса и является одной из наиболее проблемных групп синапсид. Поскольку анатомия черепа разных таксонов очень мало различается, многие из них определяются на основе расплывчатых пропорциональных различий, включая даже хорошо известных представителей. Номинальные виды отличаются преимущественно признаками, которые, как известно, весьма изменчивы в зависимости от возраста особи, включая размер глазницы , длину морды и количество послеклыковых зубов. Таким образом, возможно, что некоторые таксоны являются синонимами друг друга и представляют разные стадии развития. [ 28 ]
Среди первых попыток организовать кладу были предприняты британским зоологом Дэвидом Мередитом Сирсом Уотсоном и американским палеонтологом Альфредом Ромером в 1956 году, которые разделили ее на двадцать семейств, представители трех из которых (Burnetiidae, Hipposauridae и Phthinosuchidae) не рассматриваются. горгонопсиан больше нет. [ 6 ] В 1970 и 1989 годах, рассматривая преимущественно африканские таксоны, Сигоньо-Рассел опубликовал обширную монографию о Gorgonopsia (определяя ее как инфраотряд ) и признал только два семейства: Watongiidae и Gorgonopidae. [ 29 ] Ватонгия была перенесена в Varanopidae в 2004 году канадскими палеонтологами Робертом Р. Рейсом и Мишелем Лореном . [ 30 ] Сигоньо-Рассел разделил Gorgonopidae на три подсемейства — Gorgonopsinae, Rubidgeinae и Inostranceviinae — и сократил количество родов до двадцати трех. [ 29 ] В 2002 году российский палеонтолог Михаил Феодосиевич Ивахненко , рассматривая российские таксоны, вместо этого посчитал горгонопсию подотрядом и сгруппировал ее вместе с диноцефалиями в отряд «Горгодонтия». Он разделил горгонопсию на надсемейство «Gorgonopioidea» (семейства Gorgonopidae, Cyonosauridae и Galesuchidae) и «Rubidgeoidea» (Rubidgeidae, Phtinosuchidae и Inostranceviidae). [ 31 ] В 2007 году биолог Ева В.И. Гебауэр в своем всестороннем обзоре горгонопсии (ее докторская диссертация) отвергла модель Ивахненко в пользу модели Сигоньо-Рассела. [ 7 ] : 57 и далее сократил число родов до четырнадцати в дополнение к русским родам: Aloposaurus , Cyonosaurus , Aelurosaurus , Sauroctonus , Scylacognathus , Eoarctops , Gorgonops , " Dixeya " nasuta (под неофициальным номеном nudum " Njalila "), Lycaops , Aelctothus , Aelctothus. , Сикозавр , Клеландина и Рубиджея . [ 7 ] : 244 В целом модель Сигоньо-Рассела поддерживается, но нет единого мнения относительно того, какие роды к каким подсемействам можно отнести. [ 6 ] В 2015 году американский палеонтолог Кристиан Ф. Каммерер и его коллеги переописали эрифостому (которая была обозначена как неопределенный териодонт) как горгонопсиан. [ 32 ] и погрузил Scylacognathus , а в следующем году Eoarctops . в него [ 6 ]

Первая филогения (генеалогическое древо) представителей Gorgonopsia была опубликована в 2016 году американским палеонтологом Кристианом Ф. Каммерером, который специально исследовал Rubidgeinae и заново описал как подсемейство, так и девять отнесенных им видов (сократив их число с тридцати -шесть видов). Каммерер также воскресил Диногоргона , Леонтозавра , Рухухуцербера и Смилезавра . Каммерер не был уверен, представляют ли Leontosaurus , Clelandina , Dinogorgon и Rubidgea один и тот же таксон или нет (для которого Dinogorgon имеет приоритет ), но он решил классифицировать их всех в трибу Rubidgeini в ожидании дальнейшего изучения. [ 6 ] В 2018 году Каммерер и российский палеонтолог Владимир Масютин определили новый род Nochnitsa как самый базовый из известных горгонопсий и обнаружили, что все российские таксоны (кроме Вяткогогоргона , который находится во внекладе) образуют совершенно отдельную кладу от африканских таксонов. [ 15 ] Также в 2018 году палеобиолог Ева-Мария Бендель, Каммерер и коллеги воскресили цинариопсов . [ 1 ] В 2022 году Каммерер и его коллега-палеонтолог Брюс С. Рубидж описали Форция из Южной Африки. [ 33 ]
| |||||||||||||||||||||
| Филогения горгонопсии по Каммереру и Рубиджу (2022) [ 33 ] |
Эволюция
[ редактировать ]Synapsida традиционно разделяется на базальную Pelycosauria и производную Therapsida. К первым относятся хладнокровные существа с раскидистой походкой и предположительно более низким метаболизмом, возникшие в верхнем каменноугольном периоде . В середине-конце 20-го века американский палеонтолог Эверетт К. Олсон исследовал разнообразие синапсид в среднепермских формациях Сан-Анджело , Цветочный горшок и Чикаша в Северной Америке и отметил, что разнообразие пеликозавров в этих формациях сократилось с шести до трех, и что они сосуществовали с несколькими фрагментарными экземплярами, которые он интерпретировал как терапсиды. Затем он предположил, что адаптивный сдвиг от уровня пеликозавров к уровню терапсидов произошел в средней перми (« Вымирание Олсона »); однако с тех пор классификация этих «терапсидов» и возраст образований были поставлены под сомнение. Таким образом, точное время захвата терапсидов неясно, но шесть основных групп терапсидов ( Biarmosuchia , Dinocephalia , Anomodontia , Gorgonopsia, Therocephalia и Cynodontia) развились 265 миллионов лет назад во время Вордиан . [ 34 ] Самый старый экземпляр горгонопсии представляет собой частичную морду, принадлежащую неопределенному роду из Eodicynodon зоны объединения в бассейне Кару, датируемую примерно вордианским периодом. Phorcys, Кару тапиноцефалов известный из самого нижнего конца зоны сборки , датируемый примерно чуть позже границей Ворда и Капитана, является старейшим идентифицируемым таксоном горгонопсов. [ 33 ]
| Сторона синапса [ 34 ] |

Пермь постепенно становилась все суше и суше. В верхнем карбоне и нижней перми пеликозавры, по-видимому, цеплялись за места обитания вечно влажных угольных болот вблизи экватора (окаменелости известны в пределах 10 ° по обе стороны от палеоэкватора); за этим примерно до 30° простиралась обширная пустыня, простиравшаяся до самого побережья, отделявшая болота от областей с умеренным климатом. К средней перми экваториальные леса перешли на сезонную влажную/сухую систему, но болота были связаны с зонами умеренного климата через прибрежные проходы вдоль Восточной Пангеи, что позволило осуществить межконтинентальную миграцию из того, что сейчас является Южной Африкой, в то, что сейчас является Россией. . Терапсиды, по-видимому, развились в этом сезонно влажном/сухом ландшафте, распространившись даже на умеренные зоны. На тот момент синапсиды были единственными крупными наземными животными в их среде обитания; а пеликозавры, возможно, не смогли адаптироваться к засушливости. Примерно во время вымирания пеликозавров терапсиды испытали серьезную адаптивную тенденцию. радиация (все хищники) продолжалась и в верхнюю пермь. [ 34 ]
На протяжении всей средней перми доминирующими животными в своих экосистемах были зачастую гигантские диноцефалы. Они исчезают из летописи окаменелостей во время капитанского массового вымирания, вызванного вулканической активностью, которая образовала китайские ловушки Эмэйшань . Точная причина их вымирания неясна, но на смену им пришли горгонопсии и дицинодонты (которые начали сильно увеличиваться в размерах) и более мелкие тероцефалы. [ 35 ] Рубиджи были наиболее развитыми горгонопсианами и, следовательно, самыми массивными и тяжело сложенными. [ 6 ]
Палеобиология
[ редактировать ]Кусать
[ редактировать ]
Горгонопсианы, вероятно, были активными хищниками. Рубиджины имеют особенно крепкий череп среди горгонопсов, сравнимый с черепами огромных макрохищников, которые используют свои черепа в качестве основного оружия, таких как мозазавры или некоторые динозавры -тероподы . Менее крепкие горгонопсы с более длинными клыками и гораздо более слабым укусом, такие как Smilesaurus или Inostrancevia , вместо этого, вероятно, использовали свои клыки для рубящих ударов, что гораздо больше похоже на саблезубых кошек. Пост-клыки у Clelandina были заменены гладким гребнем, в отличие от дицинодонтов, у которых есть лезвийный ороговевший гребень, и они, возможно, преимущественно преследовали добычу, которую могли проглотить целиком. Таксоны горгонопсий действительно сосуществовали друг с другом - целых семь одновременно - и тот факт, что некоторые рубиджины обладают постклыками, в то время как некоторые другие современные виды не позволяют предположить, что они практиковали разделение ниш и преследовали разные объекты добычи. [ 6 ]
Обычно считалось, что удлиненные клыки сыграли важную роль в их охотничьей тактике. горгонопсианов Шарнир челюсти имел двойные суставы и состоял из подвижных и вращающихся костей, что позволяло им невероятно широко открывать рот — возможно, более чем на 90° — без необходимости расшатывать челюсть. [ 36 ] В качестве альтернативы было высказано предположение (впервые в 2002 году биологами Блэром Ван Валкенбургом и Тайсоном Секко, хотя и в отношении кошек), что сабли развились в первую очередь в результате полового отбора как формы демонстрации спаривания . Это наблюдается у некоторых современных видов оленей, но это трудно проверить, учитывая отсутствие живых саблезубых синапсидных хищников. Считается, что у саблезубых кошек длинносаблезубые («кинтозубые») таксоны были охотниками-преследователями , тогда как короткозубые («ятаганзубые») таксоны, как полагают, были хищниками, устраивавшими засады . [ 37 ] Предполагается, что среди короткозубых кошек эти хищники убивали удачным ударом в горло после схватки добычи, но горгонопсии, возможно, были менее точны в нанесении укусов, вооруженные рептильными челюстями и расположением зубов. Вместо этого горгонопсианы, возможно, использовали тактику укуса и отступления: хищник устраивал засаду на свою добычу и наносил ей значительный и изнурительный укус, а затем следовал за жертвой, пытавшейся убежать, прежде чем погибнуть от раны, после чего горгонопсиан наносил удары. смертельный укус. Поскольку постклыки редуцированы или полностью отсутствуют, мясо пришлось бы насильно отрывать от туши и проглатывать целиком. [ 36 ] Предполагается, что эта стратегия «прокола-вытягивания» использовалась динозаврами-тероподами. [ 10 ]

Горгонопсии, наряду с другими ранними хищниками, а также крокодилами, преимущественно полагались на «кинетико-инерционную систему» (КИ) кусания добычи, при которой крыловидные и височные мышцы быстро сжимали челюсти, используя импульс и энергию кинетическую челюсти и зубы, чтобы схватить жертву. Хищные млекопитающие, в том числе саблезубые кошки, вместо этого полагаются в основном на «систему статического давления» (SP), где височные и жевательные мышцы создают сильную силу укуса, чтобы убить добычу. [ 11 ] Височная и жевательная мышца разделились только у млекопитающих, а у горгонопсов вместо этого была мышца, тянущаяся от нижней части крыши черепа назад к чешуйчатой кости (в задней части черепа) и через скулы. Часть, прикрепленная к щекам, стабилизировала челюстную кость и позволяла ей двигаться из стороны в сторону при смыкании. Это могло быть очень важно при кусании, поскольку скулы становятся сильнее вместе с удлинением клыков. [ 7 ] : 278
Меньшие горгонопсии, такие как цианозавр (который на самом деле может представлять собой молодь другого вида), имели изящные черепа и сабли и, возможно, вели себя во многом подобно шакалам и лисицам . Более крупные горгонопсии, такие как горгонопс , имели длинную крепкую морду с сильно расширенными щеками, которые поддерживали сильные крыловидные кости и мощный укус KI. среднего размера Арктогнат имел коробчатый череп и, как следствие, мощную морду, которая позволяла совершать сильные изгибы и скручивания, а также комбинацию элементов укуса KI и SP. Даже более крупные горгонопсианы, такие как Arctops , имели более короткую и выпуклую морду, как более ранний сфенекодонт Dimetrodon , и могли быстро сжимать челюсти, открывая широкий зев (что было необходимо, учитывая длинные клыки). Еще более крупные Rubidgeinae имели чрезвычайно мощные, массивные, укрепленные черепа, с широкой мордой, сильно выступающими щеками и чрезвычайно длинными зубами; сабли Rubidgea atrox длиннее, чем зубы Тираннозавр . [ 11 ]
В отличие от хищных млекопитающих, у горгонопсов (и тероцефалов) были редуцированы или полностью отсутствовали постклыки, и челюсть, вероятно, не могла оказывать срезающее давление, необходимое для раздавливания костей и доступа к костному мозгу . [ 38 ] Во многом неясно, развился ли вообще костный мозг в пермских синапсидах (рыбы и многие амфибии в настоящее время лишены его), но в 2021 году было показано, что раннепермские амфибии Seymouria и Discosauriscus, вероятно, имели гемопоэтические (продуцирующие эритроциты) ) костный мозг в конечностях. [ 39 ]
Передвижение
[ редактировать ]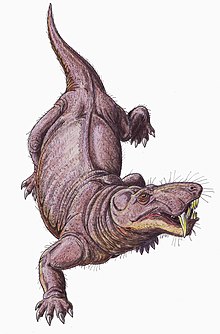
Считается, что горгонопсианы были строго наземными животными . [ 40 ] Считается, что они могли двигаться прямой походкой, подобной той, которую используют крокодилы, конечности располагались почти вертикально, а не горизонтально, как при раскинутой походке ящериц. Суставная впадина лопатки сильно наклонена назад, поэтому конечности имели ограниченное движение вперед и, возможно, имели короткую длину шага. Ящерицы часто двигают позвоночником из стороны в сторону, чтобы увеличить длину шага, но более вертикально ориентированные фасеточные суставы, соединяющие позвонки у горгонопсов, сделали бы позвоночник более жестким и стабильным, затрудняя такое движение. [ 7 ] : 259–260
Плечевой сустав горгонопсий имеет весьма необычную конфигурацию. Головка плечевой кости, которая соединяется с плечом, длиннее суставной впадины, поэтому не могла войти в полость. Следовательно, они могли быть прикреплены к большой массе хряща, при этом плечевая кость совершала перекатывающееся движение по суставной впадине. Теоретически это может привести к тому, что угол между плечевой костью и гленоидом может составлять от 80 до 145 °, если смотреть на животное. Если бы угол находился на нижнем конце, это был бы довольно прочный сустав, позволяющий дельтовидным мышцам прилагать большую силу через переднюю конечность, например, при прижимании сопротивляющейся добычи или удерживании туши при отрывании плоти. Если бы плечевая кость была расположена под большим углом, это могло бы позволить увеличить разгибание вперед и назад (вдоль длинной оси) и, следовательно, увеличить длину шага, что полезно при атаке или коротких погонях. Лопатка расширяется в стороны животного (выступает в стороны), обеспечивая также большое прикрепление дельтовидных мышц. Все лопаточно-плечевые мышцы имели сильно развитые крепления, особенно дельтовидные. При разгибании передних конечностей дельтовидные мышцы могли поднимать переднюю сторону (передний край) плечевой кости, а клювовидно-плечевая мышца опускала заднюю сторону (задний край). При втягивании передней конечности грудная мышца могла толкать передний край вниз, а подлопаточная мышца тянула задний край вверх. [ 7 ] : 260–264
Тазовый сустав имеет обычную шаровидную конфигурацию. Несколько уплощенная головка бедренной кости теоретически могла входить в тазобедренный сустав под разными углами. В 1982 году палеонтолог Том С. Кемп предположил, что ранние териодонты, в том числе горгонопсианы, могли располагать бедренную кость как под горизонтальным углом при раскинутой походке, так и под более вертикальным углом при прямой походке. Он сравнил двигательные привычки этих существ с двигательными привычками крокодилов, которые на коротких дистанциях используют раскидистую походку, но переходят на прямую походку во время бега или перемещения на большие расстояния. Хотя бедро экземпляра GPIT/RE/7113 анатомически занимает промежуточное положение между диметродоном и млекопитающими (подвздошная кость больше расширена в направлении к голове, чем к хвосту, а лобковая кость несколько уменьшена), лобково-бедренная мышца (крупная мышца, несущаяся только у рептилий, который проходит от таза до бедренной кости) широко прикреплен к нижней стороне лобковой и седалищной кости , что позволило бы ему производить сильную приводящая сила (притягивание ног ближе к телу), полезная при раскинутой походке. Также возможно, что горгонопсианы в основном задействовали эту мышцу во время борьбы с добычей. [ 7 ] : 264–270 Голени относительно короткие по сравнению с бедренными костями, что позволяет предположить, что горгонопсианы не были хорошо приспособлены к бегу на длинные дистанции. [ 7 ] : 298–299

Что касается расположения ступней на земле, то горгонопсии — единственные ранние терапсиды, у которых наблюдается эктатаксония (последний палец имеет наибольший вес), гомоподия (следы ног и ладоней выглядят одинаково) и полуплантиградия (в некоторой степени ноги стояли плашмя на земле). [ 41 ] Эти адаптации, возможно, сделали горгонопсов более быстрыми и проворными, чем их жертва. [ 36 ] У горгонопсианцев были довольно проворные пальцы, что свидетельствовало об их способности хватать как руки, так и ноги, возможно, для захвата борющейся добычи, чтобы предотвратить чрезмерную нагрузку и, как следствие, перелом или поломку клыков, когда они погружались в жертву. [ 13 ]
Чувства
[ редактировать ]
В отличие от эутериодонтов, но, как и у некоторых современных экзотермических существ, все горгонопсии обладали шишковидным глазом на макушке головы, который используется для определения дневного света (и, следовательно, оптимальной температуры для активности). Вполне возможно, что другие териодонты утратили это из-за эволюции либо эндотермии, присущих светочувствительным ганглиозным клеткам сетчатки в глазах – в тандеме с потерей цветового зрения и переходом к ночной жизни – либо того и другого. [ 42 ] Долгое время считалось, что ночное поведение зародилось у млекопитающих ( «ночное узкое место» ), но большой размер орбиты и наличие склеротических колец во многих ранних синапсидах, возникших еще в каменноугольном периоде, позволяют предположить, что способность выходить на улицу в низко- Условия освещения возникли гораздо раньше. Основываясь на этих аспектах, экземпляр SAM-PK-K10034 мог обладать мезопическим зрением , а также цианозавра скотопическим или фотопическим зрением . [ 43 ] Диаметр склеротических колец маленькой Вяткогоргоны пропорционально велик: внутренний диаметр 1,5 см (0,6 дюйма) и внешний диаметр 2,3 см (0,9 дюйма) по сравнению с диаметром орбиты 2,8 см (1,1 дюйма). сам по себе, что позволяет предположить, что он совершал преимущественно ночные экскурсии. [ 15 ] Среди горгонопсий рубиджейн Clelandina имеет необычно маленькие склеротические кольца, что указывает на то, что он обладал фотопическим зрением и вел строго дневной образ жизни ; Каммерер предположил, что разделение ниш среди рубидгеинов (поскольку на одной территории сосуществовало до семи различных таксонов) отчасти приняло форму активности разных видов в разное время суток, но склеротические кольца только у Clelandina среди это подсемейство было идентифицировано, что делает эту гипотезу весьма спекулятивной. [ 6 ]

Горгонопсии имеют довольно короткую носовую полость, как и пеликозавры, но с обильными продольными гребнями за внутренними ноздрями (которые соединяют носовую полость с горлом); поскольку вдыхаемый воздух не проходил через них, их обычно интерпретируют как обонятельные носовые раковины, и они давали горгонопсиям довольно сильно развитое обоняние. [ 44 ] Горгонопсианы обладали вомероназальным органом («органом Якобсона») — частью добавочной обонятельной системы — который располагался у основания носовой перегородки ; в отличие от дицинодонтов и тероцефалов, по-видимому, существовал канал, соединяющий орган со ртом, что указывает на то, что у горгонопсов он функционировал. [ 45 ]
Ранние териодонты (включая горгонопсианов), возможно, обладали барабанной перепонкой , в отличие от более ранних пеликозавров, о чем свидетельствует уменьшение соединения между квадратной костью (в шарнире челюсти) и крыловидной костью (в районе неба ), что позволяло квадратной кости независимо вибрировать. степень. Это могло позволить обнаруживать передающиеся по воздуху звуки с низкой амплитудой менее 1 мм (0,039 дюйма), но барабанная перепонка поддерживалась бы хрящом или связками , а не костью. Если это верно, то постдентальные кости (которые у ранних млекопитающих образуют кости среднего уха ) должны были бы отделиться от зубной кости (челюстной кости); летопись окаменелостей горгонопсов, по-видимому, указывает на то, что постдентально-дентарная связь уменьшилась. Хотя, учитывая специализацию, необходимую для кусания, состояние изолирующей квадратной кости у горгонопсов можно альтернативно объяснить как стрептостилия (поворотная квадратная кость), позволяющая расширить зияние, а не облегчить слух. [ 3 ]
Терморегуляция
[ редактировать ]Между более ранними пеликозаврами и терапсидами произошел серьезный анатомический сдвиг, который, как предполагается, был связан с усилением метаболизма и возникновением гомеотермии (поддержания высокой температуры тела). Эволюция вторичного неба и отделение рта от полости носа могли привести к повышению эффективности вентиляции, связанной с высоким уровнем аэробной активности; у горгонопсов не было костного вторичного неба, но, возможно, оно было из мягких тканей. Тем не менее, вторичное небо могло вместо этого способствовать употреблению большого количества пищи за один раз, а не вентиляции. Предполагается, что реорганизация скелета (от раскидистой походки к парасагиттальной) свидетельствует о наличии диафрагмы и , следовательно, об усилении вентиляции для аэробной активности; но вместо этого это могло быть связано с увеличением ускорения или ловкости, что не обязательно означает интенсивную аэробную активность, как у крокодилов. Фиброзная пластинчатая кортикальная кость, которая была у всех ранних терапсидов, указывает на повышенную скорость роста, но это может не быть связано со скоростью метаболизма. [ 46 ]
Современные крупные рептилии естественным образом выделяют тепло тела медленнее, чем более мелкие, и считаются «инерционными гомеотермами», но поддерживают низкую температуру тела 25–30 ° C (77–86 ° F). Если бы терапсидам требовалась более высокая температура тела — 35–40 °C (95–104 °F), они должны были бы либо быть эндотермами (генерирующими собственное тепло тела), либо иметь больший контроль над потерей тепла (то есть лучше гомеотермия). Парасагиттальная походка, возможно, помогла последнему, поскольку она удерживала бы большую часть тела над землей, а также позволяла крови оставаться в брюшной полости вместо того, чтобы циркулировать через придатки, что уменьшило бы передачу тепла к земле. и стабилизировать внутреннюю температуру . Уменьшенный хвост также уменьшил бы общую площадь поверхности животного, что еще больше свело бы к минимуму потери тепла. [ 47 ] Среди терапсидов только эутериодонты (не горгонопсианы) имеют дыхательные носовые раковины , которые помогают удерживать влагу при вдыхании большого количества воздуха, и их эволюция обычно связана с началом «млекопитающих» темпов потребления кислорода и возникновением эндотермии. [ 44 ]
Если горгонопсианы были инерционными гомеотермами, не исключено, что у них были волосы. Морда обычно пронизана отверстиями (небольшими отверстиями, которые сообщаются с кровеносными сосудами), что потенциально может указывать на наличие дряблой кожи (в отличие от чешуи), волос, различных кожных желез (таких как потовые железы ) и усов ; однако у некоторых рептилий имеется аналогичное расположение отверстий, которые вместо этого связаны с развитием зубов, а не с кожей. [ 48 ]
Палеопатология
[ редактировать ]

Переднелатеральная часть левой лучевой кости (кость предплечья) горгонопсианского экземпляра NHCC LB396 представляет собой круглое костное поражение с шипами неправильной или радиальной формы , состоящими из кортикальной кости, окруженными тонким слоем поднадкостничной кости , которая быстро росла на одном участке. вегетационный период. Это согласуется с периоститом , скорее всего, возникшим из-за поднадкостничной гематомы . Это специфическое состояние, а также быстрая скорость роста больше напоминают млекопитающих и динозавров, чем крокодилов или варанов . Среди ранних синапсид единственной отмеченной патологией является остеомиелит в нескольких группах пеликозавров. [ 49 ]
Лабиальная (губа/щека) сторона корня зуба функционального клыка RB382 представляет до 8 поражений, группирующихся вдоль средней линии зуба, которые напоминают миниатюрные зубы с пульпой , дентином и тонким эмалевым покрытием. Они имеют примерно круглую форму — диаметр варьируется от 0,3–3,9 мм (0,012–0,154 дюйма), хотя становятся менее круглыми примерно в средней точке корня, пока не пройдут шейку зуба . Это примерно соответствует заболеванию человека одонтоме , наиболее частому типу одонтогенной опухоли , которая ранее в летописи окаменелостей распространилась всего на несколько миллионов лет назад. RB382 возрастом 255 миллионов лет представляет собой старейший известный случай одонтомы. [ 50 ]
Морда взрослой особи SAM-PK-11490 неопределенного среднепермского вида горгонопсов имеет встроенный зуб неопознанного животного. На кости вокруг зуба образовалась мозоль, что указывает на то, что он зажил и человек пережил нападение. Это произошло либо от хищника, а именно от биармозуха , тероцефала или другого горгонопсиса, либо от внутривидового кусания лица, что обычно наблюдается у социальных хищников, таких как большие кошки или вараны, и это было предложено для нескольких вымерших линий, таких как тероподы. , водные рептилии и саблезубые кошки. Социальный укус предназначен для утверждения доминирования или облегчения размножения, и, если это так, можно предположить, что по крайней мере некоторые среднепермские горгонопсии были социальными хищниками. Изначально зуб не был замечен, поэтому неясно, насколько распространена эта патология на самом деле. [ 51 ]
Палеоэкология
[ редактировать ]Палеосреда
[ редактировать ]
После вымирания диноцефалов и (в Южной Африке) базальных тероцефалов Scylacosauridae и Lycosuchidae , горгонопсии превратились из мелких и необычных форм в крупных высших хищников . [ с ] На протяжении средней и верхней перми в Южной Африке дицинодонты были наиболее распространенными животными, тогда как парейазавры Deltavjatia и Scutosaurus были наиболее многочисленными в горгонопсиеносных русских формациях. [ 52 ] В течение верхней перми южноафриканская группа Бофорта представляла собой полузасушливую холодную степь с большими сезонными ( эфемерными ) реками и поймами, истощающими источники воды гораздо дальше на север, в море Кару, с некоторыми случаями ливневых паводков после внезапных сильных дождей; [ 53 ] [ 54 ] Распределение карбонатов соответствует современным отложениям калише , которые образуются в климате со средней температурой 16–20 ° C (61–68 ° F) и 100–500 мм (3,9–19,7 дюйма) сезонных осадков. [ 54 ] Саларевская свита, содержащая горгонопсии, на западе России также, вероятно, отложилась в полузасушливой среде с большим количеством сезонных осадков и характеризовалась гигрофитными и галофитными растениями в прибрежных районах, а также более засухоустойчивыми хвойными породами на возвышенностях. [ 55 ] Формация Моради представляла собой засушливую пустыню, в которой в основном доминировали капториниды рептилии- Морадизавра и парейазавр Буностегос . [ 20 ] Здесь растут вольтцианские хвойные деревья , и с экологической точки зрения его сравнивают с внутренней пустыней Намиб или бассейном озера Эйр . [ 56 ]
Вымирание
[ редактировать ]Большинство горгонопсов вымерло в конце верхней перми во время пермско-триасового вымирания , которое было в первую очередь вызвано вулканизмом, образовавшим Сибирские траппы . Результирующий массивный скачок выбросов парниковых газов вызвал быструю засушливость из-за: скачка температуры (до 8–10 °C на экваторе, со средней экваториальной температурой 32–35 °C или 90–95 °F в начале триасовый период), кислотные дожди (с pH всего лишь 2 или 3 во время извержения и 4 во всем мире, а также последующее обезлесение в течение первых 10 миллионов лет триаса), частые лесные пожары (хотя они уже были довольно распространены на протяжении всего пермского периода). ) и потенциальное разрушение озонового слоя (возможно, кратковременное увеличение бомбардировки УФ-излучением на 400% на экваторе и на 5000% на полюсах). [ 57 ] Возможный экземпляр цианозавра позволяет предположить, что некоторые более мелкие таксоны, возможно, дожили до поздней перми или раннего триаса . [ 58 ] Среди терапсидов мелкие тероцефалы и крупные травоядные аномодонты сумели пересечь границу перми и триаса и дожили соответственно до среднего и верхнего триаса , но в юрский период дожили только мелкие виды цинодонтов, потомками которых будут млекопитающие. Ниши, оставленные горгонопсианами открытыми, в конечном итоге были заполнены архозаврами ( включая крокодилов и динозавров ) на ранних стадиях триаса . [ 59 ]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ ce, мозжечок ; cnI, обонятельный нерв ; cnV +vcm — тройничный нерв и медиальная вена головы ; cnVI, отводящий нерв ; cnVII, лицевой нерв ; cnIX-XI, языкоглоточный и блуждающе-добавочный нервы ; cnXII — подъязычный нерв ; en, эпифизарный нерв ; фб, передний мозг ; fcl, флоккул ; ibic, внутренняя ветвь внутренней сонной артерии ; долька, левая обонятельная луковица ; об, обонятельная луковица; от, обонятельный тракт ; пг, гипофиз ; pgll — боковые доли гипофиза; pf — изгиб моста ; роба, правая обонятельная луковица; ВЦ, Видианский канал ; vc=spa, видиан канал, где клиновидно-небная артерия проходит ; VCD, спинная вена головы
- ^ В более поздних классификациях семейство Gorgonopsidae обычно определяется так, чтобы исключить базальных представителей Gorgonopsia. [ 7 ] : 242–244
- ^ Публикация в 2022 году официального описания рода Phorcys и аналогичного неопределенного экземпляра показывает, что некоторые горгонопсии уже в средней перми были крупнее по размеру, но не достигли, однако, размеров, подобных Inostrancevia или Rubidgeines . Однако крупные горгонопсии, такие как Phorcys и другие крупные терапсиды, возможно, оставили разделение ниши более мелким представителям после вымирания капитанов . [ 33 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Бендель, Ева-Мария; Каммерер, Кристиан Ф.; Карджилов, Николай; Фернандес, Винсент; Фрёбиш, Йорг (2018). «Краниальная анатомия горгонопсиана Cynariopsrobustus на основе КТ-реконструкции» . ПЛОС ОДИН . 13 (11): e0207367. Бибкод : 2018PLoSO..1307367B . дои : 10.1371/journal.pone.0207367 . ПМК 6261584 . PMID 30485338 .
- ^ Протеро, Дональд Р. (18 апреля 2022 г.). «20. Синапсиды: происхождение млекопитающих». Эволюция позвоночных: от истоков до динозавров и далее . Бока-Ратон: CRC Press. дои : 10.1201/9781003128205-4 . ISBN 9780367473167 . S2CID 246318785 .
- ^ Jump up to: а б с Лорен, Мишель (1998). «Новые данные о краниальной анатомии Lycaenops (Synapsida, Gorgonopsidae) и размышления о возможном наличии стрептостилии у горгонопсов». Журнал палеонтологии позвоночных . 18 (4): 765–776. Бибкод : 1998JVPal..18..765L . дои : 10.1080/02724634.1998.10011105 .
- ^ Jump up to: а б Суинтон, МЫ (1954). Ископаемые амфибии и рептилии . Британский музей (естественная история). п. 33.
- ^ Ангельчик, Кеннет Д.; Каммерер, Кристиан Ф. (2018). «Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». В Захосе, Фрэнк Э.; Ашер, Роберт Дж. (ред.). Эволюция, разнообразие и систематика млекопитающих . Берлин: Де Грюйтер. ISBN 9783110275902 .
- ^ Jump up to: а б с д и ж г час я дж к Каммерер, Кристиан Ф. (2016). Сьюс, Ханс-Дитер (ред.). «Систематика Rubidgeinae (Therapsida: Gorgonopsia)» . ПерДж . 4 : е1608. дои : 10.7717/peerj.1608 . ПМЦ 4730894 . ПМИД 26823998 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Гебауэр, Ева VI (2007). Филогения и эволюция горгонопсии с особым упором на череп и скелет GPIT/RE/7113 (PDF) (доктор философии). Тюбингенский университет Эберхарда-Карла. Архивировано из оригинала 22 июля 2012 г.
- ^ Jump up to: а б Араужо, Рикардо; Фернандес, Винсент; Полцин, Майкл Дж.; Фрёбиш, Йорг; и Мартинс, Руи М.С. (2017). «Аспекты палеобиологии и эволюции горгонопсов: данные о базальном черепе, затылке, костном лабиринте, сосудистой сети и нейроанатомии» . ПерДж . 5 : е3119. дои : 10.7717/peerj.3119 . ПМК 5390774 . ПМИД 28413721 .
- ^ Jump up to: а б Кермак, Дорис М. и Кермак, Кеннет А. (1984). «Зубные ряды, замена зубов и артикуляция челюстей». Эволюция характеров млекопитающих . стр. 71–79. дои : 10.1007/978-1-4684-7817-4_5 . ISBN 978-1-4684-7817-4 .
- ^ Jump up to: а б Уитни, MR; ЛеБлан, ARH; Рейнольдс, Арканзас; и Бринк, Канзас (2020). «Конвергентные зубные адаптации в зубцах гиперплотоядных синапсидов и динозавров» . Письма по биологии . 16 (12). дои : 10.1098/rsbl.2020.0750 . ПМЦ 7775981 . ПМИД 33321067 .
- ^ Jump up to: а б с Ван Валкенбург, Блэр и Дженкинс, Ян (2002). «Эволюционные закономерности в истории пермо-триасовых и кайнозойских синапсидных хищников». Документы Палеонтологического общества . 8 : 273–275. дои : 10.1017/S1089332600001121 .
- ^ Jump up to: а б Кермак, К.А. (1956). «Замена зубов у млекопитающих рептилий подотрядов Gorgonopsia и Therocephalia». Философские труды Лондонского королевского общества B: Биологические науки . 240 (670): 95–133. Бибкод : 1956РСПТБ.240...95К . дои : 10.1098/rstb.1956.0013 .
- ^ Jump up to: а б Кюммелл, SB; Фрей, Э. (2014). «Диапазон движения в луче I Manus и Pes и цепкость аутоподий в ранней перми и позднем мелу неаномодонтных синапсид» . ПЛОС ОДИН . 9 (12): e113911. Бибкод : 2014PLoSO...9k3911K . дои : 10.1371/journal.pone.0113911 . ПМЦ 4269487 . ПМИД 25517726 .
- ^ Jump up to: а б Оуэн, Ричард (1986). Описательный и иллюстрированный каталог ископаемых рептилий Южной Африки в коллекции Британского музея . Отпечатано по заказу попечителей. стр. 27–29.
- ^ Jump up to: а б с д и Каммерер, Кристиан Ф. и Масютин, Владимир (2018). Янг, Марк (ред.). «Горгонопсиевые терапсиды ( Ночница gen. nov. и Вяткогоргон ) из пермского местонахождения Котельнич России» . ПерДж . 6 : е4954. дои : 10.7717/peerj.4954 . ПМЦ 5995105 . ПМИД 29900078 .
- ^ Янг, Чанг К. (1979). «Позднепермская фауна из Цзиюаня, Хэнань». Позвоночные Палазиатские . 17 (2): 99–113.
- ^ Сигоньо-Рассел, Дениз ; Сунь, Ай-Линь (1981). «Краткий обзор китайских синапсидов». Геобиос . 14 (2): 276. Бибкод : 1981Geobi..14..275S . дои : 10.1016/S0016-6995(81)80012-5 . S2CID 129556772 .
- ^ Лю, Цзюнь; Ян, Ван (сентябрь 2022 г.). «Горгонопсиан из формации Утунгоу (чансин, пермь) Турфанского бассейна, Синьцзян, Китай» . Палеомир . 31 (3): 383–388. дои : 10.1016/j.palwor.2022.04.004 .
- ^ Рэй, Сангамитра; Бандиопадхьяй, Сасвати (март 2003 г.). «Позднепермское сообщество позвоночных долины Пранхита-Годавари, Индия» . Журнал азиатских наук о Земле . 21 (6): 643–654. Бибкод : 2003JAESc..21..643R . дои : 10.1016/S1367-9120(02)00050-0 . S2CID 140601673 . Проверено 24 сентября 2022 г.
- ^ Jump up to: а б Смайли, Тара М.; Сидор, Кристиан А.; Мага, Абдулай; Иде, Умару (2008). «Фауна позвоночных верхней перми Нигера. VI. Первые свидетельства горгонопсиевого терапсида» . Журнал палеонтологии позвоночных . 28 (2): 543–547. Бибкод : 2008JVPal..28..543S . doi : 10.1671/0272-4634(2008)28[543:TVFOTU]2.0.CO;2 . S2CID 131082914 .
- ^ tectus в Чарльтоне Т. Льюисе (1891) Элементарный латинский словарь , Harper & Brothers
- ^ Лидеккер, Ричард (1890). Каталог ископаемых рептилий и амфибий в Британском музее (Естественная история) Часть IV . Британский музей (естественная история). п. 111.
- ^ Сили, Гарри Г. (1895). «Исследования по строению, организации и классификации ископаемых рептилий. — Часть IX. Раздел 1. О терозухиях» . Анналы и журнал естественной истории . 13 (6): 375. дои : 10.1080/00222939408677718 .
- ^ Осборн, Генри Ф. (1903). «Подклассы рептилий Diapsida и Synapsida и ранняя история диаптозаврий. Том I. Часть VIII» (PDF) . Мемуары Американского музея естественной истории . 1 : 451–466.
- ^ Брум, Роберт (1905). «Об употреблении термина Аномодонтия» . Записи музея Олбани . 1 (4): 266–269.
- ^ Брум, Роберт (1913). «О горгонопсиях, подотряде млекопитающих рептилий» . Труды Лондонского зоологического общества . 1913 (2): 225–230. дои : 10.1111/j.1469-7998.1913.tb07574.x .
- ^ Уилли, Алистер (2003). «Обзор голотипов терапсида Роберта Брума: выдержали ли они испытание временем?» (PDF) . Палеонтология Африканская . 39 (39): 2.
- ^ Чемберс, Кристиан Ф. (2014), «Териодонтия: Введение», в Чемберс, Кристиан Ф.; Ангельчик, Кеннет Д.; Фрёбиш, Йорг (ред.), Ранняя эволюционная история Synapsida , Палеобиология и палеоантропология позвоночных, Дордрехт : Springer, стр. 165–169, номер домена : 10.1007/978-94-007-6841-3_10 , ISBN. 978-94-007-6841-3
- ^ Jump up to: а б Сигоньо-Рассел, Дениз (1989). Велнхофер, Питер (ред.). Териодонтия I: Фтинозухия, Биармозухия, Эотитанозухия, Горгонопсия . Энциклопедия палеогерпетологии. Том. 17 Б/И. Штутгарт: Густав Фишер Верлаг. ISBN 978-3437304873 .
- ^ Рейс, Роберт Р.; Лорен, Мишель (2004). «Переоценка загадочного пермского синапсида Watongia и его стратиграфического значения» . Канадский журнал наук о Земле . 41 (4): 377–386. Бибкод : 2004CaJES..41..377R . дои : 10.1139/e04-016 . S2CID 129233127 .
- ^ Ивахненко, Михаил Федорович (2002). «Таксономия восточноевропейских горгонопий (Therapsida)». Палеонтологический журнал . 36 (3): 283–292. ISSN 0031-0301 .
- ^ Каммерер, Кристиан Ф.; Смит, Роджер М.Х.; Дэй, Майкл О.; Рубидж, Брюс С. (2015). «Новые сведения о морфологии и стратиграфическом ареале среднепермского горгонопса Eriphostoma microdon Broom, 1911». Статьи по палеонтологии . 1 (2): 201–221. Бибкод : 2015PPal....1..201K . дои : 10.1002/spp2.1012 . S2CID 128762256 .
- ^ Jump up to: а б с д Каммерер, Кристиан Ф.; Рубидж, Брюс С. (2022). «Самые ранние горгонопсии из бассейна Кару в Южной Африке». Журнал африканских наук о Земле . 194 : 104631. Бибкод : 2022JAfES.19404631K . doi : 10.1016/j.jafrearsci.2022.104631 . S2CID 249977414 .
- ^ Jump up to: а б с Кемп, Том С. (2006). «Происхождение и раннее распространение терапсидных млекопитающих рептилий: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. дои : 10.1111/j.1420-9101.2005.01076.x . ПМИД 16780524 . S2CID 3184629 .
- ^ Дэй, Майкл О.; Рамезани, Джахандар; Боуринг, Сэмюэл А.; Сэдлер, Питер М.; Эрвин, Дуглас Х.; Абдала, Фернандо; и Рубидж, Брюс С. (2015). «Когда и как произошло массовое вымирание наземных видов в середине пермского периода? Свидетельства находок четвероногих в бассейне Кару, Южная Африка» . Труды Королевского общества Б. 282 (1811): 20150834. doi : 10.1098/rspb.2015.0834 . ПМЦ 4528552 . ПМИД 26156768 .
- ^ Jump up to: а б с Энтони, Морис (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. стр. 100-1 204–207. ISBN 978-0-253-01042-1 .
- ^ Рандау, Марсела; Карбоне, Крис; и Терви, Сэмюэл Т. (2013). Эванс, Алистер Роберт (ред.). «Собачья эволюция саблезубых хищников: естественный отбор или половой отбор?» . ПЛОС ОДИН . 8 (8): е72868. Бибкод : 2013PLoSO...872868R . дои : 10.1371/journal.pone.0072868 . ПМЦ 3738559 . ПМИД 23951334 .
- ^ Крукшанк, ARI (1973). «Образ жизни горгонопсианцев» (PDF) . Палеонтологическая африканская . 15 (1–2): 65–67.
- ^ Эстефа, Хорди; Таффоро, Пол; Клемент, Элис М.; Клембара, Йозеф; Недзведский, Гжегож; Берруйер, Камилла; и Санчес, Софи (2021). Таутц, Дитхард (ред.). «Новый свет пролил свет на раннюю эволюцию пластинок роста костей конечностей и костного мозга» . электронная жизнь . 10 : 51581. doi : 10.7554/eLife.51581 . ПМЦ 7924947 . PMID 33648627 .
- ^ Крылов А.; Жермен, Д.; Кановиль, А.; Винсент, П.; Саш, М. и Лорен, М. (2008). «Эволюция костной микроанатомии большеберцовой кости четвероногих и ее использование в палеобиологических выводах» (PDF) . Журнал эволюционной биологии . 21 (3): 807–826. дои : 10.1111/j.1420-9101.2008.01512.x . ПМИД 18312321 .
- ^ Маркетти, Лоренцо; Кляйн, Хендрик; Бухвиц, Майкл; Рончи, Аусонио; Смит, Роджер М.Х.; Де Клерк, Уильям Дж.; Шишио, Лара; Гроеневальд, Гидеон Х. (2019). «Следы пермско-триасовых позвоночных из Южной Африки: ихнотаксономия, производители и биостратиграфия в условиях двух крупных фаунистических кризисов» . Исследования Гондваны . 72 : 159. Бибкод : 2019GondR..72..139M . дои : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Бенуа, Жюльен; Абдала, Фернандо; Мангер, Пол Р.; и Рубидж, Брюс С. (2016). «Шестое чувство у предшественников млекопитающих: изменчивость теменного отверстия и эволюция шишковидного глаза у южноафриканских пермо-триасовых эвтериодонтных терапсидов» . Acta Palaeontologica Polonica . 61 (4): 777–789. дои : 10.4202/app.00219.2015 . S2CID 59143925 .
- ^ Ангельчик К.Д. и Шмитц Л. (2014). «Ночной образ жизни в синапсидах предшествует возникновению млекопитающих более чем на 100 миллионов лет» . Труды Королевского общества Б. 281 (1793). дои : 10.1098/rspb.2014.1642 . ПМК 4173690 . ПМИД 25186003 .
- ^ Jump up to: а б Хиллениус, Виллем Дж. (1992). Носовые раковины и эволюция эндотермии млекопитающих (доктор философии). Государственный университет Орегона. стр. 74–76. HDL : 1957/35986 .
- ^ Хиллениус, Виллем Дж. (2000). «Септомаксилла синапсид немлекопитающих: корреляты мягких тканей и новая функциональная интерпретация». Журнал морфологии . 245 (1): 29–50. doi : 10.1002/1097-4687(200007)245:1<29::AID-JMOR3>3.0.CO;2-B . ПМИД 10861830 . S2CID 22452028 .
- ^ Кемп, Т.С. (2006). «Происхождение эндотермии млекопитающих: парадигма эволюции сложной биологической структуры» (PDF) . Зоологический журнал Линнеевского общества . 147 (4): 473–488. дои : 10.1111/j.1096-3642.2006.00226.x .
- ^ Тернер, Дж. Скотт и Трейси, К. Ричард (1986). «Размер тела, гомеотермия и контроль тепла тела у млекопитающих рептилий». В Хоттоне, Николай III; Маклин, Пол Д.; Рот, Ян Дж.; и Рот, Э. Кэрол (ред.). Экология и биология млекопитающих рептилий (PDF) . Издательство Смитсоновского института. Архивировано из оригинала (PDF) 20 января 2021 г. Проверено 26 сентября 2020 г.
- ^ Ван Велан, Ли (сентябрь 1960 г.). «Терапсиды как млекопитающие». Эволюция . 14 (3): 304–313. дои : 10.2307/2405973 . JSTOR 2405973 .
{{cite journal}}: CS1 maint: дата и год ( ссылка ) - ^ Като, Кайл М.; Рега, Элизабет А.; Сидор, Кристиан А.; и Хаттенлокер, Адам К. (2020). «Исследование поражения костей горгонопсии (Synapsida) из перми Замбии и периостальных реакций у ископаемых четвероногих, не являющихся млекопитающими» . Философские труды Королевского общества Б. 375 (1793). дои : 10.1098/rstb.2019.0144 . ПМК 7017433 . ПМИД 31928188 .
- ^ Уитни, Меган Р.; Моуз, Ларри; и Сидор, Кристиан А. (2017). «Одонтома у предка млекопитающих возрастом 255 миллионов лет» . JAMA Онкология . 3 (7): 998–1000. дои : 10.1001/jamaoncol.2016.5417 . ПМЦ 5824274 . ПМИД 27930769 .
- ^ Бенуа, Жюльен; Браунинг, Клэр; Нортон, Люк А. (2021). «Первый заживший след укуса и вросший зуб в морду среднепермского горгонопсина (Synapsida: Therapsida)» . Границы экологии и эволюции . 6 : 699298. дои : 10.3389/fevo.2021.699298 .
- ^ Каммерер, Кристиан Ф. и Масютин, Владимир (2018). Сьюс, Ханс-Дитер (ред.). «Новый тероцефал ( Gorynychus masyutinae gen. et sp. nov.) из пермского местонахождения Котельнич, Кировская область, Россия» . ПерДж . 6 : е4933. дои : 10.7717/peerj.4933 . ПМК 5995100 . ПМИД 29900076 .
- ^ Уилсон, Эндрю; Флинт, Стивен; Пайенберг, Тобиас; Тохвер, Эрик; Ланчи, Лука (2014). «Архитектурные стили и седиментология речной группы Нижнего Бофорта, бассейн Кару, Южная Африка». Журнал осадочных исследований . 84 (4): 326–348. Бибкод : 2014JSedR..84..326W . дои : 10.2110/jsr.2014.28 . S2CID 129643680 .
- ^ Jump up to: а б Смит, Роджер М.Х. (1987). «Морфология и история отложений эксгумированных пермских острий на юго-западе Кару, Южная Африка». Журнал осадочных исследований . 57 (1): 19–20. дои : 10.1306/212F8A8F-2B24-11D7-8648000102C1865D . S2CID 130393112 .
- ^ Якименко Елена; Иноземцев, Святослав А.; Наугольных, Сергей В. (2004). «Палеопочвы верхней перми (саларевская свита) центральной части Русской платформы: палеоэкология и палеосреда» . Мексиканский журнал геологических наук . 21 (1): 110–119.
- ^ Луй, Синди В.; Ранкс, Стефани Л.; Чейни, Дэн С.; Санчес, Софи; Штайер, Жан-Себастьен; Смит, Роджер М.Х.; Сидор, Кристиан А.; Майерс, Тимоти С.; Иде, Умаро; Табор, Нил Дж. (июнь 2016 г.). «Биологические и физические доказательства крайней сезонности в центральной пермской Пангее» . Палеогеография, Палеоклиматология, Палеоэкология . 451 : 210–226. Бибкод : 2016PPP...451..210L . дои : 10.1016/j.palaeo.2016.02.016 . S2CID 131336926 .
- ^ Бентон, MJ (2018). «Массовые вымирания, вызванные гипертермией: модели убийств во время массового вымирания в пермско-триасовом периоде» . Философские труды Королевского общества А. 376 (2130). Бибкод : 2018RSPTA.37670076B . дои : 10.1098/rsta.2017.0076 . ПМК 6127390 . ПМИД 30177561 .
- ^ Бенуа, Жюльен; Каммерер, Кристиан Ф.; Доллман, Кэтлин; Гроеневальд, Дэвид П.; Смит, Роджер М.Х. (2024). «Пережили ли горгонопсианы «Великое вымирание» в конце перми? Повторная оценка трех экземпляров горгонопсиан (Therapsida, Theriodontia), обнаруженных в триасовой зоне сборки Lystrosaurus declivis , бассейн Кару, Южная Африка». Палеогеография, Палеоклиматология, Палеоэкология . 638 : 112044. Цифровой код : 2024PPP...63812044B . дои : 10.1016/j.palaeo.2024.112044 . S2CID 267080711 .
- ^ Сукиас, Роланд Б.; Батлер, Ричард Дж.; и Бенсон, Роджер Б.Дж. (2012). «Расцвет динозавров показывает, что основные изменения размеров тела вызваны пассивными процессами эволюции признаков» . Труды Королевского общества Б. 279 (1736): 2180–2187. дои : 10.1098/rspb.2011.2441 . ПМК 3321709 . ПМИД 22298850 . S2CID 1231897 .
Внешние ссылки
[ редактировать ] СМИ, связанные с горгонопсией, на Викискладе?
СМИ, связанные с горгонопсией, на Викискладе?  Данные, относящиеся к горгонопсии , в Wikispecies
Данные, относящиеся к горгонопсии , в Wikispecies







| Базы данных органов управления : Национальные |
|---|